
Moscatilin Inhibits Metastatic Behavior of Human Hepatocellular Carcinoma Cells: A Crucial Role of uPA Suppression via Akt/NF-κB-Dependent Pathway

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
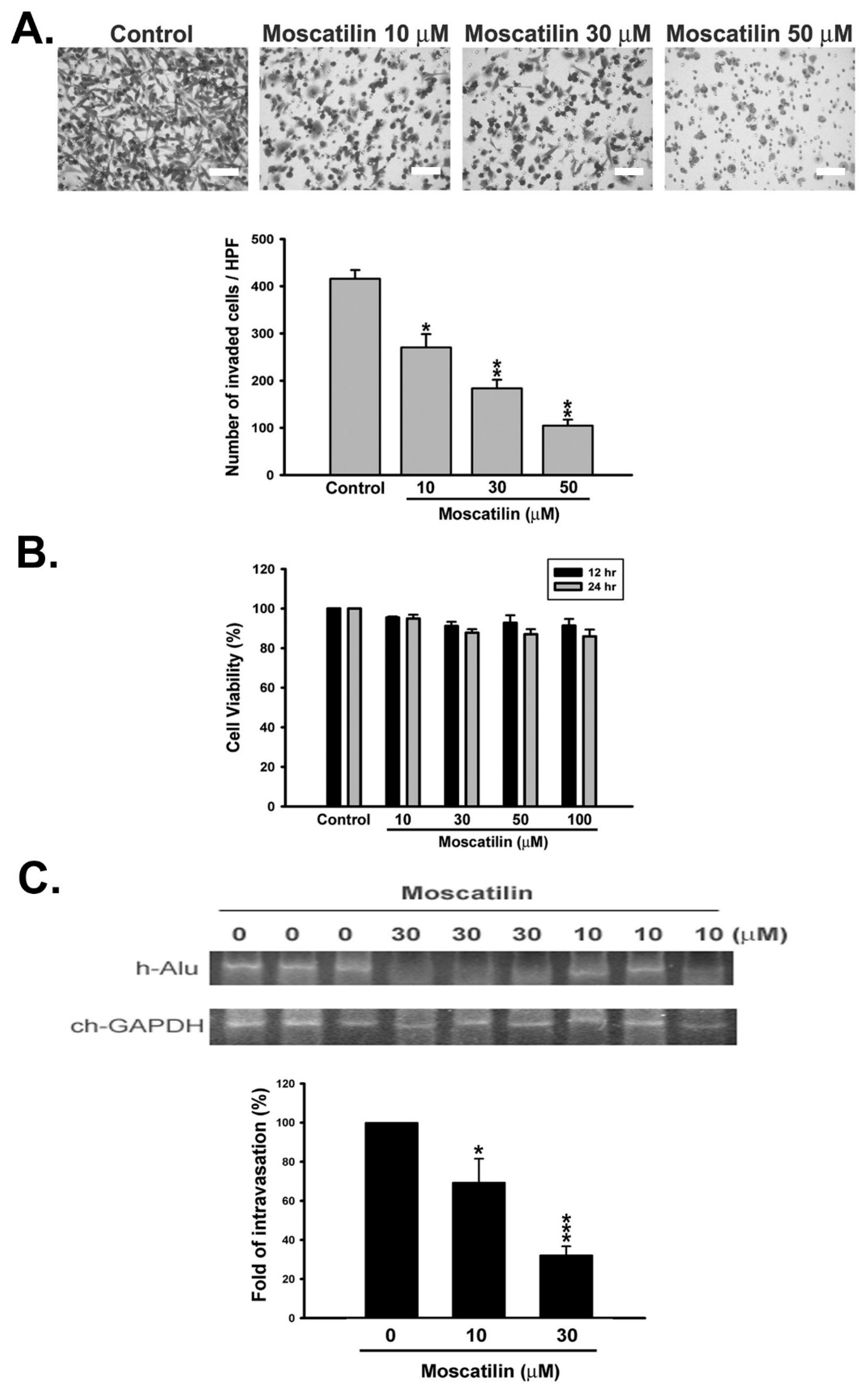
2.1. Moscatilin Inhibited Metastatic Behavior of HCC Cells In Vitro and In Vivo
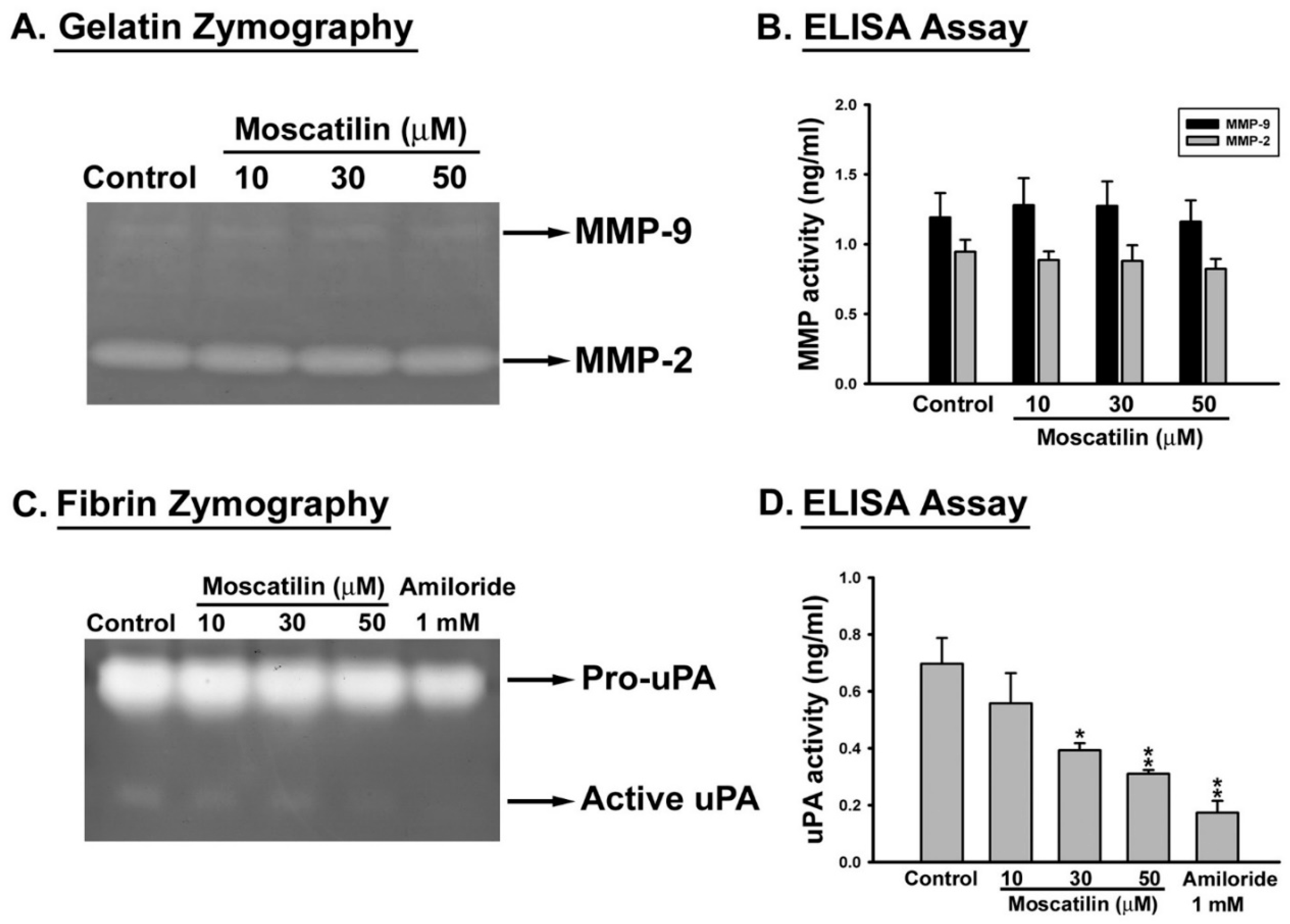
2.2. Moscatilin Decreased the Proteolytic Activity of uPA
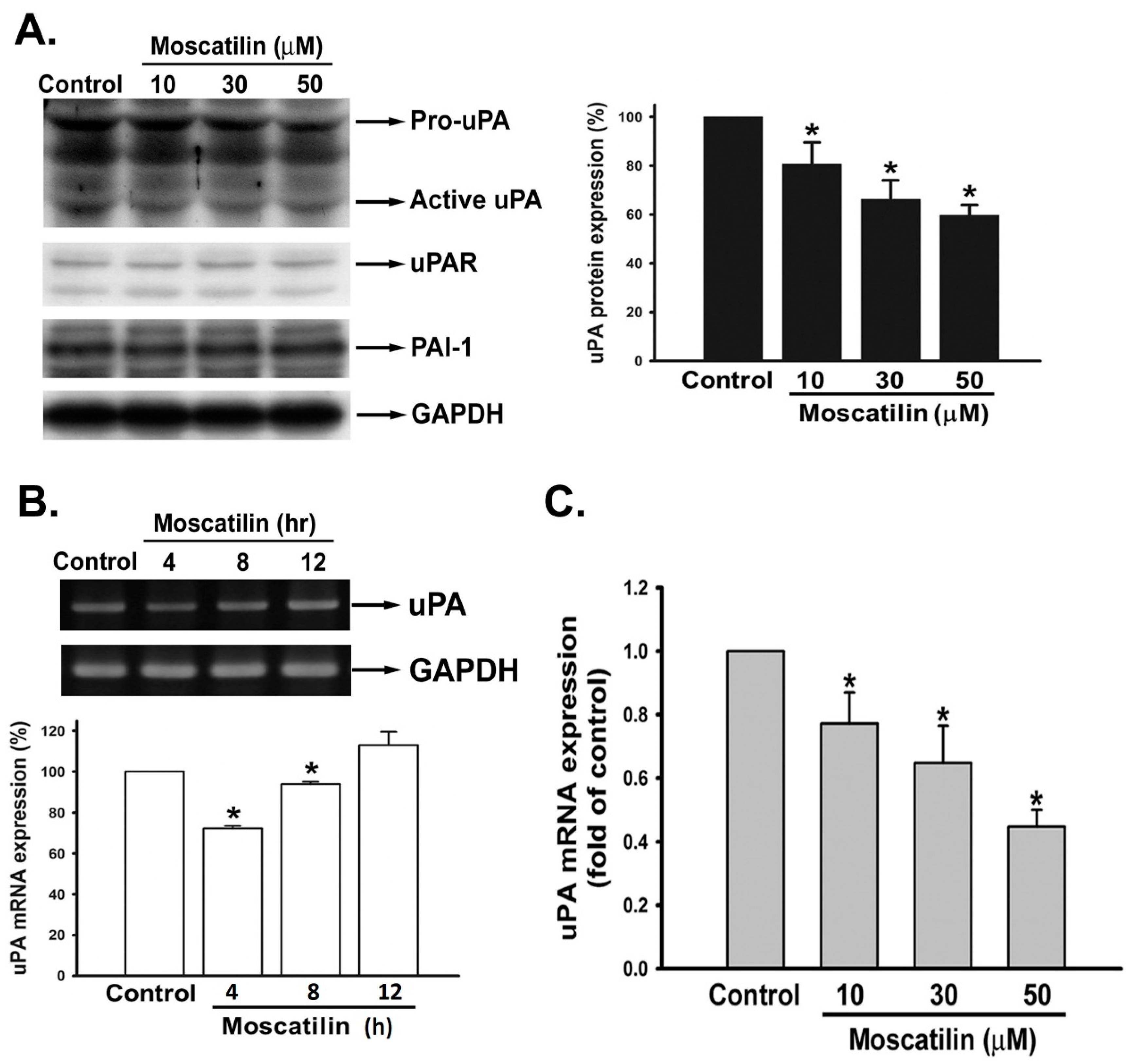
2.3. Moscatilin Suppressed uPA Expression on the Transcriptional Level
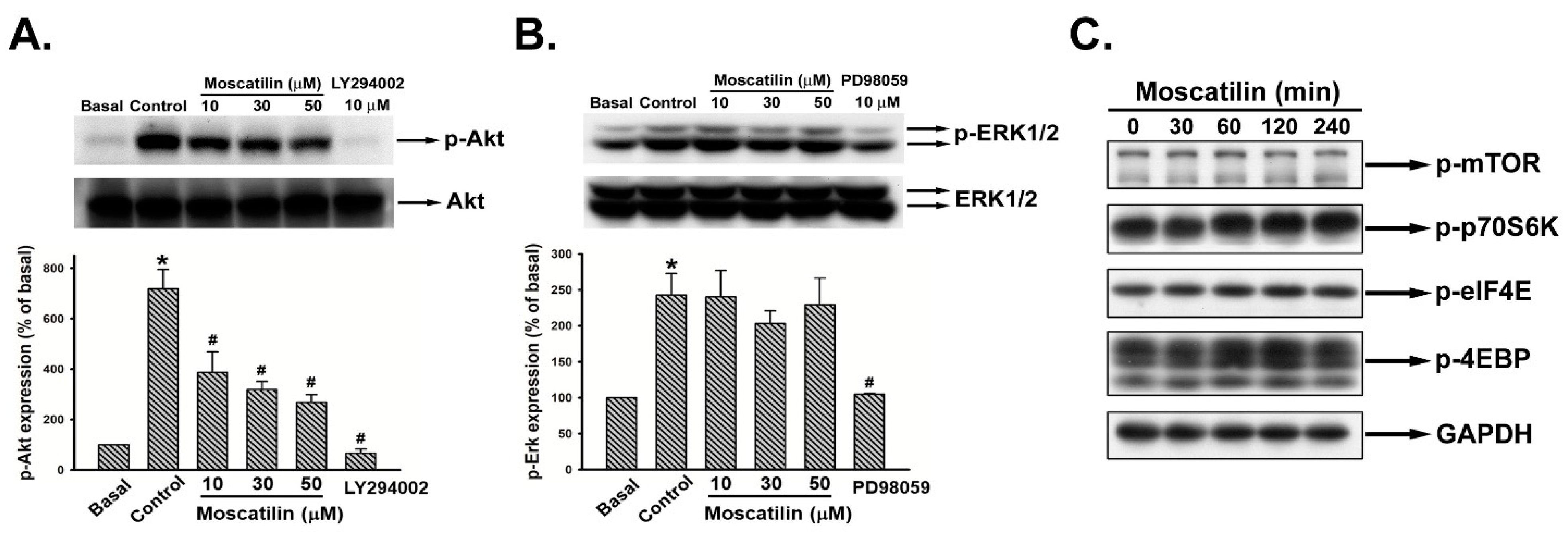
2.4. Moscatilin Reduced Akt Rather Than ERK1/2 Phosphorylation
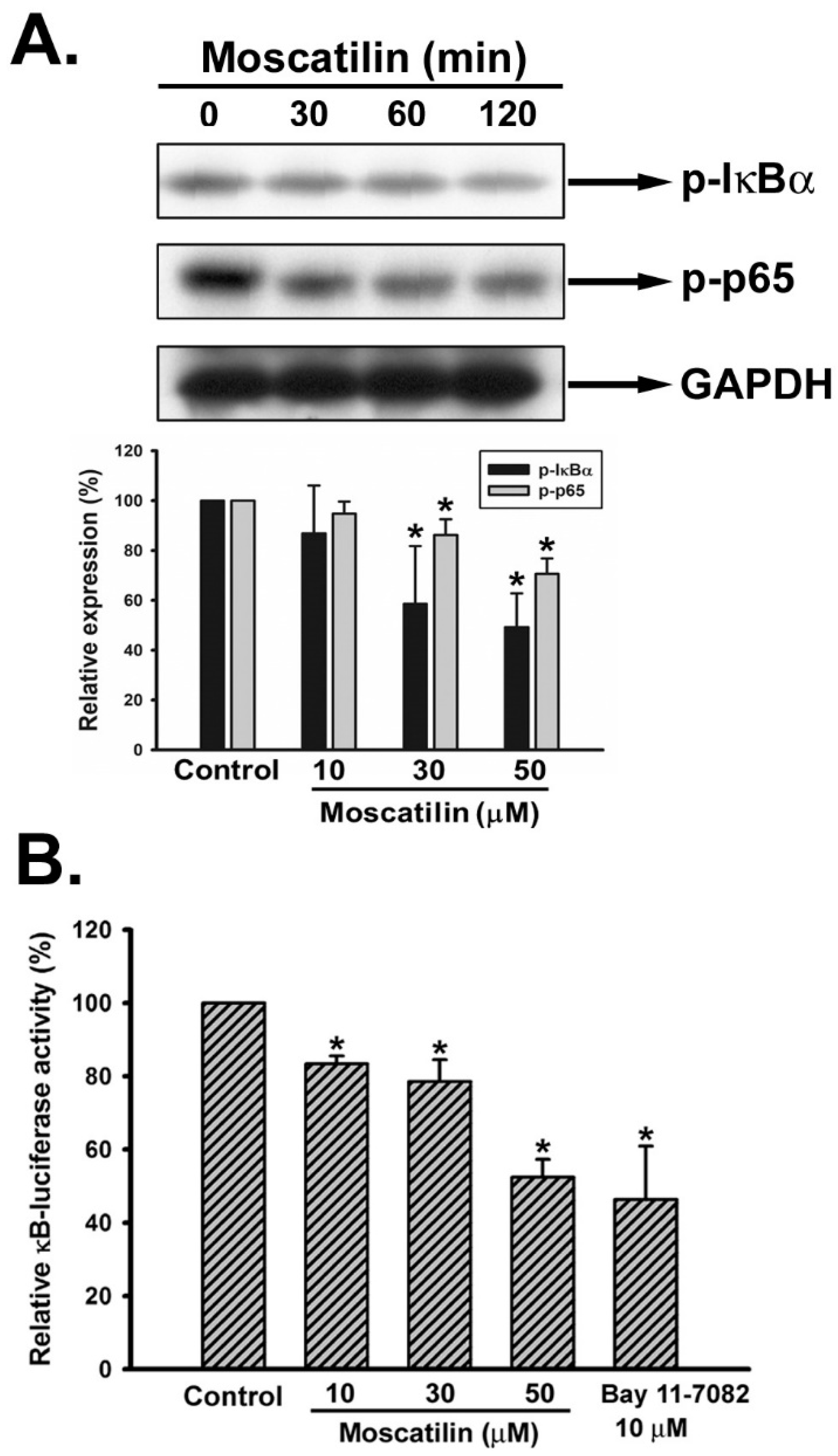
2.5. Moscatilin Repressed the Activation of NF-κB
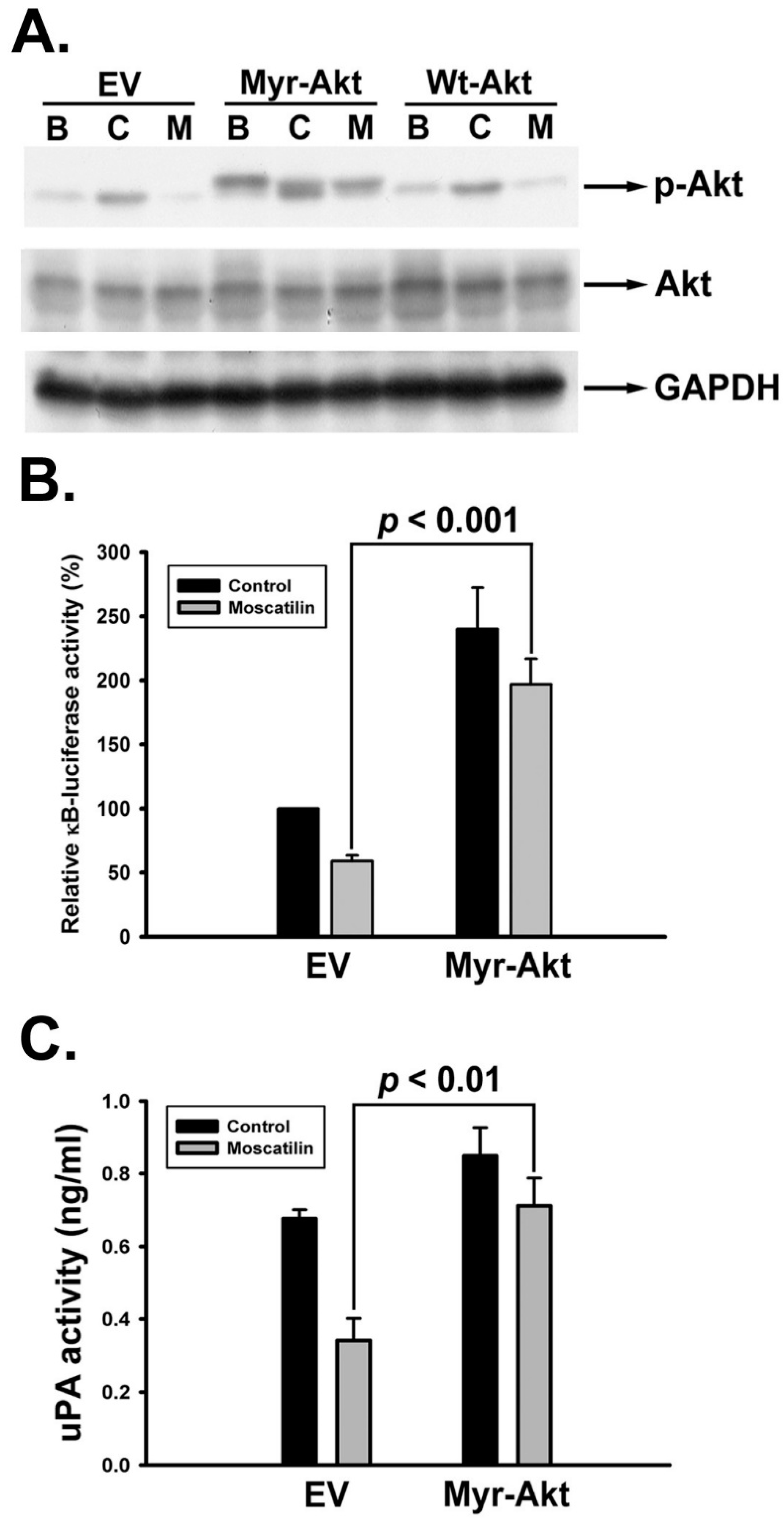
2.6. Moscatilin Impeded uPA Activation through Akt/NF-κB Signaling Pathway
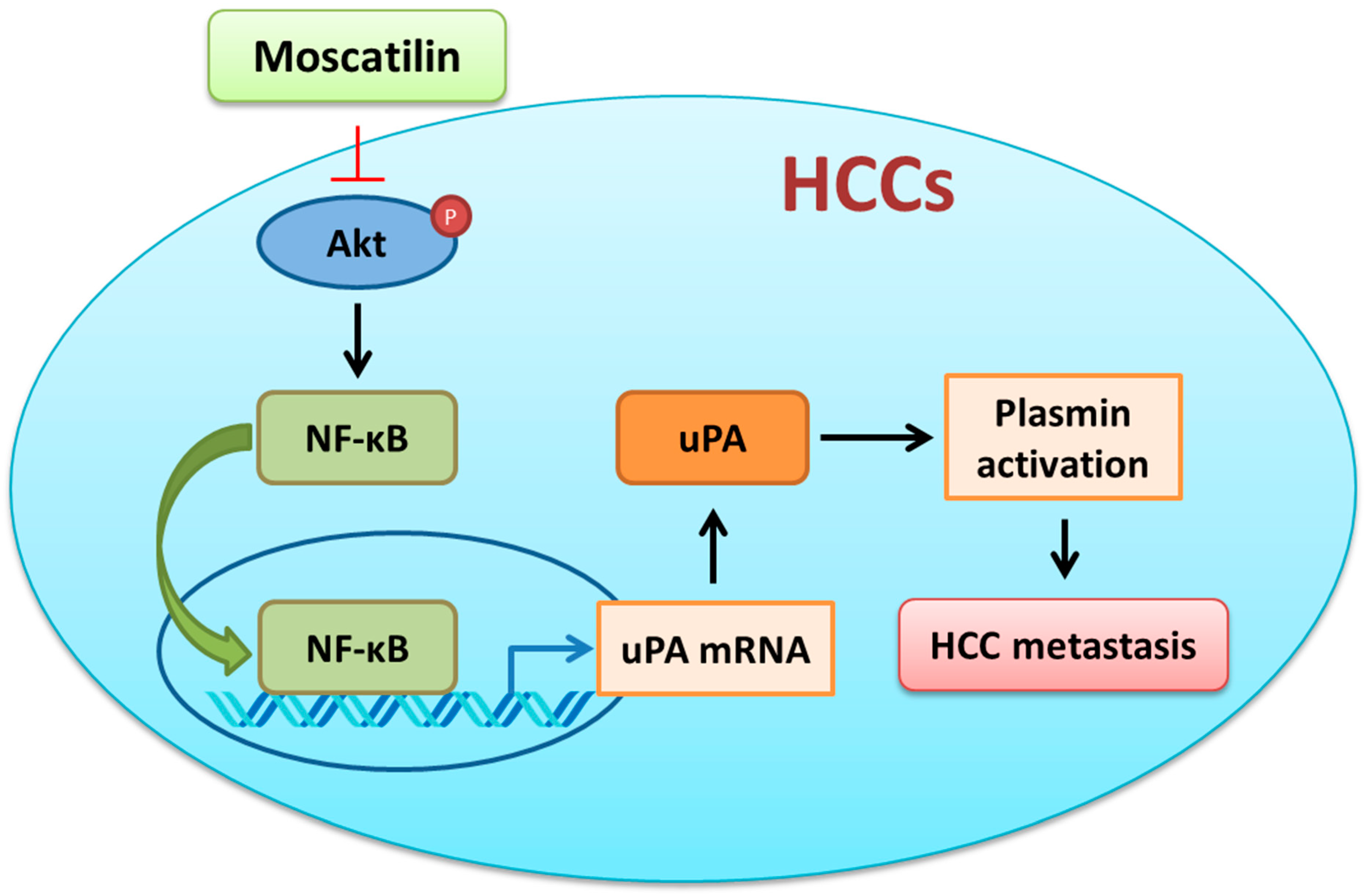
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture and Cell Viability Analysis
4.3. In Vitro Invasion Analysis
4.4. In Vivo Invasion Analysis
4.5. MMP-2, 9 and uPA Activity Assay
4.6. Western Blot Analysis
4.7. Reverse Transcription Polymerase Chain Reaction (RT-PCR) and TaqMan Quantitative Real-Time PCR Analysis
4.8. Reporter Gene Assay
4.9. Plasmids and Transient Transfection Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghouri, Y.A.; Mian, I.; Rowe, J.H. Review of hepatocellular carcinoma: Epidemiology, etiology, and carcinogenesis. J. Carcinog. 2017, 16, 1. [Google Scholar] [PubMed]
- Omata, M.; Cheng, A.L.; Kokudo, N.; Kudo, M.; Lee, J.M.; Jia, J.; Tateishi, R.; Han, K.H.; Chawla, Y.K.; Shiina, S.; et al. Asia-Pacific clinical practice guidelines on the management of hepatocellular carcinoma: A 2017 update. Hepatol. Int. 2017, 11, 317–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Serag, H.B. Epidemiology of viral hepatitis and hepatocellular carcinoma. Gastroenterology 2012, 142, 1264–1273.e1. [Google Scholar] [CrossRef] [Green Version]
- Regimbeau, J.M.; Kianmanesh, R.; Farges, O.; Dondero, F.; Sauvanet, A.; Belghiti, J. Extent of liver resection influences the outcome in patients with cirrhosis and small hepatocellular carcinoma. Surgery 2002, 131, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Valastyan, S.; Weinberg, R.A. Tumor metastasis: Molecular insights and evolving paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef]
- Dass, K.; Ahmad, A.; Azmi, A.S.; Sarkar, S.H.; Sarkar, F.H. Evolving role of uPA/uPAR system in human cancers. Cancer Treat. Rev. 2008, 34, 122–136. [Google Scholar] [CrossRef]
- Nelson, A.R.; Fingleton, B.; Rothenberg, M.L.; Matrisian, L.M. Matrix metalloproteinases: Biologic activity and clinical implications. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2000, 18, 1135–1149. [Google Scholar] [CrossRef]
- Jones, J.L.; Glynn, P.; Walker, R.A. Expression of MMP-2 and MMP-9, their inhibitors, and the activator MT1-MMP in primary breast carcinomas. J. Pathol. 1999, 189, 161–168. [Google Scholar] [CrossRef]
- Rao, J.S.; Steck, P.A.; Mohanam, S.; Stetler-Stevenson, W.G.; Liotta, L.A.; Sawaya, R. Elevated levels of M(r) 92,000 type IV collagenase in human brain tumors. Cancer Res. 1993, 53 (Suppl. 10), 2208–2211. [Google Scholar]
- Sehgal, I.; Baley, P.A.; Thompson, T.C. Transforming growth factor beta1 stimulates contrasting responses in metastatic versus primary mouse prostate cancer-derived cell lines in vitro. Cancer Res. 1996, 56, 3359–3365. [Google Scholar]
- Kuyvenhoven, J.P.; van Hoek, B.; Blom, E.; van Duijn, W.; Hanemaaijer, R.; Verheijen, J.H.; Lamers, C.B.; Verspaget, H.W. Assessment of the clinical significance of serum matrix metalloproteinases MMP-2 and MMP-9 in patients with various chronic liver diseases and hepatocellular carcinoma. Thromb. Haemost. 2003, 89, 718–725. [Google Scholar] [CrossRef] [PubMed]
- McMahon, B.J.; Kwaan, H.C. Components of the Plasminogen-Plasmin System as Biologic Markers for Cancer. Adv. Exp. Med. Biol. 2015, 867, 145–156. [Google Scholar]
- O’Rourke, J.; Jiang, X.; Hao, Z.; Cone, R.E.; Hand, A.R. Distribution of sympathetic tissue plasminogen activator (tPA) to a distant microvasculature. J. Neurosci. Res. 2005, 79, 727–733. [Google Scholar] [CrossRef]
- Mekkawy, A.H.; Pourgholami, M.H.; Morris, D.L. Involvement of urokinase-type plasminogen activator system in cancer: An overview. Med. Res. Rev. 2014, 34, 918–956. [Google Scholar] [CrossRef] [PubMed]
- Stack, M.S.; Johnson, D.A. Human mast cell tryptase activates single-chain urinary-type plasminogen activator (pro-urokinase). J. Biol. Chem. 1994, 269, 9416–9419. [Google Scholar] [CrossRef]
- Goretzki, L.; Schmitt, M.; Mann, K.; Calvete, J.; Chucholowski, N.; Kramer, M.; Gunzler, W.A.; Janicke, F.; Graeff, H. Effective activation of the proenzyme form of the urokinase-type plasminogen activator (pro-uPA) by the cysteine protease cathepsin L. FEBS Lett. 1992, 297, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Rabbani, S.A.; Xing, R.H. Role of urokinase (uPA) and its receptor (uPAR) in invasion and metastasis of hormone-dependent malignancies. Int. J. Oncol. 1998, 12, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Ellis, V.; Behrendt, N.; Dano, K. Plasminogen activation by receptor-bound urokinase. A kinetic study with both cell-associated and isolated receptor. J. Biol. Chem. 1991, 266, 12752–12758. [Google Scholar] [CrossRef]
- Hienert, G.; Kirchheimer, J.C.; Pfluger, H.; Binder, B.R. Urokinase-type plasminogen activator as a marker for the formation of distant metastases in prostatic carcinomas. J. Urol. 1988, 140, 1466–1469. [Google Scholar] [CrossRef]
- Foekens, J.A.; Schmitt, M.; van Putten, W.L.; Peters, H.A.; Bontenbal, M.; Janicke, F.; Klijn, J.G. Prognostic value of urokinase-type plasminogen activator in 671 primary breast cancer patients. Cancer Res. 1992, 52, 6101–6105. [Google Scholar]
- Tanaka, N.; Fukao, H.; Ueshima, S.; Okada, K.; Yasutomi, M.; Matsuo, O. Plasminogen activator inhibitor 1 in human carcinoma tissues. Int. J. Cancer 1991, 48, 481–484. [Google Scholar] [CrossRef] [PubMed]
- De Bruin, P.A.; Griffioen, G.; Verspaget, H.W.; Verheijen, J.H.; Lamers, C.B. Plasminogen activators and tumor development in the human colon: Activity levels in normal mucosa, adenomatous polyps, and adenocarcinomas. Cancer Res. 1987, 47, 4654–4657. [Google Scholar]
- Kuhn, W.; Pache, L.; Schmalfeldt, B.; Dettmar, P.; Schmitt, M.; Janicke, F.; Graeff, H. Urokinase (uPA) and PAI-1 predict survival in advanced ovarian cancer patients (FIGO III) after radical surgery and platinum-based chemotherapy. Gynecol. Oncol. 1994, 55 (3 Pt 1), 401–409. [Google Scholar] [CrossRef]
- Zheng, Q.; Tang, Z.Y.; Xue, Q.; Shi, D.R.; Song, H.Y.; Tang, H.B. Invasion and metastasis of hepatocellular carcinoma in relation to urokinase-type plasminogen activator, its receptor and inhibitor. J. Cancer Res. Clin. Oncol. 2000, 126, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.N.; Pan, S.L.; Peng, C.Y.; Huang, D.Y.; Guh, J.H.; Chen, C.C.; Shen, C.C.; Teng, C.M. Moscatilin repressed lipopolysaccharide-induced HIF-1alpha accumulation and NF-kappaB activation in murine RAW264.7 cells. Shock 2010, 33, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Kowitdamrong, A.; Chanvorachote, P.; Sritularak, B.; Pongrakhananon, V. Moscatilin inhibits lung cancer cell motility and invasion via suppression of endogenous reactive oxygen species. BioMed Res. Int. 2013, 2013, 765894. [Google Scholar] [CrossRef] [Green Version]
- Tsai, A.C.; Pan, S.L.; Liao, C.H.; Guh, J.H.; Wang, S.W.; Sun, H.L.; Liu, Y.N.; Chen, C.C.; Shen, C.C.; Chang, Y.L.; et al. Moscatilin, a bibenzyl derivative from the India orchid Dendrobrium loddigesii, suppresses tumor angiogenesis and growth in vitro and in vivo. Cancer Lett. 2010, 292, 163–170. [Google Scholar] [CrossRef]
- Chen, C.A.; Chen, C.C.; Shen, C.C.; Chang, H.H.; Chen, Y.J. Moscatilin induces apoptosis and mitotic catastrophe in human esophageal cancer cells. J. Med. Food 2013, 16, 869–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, H.C.; Chang, L.H.; Peng, C.Y.; Chang, Y.L.; Chen, C.C.; Shen, C.C.; Teng, C.M.; Pan, S.L. Moscatilin inhibits migration and metastasis of human breast cancer MDA-MB-231 cells through inhibition of Akt and Twist signaling pathway. J. Mol. Med. (Berl.) 2013, 91, 347–356. [Google Scholar] [CrossRef]
- Busaranon, K.; Plaimee, P.; Sritularak, B.; Chanvorachote, P. Moscatilin inhibits epithelial-to-mesenchymal transition and sensitizes anoikis in human lung cancer H460 cells. J. Nat. Med. 2016, 70, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, N.; Mohanam, S.; Gujrati, M.; Olivero, W.C.; Dinh, D.H.; Rao, J.S. Downregulation of uPA inhibits migration and PI3k/Akt signaling in glioblastoma cells. Oncogene 2003, 22, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Iiizumi, M.; Liu, W.; Pai, S.K.; Furuta, E.; Watabe, K. Drug development against metastasis-related genes and their pathways: A rationale for cancer therapy. Biochim. Biophys. Acta 2008, 1786, 87–104. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Abbruzzese, J.L.; Evans, D.B.; Chiao, P.J. Overexpression of urokinase-type plasminogen activator in pancreatic adenocarcinoma is regulated by constitutively activated RelA. Oncogene 1999, 18, 4554–4563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novikova, M.V.; Khromova, N.V.; Kopnin, P.B. Components of the Hepatocellular Carcinoma Microenvironment and Their Role in Tumor Progression. Biochemistry 2017, 82, 861–873. [Google Scholar] [CrossRef] [PubMed]
- Choong, P.F.; Nadesapillai, A.P. Urokinase plasminogen activator system: A multifunctional role in tumor progression and metastasis. Clin. Orthop. Relat. Res. 2003, (Suppl. 415), S46–S58. [Google Scholar] [CrossRef]
- Higazi, A.; Cohen, R.L.; Henkin, J.; Kniss, D.; Schwartz, B.S.; Cines, D.B. Enhancement of the enzymatic activity of single-chain urokinase plasminogen activator by soluble urokinase receptor. J. Biol. Chem. 1995, 270, 17375–17380. [Google Scholar] [CrossRef] [Green Version]
- Andreasen, P.A.; Georg, B.; Lund, L.R.; Riccio, A.; Stacey, S.N. Plasminogen activator inhibitors: Hormonally regulated serpins. Mol. Cell. Endocrinol. 1990, 68, 1–19. [Google Scholar] [CrossRef]
- Irigoyen, J.P.; Munoz-Canoves, P.; Montero, L.; Koziczak, M.; Nagamine, Y. The plasminogen activator system: Biology and regulation. Cell. Mol. Life Sci. Cmls 1999, 56, 104–132. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, Y.; Medcalf, R.L.; Munoz-Canoves, P. Transcriptional and posttranscriptional regulation of the plasminogen activator system. Thromb. Haemost. 2005, 93, 661–675. [Google Scholar] [PubMed]
- Benasciutti, E.; Pages, G.; Kenzior, O.; Folk, W.; Blasi, F.; Crippa, M.P. MAPK and JNK transduction pathways can phosphorylate Sp1 to activate the uPA minimal promoter element and endogenous gene transcription. Blood 2004, 104, 256–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irigoyen, J.P.; Besser, D.; Nagamine, Y. Cytoskeleton reorganization induces the urokinase-type plasminogen activator gene via the Ras/extracellular signal-regulated kinase (ERK) signaling pathway. J. Biol. Chem. 1997, 272, 1904–1909. [Google Scholar] [CrossRef] [Green Version]
- Hansen, S.K.; Nerlov, C.; Zabel, U.; Verde, P.; Johnsen, M.; Baeuerle, P.A.; Blasi, F. A novel complex between the p65 subunit of NF-kappa B and c-Rel binds to a DNA element involved in the phorbol ester induction of the human urokinase gene. EMBO J. 1992, 11, 205–213. [Google Scholar] [CrossRef]
- Cai, Z.; Tchou-Wong, K.M.; Rom, W.N. NF-kappaB in lung tumorigenesis. Cancers 2011, 3, 4258–4268. [Google Scholar] [CrossRef] [Green Version]
- Pan, G.; Zhu, Z.; Huang, J.; Yang, C.; Yang, Y.; Wang, Y.; Tuo, X.; Su, G.; Zhang, X.; Yang, Z.; et al. Semaphorin 5A promotes gastric cancer invasion/metastasis via urokinase-type plasminogen activator/phosphoinositide 3-kinase/protein kinase B. Dig. Dis. Sci. 2013, 58, 2197–2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.C.; Chen, C.; Chung, C.H.; Wang, P.C.; Wu, N.L.; Cheng, J.K.; Lai, Y.W.; Sun, H.L.; Peng, C.Y.; Tang, C.H.; et al. Inhibitory effects of butein on cancer metastasis and bioenergetic modulation. J. Agric. Food Chem. 2014, 62, 9109–9117. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, C.-L.; Weng, M.-S.; Chen, W.-C.; Chien, K.-T.; Chi, C.-W.; Chung, C.-H.; Huang, C.-W.; Wang, P.-C.; Chen, C.-C.; Tsai, A.-C.; et al. Moscatilin Inhibits Metastatic Behavior of Human Hepatocellular Carcinoma Cells: A Crucial Role of uPA Suppression via Akt/NF-κB-Dependent Pathway. Int. J. Mol. Sci. 2021, 22, 2930. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22062930
Yu C-L, Weng M-S, Chen W-C, Chien K-T, Chi C-W, Chung C-H, Huang C-W, Wang P-C, Chen C-C, Tsai A-C, et al. Moscatilin Inhibits Metastatic Behavior of Human Hepatocellular Carcinoma Cells: A Crucial Role of uPA Suppression via Akt/NF-κB-Dependent Pathway. International Journal of Molecular Sciences. 2021; 22(6):2930. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22062930
Chicago/Turabian StyleYu, Chen-Lin, Meng-Shih Weng, Wei-Cheng Chen, Kai-Ting Chien, Chih-Wen Chi, Ching-Hu Chung, Chia-Wen Huang, Po-Chuan Wang, Chien-Chih Chen, An-Chi Tsai, and et al. 2021. "Moscatilin Inhibits Metastatic Behavior of Human Hepatocellular Carcinoma Cells: A Crucial Role of uPA Suppression via Akt/NF-κB-Dependent Pathway" International Journal of Molecular Sciences 22, no. 6: 2930. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22062930