
Prognostic Value of microRNA-221/2 and 17-92 Families in Primary Glioblastoma Patients Treated with Postoperative Radiotherapy

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Patient Characteristics
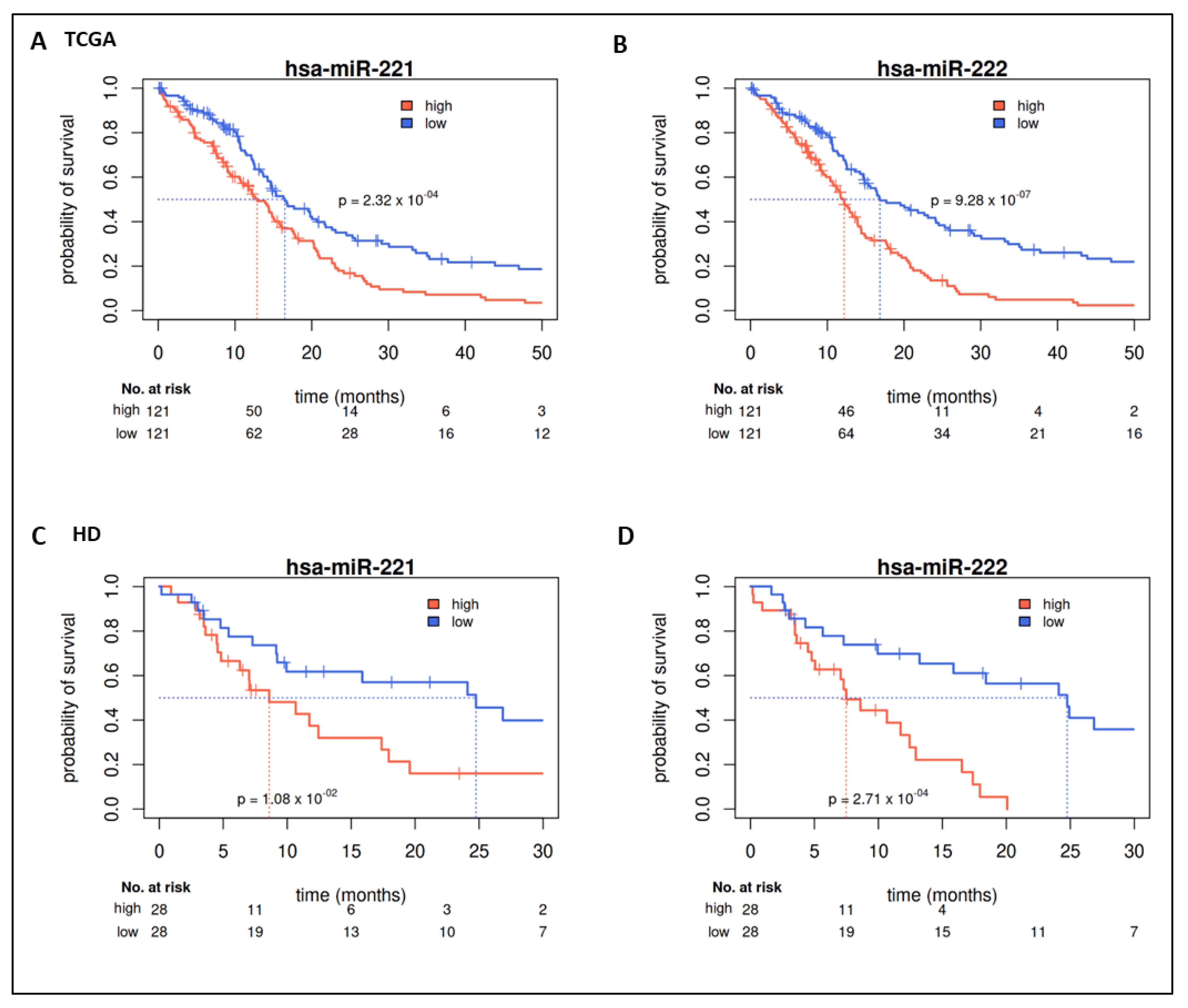
2.2. Prognostic Value of miRs in TCGA
2.3. Validation in HD Cohort
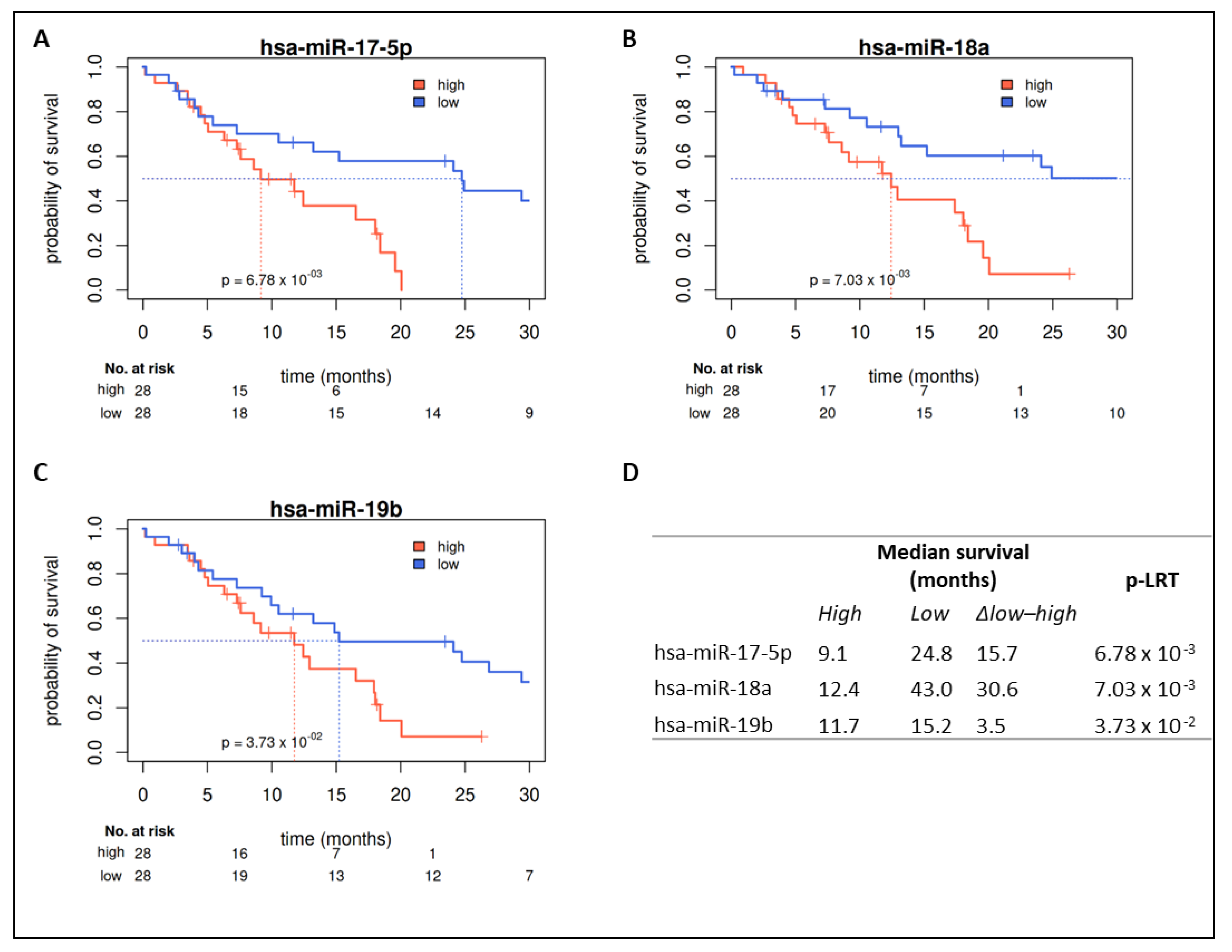
2.4. Prognostic Value in Covariate Stratified Subcohorts
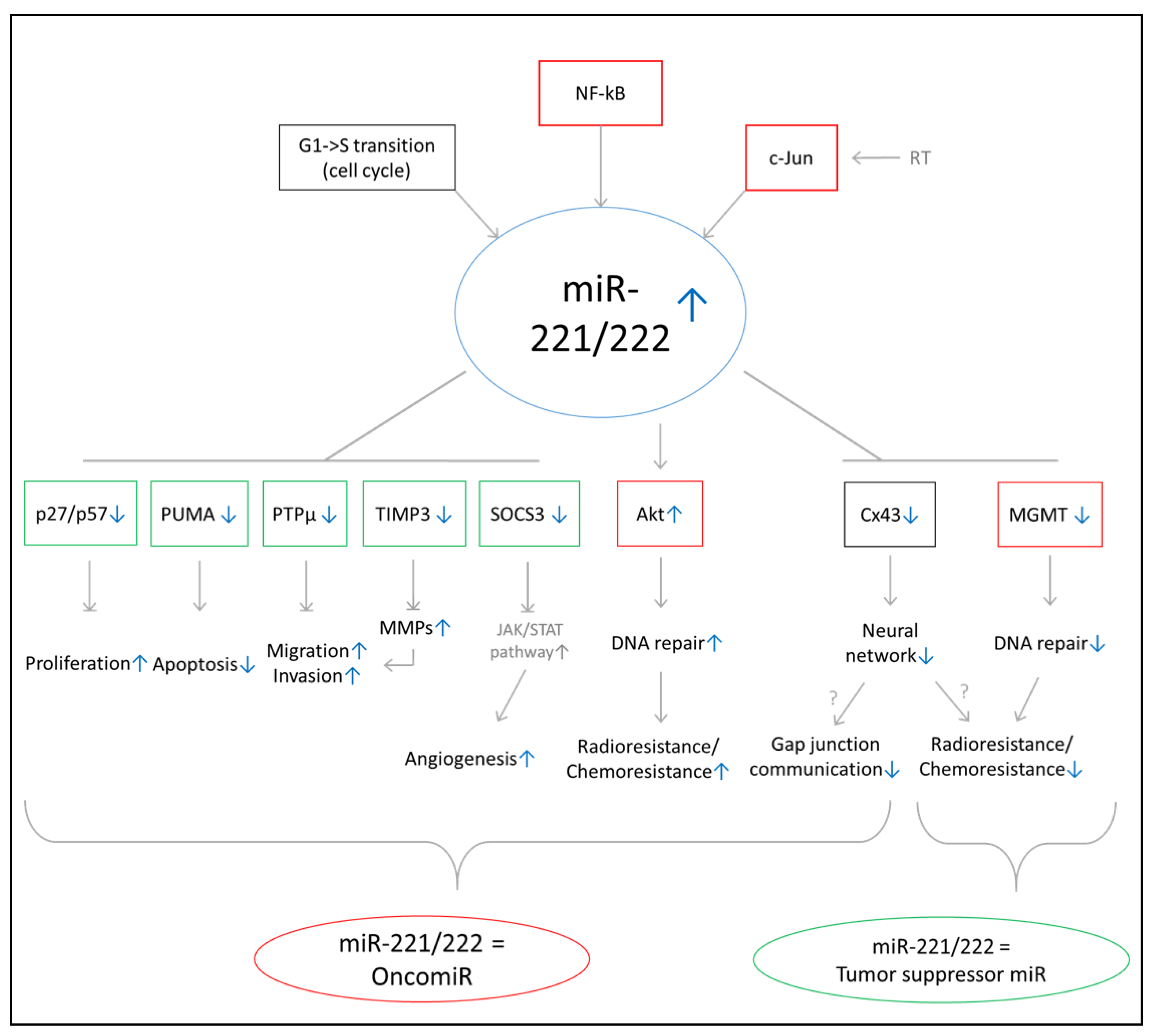
3. Discussion
4. Materials and Methods
4.1. TCGA GBM Collective
4.2. Heidelberg GBM Collective
4.2.1. Patients and Tumor Samples
4.2.2. Ethics Approval and Consent to Participate
4.2.3. Patient Material
4.2.4. Nucleic Acid Isolation
4.2.5. microRNA Analysis
4.3. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GBM | glioblastoma multiforme |
| HD | Heidelberg |
| MGMT | O-6-methylguanine-DNA methyltransferase |
| miR | microRNA |
| OS | overall survival |
| RT | radiotherapy |
| TCGA | The Cancer Genome Atlas |
References
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostrom, Q.T.; Gittleman, H.; Fulop, J.; Liu, M.; Blanda, R.; Kromer, C.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Central Nervous System Tumors Diagnosed in the United States in 2008–2012. Neuro Oncol. 2015, 17, iv1–iv62. [Google Scholar] [CrossRef]
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.D.; Green, S.B.; Byar, D.P.; Alexander, E., Jr.; Batzdorf, U.; Brooks, W.H.; Hunt, W.E.; MacCarty, C.S.; Mahaley, M.S., Jr.; Mealey, J., Jr.; et al. Randomized comparisons of radiotherapy and nitrosoureas for the treatment of malignant glioma after surgery. N. Engl. J. Med. 1980, 303, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Laperriere, N.; Zuraw, L.; Cairncross, G.; Cancer Care Ontario Practice Guidelines Initiative Neuro-Oncology Disease Site Group. Radiotherapy for newly diagnosed malignant glioma in adults: A systematic review. Radiother. Oncol. 2002, 64, 259–273. [Google Scholar] [CrossRef]
- Deutsche Gesellschaft für Neurologie Leitlinien für Diagnostik und Therapie in der Neurologie: Gliome 2015. Available online: https://www.awmf.org/uploads/tx_szleitlinien/030-099l_S2k_Gliome_2015-06-abgelaufen.pdf (accessed on 1 December 2020).
- Sanson, M.; Marie, Y.; Paris, S.; Idbaih, A.; Laffaire, J.; Ducray, F.; El Hallani, S.; Boisselier, B.; Mokhtari, K.; Hoang-Xuan, K.; et al. Isocitrate dehydrogenase 1 codon 132 mutation is an important prognostic biomarker in gliomas. J. Clin. Oncol. 2009, 27, 4150–4154. [Google Scholar] [CrossRef] [PubMed]
- Roa, W.; Brasher, P.M.; Bauman, G.; Anthes, M.; Bruera, E.; Chan, A.; Fisher, B.; Fulton, D.; Gulavita, S.; Hao, C.; et al. Abbreviated course of radiation therapy in older patients with glioblastoma multiforme: A prospective randomized clinical trial. J. Clin. Oncol. 2004, 22, 1583–1588. [Google Scholar] [CrossRef]
- Keime-Guibert, F.; Chinot, O.; Taillandier, L.; Cartalat-Carel, S.; Frenay, M.; Kantor, G.; Guillamo, J.-S.; Jadaud, E.; Colin, P.; Bondiau, P.-Y.; et al. Radiotherapy for glioblastoma in the elderly. N. Engl. J. Med. 2007, 356, 1527–1535. [Google Scholar] [CrossRef]
- Perry, J.R.; Laperriere, N.; O’Callaghan, C.J.; Brandes, A.A.; Menten, J.; Phillips, C.; Fay, M.; Nishikawa, R.; Cairncross, J.G.; Roa, W.; et al. Short-Course Radiation plus Temozolomide in Elderly Patients with Glioblastoma. N. Engl. J. Med. 2017, 376, 1027–1037. [Google Scholar] [CrossRef] [Green Version]
- Malmström, A.; Grønberg, B.H.; Marosi, C.; Stupp, R.; Frappaz, D.; Schultz, H.; Abacioglu, U.; Tavelin, B.; Lhermitte, B.; Hegi, M.E.; et al. Temozolomide versus standard 6-week radiotherapy versus hypofractionated radiotherapy in patients older than 60 years with glioblastoma: The Nordic randomised, phase 3 trial. Lancet Oncol. 2012, 13, 916–926. [Google Scholar] [CrossRef]
- Wick, W.; Platten, M.; Meisner, C.; Felsberg, J.; Tabatabai, G.; Simon, M.; Nikkhah, G.; Papsdorf, K.; Steinbach, J.P.; Sabel, M.; et al. Temozolomide chemotherapy alone versus radiotherapy alone for malignant astrocytoma in the elderly: The NOA-08 randomised, phase 3 trial. Lancet Oncol. 2012, 13, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Parsons, D.W.; Jin, G.; McLendon, R.; Rasheed, B.A.; Yuan, W.; Kos, I.; Batinic-Haberle, I.; Jones, S.; Riggins, G.J.; et al. IDH1 and IDH2 mutations in gliomas. N. Engl. J. Med. 2009, 360, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Parsons, D.W.; Jones, S.; Zhang, X.; Lin, J.C.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Siu, I.M.; Gallia, G.L.; et al. An integrated genomic analysis of human glioblastoma multiforme. Science 2008, 321, 1807–1812. [Google Scholar] [CrossRef] [Green Version]
- Krex, D.; Klink, B.; Hartmann, C.; von Deimling, A.; Pietsch, T.; Simon, M.; Sabel, M.; Steinbach, J.P.; Heese, O.; Reifenberger, G.; et al. Long-term survival with glioblastoma multiforme. Brain 2007, 130, 2596–2606. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; Felsberg, J.; Hartmann, C.; Berger, H.; Steinbach, J.P.; Schramm, J.; Westphal, M.; Schackert, G.; Simon, M.; Tonn, J.C.; et al. Molecular predictors of progression-free and overall survival in patients with newly diagnosed glioblastoma: A prospective translational study of the German Glioma Network. J. Clin. Oncol. 2009, 27, 5743–5750. [Google Scholar] [CrossRef] [Green Version]
- Weller, M.; Wick, W.; von Deimling, A. Isocitrate dehydrogenase mutations: A challenge to traditional views on the genesis and malignant progression of gliomas. Glia 2011, 59, 1200–1204. [Google Scholar] [CrossRef]
- Capper, D.; Jones, D.T.W.; Sill, M.; Hovestadt, V.; Schrimpf, D.; Sturm, D.; Koelsche, C.; Sahm, F.; Chavez, L.; Reuss, D.E.; et al. DNA methylation-based classification of central nervous system tumours. Nature 2018, 555, 469–474. [Google Scholar] [CrossRef]
- Knoll, M.; Debus, J.; Furkel, J.; Warta, R.; Bougatf, N.; Rapp, C.; Brors, B.; Wick, W.; Unterberg, A.; Herold-Mende, C.; et al. Glioblastoma evolution pattern under surgery and radio(chemo)therapy (RCHT) to identify novel methylome based glioma subtypes. J. Clin. Oncol. 2019, 37, 2012. [Google Scholar] [CrossRef]
- Geisenberger, C.; Mock, A.; Warta, R.; Rapp, C.; Schwager, C.; Korshunov, A.; Nied, A.K.; Capper, D.; Brors, B.; Jungk, C.; et al. Molecular profiling of long-term survivors identifies a subgroup of glioblastoma characterized by chromosome 19/20 co-gain. Acta Neuropathol. 2015, 130, 419–434. [Google Scholar] [CrossRef]
- Hussein, K. Pathobiologie des microRNA-Systems. Pathologe 2012, 33, 70–78. [Google Scholar] [CrossRef]
- Brower, J.V.; Clark, P.A.; Lyon, W.; Kuo, J.S. MicroRNAs in cancer: Glioblastoma and glioblastoma cancer stem cells. Neurochem. Int. 2014, 77, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Almog, N.; Ma, L.; Raychowdhury, R.; Schwager, C.; Erber, R.; Short, S.; Hlatky, L.; Vajkoczy, P.; Huber, P.E.; Folkman, J.; et al. Transcriptional switch of dormant tumors to fast-growing angiogenic phenotype. Cancer Res. 2009, 69, 836–844. [Google Scholar] [CrossRef] [Green Version]
- Almog, N.; Ma, L.; Schwager, C.; Brinkmann, B.G.; Beheshti, A.; Vajkoczy, P.; Folkman, J.; Hlatky, L.; Abdollahi, A. Consensus micro RNAs governing the switch of dormant tumors to the fast-growing angiogenic phenotype. PLoS ONE 2012, 7, e44001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazurek, M.; Grochowski, C.; Litak, J.; Osuchowska, I.; Maciejewski, R.; Kamieniak, P. Recent Trends of microRNA Significance in Pediatric Population Glioblastoma and Current Knowledge of Micro RNA Function in Glioblastoma Multiforme. Int. J. Mol. Sci. 2020, 21, 3046. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature 2008, 455, 1061–1068. [CrossRef]
- Garofalo, M.; Quintavalle, C.; Romano, G.; Croce, C.M.; Condorelli, G. miR221/222 in cancer: Their role in tumor progression and response to therapy. Curr. Mol. Med. 2012, 12, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zheng, J.; Chen, L.; Diao, H.; Liu, Y. Predictive and Prognostic Roles of Abnormal Expression of Tissue miR-125b, miR-221, and miR-222 in Glioma. Mol. Neurobiol. 2016, 53, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Lakomy, R.; Sana, J.; Hankeova, S.; Fadrus, P.; Kren, L.; Lzicarova, E.; Svoboda, M.; Dolezelova, H.; Smrcka, M.; Vyzula, R.; et al. MiR-195, miR-196b, miR-181c, miR-21 expression levels and O-6-methylguanine-DNA methyltransferase methylation status are associated with clinical outcome in glioblastoma patients. Cancer Sci. 2011, 102, 2186–2190. [Google Scholar] [CrossRef]
- Ciafrè, S.A.; Galardi, S.; Mangiola, A.; Ferracin, M.; Liu, C.G.; Sabatino, G.; Negrini, M.; Maira, G.; Croce, C.M.; Farace, M.G. Extensive modulation of a set of microRNAs in primary glioblastoma. Biochem. Biophys. Res. Commun. 2005, 334, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Conti, A.; Aguennouz, M.; La Torre, D.; Tomasello, C.; Cardali, S.; Angileri, F.F.; Maio, F.; Cama, A.; Germano, A.; Vita, G.; et al. miR-21 and 221 upregulation and miR-181b downregulation in human grade II-IV astrocytic tumors. J. Neurooncol. 2009, 93, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Quintavalle, C.; Garofalo, M.; Zanca, C.; Romano, G.; Iaboni, M.; del Basso De Caro, M.; Martinez-Montero, J.C.; Incoronato, M.; Nuovo, G.; Croce, C.M.; et al. miR-221/222 overexpession in human glioblastoma increases invasiveness by targeting the protein phosphate PTPmu. Oncogene 2012, 31, 858–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhang, J.; Hao, J.; Shi, Z.; Wang, Y.; Han, L.; Yu, S.; You, Y.; Jiang, T.; Wang, J.; et al. High level of miR-221/222 confers increased cell invasion and poor prognosis in glioma. J. Transl. Med. 2012, 10, 119. [Google Scholar] [CrossRef] [Green Version]
- le Sage, C.; Nagel, R.; Egan, D.A.; Schrier, M.; Mesman, E.; Mangiola, A.; Anile, C.; Maira, G.; Mercatelli, N.; Ciafre, S.A.; et al. Regulation of the p27(Kip1) tumor suppressor by miR-221 and miR-222 promotes cancer cell proliferation. EMBO J. 2007, 26, 3699–3708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, R.; Zaidi, S.K.; Liu, C.G.; Stein, J.L.; van Wijnen, A.J.; Croce, C.M.; Stein, G.S. MicroRNAs 221 and 222 bypass quiescence and compromise cell survival. Cancer Res. 2008, 68, 2773–2780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Kang, C.; You, Y.; Pu, P.; Yang, W.; Zhao, P.; Wang, G.; Zhang, A.; Jia, Z.; Han, L.; et al. Co-suppression of miR-221/222 cluster suppresses human glioma cell growth by targeting p27kip1 in vitro and in vivo. Int. J. Oncol. 2009, 34, 1653–1660. [Google Scholar] [PubMed] [Green Version]
- Zhang, C.Z.; Zhang, J.X.; Zhang, A.L.; Shi, Z.D.; Han, L.; Jia, Z.F.; Yang, W.D.; Wang, G.X.; Jiang, T.; You, Y.P.; et al. MiR-221 and miR-222 target PUMA to induce cell survival in glioblastoma. Mol. Cancer 2010, 9, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.H.; Liu, Y.; Xiao, L.M.; Chen, L.K.; Zheng, S.Y.; Zeng, E.M.; Li, D.H.; Li, Y.P. Silencing microRNA-221/222 cluster suppresses glioblastoma angiogenesis by suppressor of cytokine signaling-3-dependent JAK/STAT pathway. J. Cell. Physiol. 2019, 234, 22272–22284. [Google Scholar] [CrossRef]
- Quintavalle, C.; Mangani, D.; Roscigno, G.; Romano, G.; Diaz-Lagares, A.; Iaboni, M.; Donnarumma, E.; Fiore, D.; De Marinis, P.; Soini, Y.; et al. MiR-221/222 target the DNA methyltransferase MGMT in glioma cells. PLoS ONE 2013, 8, e74466. [Google Scholar] [CrossRef] [Green Version]
- Munoz, J.L.; Bliss, S.A.; Greco, S.J.; Ramkissoon, S.H.; Ligon, K.L.; Rameshwar, P. Delivery of Functional Anti-miR-9 by Mesenchymal Stem Cell-derived Exosomes to Glioblastoma Multiforme Cells Conferred Chemosensitivity. Mol. Ther. Nucleic Acids 2013, 2, e126. [Google Scholar] [CrossRef] [PubMed]
- Osswald, M.; Jung, E.; Sahm, F.; Solecki, G.; Venkataramani, V.; Blaes, J.; Weil, S.; Horstmann, H.; Wiestler, B.; Syed, M.; et al. Brain tumour cells interconnect to a functional and resistant network. Nature 2015, 528, 93–98. [Google Scholar] [CrossRef]
- Gielen, P.R.; Aftab, Q.; Ma, N.; Chen, V.C.; Hong, X.; Lozinsky, S.; Naus, C.C.; Sin, W.C. Connexin43 confers Temozolomide resistance in human glioma cells by modulating the mitochondrial apoptosis pathway. Neuropharmacology 2013, 75, 539–548. [Google Scholar] [CrossRef]
- Sun, B.; Zhao, X.; Ming, J.; Liu, X.; Liu, D.; Jiang, C. Stepwise detection and evaluation reveal miR-10b and miR-222 as a remarkable prognostic pair for glioblastoma. Oncogene 2019, 38, 6142–6157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yerukala Sathipati, S.; Huang, H.L.; Ho, S.Y. Estimating survival time of patients with glioblastoma multiforme and characterization of the identified microRNA signatures. BMC Genom. 2016, 17, 1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delfino, K.R.; Serao, N.V.; Southey, B.R.; Rodriguez-Zas, S.L. Therapy-, gender- and race-specific microRNA markers, target genes and networks related to glioblastoma recurrence and survival. Cancer Genom. Proteom. 2011, 8, 173–183. [Google Scholar]
- Chen, Y.Y.; Ho, H.L.; Lin, S.C.; Ho, T.D.; Hsu, C.Y. Upregulation of miR-125b, miR-181d, and miR-221 Predicts Poor Prognosis in MGMT Promoter-Unmethylated Glioblastoma Patients. Am. J. Clin. Pathol. 2018, 149, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Pang, B.; Xin, T.; Guo, H.; Xing, Y.; Xu, S.; Feng, B.; Liu, B.; Pang, Q. Plasma miR-221/222 Family as Novel Descriptive and Prognostic Biomarkers for Glioma. Mol. Neurobiol. 2015. [Google Scholar] [CrossRef]
- Swellam, M.; Ezz El Arab, L.; Al-Posttany, A.S.; Said, B.S. Clinical impact of circulating oncogenic MiRNA-221 and MiRNA-222 in glioblastoma multiform. J. Neurooncol. 2019, 144, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Shen, J.; Hodges, T.R.; Song, R.; Fuller, G.N.; Heimberger, A.B. Serum microRNA profiling in patients with glioblastoma: A survival analysis. Mol. Cancer 2017, 16, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Liu, S.; Sun, G.P.; Wang, F.; Zou, Y.F.; Jiao, Y.; Ning, J.; Xu, J. Prognostic significance of microRNA-221/222 expression in cancers: Evidence from 1,204 subjects. Int. J. Biol. Markers 2014, 29, e129–e141. [Google Scholar] [CrossRef] [PubMed]
- Ernst, A.; Campos, B.; Meier, J.; Devens, F.; Liesenberg, F.; Wolter, M.; Reifenberger, G.; Herold-Mende, C.; Lichter, P.; Radlwimmer, B. De-repression of CTGF via the miR-17-92 cluster upon differentiation of human glioblastoma spheroid cultures. Oncogene 2010, 29, 3411–3422. [Google Scholar] [CrossRef] [Green Version]
- Lavon, I.; Zrihan, D.; Granit, A.; Einstein, O.; Fainstein, N.; Cohen, M.A.; Cohen, M.A.; Zelikovitch, B.; Shoshan, Y.; Spektor, S.; et al. Gliomas display a microRNA expression profile reminiscent of neural precursor cells. Neuro Oncol. 2010, 12, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Malzkorn, B.; Wolter, M.; Liesenberg, F.; Grzendowski, M.; Stuhler, K.; Meyer, H.E.; Reifenberger, G. Identification and functional characterization of microRNAs involved in the malignant progression of gliomas. Brain Pathol. 2010, 20, 539–550. [Google Scholar] [CrossRef]
- Dews, M.; Fox, J.L.; Hultine, S.; Sundaram, P.; Wang, W.; Liu, Y.Y.; Furth, E.; Enders, G.H.; El-Deiry, W.; Schelter, J.M.; et al. The myc-miR-17~92 axis blunts TGFβ signaling and production of multiple TGFβ-dependent antiangiogenic factors. Cancer Res. 2010, 70, 8233–8246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yang, B.B. Stress response of glioblastoma cells mediated by miR-17-5p targeting PTEN and the passenger strand miR-17-3p targeting MDM2. Oncotarget 2012, 3, 1653–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Pan, J.Q.; Luo, L.; Ning, X.J.; Ye, Z.P.; Yu, Z.; Li, W.S. NF-kappaB induces miR-148a to sustain TGF-β/Smad signaling activation in glioblastoma. Mol. Cancer 2015, 14, 2. [Google Scholar] [CrossRef] [Green Version]
- Comincini, S.; Allavena, G.; Palumbo, S.; Morini, M.; Durando, F.; Angeletti, F.; Pirtoli, L.; Miracco, C. microRNA-17 regulates the expression of ATG7 and modulates the autophagy process, improving the sensitivity to temozolomide and low-dose ionizing radiation treatments in human glioblastoma cells. Cancer Biol. Ther. 2013, 14, 574–586. [Google Scholar] [CrossRef]
- Song, Y.; Wang, P.; Zhao, W.; Yao, Y.; Liu, X.; Ma, J.; Xue, Y.; Liu, Y. MiR-18a regulates the proliferation, migration and invasion of human glioblastoma cell by targeting neogenin. Exp. Cell Res. 2014, 324, 54–64. [Google Scholar] [CrossRef]
- Fox, J.L.; Dews, M.; Minn, A.J.; Thomas-Tikhonenko, A. Targeting of TGFβ signature and its essential component CTGF by miR-18 correlates with improved survival in glioblastoma. RNA 2013, 19, 177–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuziwara, C.S.; Kimura, E.T. Insights into Regulation of the miR-17-92 Cluster of miRNAs in Cancer. Front. Med. 2015, 2, 64. [Google Scholar] [CrossRef] [Green Version]
- Mu, P.; Han, Y.C.; Betel, D.; Yao, E.; Squatrito, M.; Ogrodowski, P.; de Stanchina, E.; D’Andrea, A.; Sander, C.; Ventura, A. Genetic dissection of the miR-17~92 cluster of microRNAs in Myc-induced B-cell lymphomas. Genes Dev. 2009, 23, 2806–2811. [Google Scholar] [CrossRef] [Green Version]
- Inomata, M.; Tagawa, H.; Guo, Y.M.; Kameoka, Y.; Takahashi, N.; Sawada, K. MicroRNA-17-92 down-regulates expression of distinct targets in different B-cell lymphoma subtypes. Blood 2009, 113, 396–402. [Google Scholar] [CrossRef]
- He, L.; Thomson, J.M.; Hemann, M.T.; Hernando-Monge, E.; Mu, D.; Goodson, S.; Powers, S.; Cordon-Cardo, C.; Lowe, S.W.; Hannon, G.J.; et al. A microRNA polycistron as a potential human oncogene. Nature 2005, 435, 828–833. [Google Scholar] [CrossRef]
- Fontana, L.; Fiori, M.E.; Albini, S.; Cifaldi, L.; Giovinazzi, S.; Forloni, M.; Boldrini, R.; Donfrancesco, A.; Federici, V.; Giacomini, P.; et al. Antagomir-17-5p abolishes the growth of therapy-resistant neuroblastoma through p21 and BIM. PLoS ONE 2008, 3, e2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, G.Q.; Wei, N.L.; Mu, L.Y.; Wang, X.Q.; Zhang, Y.N.; Zhou, W.N.; Pan, Y.W. A 4-miRNAs signature predicts survival in glioblastoma multiforme patients. Cancer Biomark. 2017, 20, 443–452. [Google Scholar] [CrossRef]
- Chen, Q.; Si, Q.; Xiao, S.; Xie, Q.; Lin, J.; Wang, C.; Chen, L.; Chen, Q.; Wang, L. Prognostic significance of serum miR-17-5p in lung cancer. Med. Oncol. 2013, 30, 353. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhang, R.; Qu, X.; Zhao, M.; Zhang, S.; Wu, H.; Jianyong, L.; Chen, L. MiR-15a, miR-16-1 and miR-17-92 cluster expression are linked to poor prognosis in multiple myeloma. Leuk. Res. 2012, 36, 1505–1509. [Google Scholar] [CrossRef] [PubMed]
- Motoyama, K.; Inoue, H.; Takatsuno, Y.; Tanaka, F.; Mimori, K.; Uetake, H.; Sugihara, K.; Mori, M. Over- and under-expressed microRNAs in human colorectal cancer. Int. J. Oncol. 2009, 34, 1069–1075. [Google Scholar]
- Schulte, J.H.; Horn, S.; Otto, T.; Samans, B.; Heukamp, L.C.; Eilers, U.C.; Krause, M.; Astrahantseff, K.; Klein-Hitpass, L.; Buettner, R.; et al. MYCN regulates oncogenic MicroRNAs in neuroblastoma. Int. J. Cancer 2008, 122, 699–704. [Google Scholar] [CrossRef]
- Xu, X.L.; Jiang, Y.H.; Feng, J.G.; Su, D.; Chen, P.C.; Mao, W.M. MicroRNA-17, microRNA-18a, and microRNA-19a are prognostic indicators in esophageal squamous cell carcinoma. Ann. Thorac. Surg. 2014, 97, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Tang, J.Q.; Tian, M.L.; Li, H.; Wang, X.; Wu, T.; Zhu, J.; Huang, S.J.; Wan, Y.L. Prognostic values of the miR-17-92 cluster and its paralogs in colon cancer. J. Surg. Oncol. 2012, 106, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Debus, J.; Abdollahi, A. For the next trick: New discoveries in radiobiology applied to glioblastoma. Am. Soc. Clin. Oncol. Educ. B 2014, e95–e99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiblak, S.; Tang, Z.; Campos, B.; Gal, Z.; Unterberg, A.; Debus, J.; Herold-Mende, C.; Abdollahi, A. Radiosensitivity of Patient-Derived Glioma Stem Cell 3-Dimensional Cultures to Photon, Proton, and Carbon Irradiation. Int. J. Radiat. Oncol. Biol. Phys. 2016, 95, 112–119. [Google Scholar] [CrossRef]
- Chiblak, S.; Tang, Z.; Lemke, D.; Knoll, M.; Dokic, I.; Warta, R.; Moustafa, M.; Mier, W.; Brons, S.; Rapp, C.; et al. Carbon irradiation overcomes glioma radioresistance by eradicating stem cells and forming an antiangiogenic and immunopermissive niche. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Debus, C.; Waltenberger, M.; Floca, R.; Afshar-Oromieh, A.; Bougatf, N.; Adeberg, S.; Heiland, S.; Bendszus, M.; Wick, W.; Rieken, S.; et al. Impact of (18)F-FET PET on Target Volume Definition and Tumor Progression of Recurrent High Grade Glioma Treated with Carbon-Ion Radiotherapy. Sci. Rep. 2018, 8, 7201. [Google Scholar] [CrossRef]
- Hartmann, C.; Meyer, J.; Balss, J.; Capper, D.; Mueller, W.; Christians, A.; Felsberg, J.; Wolter, M.; Mawrin, C.; Wick, W.; et al. Type and frequency of IDH1 and IDH2 mutations are related to astrocytic and oligodendroglial differentiation and age: A study of 1,010 diffuse gliomas. Acta Neuropathol. 2009, 118, 469–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capper, D.; Weissert, S.; Balss, J.; Habel, A.; Meyer, J.; Jäger, D.; Ackermann, U.; Tessmer, C.; Korshunov, A.; Zentgraf, H.; et al. Characterization of R132H mutation-specific IDH1 antibody binding in brain tumors. Brain Pathol. 2010, 20, 245–254. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. 2018. Available online: https://www.r-project.org/ (accessed on 1 December 2019).
- Therneau, T.; Grambsch, P. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000. [Google Scholar]
- Knoll, M. dataAnalysisMisc: Collection of Functions for Daily Tasks. R Package Version 0.99.11. Available online: http://github.com/mknoll/dataAnalysisMisc (accessed on 14 December 2019).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TCGA | HD | ||||
|---|---|---|---|---|---|
| Feature | N | % | N | % | |
| All | 482 | 100 | 109 | 100 | |
| Median OS (months) | (95% CI) | 14.0 (12.7–15.0) | 15.2 (10.7–24.1) | ||
| Median FU (months) | 55.7 | 66.7 | |||
| Age (years) | median (range) | 59 (0–89) | 65 (20–86) | ||
| Age category I | |||||
| <60 years | 249 | 51.7 | 43 | 39.4 | |
| ≥60 years | 233 | 48.3 | 66 | 60.6 | |
| Age category II | |||||
| <50 years | 120 | 24.9 | 20 | 18.3 | |
| 50–59 years | 129 | 26.8 | 23 | 21.1 | |
| 60–69 years | 134 | 27.8 | 38 | 34.9 | |
| ≥70 years | 99 | 20.5 | 28 | 25.7 | |
| Sex | |||||
| female | 186 | 38.6 | 38 | 34.9 | |
| male | 296 | 61.4 | 71 | 65.0 | |
| KPI | |||||
| <80 | 89 | 18.5 | 26 | 23.9 | |
| ≥80 | 279 | 57.9 | 43 | 39.4 | |
| N/A | 114 | 23.7 | 40 | 36.7 | |
| MGMT promotor | |||||
| hypo-methylated | 135 | 28.0 | 38 | 34.9 | |
| methylated | 50 | 10.4 | 43 | 39.4 | |
| N/A | 297 | 61.6 | 28 | 25.7 | |
| IDH1 | |||||
| mutation | 29 | 6.0 | 0 | 0 | |
| wildtype | 328 | 68.0 | 106 | 97.2 | |
| N/A | 125 | 25.9 | 3 | 2.8 | |
| TMZ | |||||
| no | 202 | 41.9 | 22 | 20.2 | |
| yes | 280 | 58.1 | 87 | 79.8 | |
| PORT | |||||
| yes | 308 | 63.9 | 95 | 87.2 | |
| no | 16 | 3.3 | 6 | 5.5 | |
| N/A | 158 | 32.8 | 8 | 7.3 |
| Median Survival (Months) | ||||
|---|---|---|---|---|
| High | Low | ΔLow–High | p-LRT | |
| hsa-miR-222 | 12.2 | 16.9 | 4.7 | 9.28 × 10−7 |
| hsa-miR-148a | 12.6 | 17.5 | 5.1 | 5.05 × 10−5 |
| hsa-miR-221 | 12.9 | 16.5 | 3.6 | 2.32 × 10−4 |
| hsa-miR-200a | 13.7 | 15.9 | 2.2 | 9.01 × 10−4 |
| hsa-miR-106a | 16.9 | 12.5 | −4.4 | 1.28 × 10−3 |
| hsa-miR-212 | 13.6 | 15.9 | 2.3 | 1.50 × 10−3 |
| hsa-miR-200b | 12.4 | 16.8 | 4.4 | 2.40 × 10−3 |
| hsa-miR-17-3p | 16.5 | 12.2 | −4.3 | 2.40 × 10−3 |
| hsa-miR-183 | 15.9 | 11.3 | −4.6 | 2.47 × 10−3 |
| hsa-miR-140 | 15.6 | 12.2 | −3.4 | 2.89 × 10−3 |
| hsa-miR-340 | 16.0 | 13.8 | −2.2 | 3.18 × 10−3 |
| hsa-miR-21 | 12.6 | 14.8 | 2.2 | 3.20 × 10−3 |
| hsa-miR-34b | 14.2 | 16.5 | 2.3 | 3.57 × 10−3 |
| hsa-miR-19b | 16.9 | 12.9 | −4.0 | 3.59 × 10−3 |
| hsa-miR-34a | 14.4 | 15.9 | 1.5 | 3.99 × 10−3 |
| hsa-miR-19a | 16.9 | 12.6 | −4.3 | 5.03 × 10−3 |
| hsa-miR-17-5p | 16.9 | 13.8 | −3.1 | 5.15 × 10−3 |
| hsa-miR-20a | 16.9 | 12.9 | −4.0 | 6.85 × 10−3 |
| hsa-miR-487a | 13.3 | 15.0 | 1.7 | 9.81 × 10−3 |
| hsa-miR-382 | 11.5 | 15.0 | 3.5 | 1.11 × 10−2 |
| Number of Samples (Method, Tissue) | miR-221 HR (95% CI) | p Value | miR-222 HR (95% CI) | p Value | |
|---|---|---|---|---|---|
| Delfino 2011 [47] | 253 (TCGA, microarray) | 0.41 (0.22–0.75) * | 0.0298 | 2.14 (1.51–3.03) | <0.0001 |
| Wang 2014 [52] | 1204 (meta-analysis) + | 1.91 (1.28–2.85) | 0.002 | 2.15 (1.51–3.06) | <0.0001 |
| Zhang 2015 [49] | 50 (qRT-PCR, plasma samples) | 2.40 (1.42–4.05) | N/A # | 2.81 (1.70–4.65) | N/A # |
| Li 2016 [30] | 54 (qRT-PCR) | 2.18 (1.02–4.65) | 0.044 | 2.13 (1.01–4.48) | 0.043 |
| Yerukala 2016 [46] | 247 (TCGA, microarray) | MED = 0.129 & | MED = 0.797 & | ||
| Zhao 2017 [51] | 106 (microarray, serum samples) | N/A | 1.71 (1.07–3.63) % | 0.038 | |
| Chen 2018 [48] | 114 (qRT-PCR) | N/A Survival benefit when miR ↓ | 0.027 § | N/A | 0.796 § |
| Swellam 2019 [50] | 20 (qRT-PCR, blood samples) | N/A Survival benefit when miR ↓ | 0.002 | N/A Survival benefit when miR ↓ | 0.001 |
| Sun 2019 [45] | 458 (TCGA, microarray) | N/A | 1.28 (1.18–1.38) | <0.001 | |
| TCGA | 482 (TCGA, microarray) | 1.31 (1.17–1.46) | <0.001 | 1.28 (1.18–1.38) | <0.001 |
| HD | 109 (HD, qRT-PCR) | 1.18 (1.04–1.34) | 0.008 | 1.21 (1.08–1.35) | 0.001 |
| Number of Samples (Method, Tissue) | Correlation of | HR | p Value | ||
|---|---|---|---|---|---|
| miR Expression | Survival | ||||
| Fox 2013 [61] | N/A (TCGA, microarray) | miR-18a↑ (+TGFβ signature↓) | ↑ | N/A | 0.035 |
| Yuan 2017 [67] | 48 (qRT-PCR) | miR signature incl. miR-17-5p↑ * | ↑ * | N/A | 0.0012 * |
| Zhao 2017 [51] | 106 (microarray, serum samples) | miR-17-5p↑ | ↓ | 1.7 (1.05–4.01) % | 0.043% |
| miR-20a↑ | ↓ | 1.69 (1.06–3.79) % | 0.04% | ||
| TCGA | 482 (microarray) | miR-17-3p↑ | ↑ | 0.75 (0.61–0.91) | 0.004 |
| miR-17-5p↑ | ↑ | 0.78 (0.68–0.9) | 0.001 | ||
| miR-18a↑ | ↑ | 0.8 (0.67–0.96) | 0.015 | ||
| miR-19a↑ | ↑ | 0.82 (0.72–0.94) | 0.003 | ||
| miR-19b↑ | ↑ | 0.85 (0.74–0.96) | 0.011 | ||
| miR-20a↑ | ↑ | 0.81 (0.71–0.92) | 0.001 | ||
| miR-92↑ | ↑ | 0.83 (0.71–0.97) | 0.021 | ||
| HD | 109 (qRT-PCR) | miR-17-5p↑ | ↓ | 1.12 (1.00–1.24) | 0.044 |
| miR-18a↑ | ↓ | 1.11 (1.02–1.22) | 0.019 | ||
| miR-19b↑ | ↓ | 1.1 (1.00–1.21) | 0.052 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schnabel, E.; Knoll, M.; Schwager, C.; Warta, R.; Mock, A.; Campos, B.; König, L.; Jungk, C.; Wick, W.; Unterberg, A.; et al. Prognostic Value of microRNA-221/2 and 17-92 Families in Primary Glioblastoma Patients Treated with Postoperative Radiotherapy. Int. J. Mol. Sci. 2021, 22, 2960. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22062960
Schnabel E, Knoll M, Schwager C, Warta R, Mock A, Campos B, König L, Jungk C, Wick W, Unterberg A, et al. Prognostic Value of microRNA-221/2 and 17-92 Families in Primary Glioblastoma Patients Treated with Postoperative Radiotherapy. International Journal of Molecular Sciences. 2021; 22(6):2960. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22062960
Chicago/Turabian StyleSchnabel, Elena, Maximilian Knoll, Christian Schwager, Rolf Warta, Andreas Mock, Benito Campos, Laila König, Christine Jungk, Wolfgang Wick, Andreas Unterberg, and et al. 2021. "Prognostic Value of microRNA-221/2 and 17-92 Families in Primary Glioblastoma Patients Treated with Postoperative Radiotherapy" International Journal of Molecular Sciences 22, no. 6: 2960. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22062960