1. Introduction
Breast cancer (Online Mendelian Inheritance in Man, OMIM No. 114480) is the most frequent cancer in females worldwide [
1], with 2.1 million new cases diagnosed and more than 620,000 individuals deceased in 2018 [
2]. Treatment of so many patients is a serious burden for healthcare systems and calls for an individualized approach. Imbalances in absorption, distribution, metabolism, and excretion of drugs used for cancer therapy influence the drug efficacy and cause adverse drug reactions in some patients. Thus, inter-individual genetic variations in drug-metabolizing enzymes became important in the clinical setting [
3]. However, prognostic and predictive biomarkers for precision therapy of breast cancer, e.g., pharmacogenetic variants, are still missing [
4].
Cytochromes P450 (CYPs) represent a large superfamily of membrane hemoproteins classified into 18 families in humans [
5,
6]. CYP monooxygenases oxidize or reduce a broad range of physiological substrates, e.g., sterols and fatty acids [
7] and xenobiotics, including drugs [
8], and together with ATP-binding transporters, represent the majority of known pharmacogenes [
4]. However, of the total number of 57 human CYP coding genes with putative enzymatic activities, only 15 seem to metabolize drugs to date [
9], and genetic polymorphisms of
CYP2C9,
CYP2C19, and
CYP2D6 belong to the most frequently studied in pharmacogenomics [
6]. Although the rest of the CYPs are not involved in drug metabolism perturbations in the homeostasis of steroid hormones, e.g., estrogen may also influence the prognosis and therapy outcome of the patients [
10]. Moreover, inhibitors of CYP19 aromatase are frequently used for endocrine therapy of breast cancer patients, while the role of genetic variability of the target gene in the treatment efficacy or adverse effects is yet unexploited for therapeutic decisions [
11].
Our pilot pharmacogenomics study followed germline alterations in 509 selected genes and their potential for prognosis and prediction of response to therapy in breast cancer patients [
12]. We provided a proof-of-the principle for the study design and established bioinformatics methodology for variant prioritization. Out of all genes whose coding and regulatory sequences were screened by the next-generation sequencing (NGS) approach, only a few variants were validated in the replication phase. We down-sampled the first-phase results by the synthesis of in silico predictions and statistically significant clinical associations. This strict process resulted in considerably smaller numbers of variants for replication, and some potentially useful biomarkers of prognosis or prediction of therapy outcome potentially remained unexplored.
In the present study, we used less-strict criteria for investigating the importance of germline genetic variability in coding, untranslated regions (UTR), and adjacent regions of all human members of the CYP superfamily for prognosis and response to the neoadjuvant cytotoxic therapy (NACT) of breast cancer patients. First, we correlated variants with the response of patients to NACT and disease-free survival (DFS). We then thoroughly reviewed haplotypes and gene dosage and corrected results for multiple testing. This study has not addressed the functional relevance to enable also the identification of purely correlative biomarkers. Prioritized variants underwent confirmation in a large cohort of breast cancer patients from a single population. Taken together, the present study brings a more detailed view of the relevance of germline genetic variability of CYPs for breast cancer prognosis and therapy outcome.
3. Discussion
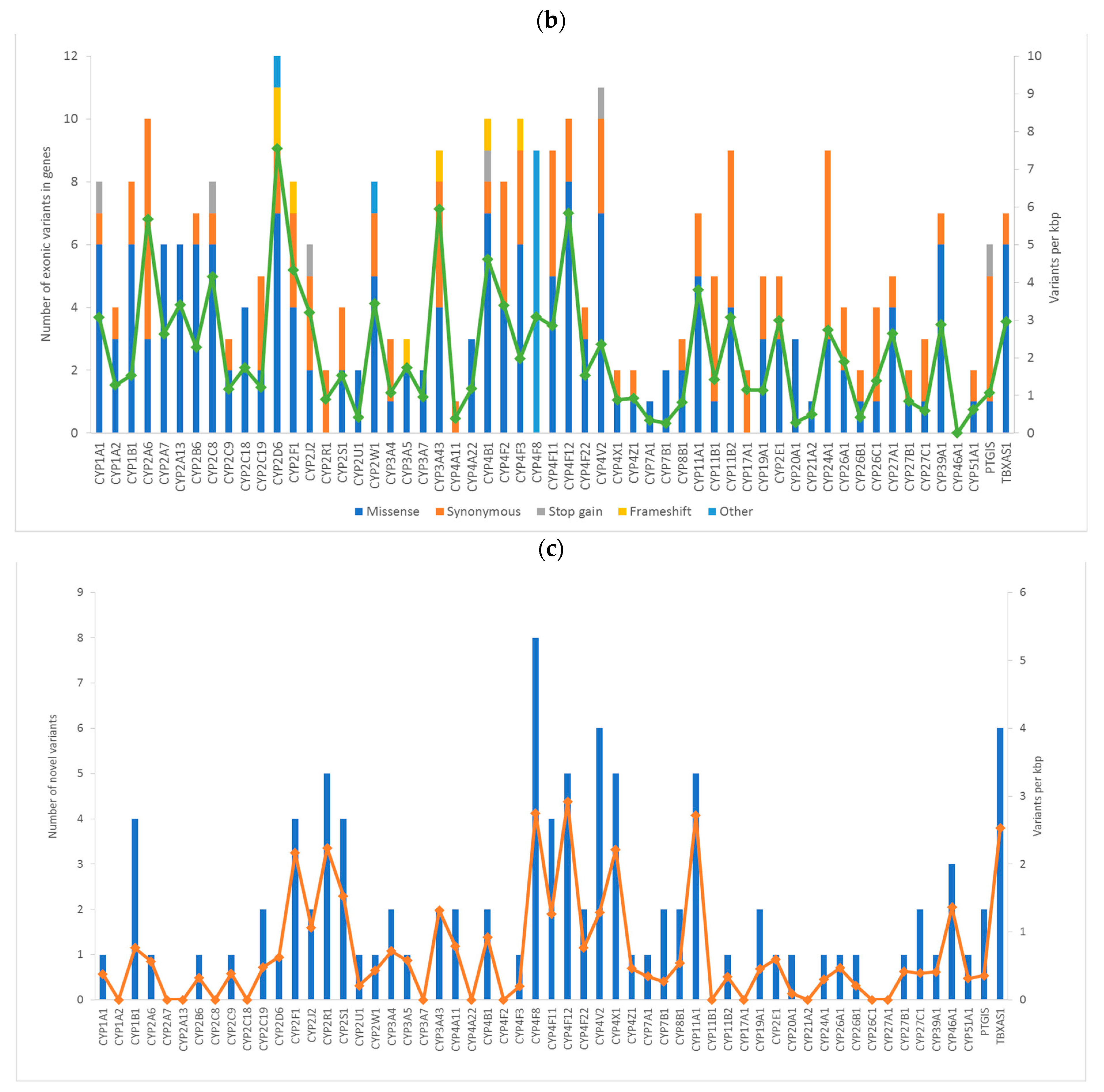
We analyzed associations of genetic variants in all human CYP monooxygenases with chemotherapy outcome and survival of breast cancer patients. Firstly, we genotyped all coding sequences and surrounding areas using the next-generation sequencing. Variants significantly associated with response to NACT and DFS of the patients using Chi-square and log-rank tests with permutations were further assessed in a large cohort of breast cancer patients (n = 802) by competitive allele-specific PCR. Of the total number of 22 variants selected for validation, the results were confirmed, in a large cohort, for six of them.
In total, we found 1274 variants in a set of 105 breast cancer patients used for the evaluation phase. We found 14 LOF variants. Of these, six were stop-gain mutations, and eight were frameshift indels. However, the MAF of these variants was too low to maintain the statistical power for correlation with clinical data precluding their further study. Thus, we evaluated 22 common variants in a large confirmation cohort of patients and assessed their associations with DFS and response to NACT.
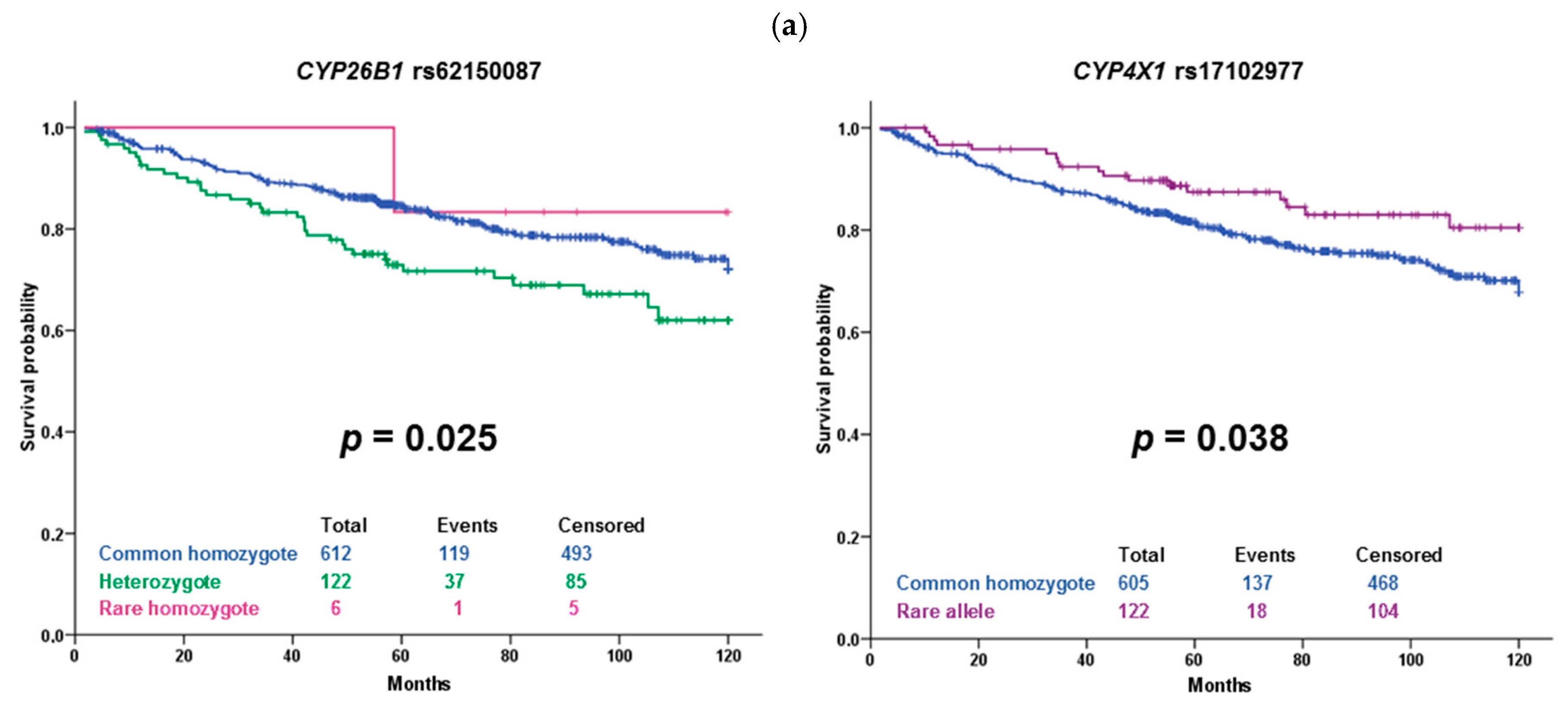
The substitution rs17102977 in
CYP4X1 intron is associated with both response of the patients to the NACT and DFS of hormonally treated patients. It is also prognostic in patients unselected according to the therapy. The association of rs17102977 with the response to NACT was significant after correction to multiple testing and thus can be considered validated in both datasets. Patients carrying the rare allele G were more often poorly responding to chemotherapy than the patients with the wild-type genotype AA. Intriguingly, patients bearing rare alleles had longer DFS than wild-type patients. Although the frequency of the rare allele in the European non-Finnish population in GnomAD is 0.08, we found no record for this variant in the scientific literature (PubMed). The gene
CYP4X1 encodes an orphan CYP enzyme expressed mainly in the brain, aorta, or breast [
13,
14]. According to recent studies, this enzyme catalyzes epoxidation of endogenous cannabinoid anandamide and arachidonic acid [
15,
16]. Its important paralog is
CYP4Z1 [
17]. The role of
CYP4X1 in cancer has been proposed as well. Protein expression of CYP4X1 was associated with increasing tumor grade in tissue microarrays from 170 breast cancer patients detected by immunohistochemistry [
18]. Recently, a lower gene expression of
CYP4X1 was associated with shorter overall survival of Chinese gastric cancer patients treated with capecitabine and oxaliplatin [
19]. Two SNPs in
CYP4X1 were also associated with the early onset of Creutzfeldt-Jakob disease in Italian patients [
20]. The endocannabinoid system is involved in various physiological processes, including inflammation, immunomodulation, or suppression of different cancers, e.g., breast cancer [
21]. Therefore, CYP4X1 might play a role in response to cancer chemotherapy through physiological processes. The role of rs17102977 in cancer is unknown. According to the GTEx portal (
https://www.gtexportal.org) (Access on: 10 February 2021), the expression quantitative trait loci (eQTL) analysis showed that rs17102977 was significantly associated with
CYP4A22 gene expression in the brain, but not in the breast tissue. No significant association with
CYP4X1 gene expression was found. Therefore, further elucidation of the function of rs17102977 and mainly the whole gene locus 1p33 containing also
CYP4B1,
CYP4A11, CYP4Z1, and
CYP4A22 genes is needed. This locus also contains mitogen-activated protein kinase
MNK1, frequently studied in cancer research.
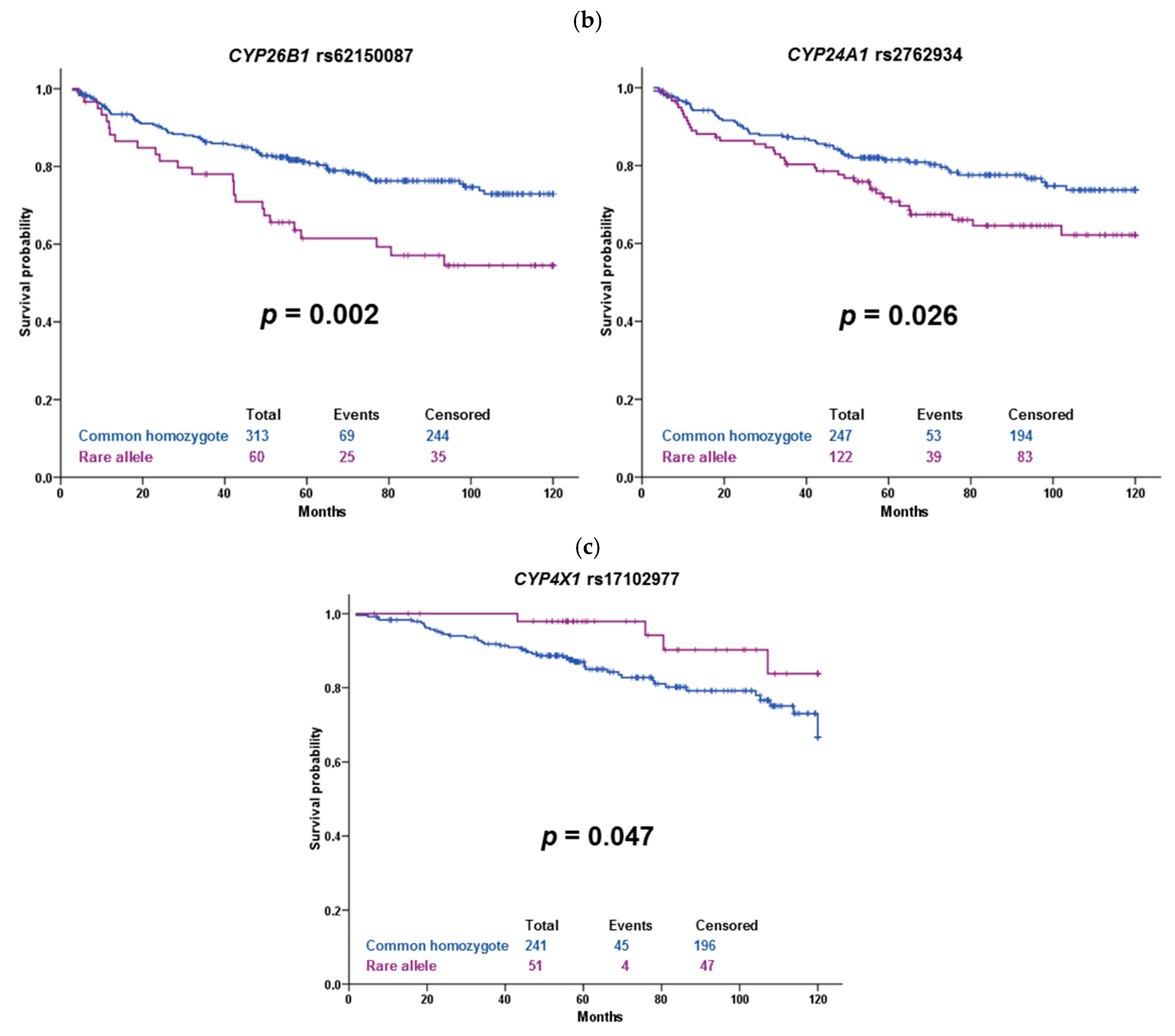
The variant rs62150087 (500 bp downstream of
CYP26B1) was associated with DFS in our sets of patients. Carriers of the rare allele G treated with cytotoxic therapy had significantly shorter DFS than wild-type patients, but the observed
p-value of 0.002 does not guarantee a true positive association after correction for multiple testing. However, this borderline significant result is interesting. A genome-wide association study (GWAS) on the Chinese population identified
CYP26B1 as a candidate gene for esophageal squamous cell carcinoma risk. According to the authors, a variant rs138478634 in
CYP26B1 influences the risk through catabolism of an anticancer nutrient all-
trans retinoic acid [
22]. Considering the ability of CYP26B1 to metabolize retinoic acid, there is support for the observed effect of rs62150087 on worse patient survival. However, the influence of this downstream variant on enzyme function or expression is not known, and no significant eQTLs were found in any of the available tissues.
Among other associations found, the variant rs1056827 in
CYP1B1 was associated with the response of the patients to NACT. Homozygotes of the variant allele A were more often poorly responding to chemotherapy than wild-type patients or heterozygotes. This SNP 355G>T changing Alanine to Serine in codon 119 is associated with susceptibility to colorectal [
23,
24], breast cancer [
25], and primary open-angle glaucoma [
26]. According to ClinVar, this variant is considered benign, and thus no clear explanation of the observed association is available now. Also, the link between breast cancer risk and therapy response is unclear. CYP1B1 oxidizes estrogen as well as a wide variety of xenobiotics [
14,
27]. This fact perhaps could help to explain associations with both risk and therapy outcome and focus further studies. Similarly, synonymous variant rs593421 in
CYP4F12 has no clear support for an observed association with response.
CYP4F12 was cloned originally from the intestine and liver, its recognized substrates are fatty acids [
13,
14]. Also, intronic variant rs3819733 in thromboxane synthase (
TBXAS1) was associated with response to NACT in our patients. In the literature, lower
TBXAS1 expression was associated with higher grade and poor prognosis of breast cancer patients [
28]. On the contrary, high expression was associated with worse overall survival of patients with diffuse low-grade glioma [
29]. CYP24A1 is responsible for vitamin D metabolism [
30]. SNP rs2762934 in an intronic region of
CYP24A1 was associated with an increased risk of breast cancer [
31], ischemic stroke [
32], and hypertension [
33]. We have seen an association of rs2762934 with DFS of patients treated with cytotoxic therapy, especially in the TNBC subgroup. Taken together, these associations are interesting but should be treated with caution because of their low significance level after FDR correction, as well as considering vague support in the literature.
The present study has some limitations. First, the modest size of the evaluation set may be seen as a study limitation, because it precludes assessment of the importance of very rare (MAF < 0.001) and rare (<0.01) variants. In light of the recently acknowledged contribution of rare variants to inter-individual variability in drug response [
3], this limitation needs attention in future studies in precision oncology. On the other hand, we consider ethnical homogeneity and robustness of clinical follow-up as notable study benefits. Moreover, we have employed the multivariate analyses adjusted to major disease characteristics and stratified patients according to intrinsic molecular subtypes to circumvent the frequently overlooked issue of non-homogeneity from the clinical point of view. Major conclusions of the study remained unchanged. Up to that, further elucidations are needed to explore additional genetic components, e.g., non-coding sequences, copy numbers and structural variations, somatic mutations, etc., of the CYP superfamily in breast cancer.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}