
Extracellular Vesicles from Mesenchymal Stromal Cells for the Treatment of Inflammation-Related Conditions

 , , and
, , and
Abstract
:1. Introduction
2. The Fundamentals of Extracellular Vesicles
3. Extracellular Vesicles from Mesenchymal Stem Cells for the Treatment of Osteoarthritis
4. Extracellular Vesicles from Mesenchymal Stem Cells for the Treatment of Rheumatoid Arthritis
5. Extracellular Vesicles from Mesenchymal Stem Cells for the Treatment of Alzheimer’s Disease
6. Extracellular Vesicles from Mesenchymal Stem Cells for the Treatment of Cardio Vascular Disease
7. Extracellular Vesicles from Mesenchymal Stem Cells for the Treatment of Preeclampsia
8. Limitations and Future Perspectives of Using Extracellular Vesicles from Mesenchymal Stem Cells for the Treatment of Inflammation-Related Conditions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AβO | amyloid-β oligomer |
| AD | Alzheimer’s disease |
| ATCC | American Type Culture Collection |
| bFGF | basic fibroblast growth factor |
| CIA | collagen-induced arthritis |
| CNS | central nervous system |
| DMM | destabilisation of the medial meniscus |
| ECM | extracellular matrix |
| ESC | embryonic stem cell |
| EV | extracellular vesicle |
| FLS | fibroblast-like synoviocyte |
| GAG | glycosaminoglycan |
| GPI | glucose-6-phosphate isomerase |
| HGF | hepatocyte growth factor |
| HUVEC | human umbilical vein endothelial cell |
| I/R | ischaemia-reperfusion |
| IDE | insulin-degrading enzyme |
| IFN | interferon |
| IL | interleukin |
| IMTP | ischemic myocardium-targeting peptide |
| iNOS | inducible nitric oxide synthase |
| iPSC | induced pluripotent stem cell |
| LAD | left anterior descending |
| lncRNA | long noncoding RNA |
| LPS | lipopolysaccharide |
| LVEF | left ventricular ejection fraction |
| LVFS | left ventricular fractional shortening |
| LVSP | left ventricular systolic pressure |
| LVESD | left ventricular end systolic diameter |
| LVEDD | left ventricular end diastolic diameter |
| MI | myocardial infarction |
| MIA | monosodium iodoacetate |
| MMP | matrix metalloproteinase |
| MSC | mesenchymal stromal cell |
| MV | microvesicle |
| MWM | Morris water maze |
| NEP | neprilysin |
| NO | nitric oxide |
| NOR | Novel Object Recognition |
| NSC | neural stem cell |
| OA | osteoarthritis |
| PGE2 | prostaglandin E2 |
| PBS | phosphate buffered saline |
| PCNA | proliferating cell nuclear antigen |
| RA | rheumatoid arthritis |
| ROS | reactive oxygen species |
| s-GAG | sulfated glycosaminoglycan |
| SAP | self-assembling peptide |
| TGF | transforming growth factor |
| TNF | tumour necrosis factor |
| TRAP | tartrate-resistant acid phosphatase |
| VEGF | vascular endothelial growth factor |
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netea, M.G.; Balkwill, F.; Chonchol, M.; Cominelli, F.; Donath, M.Y.; Giamarellos-Bourboulis, E.J.; Golenbock, D.; Gresnigt, M.S.; Heneka, M.T.; Hoffman, H.M.; et al. A guiding map for inflammation. Nat. Immunol. 2017, 18, 826–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathan, C.; Ding, A. Nonresolving inflammation. Cell 2010, 140, 871–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McInnes, I.B.; Schett, G. Pathogenetic insights from the treatment of rheumatoid arthritis. Lancet 2017, 389, 2328–2337. [Google Scholar] [CrossRef] [Green Version]
- Falk, E. Pathogenesis of atherosclerosis. J. Am. Coll. Cardiol. 2006, 47 (Suppl. 8), C7–C12. [Google Scholar] [CrossRef] [Green Version]
- Zotova, E.; Nicoll, J.A.; Kalaria, R.; Holmes, C.; Boche, D. Inflammation in Alzheimer’s disease: Relevance to pathogenesis and therapy. Alzheimer Res. Ther. 2010, 2, 1–9. [Google Scholar] [CrossRef]
- Cho, W.C.; Kwan, C.K.; Yau, S.; So, P.P.; Poon, P.C.; Au, J.S. The role of inflammation in the pathogenesis of lung cancer. Expert Opin. Ther. Targets 2011, 15, 1127–1137. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and cancer: Triggers, mechanisms, and consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Nelson, W.G.; De Marzo, A.M.; DeWeese, T.L.; Isaacs, W.B. The role of inflammation in the pathogenesis of prostate cancer. J. Urol. 2004, 172, S6–S12. [Google Scholar] [CrossRef] [PubMed]
- Holgate, S.T.; Arshad, H.S.; Roberts, G.C.; Howarth, P.H.; Thurner, P.; Davies, D.E. A new look at the pathogenesis of asthma. Clin. Sci. 2010, 118, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Pickup, J.C. Inflammation and activated innate immunity in the pathogenesis of type 2 diabetes. Diabetes Care 2004, 27, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Wada, J.; Makino, H. Inflammation and the pathogenesis of diabetic nephropathy. Clin. Sci. 2013, 124, 139–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldring, M.B.; Otero, M. Inflammation in osteoarthritis. Curr. Opin. Rheumatol. 2011, 23, 471–478. [Google Scholar] [CrossRef]
- Robinson, W.H.; Lepus, C.M.; Wang, Q.; Raghu, H.; Mao, R.; Lindstrom, T.M.; Sokolove, J. Low-grade inflammation as a key mediator of the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 580. [Google Scholar] [CrossRef]
- Sokolove, J.; Lepus, C.M. Role of inflammation in the pathogenesis of osteoarthritis: Latest findings and interpretations. Ther. Adv. Musculoskelet. Dis. 2013, 5, 77–94. [Google Scholar] [CrossRef]
- Haider, L. Inflammation, iron, energy failure, and oxidative stress in the pathogenesis of multiple sclerosis. Oxidative Med. Cell. Longev. 2015, 2015, 725370. [Google Scholar] [CrossRef]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: Inflammation and the pathogenesis of depression. Trends Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Van Crombruggen, K.; Zhang, N.; Gevaert, P.; Tomassen, P.; Bachert, C. Pathogenesis of chronic rhinosinusitis: Inflammation. J. Allergy Clin. Immunol. 2011, 128, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Bringardner, B.D.; Baran, C.P.; Eubank, T.D.; Marsh, C.B. The role of inflammation in the pathogenesis of idiopathic pulmonary fibrosis. Antioxid. Redox Signal. 2008, 10, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.F.; Chen, Y.J.; Lin, Y.J.; Chen, S.A. Inflammation and the pathogenesis of atrial fibrillation. Nat. Rev. Cardiol. 2015, 12, 230–243. [Google Scholar] [CrossRef]
- Libby, P. Inflammatory mechanisms: The molecular basis of inflammation and disease. Nutr. Rev. 2007, 65, S140–S146. [Google Scholar] [CrossRef]
- Tabas, I.; Glass, C.K. Anti-inflammatory therapy in chronic disease: Challenges and opportunities. Science 2013, 339, 166–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viswanathan, S.; Shi, Y.; Galipeau, J.; Krampera, M.; Leblanc, K.; Martin, I.; Nolta, J.; Phinney, D.; Sensebe, L. Mesenchymal stem versus stromal cells: International Society for Cell & Gene Therapy (ISCT®) Mesenchymal Stromal Cell committee position statement on nomenclature. Cytotherapy 2019, 21, 1019–1024. [Google Scholar]
- Mendicino, M.; Bailey, A.M.; Wonnacott, K.; Puri, R.K.; Bauer, S.R. MSC-based product characterization for clinical trials: An FDA perspective. Cell Stem Cell 2014, 14, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Iyer, S.S.; Rojas, M. Anti-inflammatory effects of mesenchymal stem cells: Novel concept for future therapies. Expert Opin. Biol. Ther. 2008, 8, 569–581. [Google Scholar] [CrossRef]
- Murphy, M.B.; Moncivais, K.; Caplan, A.I. Mesenchymal stem cells: Environmentally responsive therapeutics for regenerative medicine. Exp. Mol. Med. 2013, 45, e54. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Lee, J.K.; Lee, H.; Carter, J.E.; Chang, J.W.; Oh, W.; Yang, Y.S.; Suh, J.-G.; Lee, B.-H.; Jin, H.K. Human umbilical cord blood-derived mesenchymal stem cells improve neuropathology and cognitive impairment in an Alzheimer’s disease mouse model through modulation of neuroinflammation. Neurobiol. Aging 2012, 33, 588–602. [Google Scholar] [CrossRef]
- Wakitani, S.; Imoto, K.; Yamamoto, T.; Saito, M.; Murata, N.; Yoneda, M. Human autologous culture expanded bone marrow mesenchymal cell transplantation for repair of cartilage defects in osteoarthritic knees. Osteoarthr. Cartil. 2002, 10, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Wang, Z.; Liu, L.; Zhang, B.; Li, B. Exosomes derived from adipose tissue, bone marrow, and umbilical cord blood for cardioprotection after myocardial infarction. J. Cell. Biochem. 2020, 121, 2089–2102. [Google Scholar] [CrossRef]
- Gupta, P.K.; Das, A.K.; Chullikana, A.; Majumdar, A.S. Mesenchymal stem cells for cartilage repair in osteoarthritis. Stem Cell Res. Ther. 2012, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Von Bahr, L.; Batsis, I.; Moll, G.; Hägg, M.; Szakos, A.; Sundberg, B.; Uzunel, M.; Ringden, O.; Le Blanc, K. Analysis of tissues following mesenchymal stromal cell therapy in humans indicates limited long-term engraftment and no ectopic tissue formation. Stem Cells 2012, 30, 1575–1578. [Google Scholar] [CrossRef]
- Barry, F. MSC therapy for osteoarthritis: An unfinished story. J. Orthop. Res. 2019, 37, 1229–1235. [Google Scholar] [CrossRef]
- Doorn, J.; Moll, G.; Le Blanc, K.; Van Blitterswijk, C.; De Boer, J. Therapeutic applications of mesenchymal stromal cells: Paracrine effects and potential improvements. Tissue Eng. Part B: Rev. 2012, 18, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.R.; Teixeira, G.Q.; Santos, S.G.; Barbosa, M.A.; Almeida-Porada, G.; Gonçalves, R.M. Mesenchymal stromal cell secretome: Influencing therapeutic potential by cellular pre-conditioning. Front. Immunol. 2018, 9, 2837. [Google Scholar] [CrossRef]
- Gnecchi, M.; Danieli, P.; Malpasso, G.; Ciuffreda, M.C. Paracrine mechanisms of mesenchymal stem cells in tissue repair. Methods Mol. Biol. 2016, 1416, 123–146. [Google Scholar] [PubMed]
- Mancuso, P.; Raman, S.; Glynn, A.; Barry, F.; Murphy, J.M. Mesenchymal stem cell therapy for osteoarthritis: The critical role of the cell secretome. Front. Bioeng. Biotechnol. 2019, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galleu, A.; Riffo-Vasquez, Y.; Trento, C.; Lomas, C.; Dolcetti, L.; Cheung, T.S.; von Bonin, M.; Barbieri, L.; Halai, K.; Ward, S.; et al. Apoptosis in mesenchymal stromal cells induces in vivo recipient-mediated immunomodulation. Sci. Transl. Med. 2017, 9, eaam7828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Witte, S.F.; Luk, F.; Sierra Parraga, J.M.; Gargesha, M.; Merino, A.; Korevaar, S.S.; Shankar, A.S.; O’Flynn, L.; Elliman, S.J.; Roy, D. Immunomodulation by therapeutic mesenchymal stromal cells (MSC) is triggered through phagocytosis of MSC by monocytic cells. Stem Cells 2018, 36, 602–615. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abels, E.R.; Breakefield, X.O. Introduction to extracellular vesicles: Biogenesis, RNA cargo selection, content, release, and uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R.F. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef] [Green Version]
- Harding, C.; Heuser, J.; Stahl, P. Endocytosis and intracellular processing of transferrin and colloidal gold-transferrin in rat reticulocytes: Demonstration of a pathway for receptor shedding. Eur. J. Cell Biol. 1984, 35, 256–263. [Google Scholar]
- Pan, B.T.; Johnstone, R.M. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983, 33, 967–978. [Google Scholar] [CrossRef]
- Rani, S.; Ryan, A.E.; Griffin, M.D.; Ritter, T. Mesenchymal stem cell-derived extracellular vesicles: Toward cell-free therapeutic applications. Mol. Ther. 2015, 23, 812–823. [Google Scholar] [CrossRef] [Green Version]
- Tsatsaronis, J.A.; Franch-Arroyo, S.; Resch, U.; Charpentier, E. Extracellular vesicle RNA: A universal mediator of microbial communication? Trends Microbiol. 2018, 26, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Van der Pol, E.; Boing, A.N.; Harrison, P.; Sturk, A.; Nieuwland, R. Classification, functions, and clinical relevance of extracellular vesicles. Pharmacol. Rev. 2012, 64, 676–705. [Google Scholar] [CrossRef] [Green Version]
- Witwer, K.W.; Buzas, E.I.; Bemis, L.T.; Bora, A.; Lasser, C.; Lotvall, J.; Nolte-’t Hoen, E.N.; Piper, M.G.; Sivaraman, S.; Skog, J.; et al. Standardization of sample collection, isolation and analysis methods in extracellular vesicle research. J. Extracell. Vesicles 2013, 2, 20360. [Google Scholar] [CrossRef]
- Lötvall, J.; Hill, A.F.; Hochberg, F.; Buzás, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar] [CrossRef]
- Grant, B.D.; Donaldson, J.G. Pathways and mechanisms of endocytic recycling. Nat. Rev. Mol. Cell Biol. 2009, 10, 597–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pols, M.S.; Klumperman, J. Trafficking and function of the tetraspanin CD63. Exp. Cell Res. 2009, 315, 1584–1592. [Google Scholar] [CrossRef] [PubMed]
- Kalra, H.; Drummen, G.P.; Mathivanan, S. Focus on extracellular vesicles: Introducing the next small big thing. Int. J. Mol. Sci. 2016, 17, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorente, A.; Skotland, T.; Sylvanne, T.; Kauhanen, D.; Rog, T.; Orlowski, A.; Vattulainen, I.; Ekroos, K.; Sandvig, K. Molecular lipidomics of exosomes released by PC-3 prostate cancer cells. Biochim. ET Biophys. Acta (BBA) Gen. Subj. 2013, 1831, 1302–1309. [Google Scholar] [CrossRef]
- Laulagnier, K.; Motta, C.; Hamdi, S.; Roy, S.; Fauvelle, F.; Pageaux, J.F.; Kobayashi, T.; Salles, J.P.; Perret, B.; Bonnerot, C.; et al. Mast cell- and dendritic cell-derived exosomes display a specific lipid composition and an unusual membrane organization. Biochem. J. 2004, 380, 161–171. [Google Scholar] [CrossRef]
- Guescini, M.; Genedani, S.; Stocchi, V.; Agnati, L.F. Astrocytes and Glioblastoma cells release exosomes carrying mtDNA. J. Neural Transm. 2010, 117, 1–4. [Google Scholar] [CrossRef]
- Waldenstrom, A.; Genneback, N.; Hellman, U.; Ronquist, G. Cardiomyocyte microvesicles contain DNA/RNA and convey biological messages to target cells. PLoS ONE 2012, 7, e34653. [Google Scholar] [CrossRef] [Green Version]
- Varderidou-Minasian, S.; Lorenowicz, M.J. Mesenchymal stromal/stem cell-derived extracellular vesicles in tissue repair: Challenges and opportunities. Theranostics 2020, 10, 5979–5997. [Google Scholar] [CrossRef]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J. Neuro Oncol. 2013, 113, 1–11. [Google Scholar] [CrossRef] [Green Version]
- El Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.-F.; Lin, S.-H.; Chen, H.-C.; Tai, C.-J.; Chang, C.-C.; Li, L.-T.; Yeh, C.-M.; Yeh, K.-T.; Chen, Y.-C.; Hsu, T.-H. CSE1L, a novel microvesicle membrane protein, mediates Ras-triggered microvesicle generation and metastasis of tumor cells. Mol. Med. 2012, 18, 1269–1280. [Google Scholar] [CrossRef]
- Tricarico, C.; Clancy, J.; D’Souza-Schorey, C. Biology and biogenesis of shed microvesicles. Small Gtpases 2017, 8, 220–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crescitelli, R.; Lasser, C.; Szabo, T.G.; Kittel, A.; Eldh, M.; Dianzani, I.; Buzas, E.I.; Lotvall, J. Distinct RNA profiles in subpopulations of extracellular vesicles: Apoptotic bodies, microvesicles and exosomes. J. Extracell. Vesicles 2013, 2, 20677. [Google Scholar] [CrossRef] [PubMed]
- Hauser, P.; Wang, S.; Didenko, V.V. Apoptotic bodies: Selective detection in extracellular vesicles. Methods Mol. Biol. 2017, 1554, 193–200. [Google Scholar]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular vesicles: Unique intercellular delivery vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montecalvo, A.; Larregina, A.T.; Shufesky, W.J.; Beer Stolz, D.; Sullivan, M.L.; Karlsson, J.M.; Baty, C.J.; Gibson, G.A.; Erdos, G.; Wang, Z. Mechanism of transfer of functional microRNAs between mouse dendritic cells via exosomes. Blood 2012, 119, 756–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegtel, D.M.; Cosmopoulos, K.; Thorley-Lawson, D.A.; van Eijndhoven, M.A.; Hopmans, E.S.; Lindenberg, J.L.; de Gruijl, T.D.; Wurdinger, T.; Middeldorp, J.M. Functional delivery of viral miRNAs via exosomes. Proc. Natl. Acad. Sci. USA 2010, 107, 6328–6333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clayton, A.; Mitchell, J.P.; Mason, M.D.; Tabi, Z. Human tumor-derived exosomes selectively impair lymphocyte responses to interleukin-2. Cancer Res. 2007, 67, 7458–7466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprague, D.L.; Elzey, B.D.; Crist, S.A.; Waldschmidt, T.J.; Jensen, R.J.; Ratliff, T.L. Platelet-mediated modulation of adaptive immunity: Unique delivery of CD154 signal by platelet-derived membrane vesicles. Blood 2008, 111, 5028–5036. [Google Scholar] [CrossRef] [Green Version]
- Chivet, M.; Hemming, F.; Fraboulet, S.; Sadoul, R. Emerging role of neuronal exosomes in the central nervous system. Front. Physiol. 2012, 3, 145. [Google Scholar] [CrossRef] [Green Version]
- Aliotta, J.M.; Pereira, M.; Li, M.; Amaral, A.; Sorokina, A.; Dooner, M.S.; Sears, E.H.; Brilliant, K.; Ramratnam, B.; Hixson, D.C. Stable cell fate changes in marrow cells induced by lung-derived microvesicles. J. Extracell. Vesicles 2012, 1, 18163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliotta, J.M.; Sanchez-Guijo, F.M.; Dooner, G.J.; Johnson, K.W.; Dooner, M.S.; Greer, K.A.; Greer, D.; Pimentel, J.; Kolankiewicz, L.M.; Puente, N. Alteration of marrow cell gene expression, protein production, and engraftment into lung by lung-derived microvesicles: A novel mechanism for phenotype modulation. Stem Cells 2007, 25, 2245–2256. [Google Scholar] [CrossRef] [Green Version]
- Rak, J.; Guha, A. Extracellular vesicles–vehicles that spread cancer genes. Bioessays 2012, 34, 489–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini-Beheshti, E.; Choi, W.; Weiswald, L.-B.; Kharmate, G.; Ghaffari, M.; Roshan-Moniri, M.; Hassona, M.D.; Chan, L.; Chin, M.Y.; Tai, I.T. Exosomes confer pro-survival signals to alter the phenotype of prostate cells in their surrounding environment. Oncotarget 2016, 7, 14639. [Google Scholar] [CrossRef] [Green Version]
- Skog, J.; Würdinger, T.; Van Rijn, S.; Meijer, D.H.; Gainche, L.; Curry, W.T.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef]
- Al-Nedawi, K.; Meehan, B.; Kerbel, R.S.; Allison, A.C.; Rak, J. Endothelial expression of autocrine VEGF upon the uptake of tumor-derived microvesicles containing oncogenic EGFR. Proc. Natl. Acad. Sci. USA 2009, 106, 3794–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidhu, S.S.; Mengistab, A.T.; Tauscher, A.N.; LaVail, J.; Basbaum, C. The microvesicle as a vehicle for EMMPRIN in tumor–stromal interactions. Oncogene 2004, 23, 956–963. [Google Scholar] [CrossRef] [Green Version]
- Parfejevs, V.; Sagini, K.; Buss, A.; Sobolevska, K.; Llorente, A.; Riekstina, U.; Abols, A. Adult stem cell-derived extracellular vesicles in cancer treatment: Opportunities and challenges. Cells 2020, 9, 1171. [Google Scholar] [CrossRef]
- Gatti, S.; Bruno, S.; Deregibus, M.C.; Sordi, A.; Cantaluppi, V.; Tetta, C.; Camussi, G. Microvesicles derived from human adult mesenchymal stem cells protect against ischaemia-reperfusion-induced acute and chronic kidney injury. Nephrol. Dial. Transplant. 2011, 26, 1474–1483. [Google Scholar] [CrossRef] [Green Version]
- Herrera, M.; Fonsato, V.; Gatti, S.; Deregibus, M.C.; Sordi, A.; Cantarella, D.; Calogero, R.; Bussolati, B.; Tetta, C.; Camussi, G. Human liver stem cell-derived microvesicles accelerate hepatic regeneration in hepatectomized rats. J. Cell. Mol. Med. 2010, 14, 1605–1618. [Google Scholar] [CrossRef] [Green Version]
- Deregibus, M.C.; Cantaluppi, V.; Calogero, R.; Lo Iacono, M.; Tetta, C.; Biancone, L.; Bruno, S.; Bussolati, B.; Camussi, G. Endothelial progenitor cell derived microvesicles activate an angiogenic program in endothelial cells by a horizontal transfer of mRNA. Blood 2007, 110, 2440–2448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratajczak, J.; Miekus, K.; Kucia, M.; Zhang, J.; Reca, R.; Dvorak, P.; Ratajczak, M. Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: Evidence for horizontal transfer of mRNA and protein delivery. Leukemia 2006, 20, 847–856. [Google Scholar] [CrossRef] [Green Version]
- De Jong, O.G.; Verhaar, M.C.; Chen, Y.; Vader, P.; Gremmels, H.; Posthuma, G.; Schiffelers, R.M.; Gucek, M.; van Balkom, B.W. Cellular stress conditions are reflected in the protein and RNA content of endothelial cell-derived exosomes. J. Extracell. Vesicles 2012, 1, 18396. [Google Scholar] [CrossRef] [PubMed]
- Lo Sicco, C.; Reverberi, D.; Balbi, C.; Ulivi, V.; Principi, E.; Pascucci, L.; Becherini, P.; Bosco, M.C.; Varesio, L.; Franzin, C. Mesenchymal stem cell-derived extracellular vesicles as mediators of anti-inflammatory effects: Endorsement of macrophage polarization. Stem Cells Transl. Med. 2017, 6, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Zhang, Z.; Hu, S.; Zhang, Z.; Chang, Z.; Huang, Z.; Liao, W.; Kang, Y. Exosomes derived from miR-92a-3p-overexpressing human mesenchymal stem cells enhance chondrogenesis and suppress cartilage degradation via targeting WNT5A. Stem Cell Res. Ther. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.C.; Yuan, T.; Zhang, Y.L.; Yin, W.J.; Guo, S.C.; Zhang, C.Q. Exosomes derived from miR-140-5p-overexpressing human synovial mesenchymal stem cells enhance cartilage tissue regeneration and prevent osteoarthritis of the knee in a rat model. Theranostics 2017, 7, 180–195. [Google Scholar] [CrossRef]
- Wu, J.; Kuang, L.; Chen, C.; Yang, J.; Zeng, W.N.; Li, T.; Chen, H.; Huang, S.; Fu, Z.; Li, J.; et al. miR-100-5p-abundant exosomes derived from infrapatellar fat pad MSCs protect articular cartilage and ameliorate gait abnormalities via inhibition of mTOR in osteoarthritis. Biomaterials 2019, 206, 87–100. [Google Scholar] [CrossRef]
- Lamichhane, T.N.; Sokic, S.; Schardt, J.S.; Raiker, R.S.; Lin, J.W.; Jay, S.M. Emerging roles for extracellular vesicles in tissue engineering and regenerative medicine. Tissue Eng. Part B Rev. 2015, 21, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Li, J.J.; Hosseini-Beheshti, E.; Grau, G.E.; Zreiqat, H.; Little, C.B. Stem cell-derived extracellular vesicles for treating joint injury and osteoarthritis. Nanomaterials 2019, 9, 261. [Google Scholar] [CrossRef] [Green Version]
- Caplan, H.; Olson, S.D.; Kumar, A.; George, M.; Prabhakara, K.S.; Wenzel, P.; Bedi, S.; Toledano-Furman, N.E.; Triolo, F.; Kamhieh-Milz, J. Mesenchymal stromal cell therapeutic delivery: Translational challenges to clinical application. Front. Immunol. 2019, 10, 1645. [Google Scholar] [CrossRef]
- Zhu, X.; Badawi, M.; Pomeroy, S.; Sutaria, D.S.; Xie, Z.; Baek, A.; Jiang, J.; Elgamal, O.A.; Mo, X.; Perle, K.L.; et al. Comprehensive toxicity and immunogenicity studies reveal minimal effects in mice following sustained dosing of extracellular vesicles derived from HEK293T cells. J. Extracell. Vesicles 2017, 6, 1324730. [Google Scholar] [CrossRef]
- Kordelas, L.; Rebmann, V.; Ludwig, A.K.; Radtke, S.; Ruesing, J.; Doeppner, T.R.; Epple, M.; Horn, P.A.; Beelen, D.W.; Giebel, B. MSC-derived exosomes: A novel tool to treat therapy-refractory graft-versus-host disease. Leukemia 2014, 28, 970–973. [Google Scholar] [CrossRef] [PubMed]
- Aghajani Nargesi, A.; Lerman, L.O.; Eirin, A. Mesenchymal stem cell-derived extracellular vesicles for kidney repair: Current status and looming challenges. Stem Cell Res. Ther. 2017, 8, 273. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cen, P.; Chen, J.; Fan, L.; Li, J.; Cao, H.; Li, L. Role of mesenchymal stem cells, their derived factors, and extracellular vesicles in liver failure. Stem Cell Res. Ther. 2017, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Casado-Díaz, A.; Quesada-Gómez, J.M.; Dorado, G. Extracellular vesicles derived from mesenchymal stem cells (MSC) in regenerative medicine: Applications in skin wound healing. Front. Bioeng. Biotechnol. 2020, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Glyn-Jones, S.; Palmer, A.; Agricola, R.; Price, A.; Vincent, T.; Weinans, H.; Carr, A. Osteoarthritis. Lancet 2015, 386, 376–387. [Google Scholar] [CrossRef]
- Varela-Eirin, M.; Loureiro, J.; Fonseca, E.; Corrochano, S.; Caeiro, J.R.; Collado, M.; Mayan, M.D. Cartilage regeneration and ageing: Targeting cellular plasticity in osteoarthritis. Ageing Res. Rev. 2018, 42, 56–71. [Google Scholar] [CrossRef]
- Malda, J.; Boere, J.; Van De Lest, C.H.; Van Weeren, P.R.; Wauben, M.H. Extracellular vesicles—new tool for joint repair and regeneration. Nat. Rev. Rheumatol. 2016, 12, 243. [Google Scholar] [CrossRef] [PubMed]
- Mora, J.C.; Przkora, R.; Cruz-Almeida, Y. Knee osteoarthritis: Pathophysiology and current treatment modalities. J. Pain Res. 2018, 11, 2189–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, R.L.; Saleh, K.J.; Wilt, T.J.; Bershadsky, B. The functional outcomes of total knee arthroplasty. J. Bone Jt. Surg. 2005, 87, 1719–1724. [Google Scholar]
- Ahmed, T.A.E.; Hincke, M.T. Strategies for articular cartilage lesion repair and functional restoration. Tissue Eng. Part B: Rev. 2010, 16, 305–329. [Google Scholar] [CrossRef]
- Akgun, I.; Unlu, M.C.; Erdal, O.A.; Ogut, T.; Erturk, M.; Ovali, E.; Kantarci, F.; Caliskan, G.; Akgun, Y. Matrix-induced autologous mesenchymal stem cell implantation versus matrix-induced autologous chondrocyte implantation in the treatment of chondral defects of the knee: A 2-year randomized study. Arch. Orthop. Trauma Surg. 2015, 135, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Vega, A.; Martín-Ferrero, M.A.; Del Canto, F.; Alberca, M.; García, V.; Munar, A.; Orozco, L.; Soler, R.; Fuertes, J.J.; Huguet, M. Treatment of knee osteoarthritis with allogeneic bone marrow mesenchymal stem cells: A randomized controlled trial. Transplantation 2015, 99, 1681–1690. [Google Scholar] [CrossRef] [PubMed]
- Galipeau, J.; Sensebe, L. Mesenchymal stromal cells: Clinical challenges and therapeutic opportunities. Cell Stem Cell 2018, 22, 824–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Arrigo, D.; Roffi, A.; Cucchiarini, M.; Moretti, M.; Candrian, C.; Filardo, G. Secretome and extracellular vesicles as new biological therapies for knee osteoarthritis: A systematic review. J. Clin. Med. 2019, 8, 1867. [Google Scholar] [CrossRef] [Green Version]
- Cosenza, S.; Ruiz, M.; Toupet, K.; Jorgensen, C.; Noel, D. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci. Rep. 2017, 7, 16214. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ding, Z.; Li, Y.; Wang, W.; Wang, J.; Yu, H.; Liu, A.; Miao, J.; Chen, S.; Wu, T.; et al. BMSCs-derived exosomes ameliorate pain via abrogation of aberrant nerve invasion in subchondral bone in lumbar facet joint osteoarthritis. J. Orthop. Res. 2020, 38, 670–679. [Google Scholar] [CrossRef]
- Vonk, L.A.; van Dooremalen, S.F.J.; Liv, N.; Klumperman, J.; Coffer, P.J.; Saris, D.B.F.; Lorenowicz, M.J. Mesenchymal stromal/stem cell-derived extracellular vesicles promote human cartilage regeneration in vitro. Theranostics 2018, 8, 906–920. [Google Scholar] [CrossRef]
- Xiang, C.; Yang, K.; Liang, Z.; Wan, Y.; Cheng, Y.; Ma, D.; Zhang, H.; Hou, W.; Fu, P. Sphingosine-1-phosphate mediates the therapeutic effects of bone marrow mesenchymal stem cell-derived microvesicles on articular cartilage defect. Transl. Res. 2018, 193, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Ragni, E.; Perucca Orfei, C.; De Luca, P.; Lugano, G.; Vigano, M.; Colombini, A.; Valli, F.; Zacchetti, D.; Bollati, V.; de Girolamo, L. Interaction with hyaluronan matrix and miRNA cargo as contributors for in vitro potential of mesenchymal stem cell-derived extracellular vesicles in a model of human osteoarthritic synoviocytes. Stem Cell Res. Ther. 2019, 10, 109. [Google Scholar] [CrossRef]
- Woo, C.H.; Kim, H.K.; Jung, G.Y.; Jung, Y.J.; Lee, K.S.; Yun, Y.E.; Han, J.; Lee, J.; Kim, W.S.; Choi, J.S.; et al. Small extracellular vesicles from human adipose-derived stem cells attenuate cartilage degeneration. J. Extracell. Vesicles 2020, 9, 1735249. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Wang, Y.; Zhao, B.; Niu, X.; Hu, B.; Li, Q.; Zhang, J.; Ding, J.; Chen, Y.; Wang, Y. Comparison of exosomes secreted by induced pluripotent stem cell-derived mesenchymal stem cells and synovial membrane-derived mesenchymal stem cells for the treatment of osteoarthritis. Stem Cell Res. Ther. 2017, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yu, D.; Liu, Z.; Zhou, F.; Dai, J.; Wu, B.; Zhou, J.; Heng, B.C.; Zou, X.H.; Ouyang, H.; et al. Exosomes from embryonic mesenchymal stem cells alleviate osteoarthritis through balancing synthesis and degradation of cartilage extracellular matrix. Stem Cell Res. Ther. 2017, 8, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Chu, W.C.; Lai, R.C.; Lim, S.K.; Hui, J.H.; Toh, W.S. Exosomes derived from human embryonic mesenchymal stem cells promote osteochondral regeneration. Osteoarthr. Cartil. 2016, 24, 2135–2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Chuah, S.J.; Lai, R.C.; Hui, J.H.P.; Lim, S.K.; Toh, W.S. MSC exosomes mediate cartilage repair by enhancing proliferation, attenuating apoptosis and modulating immune reactivity. Biomaterials 2018, 156, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yang, Y.; Li, Y.; Niu, X.; Zhao, B.; Wang, Y.; Bao, C.; Xie, Z.; Lin, Q.; Zhu, L. Integration of stem cell-derived exosomes with in situ hydrogel glue as a promising tissue patch for articular cartilage regeneration. Nanoscale 2017, 9, 4430–4438. [Google Scholar] [CrossRef]
- Liu, Y.; Zou, R.; Wang, Z.; Wen, C.; Zhang, F.; Lin, F. Exosomal KLF3-AS1 from hMSCs promoted cartilage repair and chondrocyte proliferation in osteoarthritis. Biochem. J. 2018, 475, 3629–3638. [Google Scholar] [CrossRef]
- Withrow, J.; Murphy, C.; Liu, Y.; Hunter, M.; Fulzele, S.; Hamrick, M.W. Extracellular vesicles in the pathogenesis of rheumatoid arthritis and osteoarthritis. Arthritis Res. Ther. 2016, 18, 286. [Google Scholar] [CrossRef] [Green Version]
- Catrina, A.I.; Joshua, V.; Klareskog, L.; Malmstrom, V. Mechanisms involved in triggering rheumatoid arthritis. Immunol. Rev. 2016, 269, 162–174. [Google Scholar] [CrossRef]
- Krishnamurthy, A.; Joshua, V.; Haj Hensvold, A.; Jin, T.; Sun, M.; Vivar, N.; Ytterberg, A.J.; Engstrom, M.; Fernandes-Cerqueira, C.; Amara, K.; et al. Identification of a novel chemokine-dependent molecular mechanism underlying rheumatoid arthritis-associated autoantibody-mediated bone loss. Ann. Rheum. Dis. 2016, 75, 721–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Wang, H.; Xia, Y.; Yan, F.; Lu, Y. Therapeutic potential of mesenchymal cell-derived miRNA-150-5p-expressing exosomes in rheumatoid arthritis mediated by the modulation of MMP14 and VEGF. J. Immunol. 2018, 201, 2472–2482. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, S.; Toupet, K.; Maumus, M.; Luz-Crawford, P.; Blanc-Brude, O.; Jorgensen, C.; Noel, D. Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis. Theranostics 2018, 8, 1399–1410. [Google Scholar] [CrossRef]
- Meng, H.Y.; Chen, L.Q.; Chen, L.H. The inhibition by human MSCs-derived miRNA-124a overexpression exosomes in the proliferation and migration of rheumatoid arthritis-related fibroblast-like synoviocyte cell. Bmc Musculoskelet. Disord. 2020, 21, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Zhu, L.; Iok In, I.; Chen, Y.; Jia, N.; Zhu, W. Bone marrow-derived mesenchymal stem cells-secreted exosomal microRNA-192-5p delays inflammatory response in rheumatoid arthritis. Int. Immunopharmacol. 2020, 78, 105985. [Google Scholar] [CrossRef] [PubMed]
- Headland, S.E.; Jones, H.R.; Norling, L.V.; Kim, A.; Souza, P.R.; Corsiero, E.; Gil, C.D.; Nerviani, A.; Dell’Accio, F.; Pitzalis, C.; et al. Neutrophil-derived microvesicles enter cartilage and protect the joint in inflammatory arthritis. Sci. Transl. Med. 2015, 7, 315ra190. [Google Scholar] [CrossRef]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [Green Version]
- Kinney, J.W.; Bemiller, S.M.; Murtishaw, A.S.; Leisgang, A.M.; Salazar, A.M.; Lamb, B.T. Inflammation as a central mechanism in Alzheimer’s disease. Alzheimer Dement. Transl. Res. Clin. Interv. 2018, 4, 575–590. [Google Scholar] [CrossRef]
- Chakari-Khiavi, F.; Dolati, S.; Chakari-Khiavi, A.; Abbaszadeh, H.; Aghebati-Maleki, L.; Pourlak, T.; Mehdizadeh, A.; Yousefi, M. Prospects for the application of mesenchymal stem cells in Alzheimer’s disease treatment. Life Sci. 2019, 231, 116564. [Google Scholar] [CrossRef]
- Cui, G.H.; Guo, H.D.; Li, H.; Zhai, Y.; Gong, Z.B.; Wu, J.; Liu, J.S.; Dong, Y.R.; Hou, S.X.; Liu, J.R. RVG-modified exosomes derived from mesenchymal stem cells rescue memory deficits by regulating inflammatory responses in a mouse model of Alzheimer’s disease. Immun. Ageing 2019, 16, 10. [Google Scholar] [CrossRef] [Green Version]
- De Godoy, M.A.; Saraiva, L.M.; de Carvalho, L.R.P.; Vasconcelos-Dos-Santos, A.; Beiral, H.J.V.; Ramos, A.B.; Silva, L.R.P.; Leal, R.B.; Monteiro, V.H.S.; Braga, C.V.; et al. Mesenchymal stem cells and cell-derived extracellular vesicles protect hippocampal neurons from oxidative stress and synapse damage induced by amyloid-beta oligomers. J. Biol. Chem. 2018, 293, 1957–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elia, C.A.; Tamborini, M.; Rasile, M.; Desiato, G.; Marchetti, S.; Swuec, P.; Mazzitelli, S.; Clemente, F.; Anselmo, A.; Matteoli, M.; et al. Intracerebral injection of extracellular vesicles from mesenchymal stem cells exerts reduced abeta plaque burden in early stages of a preclinical model of alzheimer’s disease. Cells 2019, 8, 1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losurdo, M.; Pedrazzoli, M.; D’Agostino, C.; Elia, C.A.; Massenzio, F.; Lonati, E.; Mauri, M.; Rizzi, L.; Molteni, L.; Bresciani, E.; et al. Intranasal delivery of mesenchymal stem cell-derived extracellular vesicles exerts immunomodulatory and neuroprotective effects in a 3xTg model of Alzheimer’s disease. Stem Cells Transl. Med. 2020, 9, 1068–1084. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, G. Bone marrow mesenchymal stem cells-derived exosomes reduce Aβ deposition and improve cognitive function recovery in mice with Alzheimer’s disease by activating sphingosine kinase/sphingosine-1-phosphate signaling pathway. Cell Biol. Int. 2020. [Google Scholar] [CrossRef]
- Ma, X.; Huang, M.; Zheng, M.; Dai, C.; Song, Q.; Zhang, Q.; Li, Q.; Gu, X.; Chen, H.; Jiang, G.; et al. ADSCs-derived extracellular vesicles alleviate neuronal damage, promote neurogenesis and rescue memory loss in mice with Alzheimer’s disease. J. Control. Release 2020, 327, 688–702. [Google Scholar] [CrossRef]
- Ding, M.; Shen, Y.; Wang, P.; Xie, Z.; Xu, S.; Zhu, Z.; Wang, Y.; Lyu, Y.; Wang, D.; Xu, L.; et al. Exosomes isolated from human umbilical cord mesenchymal stem cells alleviate neuroinflammation and reduce amyloid-beta deposition by modulating microglial activation in alzheimer’s disease. Neurochem. Res. 2018, 43, 2165–2177. [Google Scholar] [CrossRef]
- Yang, L.; Zhai, Y.; Hao, Y.; Zhu, Z.; Cheng, G. The regulatory functionality of exosomes derived from humscs in 3D culture for Alzheimer’s disease therapy. Small 2020, 16, 1906273. [Google Scholar] [CrossRef] [PubMed]
- Bodart-Santos, V.; de Carvalho, L.R.P.; de Godoy, M.A.; Batista, A.F.; Saraiva, L.M.; Lima, L.G.; Abreu, C.A.; De Felice, F.G.; Galina, A.; Mendez-Otero, R.; et al. Extracellular vesicles derived from human Wharton’s jelly mesenchymal stem cells protect hippocampal neurons from oxidative stress and synapse damage induced by amyloid-β oligomers. Stem Cell Res. Ther. 2019, 10, 332. [Google Scholar] [CrossRef]
- Reza-Zaldivar, E.E.; Hernández-Sapiéns, M.A.; Gutiérrez-Mercado, Y.K.; Sandoval-Ávila, S.; Gomez-Pinedo, U.; Márquez-Aguirre, A.L.; Vázquez-Méndez, E.; Padilla-Camberos, E.; Canales-Aguirre, A.A. Mesenchymal stem cell-derived exosomes promote neurogenesis and cognitive function recovery in a mouse model of Alzheimer’s disease. Neural Regen. Res. 2019, 14, 1626–1634. [Google Scholar]
- Li, B.; Liu, J.; Gu, G.; Han, X.; Zhang, Q.; Zhang, W. Impact of neural stem cell-derived extracellular vesicles on mitochondrial dysfunction, sirtuin 1 level, and synaptic deficits in Alzheimer’s disease. J. Neurochem. 2020, 154, 502–518. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Yang, Y.; Guo, Q.; Gao, Q.; Ding, Y.; Wang, H.; Xu, W.; Yu, B.; Wang, M.; Zhao, Y.; et al. Exosomes derived from human umbilical cord mesenchymal stem cells promote fibroblast-to-myofibroblast differentiation in inflammatory environments and benefit cardioprotective effects. Stem Cells Dev. 2019, 28, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Andrejic, O.M.; Vucic, R.M.; Pavlovic, M.; McClements, L.; Stokanovic, D.; Jevtovic-Stoimenov, T.; Nikolic, V.N. Association between Galectin-3 levels within central and peripheral venous blood, and adverse left ventricular remodelling after first acute myocardial infarction. Sci. Rep. 2019, 9, 13145. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Huang, Y.; Han, J.; Yu, L.; Li, Y.; Lu, Z.; Li, H.; Liu, Z.; Shi, C.; Duan, F.; et al. Immunomodulatory effects of mesenchymal stromal cells-derived exosome. Immunol. Res. 2016, 64, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Zhang, Y.; Lan, B.; Wang, J.; Zhang, Z.; Zhang, L.; Xiao, P.; Meng, Q.; Geng, Y.J.; Yu, X.Y.; et al. MiRNA-sequence indicates that mesenchymal stem cells and exosomes have similar mechanism to enhance cardiac repair. Biomed Res. Int. 2017, 2017, 4150705. [Google Scholar] [CrossRef]
- Liu, G.; Ma, C.; Yang, H.; Zhang, P.Y. Transforming growth factor β and its role in heart disease. Exp. Ther. Med. 2017, 13, 2123–2128. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Jin, X.; Hu, C.-F.; Li, R.; Shen, C.-X. Exosomes derived from mesenchymal stem cells rescue myocardial ischaemia/reperfusion injury by inducing cardiomyocyte autophagy via AMPK and Akt pathways. Cell. Physiol. Biochem. 2017, 43, 52–68. [Google Scholar] [CrossRef]
- Sun, X.; Shan, A.; Wei, Z.; Li, B. Intravenous mesenchymal stem cell-derived exosomes ameliorate myocardial inflammation in the dilated cardiomyopathy. Biochem. Biophys. Res. Commun. 2018, 503, 2611–2618. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, X.; Hu, J.; Chen, F.; Qiao, S.; Sun, X.; Gao, L.; Xie, J.; Xu, B. Mesenchymal stromal cell-derived exosomes attenuate myocardial ischaemia-reperfusion injury through miR-182-regulated macrophage polarization. Cardiovasc. Res. 2019, 115, 1205–1216. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Zhang, L.; Liang, C.; Liu, B.; Pan, X.; Wang, Y.; Zhang, Y.; Zhang, Y.; Xie, W.; Yan, B.; et al. Stem cell-derived exosomes prevent aging-induced cardiac dysfunction through a novel exosome/lncRNA MALAT1/NF-κB/TNF-α signaling pathway. Oxidative Med. Cell. Longev. 2019, 2019, 9739258. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Liu, Y.; Ding, X.; Li, Q.; Qiu, F.; Wang, M.; Shen, Z.; Zheng, H.; Fu, G. Bone marrow mesenchymal stem cell-secreted exosomes carrying microRNA-125b protect against myocardial ischemia reperfusion injury via targeting SIRT7. Mol. Cell. Biochem. 2020, 465, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Li, N.F.; Zhou, H.; Tang, Q. miR-133: A suppressor of cardiac remodeling? Front. Pharmacol. 2018, 9, 903. [Google Scholar] [CrossRef]
- Xu, R.; Zhang, F.; Chai, R.; Zhou, W.; Hu, M.; Liu, B.; Chen, X.; Liu, M.; Xu, Q.; Liu, N. Exosomes derived from pro-inflammatory bone marrow-derived mesenchymal stem cells reduce inflammation and myocardial injury via mediating macrophage polarization. J. Cell. Mol. Med. 2019, 23, 7617–7631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milano, G.; Biemmi, V.; Lazzarini, E.; Balbi, C.; Ciullo, A.; Bolis, S.; Ameri, P.; Di Silvestre, D.; Mauri, P.; Barile, L.; et al. Intravenous administration of cardiac progenitor cell-derived exosomes protects against doxorubicin/trastuzumab-induced cardiac toxicity. Cardiovasc. Res. 2020, 116, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Firoozi, S.; Pahlavan, S.; Ghanian, M.H.; Rabbani, S.; Barekat, M.; Nazari, A.; Pakzad, M.; Shekari, F.; Hassani, S.N.; Moslem, F.; et al. Mesenchymal stem cell-derived extracellular vesicles alone or in conjunction with a SDKP-conjugated self-assembling peptide improve a rat model of myocardial infarction. Biochem. Biophys. Res. Commun. 2020, 524, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Zhou, J.; Liang, C.; Liu, B.; Pan, X.; Zhang, Y.; Wang, Y.; Yan, B.; Xie, W.; Liu, F. Human umbilical cord mesenchymal stem cell derived exosomes encapsulated in functional peptide hydrogels promote cardiac repair. Biomater. Sci. 2019, 7, 2920–2933. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, Y.; Zhao, Z.; Meng, Q.; Yu, Y.; Sun, J.; Yang, Z.; Chen, Y.; Li, J.; Ma, T.; et al. Engineered exosomes with ischemic myocardium-targeting peptide for targeted therapy in myocardial infarction. J. Am. Heart Assoc. 2018, 7, e008737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, P.; Wang, L.; Li, Q.; Xu, J.; Xu, J.; Xiong, Y.; Chen, G.; Qian, H.; Jin, C.; Yu, Y.; et al. Combinatorial treatment of acute myocardial infarction using stem cells and their derived exosomes resulted in improved heart performance. Stem Cell Res. Ther. 2019, 10, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aneman, I.; Pienaar, D.; Suvakov, S.; Simic, T.P.; Garovic, V.D.; McClements, L. Mechanisms of key innate immune cells in early- and late-onset preeclampsia. Front. Immunol. 2020, 11, 1864. [Google Scholar] [CrossRef]
- Suvakov, S.; Richards, C.; Nikolic, V.; Simic, T.; McGrath, K.; Krasnodembskaya, A.; McClements, L. Emerging therapeutic potential of mesenchymal stem/stromal cells in preeclampsia. Curr. Hypertens. Rep. 2020, 22, 37. [Google Scholar] [CrossRef] [PubMed]
- McNally, R.; Alqudah, A.; Obradovic, D.; McClements, L. Elucidating the pathogenesis of pre-eclampsia using in vitro models of spiral uterine artery remodelling. Curr. Hypertens. Rep. 2017, 19, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalczyk, M.; Celewicz, A.; Celewicz, M.; Wozniakowska-Gondek, P.; Rzepka, R. The role of inflammation in the pathogenesis of preeclampsia. Mediat. Inflamm. 2020, 2020, 3864941. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Shi, A.; Hill, S.; Grant, C.; Kokkinos, M.I.; Murthi, P.; Georgiou, H.M.; Brennecke, S.P.; Kalionis, B. Decidual mesenchymal stem/stromal cell-derived extracellular vesicles ameliorate endothelial cell proliferation, inflammation, and oxidative stress in a cell culture model of preeclampsia. Pregnancy Hypertens. 2020, 22, 37–46. [Google Scholar] [CrossRef]
- Wang, D.; Na, Q.; Song, G.Y.; Wang, L. Human umbilical cord mesenchymal stem cell-derived exosome-mediated transfer of microRNA-133b boosts trophoblast cell proliferation, migration and invasion in preeclampsia by restricting SGK1. Cell Cycle 2020, 19, 1869–1883. [Google Scholar] [CrossRef]
- Liu, H.; Wang, F.; Zhang, Y.; Xing, Y.; Wang, Q. Exosomal microRNA-139-5p from mesenchymal stem cells accelerates trophoblast cell invasion and migration by motivation of the ERK/MMP-2 pathway via downregulation of protein tyrosine phosphatase. J. Obstet. Gynaecol. Res. 2020, 46, 2561–2572. [Google Scholar] [CrossRef]
- Xiong, Z.-H.; Wei, J.; Lu, M.-Q.; Jin, M.-Y.; Geng, H.-L. Protective effect of human umbilical cord mesenchymal stem cell exosomes on preserving the morphology and angiogenesis of placenta in rats with preeclampsia. Biomed. Pharmacother. 2018, 105, 1240–1247. [Google Scholar] [CrossRef]
- Vishnubhatla, I.; Corteling, R.; Stevanato, L.; Hicks, C.; Sinden, J. The development of stem cell-derived exosomes as a cell-free regenerative medicine. J. Circ. Biomark. 2014, 3. [Google Scholar] [CrossRef]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 2012, 1820, 940–948. [Google Scholar] [CrossRef]
- Moll, G.; Ankrum, J.A.; Kamhieh-Milz, J.; Bieback, K.; Ringdén, O.; Volk, H.-D.; Geissler, S.; Reinke, P. Intravascular mesenchymal stromal/stem cell therapy product diversification: Time for new clinical guidelines. Trends Mol. Med. 2019, 25, 149–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorgun, C.; Ceresa, D.; Lesage, R.; Villa, F.; Reverberi, D.; Balbi, C.; Santamaria, S.; Cortese, K.; Malatesta, P.; Geris, L.; et al. Dissecting the effects of preconditioning with inflammatory cytokines and hypoxia on the angiogenic potential of mesenchymal stromal cell (MSC)-derived soluble proteins and extracellular vesicles (EVs). Biomaterials 2021, 269, 120633. [Google Scholar] [CrossRef] [PubMed]
- Lener, T.; Gimona, M.; Aigner, L.; Börger, V.; Buzas, E.; Camussi, G.; Chaput, N.; Chatterjee, D.; Court, F.A.; Portillo, H.A.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar]
- Witwer, K.W.; Van Balkom, B.W.M.; Bruno, S.; Choo, A.; Dominici, M.; Gimona, M.; Hill, A.F.; De Kleijn, D.; Koh, M.; Lai, R.C.; et al. Defining mesenchymal stromal cell (MSC)-derived small extracellular vesicles for therapeutic applications. J. Extracell. Vesicles 2019, 8, 1609206. [Google Scholar] [CrossRef] [Green Version]
- Tofino-Vian, M.; Guillen, M.I.; Alcaraz, M.J. Extracellular vesicles: A new therapeutic strategy for joint conditions. Biochem. Pharmacol. 2018, 153, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Riazifar, M.; Pone, E.J.; Lotvall, J.; Zhao, W. Stem cell extracellular vesicles: Extended messages of regeneration. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 125–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardpour, S.; Ghanian, M.H.; Sadeghi-Abandansari, H.; Mardpour, S.; Nazari, A.; Shekari, F.; Baharvand, H. Hydrogel-mediated sustained systemic delivery of mesenchymal stem cell-derived extracellular vesicles improves hepatic regeneration in chronic liver failure. Acs Appl. Mater. Interfaces 2019, 11, 37421–37433. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, S.; Zhao, M.; Wang, C.; Li, L.; Yuan, Y.; Li, L.; Liao, G.; Bresette, W.; Zhang, J.; et al. Injectable extracellular vesicle-released self-assembling peptide nanofiber hydrogel as an enhanced cell-free therapy for tissue regeneration. J. Control. Release 2019, 316, 93–104. [Google Scholar] [CrossRef] [PubMed]
- van der Pol, E.; Hoekstra, A.G.; Sturk, A.; Otto, C.; van Leeuwen, T.G.; Nieuwland, R. Optical and non-optical methods for detection and characterization of microparticles and exosomes. J. Thromb. Haemost. 2010, 8, 2596–2607. [Google Scholar] [CrossRef] [PubMed]

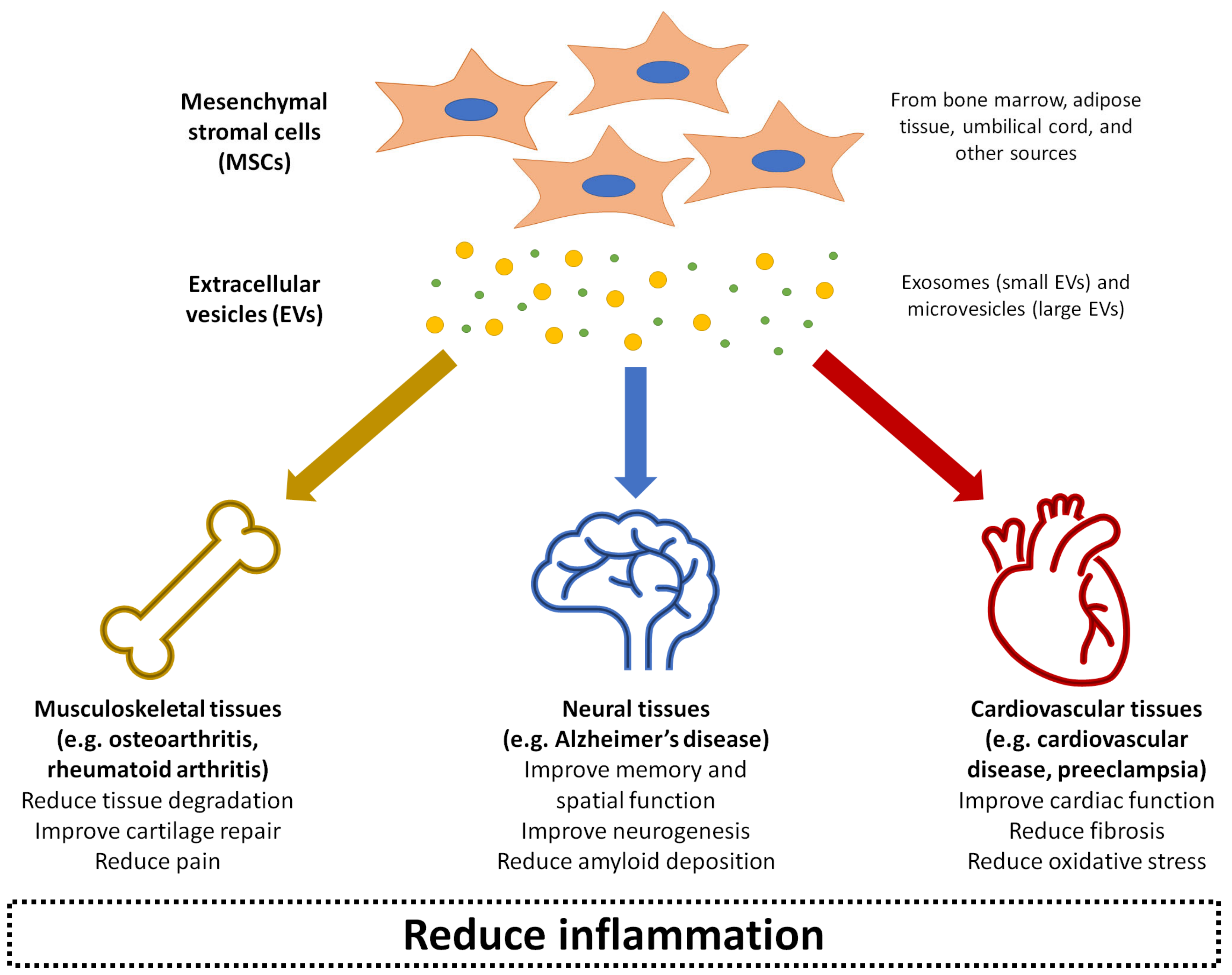

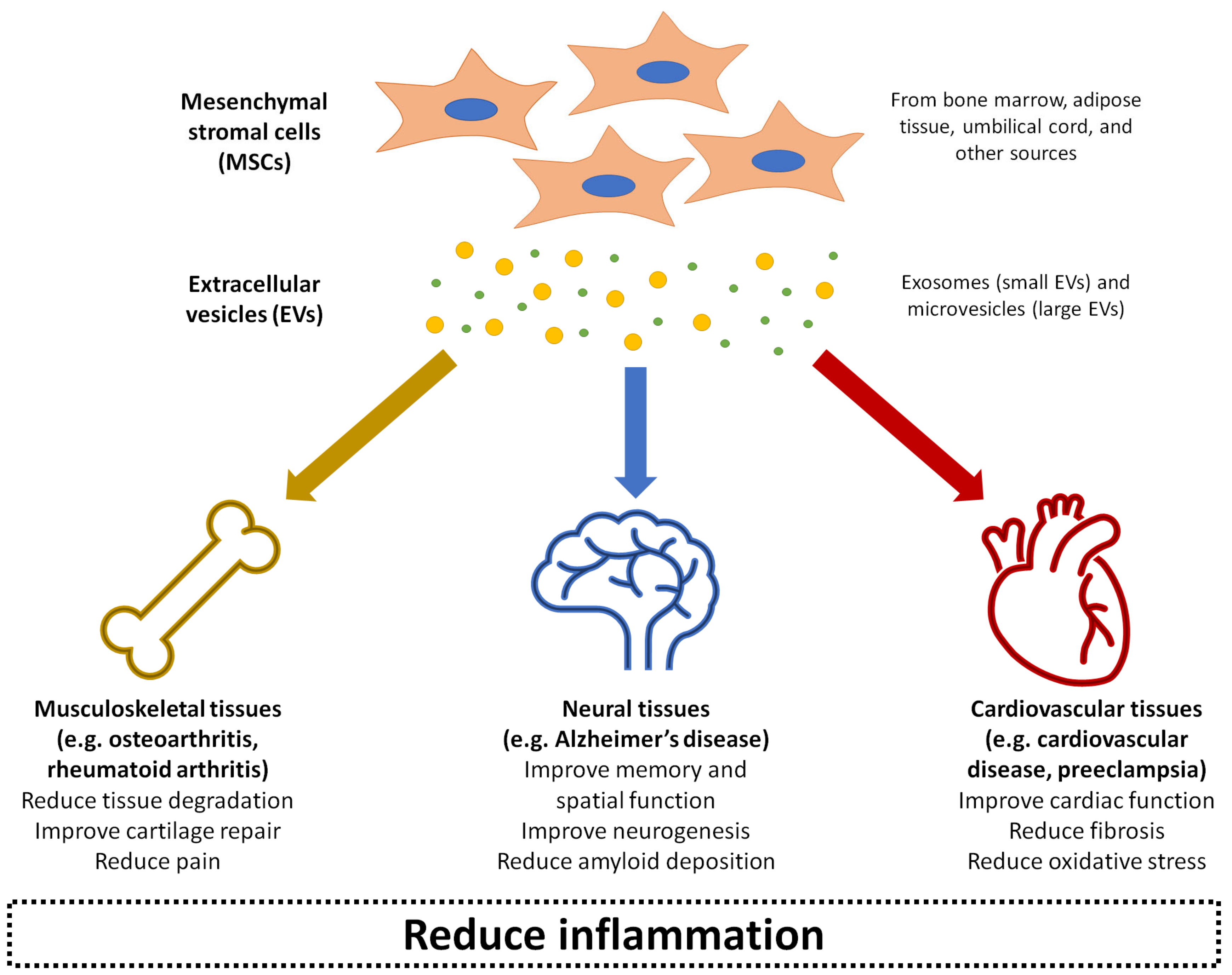
{kind=link}
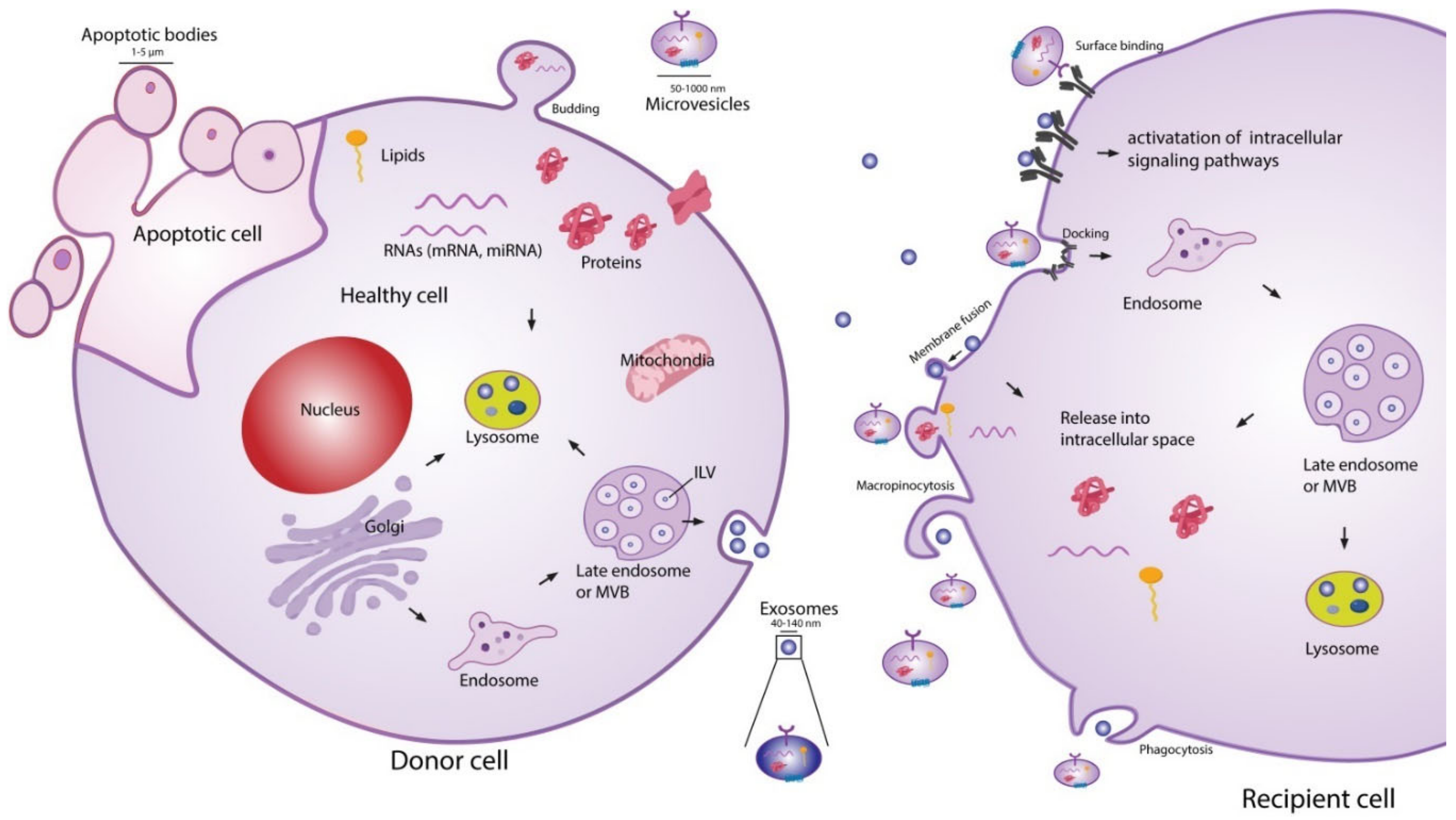
{kind=link}
| OSTEOARTHRITIS | |||||
| Study | Source of EVs | Type of EVs | Administration and Dose | Model(s) Used | Major Findings |
| Cosenza et al., 2017 [104] | Mouse bone marrow-derived MSCs | Exosomes (112 ± 6.6 nm) and MVs (223 ± 14.5 nm) | In vitro: Exosomes and MVs were applied to cells at 12.5 ng, 125 ng, or 1.25 μg In vivo: Intra-articular injection of 2.5 × 105 MSCs, 500 ng MVs or 250 ng exosomes in 5 μL saline, at seven days after OA induction | In vitro: Mouse chondrocytes treated with IL-1β (1 ng/mL) In vivo: Collagenase-induced mouse model of OA; analysis at 42 days after OA induction | In vitro: - Exosomes and MVs enhanced anabolic marker expression and decreased catabolic marker expression; at high doses, the effects of EVs on cells were similar to those treated with MSCs - Inhibited macrophage activation In vivo: - Exosomes and MVs both showed chondroprotective effects, presenting with healthy articular cartilage that had no discernible difference compared to healthy controls at 42 days - Improved subchondral bone quality and partially prevented ligament and meniscus calcification |
| Li et al., 2020 [105] | Mouse bone marrow-derived MSCs | Exosomes (120.31 ± 15.28 nm) | EVs (200 μg exosomes in 200 μL PBS) were injected weekly immediately after surgery for four weeks through the tail vein of mice | In vivo: Lumbar facet joint resection-induced mouse model of OA | - EVs had a powerful analgesic effect - Attenuated cartilage degeneration - Induced higher proteoglycan levels, and downregulation of MMP-13 and ACAN - Caused regeneration of osteochondral tissue, as well as maintenance and regeneration of subchondral bone - Blocked angiogenesis and aberrant nerve invasion |
| Liu et al., 2018 [115] | Human MSCs (from ATCC) | Exosomes (100 nm) | In vitro: Cells treated with 1, 5, or 10 µg/mL exosomes for 24 h In vivo: Intra-articular injection of exosomes from untreated MSCs and MSCs with knockdown of the lncRNA KLF3-AS1, at 21 days after OA induction | In vitro: Rat chondrocytes treated with IL-1β In vivo: Collagenase-induced rat model of OA; analysis at eight weeks | In vitro: - Partially reversed the changes induced by IL-1β treatment of cells, including reduced expression of COL2A1 and aggrecan, increased expression of MMP-13 and RUNX2, reduced migration and proliferation, and increased apoptosis In vivo: - Partially reversed cartilage degradation and expression of OA-indicative genes - Exosomes with lncRNA KLF3-AS1 knockdown had the opposite effect, suggesting that exosomal KLF3-AS1 could promote cartilage repair and chondrocyte proliferation |
| Liu et al., 2017 [114] | Human iPSC-derived MSCs | Exosomes (50–150 nm) | Exosomes were integrated into a hydrogel in a 20 μL volume at 1 × 1011 particles/mL, or given by intra-articular injection directly in the same volume and concentration | In vitro: Human chondrocytes or bone marrow-derived MSCs In vivo: Full-thickness osteochondral defects (4 mm diameter, 3 mm depth) in rabbits | In vitro: - Increased the migration of chondrocytes and MSCs - When cells were hydrogel encapsulated, exposure to exosomes improved their viability In vivo: - Exosomes suspended in the hydrogel formed in situ and integrated well with surrounding cartilage - 7 days after implantation, treated joints showed superior cell infiltration including chondrocytes, inflammatory cells, fibroblasts, and blood cells - 12 weeks after implantation, treated joints showed complete defect filling with smooth hyaline-like cartilage (high levels of organised collagen type II and low levels of collagen type I), and complete integration with native tissue |
| Lo Sicco et al., 2017 [82] | Human adipose-derived MSCs cultured in normoxic (20% O2) or hypoxic (1% O2) atmosphere | Mostly exosomes but also contained MVs (4–250 nm) | Intramuscular injection of 1 μg EVs in 20 μL PBS, injected 2 h after muscular injury and repeated after four days in muscular injury model | In vitro: Mouse bone marrow-derived macrophages In vivo: Mouse cardiotoxin-induced muscular injury model (analysis at 1, 2, and 7 days after injury) | In vitro: - MSCs cultured in normoxic or hypoxic conditions produced EVs with the same size and morphology - Both EV groups increased the expression of angiogenic molecules and induced epithelial tube formation, with hypoxic EVs being more angiogenic - Both EV groups were internalised by macrophages and increased macrophage proliferation; both demonstrated anti-inflammatory properties and ability to switch macrophages from M1 to M2 phenotype; hypoxic EVs had greater effects - Hypoxic EVs showed downregulation of 48 miRNAs and upregulation of 20 miRNAs; 4 of the upregulated miRNAs were associated with inflammatory processes (miR-223, miR-146b), proliferation and differentiation (miR-126, miR-199a) In vivo: - Both EV groups showed anti-inflammatory properties, with increase in M2 anti-inflammatory markers and decrease in M1 pro-inflammatory markers - Both EV groups showed accelerated regeneration of muscle tissue, although hypoxic EVs had a greater effect |
| Mao et al., 2018 [83] | Human bone marrow-derived MSCs | Exosomes (50–150 nm) | Intra-articular injection of exosomes from untreated MSCs and MSCs overexpressing miR-92a-3p; 15 μL at 500 μg/mL injected at 7, 14, and 21 days after OA induction | In vitro: Human chondrocytes treated with IL-1β and human MSCs undergoing chondrogenesis In vivo: Collagenase-induced rat model of OA; analysis at 28 days | In vitro: - Enhanced chondrocyte proliferation and migration - Upregulated COL2A1, SOX9 and aggrecan, and downregulated MMP-13, WNT5A, COL10A1 and RUNX2 in MSCs undergoing chondrogenesis In vivo: - Exosomes overexpressing miR-92a-3p reversed the OA-related changes (reduced COL2A1 and aggrecan in the cartilage matrix) to levels slightly below normal cartilage tissue, suggesting that they can reverse cartilage degradation in OA |
| Ragni et al., 2019 [108] | Human adipose-derived MSCs | Not specified; both small and large EVs | EVs were added to cells at a ratio of 100,000:1 (EVs:cell) for 10 days, with EVs freshly supplemented every two days | In vitro: Inflammatory OA cell model, comprising human FLSs isolated from OA patients and stimulated with IL-1β at physiological levels (25 pg/mL) | - Downregulated HAS3, MMP1, MMP13, CCL2 and CCL5 gene expression, and significantly (50%) downregulated CXCL8 expression from 2 to 10 days - EVs showed clear anti-inflammatory properties, possibly mediated by direct interaction with hyaluronan matrix and miRNA release |
| Tao et al., 2017 [84] | Human synovial membrane-derived MSCs | Exosomes (30–150 nm) | Intra-articular injection of exosomes from untreated MSCs and MSCs overexpressing miR-140-5p; 100 μL at 1 × 1011 particles/mL injected weekly from the fifth to eighth week after OA induction | In vitro: Human articular chondrocytes In vivo: Rat OA model induced through medial collateral ligament and medial meniscus transection; analysis at 12 weeks | In vitro: - Exosomes with normal miR-140-5p content increased chondrocyte proliferation and migration, but decreased ECM secretion levels - Exosomes with upregulated miR-140-5p increased chondrocyte proliferation and migration while maintaining ECM secretion levels In vivo: - Exosome with normal miR-140-5p content were mildly chondroprotective - Exosomes with upregulated miR-140-5p had much stronger chondroprotective abilities and the treated joint showed a very low level of osteochondral degeneration |
| Vonk et al., 2018 [106] | Human bone marrow-derived MSCs | Exosomes (40–150 nm) | EVs were added to cells over a period of 48 h (inflammatory model) or 28 days with repeated treatment every five days (cartilage regeneration model) | In vitro: Inflammatory model–human OA chondrocytes stimulated with TNF-α; cartilage regeneration model–human OA chondrocytes cultured in fibrin matrices | - In inflammatory model, EVs suppressed TNF-α and COX2 expression, as well as pro-inflammatory interleukins (IL-1α, IL-1β, IL-6, IL-8, IL-17) - In cartilage regeneration model, EVs improved proteoglycan content and upregulated the production of collagen type II |
| Wang et al., 2017 [111] | Human ESC-derived MSCs | Exosomes (38–169 nm) | Intra-articular injection of 1 × 106 particles in 5 μL PBS in the bilateral joints, every 3 days for 28 days after OA induction | In vitro: Mouse articular chondrocytes treated with IL-1β In vivo: DMM mouse model of OA | In vitro: - Reversed the increased synthesis of collagen type II and ADAMTS5 expression in IL-1β-treated chondrocytes In vivo: - Significant chondroprotection and attenuation of OA progression - Maintained collagen type II content and lower levels of ADAMTS5 |
| Woo et al., 2020 [109] | Human adipose- derived stem cells | Not specified; mostly small EVs (around 100 nm) | In vitro: Cells treated with 1 × 108 or 2 × 108 particles/mL In vivo: Intra-articular injection of 1 × 108 particles in a 30 μL volume per joint | In vitro: Human OA chondrocytes treated with IL-1β In vivo: MIA rat model of OA and DMM mouse model of OA | In vitro: - Reduced catabolic gene expression - Reduced MMP-1, MMP-3, MMP-13, and ADAMTS5 expression - Increased collagen type II synthesis - Reduced expression of pro-inflammatory and cartilage degradation genes In vivo: - Reduced cartilage erosion, OA progression, and proteoglycan degradation - Reduced synovial inflammation and inhibited M1 macrophage infiltration into the synovium - Reduced MMP-13 expression - Maintained adipocyte content of infrapatellar fat pad - Did not demonstrate subintimal fibrosis and neovascularisation |
| Wu et al., 2019 [85] | MSCs from human intrapatellar fat pad | Exosomes (average 121.9 nm) | In vitro: Cells treated with different concentrations (1, 5, or 10 × 108 particles/mL) in the presence or absence of IL-1β (10 ng/mL) In vivo: Intra-articular injection of 1 × 1010 particles/mL in 10μL twice a week for four or six weeks | In vitro: Human OA chondrocytes In vivo: DMM mouse model of OA | In vitro: - Increased chondrocyte viability and cell migration - Partially reversed IL-1β-induced apoptosis - Reversed IL-1β-induced changes to ADAMTS5, MMP-13 and collagen type II expression In vivo: - Attenuated cartilage degradation and promoted cartilage maintenance - Reversed DMM-induced changes to MMP-14, ADAMTS5, and collagen type II expression |
| Xiang et al., 2018 [107] | Human bone marrow-derived MSCs | MVs (average 200 nm) | In vitro: Cells were treated with 5–20 μL MVs isolated from 1 × 106 MSCs, for 12 h In vivo: Intra-articular injection of 30 μL MVs at three days after defect creation | In vitro: Human chondrocytes treated with IL-1β (10 ng/mL) In vivo: Osteochondral defects (4 mm diameter, 5 mm depth) in the medial femoral condyle of rabbits | In vitro: - Cells showed EV uptake, which was CD44 dependent - Increased proliferation potential and decreased apoptosis rate In vivo: - At 20, 40, and 60 days, MV-treated group caused gradual defect filling with hyaline-like tissue rich in collagen type II, reaching the heigh of native cartilage; PBS-treated group showed no significant regeneration, with adipose and fibrotic cells progressively filling the defect |
| Zhang et al., 2018 [113] | MSCs derived from a human embryonic cell line | Exosomes (average 100 nm) | Intra-articular injection of 100 μg exosomes in 100 μL PBS once per week for 12 weeks after OA induction | In vitro: Rat chondrocytes In vivo: Osteochondral defects (1.5 mm diameter, 1 mm depth) in the knee joint of rats; analysis at 2, 6, and 12 weeks | In vitro: - Cells showed increased migration and proliferation potential proportional to the dose of exosomes - Increased ECM synthesis and decreased apoptosis In vivo: - Promoted hyaline-like cartilage regeneration - Increased collagen type II and s-GAG content - Increased cell proliferation and apoptosis attenuation - Caused increase in cells presenting PCNA and a more rapid decrease in CP3 apoptotic cells - Increased abundance of M2 macrophages in the synovium and cartilage, and decreased M1 macrophages and inflammatory cytokines |
| Zhang et al., 2016 [112] | MSCs derived from a human embryonic cell line | Exosomes (average 100 nm) | Intra-articular injection of 100 μg exosomes in 100 μL PBS once per week for 12 weeks after OA induction | In vivo: Osteochondral defects (1.5 mm diameter, 1 mm depth) in the knee joint of rats; analysis at 6 and 12 weeks | - Attenuated cartilage degeneration - Caused regeneration of hyaline cartilage, and increased GAG and collagen type II content - Caused complete regeneration of subchondral bone |
| Zhu et al., 2017 [110] | Human synovial membrane-derived MSCs, and human iPSC-derived MSCs | Exosomes (50–150 nm) | Intra-articular injection of 1 × 1010 particles/mL in 8 μL PBS at 7, 14, and 21 days after OA induction | In vitro: Human articular chondrocytes In vivo: Collagenase-induced mouse model of OA; analysis at 28 days | In vitro: - Both exosome groups improved chondrocyte migration and proliferation, with exosomes from iPSC-derived MSC showing more dramatic improvements In vivo: - Both exosome groups showed significant improvements in reducing OA pathology compared to controls - Exosomes from iPSC-derived MSCs showed a greater regenerative potential, presenting with smooth hyaline-like cartilage, normal collagen type II localisation and healthy proteoglycan content |
| RHEUMATOID ARTHRITIS | |||||
| Study | Source of EVs | Type of EVs | Administration and Dose | Model(s) Used | Major Findings |
| Chen et al., 2018 [119] | Mouse bone marrow-derived MSCs | Exosomes (average 100 nm) | Intradermal injection of 50 μg exosomes in 100 μL PBS twice per week; exosomes were from MSCs overexpressing miR-150-5p or the control miR-67 | In vitro: FLSs isolated from RA patients, and HUVEC and FLS co-culture In vivo: CIA mouse model | In vitro: - Exosomes enriched with miR-150-5p substantially inhibited tube formation in HUVEC-FLS co-culture, as well as migration and invasion of RA FLSs, indicating that they can suppress angiogenesis - miR-150-5p from enriched exosomes suppressed the expression of MMP-14 and VEGF, but miR-67 did not have the same effects In vivo: - Exosomes enriched with miR-150-5p induced downregulation of MMP-14 and VEGF in tissue, reduction in the thickness of the hind paw, and lower clinical scores of arthritis compared to groups treated with miR-67 control exosomes or PBS - Substantially improved arthritis severity and successfully inhibited angiogenesis |
| Cosenza et al., 2018 [120] | Mouse bone marrow-derived MSCs | EVs separated into exosomes (average 120 nm, expressing CD81 and CD 9) and MVs (150–600 nm, expressing Sca-1, CD44 and CD29) | Intravenous injection of 250 ng exosomes or 600 ng MVs in CIA mouse model at 18 and 24 days after arthritis induction | In vitro: Mouse spleen T- and B-lymphocytes In vivo: Mouse delayed T hypersensitivity (DTH) model (injection into footpad at five days after immunisation; analysis at 24 h after injection), and CIA mouse model (analysis at 30 days) | In vitro: - EVs from MSCs primed with IFN-γ showed a dose-dependent effect on T-lymphocyte suppression, but their immunomodulatory effects were lost after freeze-thawing - Primed and un-primed exosomes and MVs suppressed ConA-activated splenocytes to a similar extent - MSCs exerted the strongest suppression of CD8+ IFN-γ+ cells, followed by similar levels by exosomes and MVs - MSCs, exosomes and MVS all increased the percentage of CD4+ IL-10+ Tr1 cells - Exosomes and MVs both increased the percentage of CD4+ CD25+ Treg cells, while MSCs had no effect - Isolated CD4+ and CD8+ cells treated with exosomes or MVs did not show reduced proliferation In vivo: - In DTH model, both exosomes and MVs showed anti-inflammatory effects that were dose-dependent - In CIA model, exosomes and MVs showed significant protection from developing arthritis; treatment with exosomes showed 5% rate of developing arthritis, and this 5% had very low clinical arthritis scores; treatment with MVs did not exert significant protection from arthritis symptoms, but reduced the incidence to 20% with low clinical scores - Exosomes and MVs both showed maintenance of subchondral bone, with exosomes being more effective than MVs |
| Headland et al., 2015 [123] | Human RA synovial fluid, and human neutrophils (stimulated or not stimulated with TNF-α) | MVs (0.05–1 μm; may contain both exosomes and MVs) | Intra-articular injection of 3 × 104 particles in 5 μL PBS in k/BxN-induced model at 3 days after arthritis induction, or in GPI-induced model at 21 days after arthritis induction | In vitro: Human chondrocyte micromass, and ex vivo rat cartilage explant In vivo: Inflammatory arthritis rat models induced by k/BxN (analysis at five days after induction) or GPI (analysis at 25 days after induction) | In vitro: - Protected against cartilage degradation by reducing IL-8 and PGE2 release, ECM degradation, and chondrocyte apoptosis - EVs from neutrophils showed the ability to migrate through the ECM in rat cartilage explants, and migration was increased in explants treated with IL-1β; EVs needed to remain intact to migrate and exert chondroprotective effects - Although neutrophil-derived EVs had chondroprotective effects, direct contact between neutrophils and chondrocytes induced apoptosis In vivo: - Neutrophils injected into damaged joints migrated towards zones of inflammation where they released EVs, which showed the ability to penetrate cartilage ECM - Protective effects of neutrophil-derived EVs were thought to be related to AnxA1 and FPR2/ALX interactions, which increased the production of TGF-β in chondrocytes |
| Meng et al., 2020 [121] | Human bone marrow-derived MSCs | Exosomes (approximately 100 nm) | Cells were treated with 20 μg/mL exosomes from MSCs overexpressing miR-124a | In vitro: Human RA FLS cell line | - Exosomes enriched with miR-124a showed the ability to enter cells, providing significant numbers of exosomes and high levels of miR-124a in cells - Suppressed RA FLS proliferation, inhibited wound closure healing rate at 24 h, and inhibited migration and invasion - Exosomes enriched with miR-124a arrested the cell cycle in the G0/G1 phase, compared to control exosomes at the S phase - Both miR-124a-enriched and control exosomes promoted RA FLS apoptosis |
| Zheng et al., 2020 [122] | Rat bone marrow-derived MSCs | Exosomes (101 ± 14.45 nm) | Injection of 50 μg exosomes in 100 μL PBS twice per week, starting at three weeks after the second arthritis induction procedure; exosomes were from MSCs overexpressing miR-192-5p | In vivo: CIA rat model, with two inductions 21 days apart | - Exosomes enriched with miR-192-5p showed the ability to migrate from the bloodstream to synovial tissues, where they significantly increased miR-192-5p expression and reduced RAC2 expression - Reduced TRAP activity (usually elevated in CIA model) - Significantly attenuated the elevated levels of PGE2, IL-1β, and TNF-α levels in synovial tissues, and levels of NO and iNOS in serum (usually elevated in CIA model); exosomes enriched with miR-192-5p had greater effects than control exosomes |
| ALZHEIMER’S DISEASE | |||||
| Study | Source of EVs | Type of EVs | Administration and Dose | Model(s) Used | Major Findings |
| Bodart-Santos et al., 2019 [135] | Human Wharton’s jelly mesenchymal stem cells | Not specified, contained both exosomes and MVs (mostly 50–300 nm) | Cells were cultured with 6 × 108 EV particles | In vitro: Hippocampal cells were isolated from hippocampi from 18-day old rat embryos, and conditioned or not with AβOs for 2 h | - Control cells showed low uptake of EVs, while AβO-treated cells showed much higher levels of EV uptake - EV uptake was primarily performed by astrocytes rather than neuronal cells - Once catalase in EVs was inactivated, the EVs no longer prevented the formation of ROS in AβO-treated cells - EV treatment of AβO-treated cells for 22 h prevented synapse damage |
| Cui et al., 2019 [127] | Mouse bone marrow-derived MSCs | Exosomes, tagged with rabies viral glycoprotein (RVG) which targets the CNS | Intravenous injection of tagged and untagged exosomes at 5 × 1011 particles in 100 μL PBS, once per month for four months | In vivo: APP/PS1 transgenic mouse (prone to early onset of AD) | - Conjugated RVG-exosomes travelled from the bloodstream to the cortex and hippocampus at much greater numbers than unconjugated exosomes - Conjugated exosomes greatly reduced the levels of cortex and hippocampus plaque deposition, soluble Aβ40 and Aβ42, insoluble Aβ40 and Aβ42, and GFAP expression (astrocyte marker) in the brain; unconjugated exosomes showed smaller effects - Conjugated exosomes caused improved spatial recognition and memory retention as shown through MWM test - Conjugated exosomes substantially downregulated the pro-inflammatory markers IL-1α, IL-1β and IL-6, and upregulated the anti-inflammatory markers IL-4, IL-10 and IL-12 |
| de Godoy, 2018 [128] | Rat bone marrow-derived MSCs | Not specified, contained both exosomes and MVs (mostly 50–300 nm) | Cells were cultured with 8 × 107 EV particles (corresponds to ~5000 MSCs), dose tripled in some experiments | In vitro: Hippocampal neuronal cells treated with AβOs | - Prevented AβO-induced synapse damage in neurons - EVs that had catalase (an antioxidant) removed lost the ability to protect against oxidative stress - Functional ability of EVs was maintained after cryopreservation |
| Ding et al., 2018 [133] | Human umbilical cord-derived MSCs | Exosomes (30–150 nm) | In vitro: Cells were cultured with exosomes at 30 μg/mL In vivo: Intravenous injection of 30 μg exosomes in 100 μL PBS every two weeks for eight weeks | In vitro: Mouse BV2 microglia cell line In vivo: APP/PS1 transgenic mouse | In vitro: - Cells showed alternative activation into anti-inflammatory M2 microglia, with decreased levels of pro-inflammatory cytokines (IL1β, TNF-α) and increased levels of anti-inflammatory cytokines (IL-10, TGF-β) In vivo: - Increased memory as shown through MWM test - Decreased numbers of Aβ plaques in the hippocampus and cortex, and soluble Aβ40 and Aβ42 in the brain - Greatly increased the levels of NEP and IDE (Aβ-degrading enzymes) - Decreased levels of Iba-1 positive (pro-inflammatory M1) microglia, and increased expression of markers for anti-inflammatory M2 microglia |
| Elia et al., 2019 [129] | Mouse bone marrow-derived MSCs | Not specified, contained both exosomes and MVs (mostly 50–300 nm) | Intracerebral injection of 22.4 μg EVs (1 × 109 particles) in 4 μL PBS | In vivo: APP/PS1 transgenic mouse, two age groups (three and five months; earliest signs of cognitive impairment appear at six months); analysis at 25 days after injection | - Reduced Aβ plaque area and density in the hippocampus and cortex - Reduced plaque solidarity in the neocortex (site of injection) - Lower numbers of plaques surrounded by dystrophic neurites - Introduces the possibility for intervention before clinical manifestation of AD |
| Li et al., 2020 [137] | Mouse hippocampus NSCs | Not specified (50–190 nm) | Injection of 200 μg EVs in 10 μL PBS bilaterally into the lateral ventricles, twice a week for four weeks | In vivo: APP/PS1 transgenic mouse | - Increased memory as shown through MWM test - Upregulated the expression of mitochondrial function-related factors and synaptic proteins - Significantly reduced the ratio of damaged to total synapses - Significantly reduced the levels of pro-inflammatory IL-1β, IL-4, IL-10, p65, and TNF-α, as well as Iba-1 expression compared to vehicle control - No significant differences in the levels of soluble and insoluble Aβ40 and Aβ42 between EV-treated and vehicle control groups |
| Losurdo et al., 2020 [130] | Human bone marrow-derived MSCs | Not specified, contained both exosomes and MVs (average 200 nm) | In vitro: Cells were cultured with EVs (4.5 μg/mL) from MSCs pre-conditioned with TNF-α and IFN-γ In vivo: Intranasal spurts of 5 μL EV solution (300 μg/mL) totalling 100 μL, given twice at 18 h apart | In vitro: Microglia cultures consisting of hippocampal and cortical astrocytes, treated with TNF-α and IFN-γ In vivo: Female triple-transgenic AD mouse expressing three mutant human transgenes (PS1M146V, APPSwe, and tauP301L) | In vitro: - Reduced expression of pro-inflammatory markers IL-6 and IL-1β, and increased expression of the anti-inflammatory marker IL-10 - Completely or partly attenuated the negative effects exerted by pro-inflammatory cytokine treatment In vivo: - Reduced Iba-1+ cell density, together with reduction in microglial cell body size - Reduced CD68 expression associated with the activated microglia phenotype |
| Ma et al., 2020 [132] | Human adipose MSCs | EVs (130 ± 28 nm) | In vitro: Cells were cultured with EVs for 24 h In vivo: Intranasal administration of 10 μL EVs at the protein dose of 1 mg/kg every two days for two weeks | In vitro: Primary neurons from embryonic mice, treated with Aβ1-42 oligomers or glutamate In vivo: APP/PS1 transgenic mouse | In vitro: - RNA sequencing showed neuroprotective effects of EVs, with some upregulated genes important for synaptic function, and downregulated genes related to cell death - Significantly reversed neuronal toxicity induced by Aβ1-42 oligomers or glutamate; increased cell viability In vivo: - Reduced neurologic damage in whole brain areas, and remarkably increased neurogenesis - Slightly reduced Aβ deposition and microglia activation - Rescued memory deficits as shown through NOR and Y-maze tests |
| Reza-Zaldivar et al., 2019 [136] | MSCs (from ATCC) | Exosomes (size unspecified) | Intra-peritoneal injection of 10 μg exosomes in 2 μL PBS; analysis at 14 and 28 days after injection | In vivo: C57BL/6 mouse treated with Aβ aggregates, and AD allowed to develop for 14 days before intervention | - Improved spatial learning and memory as shown through MWM test - Increased scores in NOR test - Stimulated expression of neurogenesis markers in the subventricular zone - Increased numbers of immunoreactive cells compared to PBS control, but similar numbers compared to MSC-treated group |
| Wang et al., 2020 [131] | Mouse bone marrow-derived MSCs | Exosomes (approximately 110 nm) | Injection of exosomes (50 μg in 80 μL saline) through the cauda vein, every two weeks for 16 weeks | In vivo: APP/PS1 transgenic mouse | - Significantly improved spatial learning and memory ability as shown through MWM test - Reduced amyloid levels in the cortex and hippocampus, and enhanced the expression of NeuN - Reduced Aβ1-40, Aβ1-42, BACE1, and PS1 expression, and promoted NEP expression in the brain - Effects were mediated by activating the SphK/S1P signalling pathway |
| Yang et al., 2020 [134] | Human umbilical cord MSCs | Exosomes (50–150 nm) | In vitro: Cells were cultured with 2 μg exosomes for 24 h; exosomes were isolated from MSCs cultured on 2D graphene film or 3D graphene scaffold In vivo: Exosomes were delivered by infusion into the right hippocampus, at 0.25 μL/h (2 mg protein/mL) for 14 days | In vitro: AD pathology cell model, comprising SH-SY5Y cells transfected with amyloid precursor protein (APP) gene, leading to increased production of Aβ peptides In vivo: APP/PS1 transgenic mouse | In vitro: - 3D-exosomes had greater effect in upregulating α-secretase and downregulating β-secretase to reduce levels of secreted and intracellular Aβ In vivo: - 3D exosomes were more effective at improving spatial learning and memory function, as shown through MWM test - Exosomes were mainly concentrated at the site of delivery but also distributed throughout the brain parenchyma; 3D exosomes were more effective at decreasing Aβ deposition by eliminating production and facilitating clearance of Aβ - Exosomes reduced neuroinflammation by attenuating microglial activation, and markedly reduced oxidative stress; 3D exosomes produced more pronounced effects |
| CARDIOVASCULAR DISEASE | |||||
| Study | Source of EVs | Type of EVs | Administration and Dose | Model(s) Used | Major Findings |
| Chen et al., 2020 [147] | Rat bone marrow MSCs | Exosomes (60–100 nm), enriched with miR-125b or control miR-67 | Exosomes (50 μg) were injected into the ligation zone contiguous to the left anterior free wall after left ventricle exposure | In vivo: Rat I/R model, in which the LAD was ligated for 30 min followed by reperfusion for 2 h | - Increased cell viability and inhibited inflammation, oxidative stress and apoptosis - Reduced infarct size and improved cardiac function, with increased LVEF, LVFS and LVSP - Upregulated miR-125b and anti-apoptotic Bcl-2, and downregulated pro-apoptotic factors Bax and caspase-3 - Decreased the levels of inflammatory factors IL-1β, IL-6 and TNF-α - Downregulated SIRT7 gene and protein expression |
| Firoozi et al., 2020 [151] | Human bone marrow MSCs | EVs (average 100 nm) | EVs (80 μg) were injected in a 100 μL volume, directly or encapsulated in an SAP hydrogel, into four sites of the infarct border zone after ligation | In vitro: Neonatal mouse cardiomyocytes treated with H2O2 In vivo: LAD coronary artery ligation rat model of MI | In vitro: - Protected cardiomyocytes from damage due to H2O2-induced oxidative stress In vivo: - Improved LVEF and LVFS, promoted cardiac morphological status and preserved function - Reduced fibrosis area, apoptosis and inflammation; reduced expression of pro-inflammatory CD68+ cells - Promoted angiogenesis in infarcted myocardium, with increased numbers of α-SMA+ structures - Both encapsulated and free EVs improved cardiac function, likely by reducing macrophage infiltration and increasing angiogenesis |
| Han et al., 2019 [152] | Human umbilical cord MSCs | Exosomes (size unspecified) | Exosomes (20 μg) were injected in a 20 μL volume, directly or encapsulated in a peptide hydrogel, into two different sites next to the infarcted border region after ligation | In vitro: Rat H9C2 cardiomyoblasts treated with H2O2 In vivo: LAD coronary artery ligation rat model of MI | In vitro: - Exosomes protected cell damage from H2O2-induced oxidative stress and improved cell viability In vivo: - Improved cardiac function with increased LVEF and LVFS - Reduced infarct size and length, fibrosis area, and apoptosis - Promoted angiogenesis in infarcted myocardium - Hydrogel facilitated prolonged and stable release of exosomes in the area of ischaemic myocardium - Decreased expression of TGF-β1 (pro-fibrosis), CD68 (indicative of inflammatory cell infiltration), and TNF-α (pro-inflammatory) - Increased diameter of CD31+ blood vessels and reduced number of apoptotic cells - Growth hormone releasing peptide-6 (GHRP6) was released following hydrogel degradation, activating pro-survival PI3K/Akt1 and TGF-β1 pathways and inhibiting the NF-kB pathway |
| Huang et al., 2019 [154] | Rat bone marrow MSCs | Exosomes (average 100 nm) | Exosomes (10 μg in 100 μL PBS) were injected at 3 sites around the infarct border 30 min after ligation, with or without intravenous delivery of atorvastatin-pretreated MSCs through the tail vein at day 1, 3, or 7 after MI | In vivo: LAD coronary artery ligation rat model of MI | - Exosome and MSC combinatorial treatment caused improved cardiac function with increased LVES and LVFS, reduced infarct size and collagen area, and increased neovascularisation (microvascular density in both arteriolar and capillary vessels) compared to exosomes or MSCs alone - Intramyocardial injection of exosomes 30 min after AMI combined with MSC transplantation at 3 days after AMI achieved the highest improvement in heart function - Combinatorial therapy reduced inflammation, with increased expression of the pro-survival protein Bcl-2, reduced number of apoptotic cells, and decreased levels of inflammatory cytokines IL-6 and TNF-α |
| Liu et al., 2017 [143] | Rat bone marrow-derived MSCs | Exosomes (50–100 nm) | In vitro: Cells were cultured with exosomes (10 µg/mL) for 6, 12, and 24 h In vivo: Exosomes (5 μg in 10 μL PBS) were injected into the anterior and lateral part of the visibly injured region 5 min before reperfusion | In vitro: Rat H9C2 cardiomyoblasts treated with H2O2 In vivo: Rat I/R model, in which the LAD was ligated for 30 min followed by reperfusion for 2 h | In vitro: - Enhanced cell viability, and reduced cell apoptosis and ROS production after H2O2 stimulation - Increased cell autophagy, which was regulated by AMPK/mTOR and Akt/mTOR signalling; upregulated p-AMPK/AMPK ratio and downregulated p-Akt/Akt and p-mTOR/mTOR ratio In vivo: - Reduced cardiomyocyte apoptosis and infarct size - Improved cardiac function with increased LVEF and LVFS |
| Milano et al., 2020 [150] | Human cardiac-resident mesenchymal progenitor cells (obtained from right cardiac appendage tissue) | Exosomes (mostly < 150 nm) | In vitro: Cells were pre-treated with exosomes for 1 h before inducing oxidative stress In vivo: Exosomes (3 × 1010 particles in 100 μL) were injected into the tail vein at 5, 11, and 19 days after study commencement | In vitro: Neonatal rat ventricular myocytes treated with doxorubicin (Dox) and trastuzumab (Trz) to induce oxidative stress In vivo: Rats injected with six doses of Dox (days 1–11) and six doses of Trz (days 19–28) | In vitro: - Prevented increase in ROS induced by Dox/Trz - Enriched in miR-146a-5p compared to exosomes from human dermal fibroblasts; suppressed expression of Traf6, Smad4, Irak1, Nox4, and Mpo (known target genes of miR-146a-5p) in Dox-treated cells, which might provide protection against Dox-induced cell death In vivo: - Prevented Dox/Trz effects on left ventricular dysfunction, myocardial fibrosis, CD68+ macrophage infiltration, and iNOS expression |
| Shao et al., 2017 [141] | Rat bone marrow-derived MSCs | Exosomes (size unspecified) | In vitro: Cells were cultured with exosomes for up to 48 h In vivo: Exosomes (20 μg in 20 μL PBS) were injected at two different sites beside the infarct border region after ligation | In vitro: Rat H9C2 cardiomyoblasts or BJ fibroblasts treated with TGF-β In vivo: LAD coronary artery ligation rat model of MI | In vitro: - Enhanced proliferation capacity and inhibited apoptosis in H9C2 cells - Reduced TGF-β-induced α-SMA expression and inhibited fibroblast transformation - Compared to MSCs, exosomes had lower expression of miR-21 and miR-15 - Upregulated PI3k-Akt and mTOR pathways In vivo: - Decreased infiltration of CD68+ inflammatory cells, and inhibited apoptosis - Improved cardiac function with increased LVEF and LVFS |
| Shi et al., 2019 [138] | Human umbilical cord MSCs | Exosomes (mostly 100 nm) | Exosomes (400 μg) were given by intramyocardial administration during surgery | In vitro: Rat neonatal cardiomyocytes, and cardiac fibroblasts treated with LPS In vivo: LAD coronary artery ligation rat model of MI | In vitro: - Increased myofibroblast density and improved collagen contraction - Promoted fibroblast-to-myofibroblast differentiation in inflammatory environments - Reduced cardiomyocyte apoptosis - Decreased fibroblast migration, but no effect on fibroblast proliferation - Decreased expression of IL-1β and TNF-α, and increased expression of TGF-β In vivo: - Suppressed inflammatory response and improved cardioprotective effects |
| Sun et al., 2018 [144] | Mouse bone marrow MSCs | Exosomes (average 35 nm) | Exosomes (300 μg in 200 μL PBS) were injected intravenously through tail vein seven days after disease induction | In vivo: Doxorubicin-induced mouse model of dilated cardiomyopathy | - Improved cardiac function with increased LVEF and LVFS - Attenuated cardiac dilation and reduced cardiomyocytes apoptosis - Decreased expression of pro-apoptotic protein Bax, and increased expression of pro-survival protein Bcl-2 - Decreased levels of inflammatory cytokines IL-1, IL-6 and TNF-α in serum - Reduced pro-inflammatory ILY6Chigh and M1-like F4/80+ CD11c+ macrophages, and elevated anti-inflammatory LY6Clow and M2-like F4/80+ CD206+ macrophages - Regulated macrophage polarisation through activation of the JAK2-STAT6 pathway |
| Wang et al., 2018 [153] | Mouse bone marrow MSCs | Exosomes (30–150 nm), engineered through lentiviral packaging technology | Exosomes (4 × 109 particles or 50 μg) in 100 μL were injected intravenously through the tail vein after ligation | In vitro: Hypoxia-induced rat H9C2 cardiomyoblasts In vivo: LAD coronary artery ligation mouse model of MI | In vitro: - IMTP-exosomes produced by transfected MSCs were internalised to a greater extent by hypoxia-injured H9C2 cells than blank exosomes In vivo: - IMTP-exosomes allowed prolonged delivery in the region of ischaemic myocardium - Decreased inflammation and apoptosis, with reduced expression of pro-inflammatory factors (IL-6, TNF-α, IL-1β) - Reduced M1 macrophages (TNF-α+, CD68+) and increased M2 macrophages (CD206+) - Improved revascularisation and cardiac function, with increased capillary density and number of arterioles |
| Xu et al., 2020 [29] | Human MSCs from bone marrow, adipose tissue and umbilical cord | Exosomes (mostly < 100 nm) for all MSC types | Injection of exosomes or MSCs (1.5 × 106 cells) in 150 μL PBS at the margin area of MI 30 min after ligation | In vivo: LAD coronary artery ligation rat model of MI | - Exosomes promoted angiogenesis, reduced infarct size, inhibited cardiomyocyte apoptosis, and improved microvascular density - Decreased LVESD and LVEDD, increased LVEF and LVFS, and improved cardiac function - Increased the levels of angiogenesis factors VEGF, bFGF, and HGF - Decreased adverse cardiac remodelling |
| Xu et al., 2019 [149] | Rat bone marrow-derived MSCs | Exosomes (mostly 100 nm) | Exosomes (from 1 × 106 LPS-primed or non-primed MSCs) were injected at four sites into myocardium around the infarct border zone | In vitro: Rat peritoneal macrophages In vivo: LAD coronary artery ligation mouse model of MI | In vitro: - Exosomes decreased the expression of pro-inflammatory TNF-α, IL-6 and IL-1β, and increased the expression of anti-inflammatory IL-10; LPS-primed exosomes showed greater downregulation of pro-inflammatory cytokines - LPS-primed exosomes significantly reduced pro-inflammatory M1 macrophage protein markers (CD11b, iNOS) and elevated anti-inflammatory M2 macrophage protein markers (CD206, ArgI) - LPS-primed exosomes regulated macrophage polarisation by suppressing NF-κB signalling, and activating AKT1/AKT2 signalling In vivo: - Attenuated post-infarction inflammation by mediating macrophage polarisation, reduced cardiomyocyte apoptosis, and improved cardiac function; LPS-primed exosomes had greater effects |
| Zhao et al., 2019 [145] | Mouse bone marrow MSCs | Exosomes (50–150 nm) | Exosomes (50 μg in 25 μL PBS) were injected at three different points of the peri-infarct myocardial region after reperfusion | In vivo: Mouse I/R model, where the LAD was ligated for 45 min followed by reperfusion; for macrophage depletion studies, mice were intravenously injected with 150 μL (5 mg/mL) clodronate liposomes | - Improved cardiac function; reduced infarct size, fibrosis, hypertrophy of cardiomyocytes, and myocardial injury - Reduced infiltration of inflammatory cells and inflammation of heart tissue - Decreased pro-inflammatory cytokines (IL-6) and increased anti-inflammatory cytokines (IL-10) in serum and heart tissues - Reduced M1 macrophages and M1 gene expression markers (iNOS, IL-1β, IL-6, TNF-α), and increased M2 macrophages and M2 gene expression markers (Arg1, IL-10, CD206, TGF-β) - Downregulated TLR4, and upregulated PI3k/Akt signalling pathway through miR-182 in association with immunomodulation effects |
| Zhu et al., 2019 [146] | Human umbilical cord MSCs | Exosomes (size unspecified) | Exosomes (100 μg/injection) were injected in the tail vein three times per week for six weeks | In vitro: Rat H9C2 cardiomyoblasts treated with treated with H2O2 In vivo: Mouse model of aging and cardiac dysfunction induced by D-galactose | In vitro: - Promoted cell proliferation and prevented senescence - Inhibited the activity of the cell senescence mediator NF-κB and the expression of its subunit p-p65 - Exosomes prevented cell senescence through the lncRNA MALAT1, by inhibiting the NF-κB/TNF-α pathway In vivo: - Improved cardiac function with increased LVEF and LVFS - Attenuated the effects of D-galactose and preserved telomere length - Increased the anti-aging marker TERT and decreased the aging marker p21 - Reduced the expression of pro-inflammatory TNF-α - Protective effects of exosomes were blocked by silencing the lncRNA MALAT1 |
| PREECLAMPSIA | |||||
| Study | Source of EVs | Type of EVs | Administration and Dose | Model(s) Used | Major Findings |
| Liu et al., 2020 [161] | Human umbilical cord MSCs | Exosomes (30–120 nm) | Abdominal injection of exosomes (160 μg/mL) at 0.5 mL/day from day 14 to 19 of gestation | In vitro: HTR-8/SVneo human trophoblast cell line In vivo: Rat preeclampsia model induced by L-NAME | In vitro: - Hindered cell apoptosis by inhibiting c-caspase-3 activity - Increased miR-139-5p expression in trophoblasts, showing pro-angiogenic and anti-inflammatory function - Inhibited PTEN expression and promoted ERK/MMP-2 pathway activation through miR-139-5p In vivo: - Decreased blood pressure and proteinuria - Restored miR-139-3p expression in placental tissue, and reduced expression of PTEN and MMP-2 |
| Wang et al., 2020 [160] | Human umbilical cord MSCs | Exosomes (average 130 nm) | Cells were treated with exosomes for three days; exosomes were from MSCs overexpressing miR-133b | In vitro: HPT-8 and HTR8-S/Vneo human trophoblast cell lines | - Promoted trophoblast proliferation, migration, invasion ability, and cell cycle progression, and inhibited cell apoptosis; effects were more pronounced in exosomes enriched with miR-133b overexpression - In placental tissue of preeclampsia patients, miR-133b is downregulated and SGK1 is up-regulated - Exosomal miR-133b may act by reducing SGK1 expression |
| Xiong et al., 2018 [162] | Human umbilical cord MSCs | Exosomes (30–100 nm) | Abdominal injection of low (120 µg/mL), medium (140 µg/mL), or high (160 µg/mL) dose exosomes at 0.5 mL/day for 6 days, starting on day 14 of gestation | In vivo: Rat preeclampsia model induced by L-NAME | - Decreased blood pressure and proteinuria - Dose-dependent increase in foetus number and quality, as well as microvascular density - Decreased expression of anti-angiogenic sFlt1 and increased expression of pro-angiogenic VEGF in placental tissue, with the high exosome dose showing the most prominent effects |
| Zheng et al., 2020 [159] | Decidual MSCs from human placenta | Not specified, mostly small EVs (224.2 ± 4.2 nm) | Cells were treated with EVs (100 μg/mL, pooled from 6 patient samples) in 100 μL culture medium | In vitro: HUVECs treated with LPS or human preeclampsia serum | - Increased cell attachment and proliferation, and reduced production of pro-inflammatory IL-6 in both HUVEC models - No significant effect on lipid peroxidation in LPS-treated HUVECs, but significantly reduced lipid peroxidation in preeclampsia serum-treated HUVECs |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryan, S.T.; Hosseini-Beheshti, E.; Afrose, D.; Ding, X.; Xia, B.; Grau, G.E.; Little, C.B.; McClements, L.; Li, J.J. Extracellular Vesicles from Mesenchymal Stromal Cells for the Treatment of Inflammation-Related Conditions. Int. J. Mol. Sci. 2021, 22, 3023. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063023
Ryan ST, Hosseini-Beheshti E, Afrose D, Ding X, Xia B, Grau GE, Little CB, McClements L, Li JJ. Extracellular Vesicles from Mesenchymal Stromal Cells for the Treatment of Inflammation-Related Conditions. International Journal of Molecular Sciences. 2021; 22(6):3023. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063023
Chicago/Turabian StyleRyan, Sean T., Elham Hosseini-Beheshti, Dinara Afrose, Xianting Ding, Binbin Xia, Georges E. Grau, Christopher B. Little, Lana McClements, and Jiao Jiao Li. 2021. "Extracellular Vesicles from Mesenchymal Stromal Cells for the Treatment of Inflammation-Related Conditions" International Journal of Molecular Sciences 22, no. 6: 3023. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063023