
Transcriptome Analysis Unravels Key Factors Involved in Response to Potassium Deficiency and Feedback Regulation of K+ Uptake in Cotton Roots

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
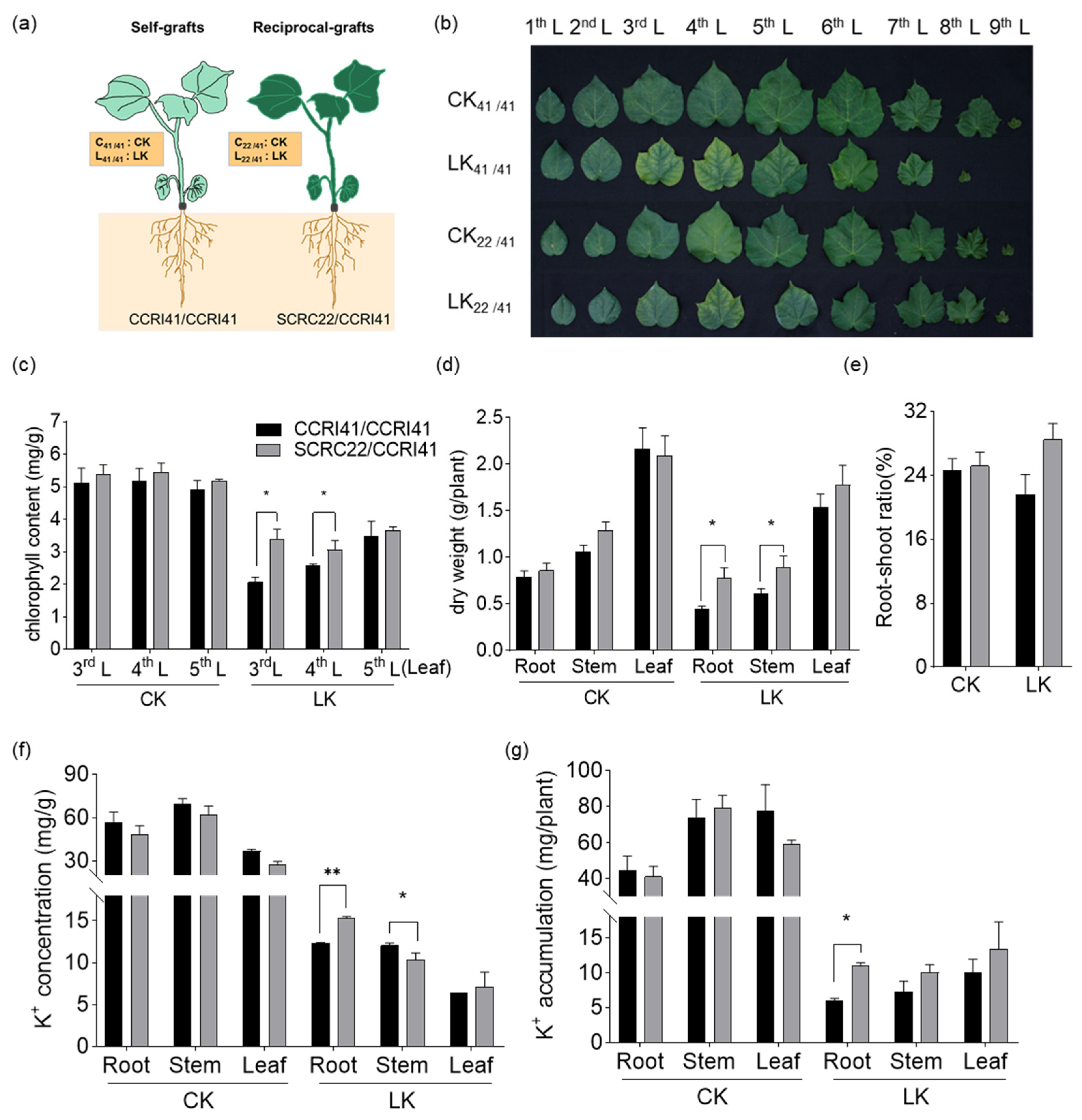
2.1. Effects of Low-K+ Stress on Phenotype, Chlorophyll Content, Biomass, and K+ Content of CCRI41/CCRI41 and SCRC22/CCRI41
2.2. Transcriptome Profiling of CCRI41/CCRI41 and SCRC22/CCRI41 in Response to K+ Deficiency
2.3. Gene Function Enrichment Analysis of DEGs in Response to K+ Deficiency
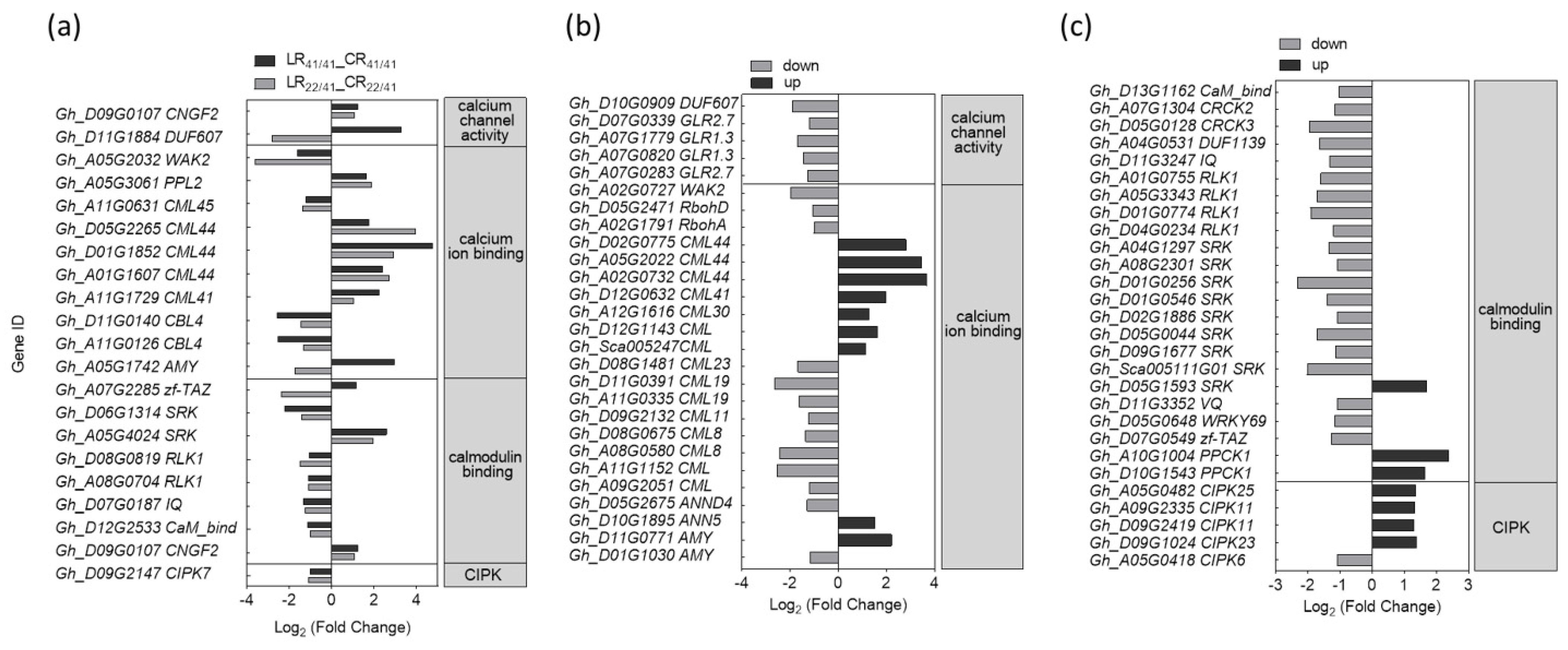
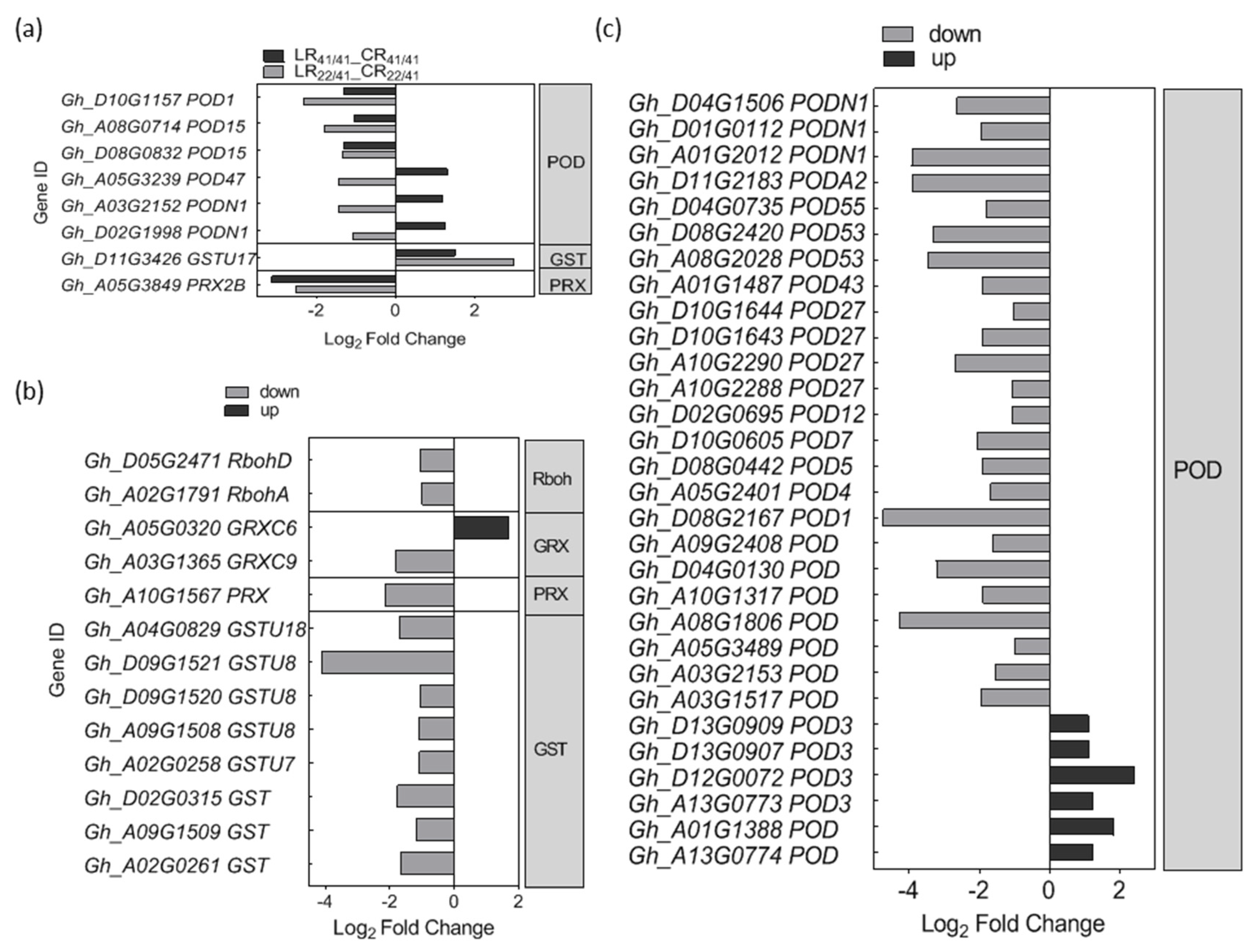
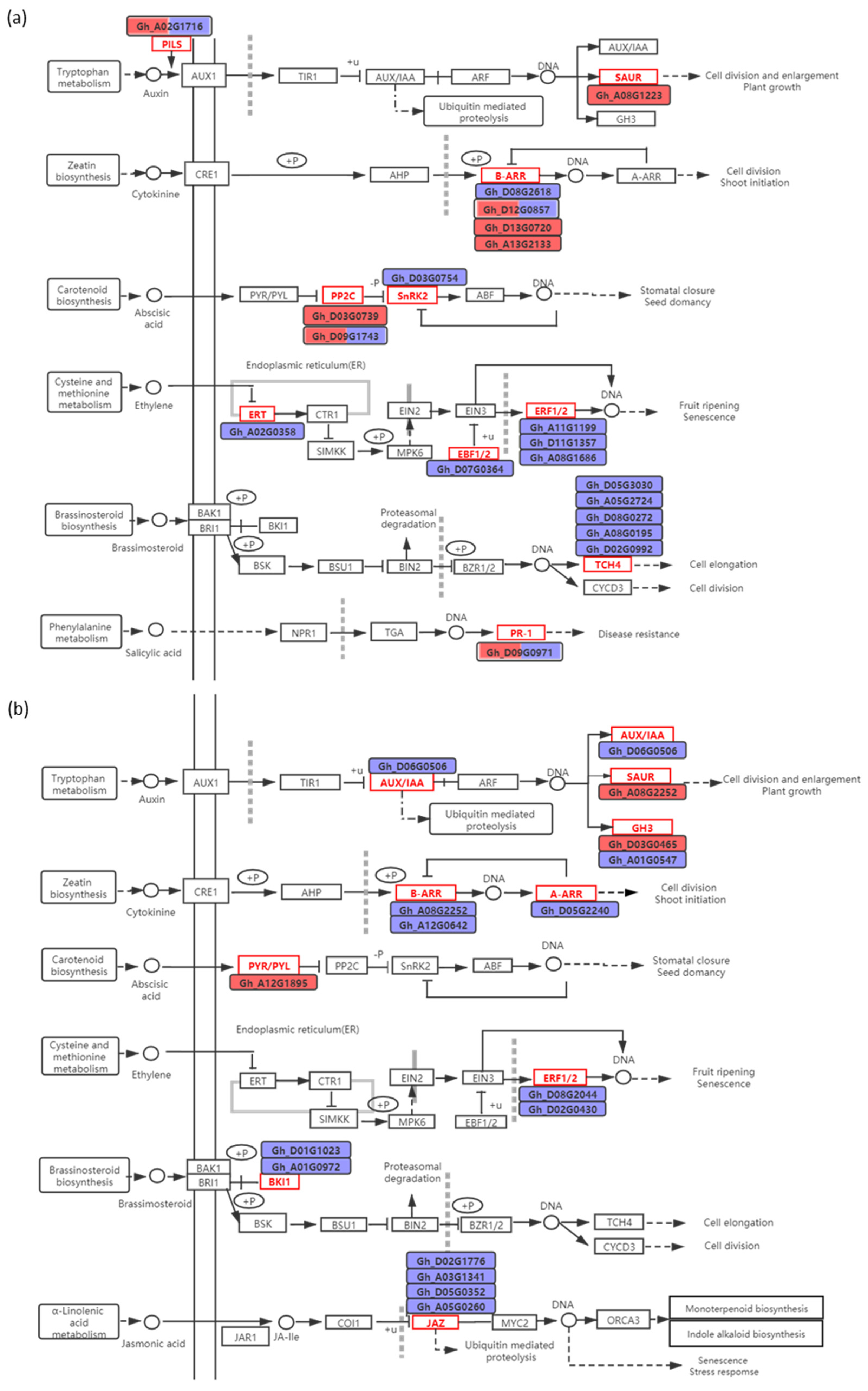
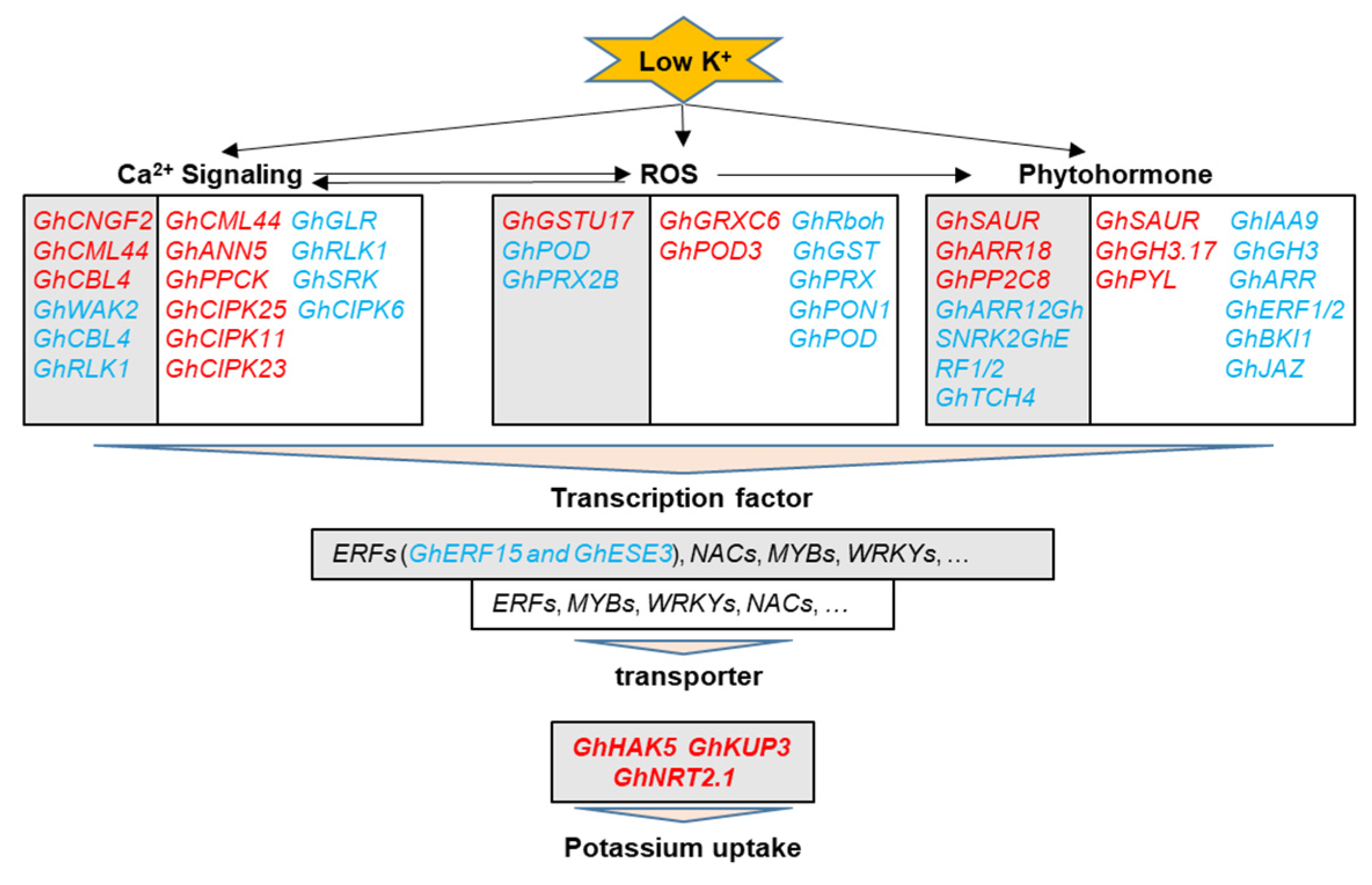
2.4. DEGs Associated with Ca2+, ROS, and Phytohormone Signaling Pathway in Response to K+ Deficiency
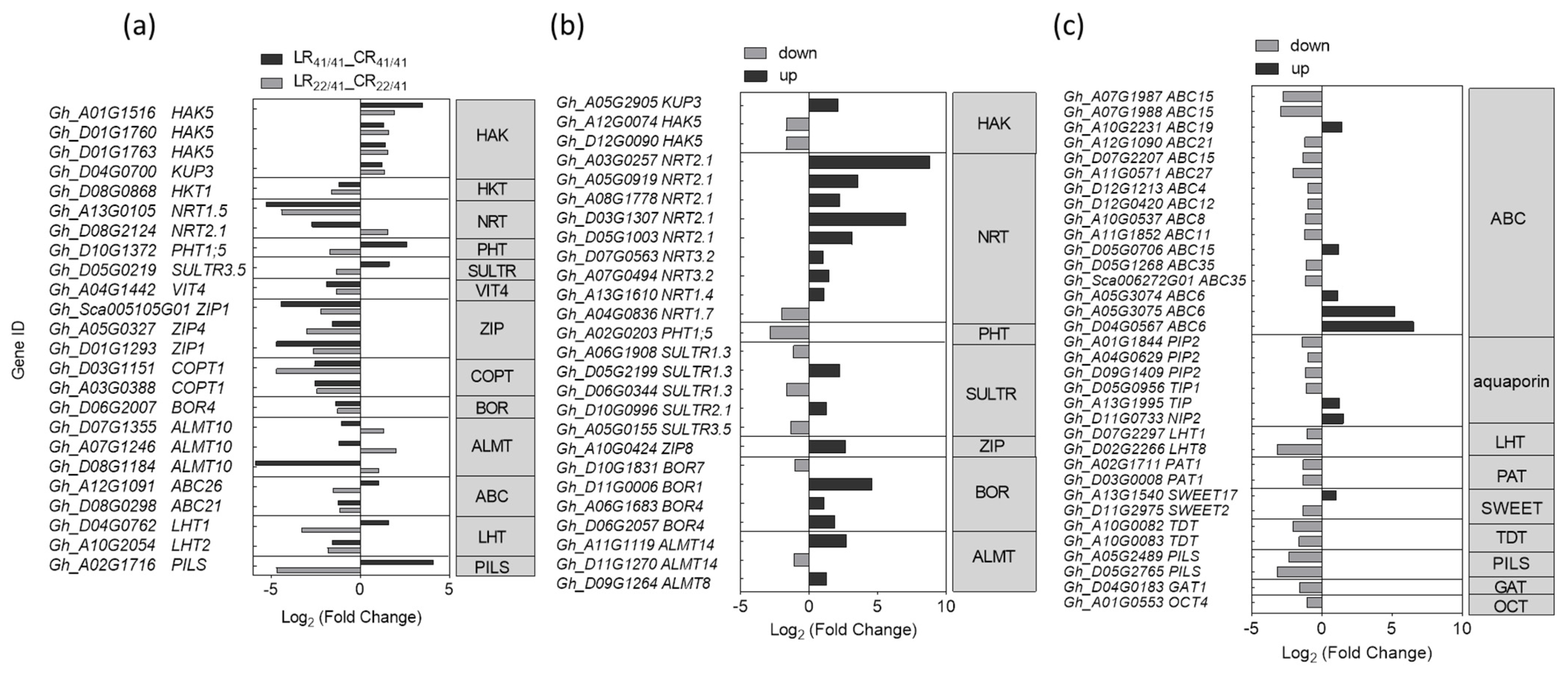
2.5. DEGs Associated with Transporters in Response to K+ Deficiency
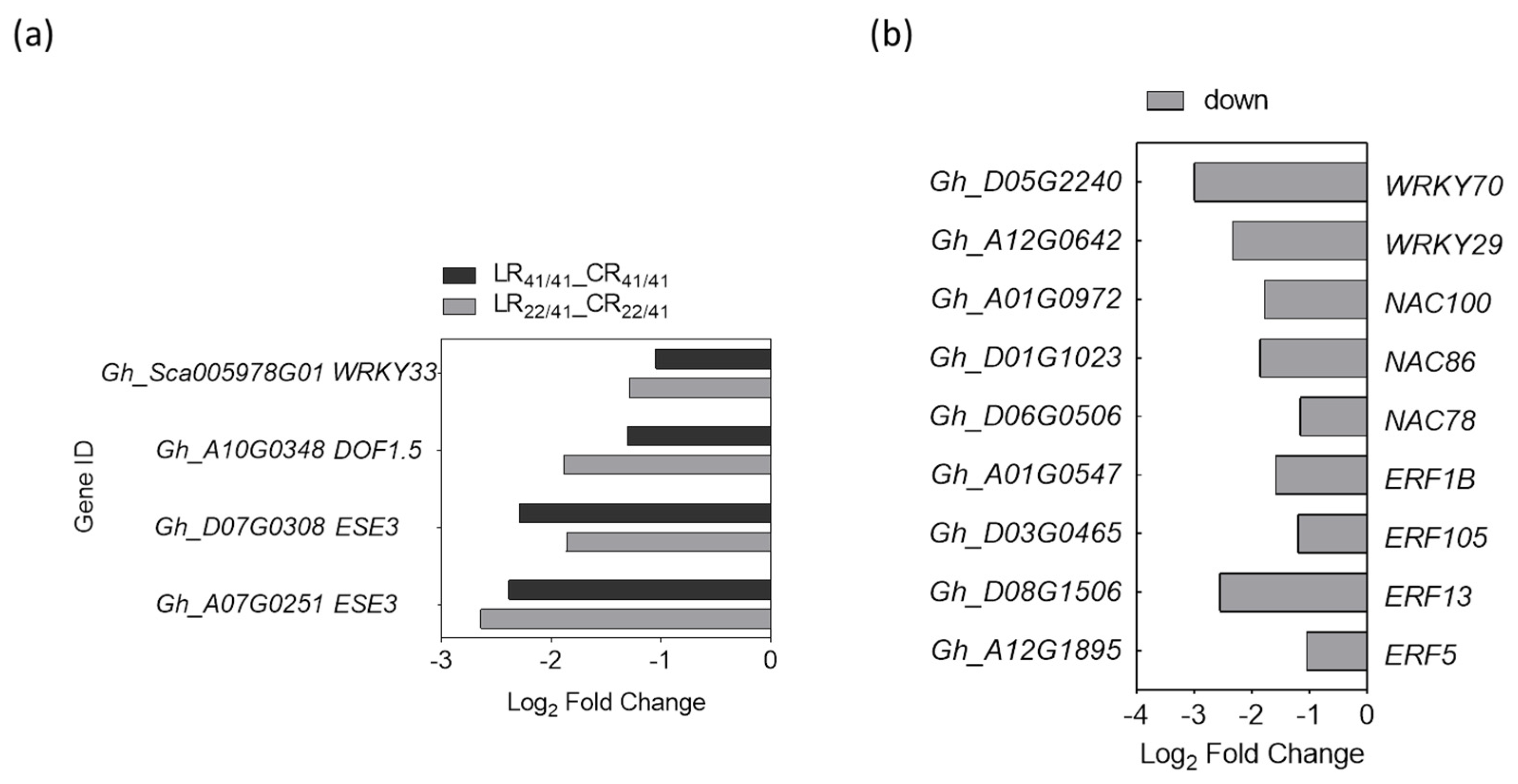
2.6. DEGs Associated with Transcription Factors (TFs) in Response to K+ Deficiency
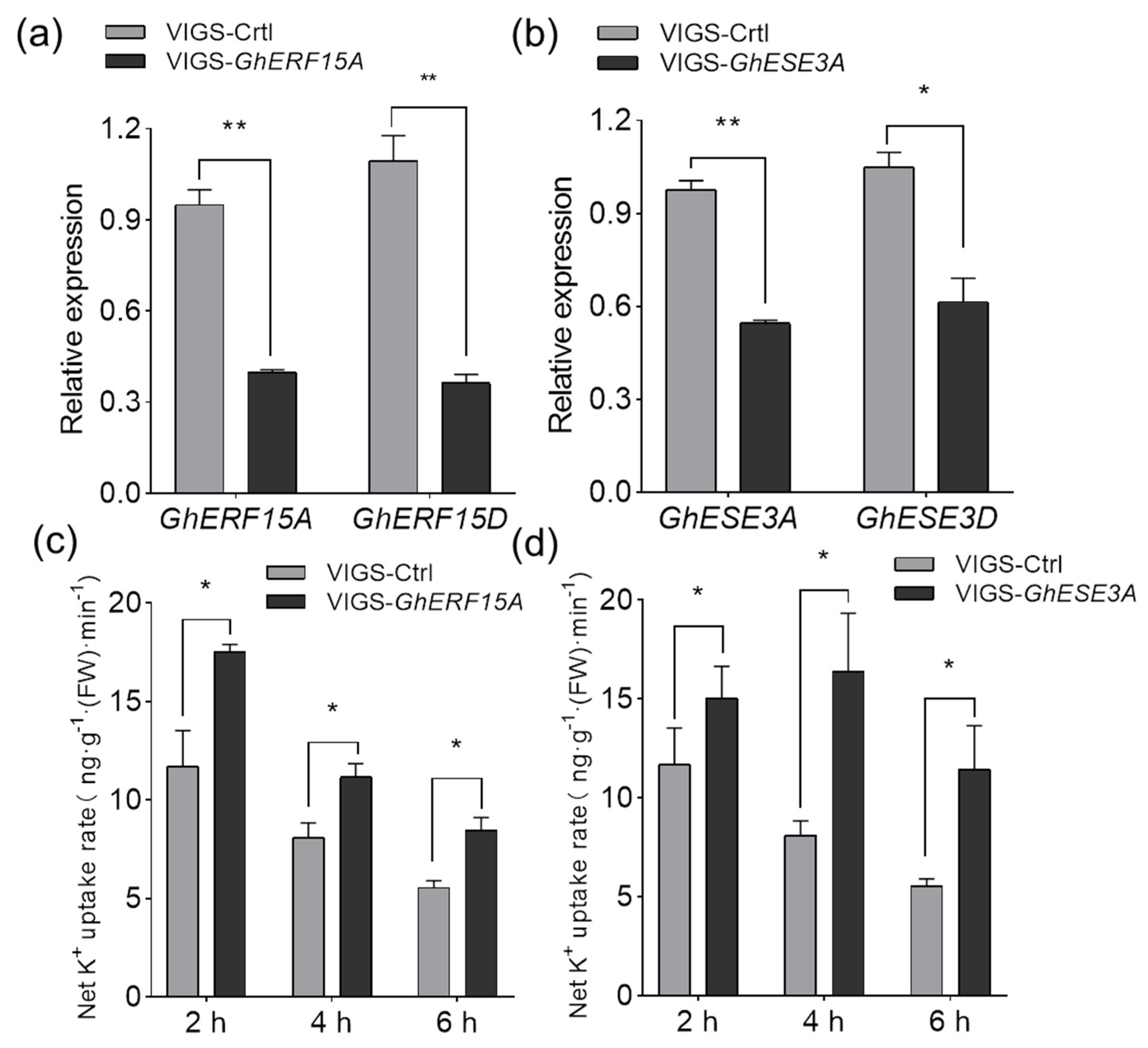
2.7. GhERF15A and GhESE3A Are Probably Involved in Negatively Regulating K+ Uptake under K+ Deficiency
3. Discussion
3.1. Ca2+ and ROS Signaling Involved in the Response to K+ Deficiency and Feedback Regulation of K+ Uptake and Root Growth
3.2. Phytohormone Signaling Involved in Response to K+ Deficiency and Feedback Regulation of K+ Uptake and Root Growth
3.3. Transporters Involved in the Response to K+ Deficiency and Feedback Regulation of K+ Uptake
3.4. Transcription Factors Involved in the Response to K+ Deficiency and Feedback Regulation of K+ Uptake and Root Growth
4. Materials and methods
4.1. Plant Materials and Growth Condition
4.2. Photography of K+ Deficiency Symptoms and Measurement of Biomass, Chlorophyll and K+ Content
4.3. RNA Isolation, Library Construction, and Sequencing
4.4. Read Mapping and Analysis of RNA-Seq
4.5. Functional Annotation of DEGs
4.6. Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR) Analysis
4.7. Prediction of TFs Binding to the Promoter of K+ Transporter GhHAK5 (Gh_D01G1760)
4.8. Detect the Transcription Pattern of Candidate Genes
4.9. Determination of Net K+ Uptake Rate in Virus-Induced Gene Silencing (VIGS ) Plants
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ahanger, M.A.; Tittal, M.; Mir, R.A.; Agarwal, R.M. Alleviation of water and osmotic stress-induced changes in nitrogen metabolizing enzymes in Triticum aestivum L. cultivars by potassium. Protoplasma 2017, 254, 1953–1963. [Google Scholar] [CrossRef] [PubMed]
- Maathuis, F.J.M. Physiological functions of mineral macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Pettigrew, W.T. Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol. Plant. 2008, 133, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Y.F.; Wu, W.H. Potassium and phosphorus transport and signaling in plants. J. Integr. Plant Biol. 2020, 63, 34–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, W.H. Potassium transport and signaling in higher plants. Annu. Rev. Plant Biol. 2013, 64, 451–476. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, W.H. Regulation of potassium transport and signaling in plants. Curr. Opin. Plant Biol. 2017, 39, 123–128. [Google Scholar] [CrossRef]
- de Bang, T.C.; Husted, S.; Laursen, K.H.; Persson, D.P.; Schjoerring, J.K. The molecular-physiological functions of mineral macronutrients and their consequences for deficiency symptoms in plants. New Phytol. 2020, 229, 5. [Google Scholar]
- Seybold, H.; Trempel, F.; Scheel, R.; Dierk, S.; Romeis, T.; Lee, J. Ca2+ signalling in plant immune response: From pattern recognition receptors to Ca2+ decoding mechanisms. New Phytol. 2014, 204, 782–790. [Google Scholar] [CrossRef]
- Steinhorst, L.; Kudla, J. Calcium and reactive oxygen species rule the waves of signaling. Plant Physiol. 2013, 163, 471–485. [Google Scholar] [CrossRef] [Green Version]
- Schulz, P.; Romeis, H.T. Calcium-Dependent Protein Kinases: Hubs in Plant Stress Signaling and Development. Plant Physiol. 2013, 163, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.K.; Shankar, A.; Nalini Chandran, A.K.; Sharma, M.; Jung, K.H.; Suprasanna, P.; Pandey, G.K. Emerging concepts of potassium homeostasis in plants. J. Exp. Bot. 2020, 71, 608–619. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.; Lee, S.; Che, Y.; Jiang, Y.; Luan, S. Mechanistic analysis of AKT1 regulation by the CBL-CIPK-PP2CA interactions. Mol. Plant 2011, 4, 527–536. [Google Scholar] [CrossRef]
- Ragel, P.; Rodenas, R.; Garcia-Martin, E.; Andres, Z.; Villalta, I.; Nieves-Cordones, M.; Rivero, R.M.; Martinez, V.; Pardo, J.M.; Quintero, F.J.; et al. The CBL-Interacting Protein Kinase CIPK23 Regulates HAK5-Mediated High-Affinity K+ Uptake in Arabidopsis Roots. Plant Physiol. 2015, 169, 2863–2873. [Google Scholar] [PubMed] [Green Version]
- Gajdanowicz, P.; Michard, E.; Sandmann, M.; Rocha, M.; Correa, L. Potassium (K+) gradients serve as a mobile energy source in plant vascular tissues. Proc. Natl. Acad. Sci. USA 2011, 108, 864–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Held, K.; Pascaud, F.; Eckert, C.; Gajdanowicz, P.; Hashimoto, K.; Corratgé-Faillie, C.; Offenborn, J.N.; Lacombe, B.; Dreyer, I.; Thibaud, J.-B.; et al. Calcium-dependent modulation and plasma membrane targeting of the AKT2 potassium channel by the CBL4/CIPK6 calcium sensor/protein kinase complex. Cell Res. 2011, 21, 1116–1130. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.H.; Yang, G.W. Signal Function Studies of ROS, Especially RBOH-Dependent ROS, in Plant Growth, Development and Environmental Stress. J. Plant Growth Regul. 2019, 39, 157–171. [Google Scholar] [CrossRef]
- An, J.; Hu, P.G.; Li, F.J.; Wu, H.H.; Shen, Y.; White, J.C.; Tian, X.L.; Li, Z.H.; Giraldo, J.P. Emerging investigator series: Molecular mechanisms of plant salinity stress tolerance improvement by seed priming with cerium oxide nanoparticles. Environ. Sci. Nano 2020, 7, 2214–2228. [Google Scholar] [CrossRef]
- Gupta, D.K.; Palma, J.M.; Corpas, F.J. Reactive Oxygen Species and Oxidative Damage in Plants Under Stress || Redox Regulation and Antioxidant Defence During Abiotic Stress: What Have We Learned from Arabidopsis and Its Relatives; Springer: Berlin/Heidelberg, Germany, 2015; pp. 83–113. [Google Scholar]
- Zhang, Z.Y.; Wang, Q.L.; Li, Z.H.; Duan, L.S.; Tian, X.L. Effect of Potassium Deficiency on Root Growth of Cotton (Gossypium hirsutum L.) Seedlings and Its Physiological Mechanisms Involved. Acta Agron. Sin. 2009, 35, 718–723. [Google Scholar] [CrossRef]
- Sustr, M.; Soukup, A.; Tylova, E. Potassium in Root Growth and Development. Plants 2019, 8, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osakabe, Y.; Arinaga, N.; Umezawa, T.; Katsura, S.; Nagamachi, K.; Tanaka, H.; Ohiraki, H.; Yamada, K.; Seo, S.U.; Abo, M. Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis. Plant Cell 2013, 25, 609–624. [Google Scholar] [CrossRef] [Green Version]
- Rigas, S.; Debrosses, G.; Haralampidis, K.; Vicente-Agullo, F.; Feldmann, K.; Grabov, A.; Dolan, L.; Hatzopoulos, P. TRH1 encodes a potassium transporter required for tip growth in Arabidopsis root hairs. Plant Cell 2001, 13, 139–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente-Agullo, F.; Rigas, S.; Desbrosses, G.; Dolan, L.; Hatzopoulos, P.; Grabov, A. Potassium carrier TRH1 is required for auxin transport in Arabidopsis roots. Plant J. Cell Mol. Biol. 2004, 40, 523–535. [Google Scholar] [CrossRef]
- Nam, Y.J.; Tran, L.S.; Kojima, M.; Sakakibara, H.; Nishiyama, R.; Shin, R. Regulatory roles of cytokinins and cytokinin signaling in response to potassium deficiency in Arabidopsis. PLoS ONE 2012, 7, e47797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.Z.; Wu, Y.F.; Liu, G.Y.; Xiao, X.H.; Wang, P.F.; Gao, T.; Xu, M.J.; Han, Q.X.; Wang, Y.H.; Guo, T.C.; et al. Large-scale Proteomics Combined with Transgenic Experiments Demonstrates An Important Role of Jasmonic Acid in Potassium Deficiency Response in Wheat and Rice. Mol. Cell Proteom. 2017, 16, 1889–1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.G.; Kim, B.G.; Cheong, Y.H.; Pandey, G.K.; Luan, S. A Ca2+ signaling pathway regulates a K+ channel for low-K response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 12625–12630. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Li, H.D.; Chen, L.Q.; Wang, Y.; Liu, L.L.; He, L.; Wu, W.H. A Protein Kinase, Interacting with Two Calcineurin B-like Proteins, Regulates K+ Transporter AKT1 in Arabidopsis. Cell 2006, 125, 1347–1360. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Lan, W.Z.; Kim, B.G.; Li, L.; Cheong, Y.H.; Pandey, G.K. A protein phosphorylation/dephosphorylation network regulates a plant potassium channel. Proc. Natl. Acad. Sci. USA 2007, 104, 15959–15964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.Y.; Shin, R.; Schachtman, D.P. Ethylene mediates response and tolerance to potassium deprivation in Arabidopsis. Plant Cell 2009, 21, 607–621. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.; Shin, R.; Schachtman, D. Expression of KT/KUP genes in Arabidopsis and the role of root hairs in K+ uptake. Plant Physiol. 2004, 134, 1135–1145. [Google Scholar] [CrossRef] [Green Version]
- Armengaud, P.; Breitling, R.; Amtmann, A. The potassium-dependent transcriptome of Arabidopsis reveals a prominent role of jasmonic acid in nutrient signaling. Plant Physiol. 2004, 136, 2556–2576. [Google Scholar] [CrossRef] [Green Version]
- Takehisa, H.; Sato, Y.; Antonio, B.A.; Nagamura, Y. Global transcriptome profile of rice root in response to essential macronutrient deficiency. Plant Signal. Behav. 2014, 8, e24409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armengaud, P.; Breitling, R.; Amtmann, A. Coronatine-Insensitive 1 (COI1) Mediates Transcriptional Responses of Arabidopsis thaliana to External Potassium Supply. Mol. Plant 2010, 3, 390–405. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.L.; Wu, W.H.; Wang, Y. Transcriptome analysis of rice root responses to potassium deficiency. BMC Plant Biol. 2012, 12, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.B.; He, X.Y.; Wu, D.Z.; Zhu, B.; Cai, S.G.; Nadira, U.A.; Jabeen, Z.; Zhang, G. Comparative Transcriptome Profiling of Two Tibetan Wild Barley Genotypes in Responses to Low Potassium. PLoS ONE 2014, 9, e100567. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.L.; Huang, Y.; Zhong, Y.Q.; Kong, Q.S.; Xie, J.J.; Niu, M.L.; Xu, Y.; Bie, Z.L. Comparative transcriptome profiling of potassium starvation responsiveness in two contrasting watermelon genotypes. Planta 2013, 239, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, Y.; Jin, Y.M.; Kan, L.P.; Dong, C.X. Transcriptome Analysis of Pyrus betulaefolia Seedling Root Responses to Short-Term Potassium Deficiency. Int. J. Mol. Sci. 2020, 21, 8857. [Google Scholar] [CrossRef]
- Gierth, M.; Maser, P.; Schroeder, J.I. The potassium transporter AtHAK5 functions in K+ deprivation-induced high-affinity K+ uptake and AKT1 K+ channel contribution to K+ uptake kinetics in Arabidopsis roots. Plant Physiol. 2005, 137, 1105–1114. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Cordero, M.A. High-affinity K+ uptake in pepper plants. J. Exp. Bot. 2005, 56, 1553–1562. [Google Scholar] [CrossRef] [Green Version]
- Nieves-Cordones, M.; Martínez-Cordero, M.A.; Martínez, V.; Rubio, F. An NH4+-sensitive component dominates high-affinity K+ uptake in tomato plants. Plant Sci. 2007, 172, 273–280. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Li, B.; Xiong, C.; Eneji, A.E.; Zhang, M.; Li, F.; Tian, X.; Li, Z. The Cotton High-Affinity K+ Transporter, GhHAK5a, Is Essential for Shoot Regulation of K+ Uptake in Root under Potassium Deficiency. Plant Cell Physiol. 2019, 60, 888–899. [Google Scholar] [CrossRef]
- Kim, M.J.; Ruzicka, D.; Shin, R.; Schachtman, D.P. The Arabidopsis AP2/ERF transcription factor RAP2.11 modulates plant response to low-potassium conditions. Mol. Plant 2012, 5, 1042–1057. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Zhang, M.L.; Ma, T.L.; Wang, Y. Phosphorylation of ARF2 Relieves Its Repression of Transcription of the K+ Transporter Gene HAK5 in Response to Low Potassium Stress. Plant Cell 2016, 28, 3005–3019. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Y.; Chao, M.N.; Wang, S.F.; Bu, J.J.; Tang, J.X.; Li, F.; Wang, Q.L.; Zhang, B.H. Proteome quantification of cotton xylem sap suggests the mechanisms of potassium-deficiency-induced changes in plant resistance to environmental stresses. Sci. Rep. 2016, 6, 21060. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.B.; Quan, X.Y.; He, X.Y.; Cai, S.G.; Ye, Z.L.; Chen, G.; Zhang, G.P. Root and leaf metabolite profiles analysis reveals the adaptive strategies to low potassium stress in barley. BMC Plant Biol. 2018, 18, 187. [Google Scholar] [CrossRef] [PubMed]
- Li, L.Q.; Lyu, C.C.; Huang, L.P.; Chen, Q.; Zhuo, W.; Wang, X.Y.; Lu, Y.F.; Zeng, F.C.; Lu, L.M. Physiology and proteomic analysis reveals root, stem and leaf responses to potassium deficiency stress in alligator weed. Sci. Rep. 2019, 9, 17366. [Google Scholar] [CrossRef] [PubMed]
- He, Y.D.; Li, R.M.; Lin, F.; Xiong, Y.; Wang, L.X.; Wang, B.Z.; Guo, J.C.; Hu, C.X. Transcriptome Changes Induced by Different Potassium Levels in Banana Roots. Plants 2019, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Hu, Y.; Jiang, W.; Fang, L.; Guan, X.; Chen, J.; Zhang, J.; Saski, C.A.; Scheffler, B.E.; Stelly, D.M.; et al. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nat. Biotechnol. 2015, 33, 531–537. [Google Scholar] [CrossRef] [Green Version]
- Véry, A.; Nieves-Cordones, M.; Daly, M.; Khan, I.; Fizames, C.; Sentenac, H. Molecular biology of K+ transport across the plant cell membrane: What do we learn from comparison between plant species? J. Plant Physiol. 2014, 171, 748–769. [Google Scholar] [CrossRef] [PubMed]
- Forieri, I.; Sticht, C.; Reichelt, M.; Gretz, N.; Hawkesford, M.J.; Malagoli, M.; Wirtz, M.; Hell, R. System analysis of metabolism and the transcriptome in Arabidopsis thaliana roots reveals differential co-regulation upon iron, sulfur and potassium deficiency. Plant Cell Environ. 2017, 40, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.C.; Magwanga, R.O.; Yang, X.; Jin, D.S.; Cai, X.Y.; Hou, Y.Q.; Wei, Y.Y.; Zhou, Z.L.; Wang, K.B.; Liu, F. Genetic regulatory networks for salt-alkali stress in Gossypium hirsutum with differing morphological characteristics. BMC Genom. 2020, 21, 15. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Xing, H.; Zeng, W.; Xu, J.; Mao, L.; Wang, L.; Feng, W.; Tao, J.; Wang, H.; Zhang, H.; et al. Genome-wide association and differential expression analysis of salt tolerance in Gossypium hirsutum L at the germination stage. BMC Plant Biol. 2019, 19, 394. [Google Scholar] [CrossRef]
- Han, M.G.; Lu, X.K.; Yu, J.; Chen, X.G.; Wang, X.G.; Malik, W.; Wang, J.J.; Wang, D.L.; Wang, S.; Guo, L.X.; et al. Transcriptome Analysis Reveals Cotton (Gossypium hirsutum) Genes That Are Differentially Expressed in Cadmium Stress Tolerance. Int. J. Mol. Sci. 2019, 20, 1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudd, J.J.; Franklin-Tong, V.E. Unravelling response-specificity in Ca2+ signalling pathways in plant cells. New Phytol. 2010, 151, 7–33. [Google Scholar] [CrossRef]
- Shin, R.; Schachtman, D. Hydrogen peroxide mediates plant root cell response to nutrient deprivation. Proc. Natl. Acad. Sci. USA 2004, 101, 8827–8832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schachtman, D.P. The Role of Ethylene in Plant Responses to K+ Deficiency. Front. Plant Sci. 2015, 6, 1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Cao, H.S.; Yang, L.; Chen, C.; Shabala, L.; Xiong, M.; Niu, M.L.; Liu, J.; Zheng, Z.H.; Zhou, L.J.; et al. Tissue-specific respiratory burst oxidase homolog-dependent H2O2 signaling to the plasma membrane H+-ATPase confers potassium uptake and salinity tolerance in Cucurbitaceae. J. Exp. Bot. 2019, 70, 5879–5893. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Maathuis, F.; Voitsekhovskaja, O. Unravelling the plant signalling machinery: An update on the cellular and genetic basis of plant signal transduction. Funct. Plant Biol. 2018, 45, 1–8. [Google Scholar] [CrossRef]
- Zhao, X.M.; Liu, Y.; Liu, X.; Jiang, J. Comparative Transcriptome Profiling of Two Tomato Genotypes in Response to Potassium-Deficiency Stress. Int. J. Mol. Sci. 2018, 19, 2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.K.; Tu, B.J.; Wang, X.; Li, Y.S.; Zhang, Q.Y.; Liu, X.B. Transcript Profile in Vegetable Soybean Roots Reveals Potential Gene Patterns Regulating K Uptake Efficiency. Agronomy 2020, 10, 1796. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, Z.J.; Yao, X.D.; Chen, J.L.; Jin, Y. Genome-wide identification and characterization of small auxin-up RNA (SAUR) gene family in plants: Evolution and expression profiles during normal growth and stress response. BMC Plant Biol. 2021, 21, 4. [Google Scholar] [CrossRef]
- Jeong, J.; Park, S.; Im, J.H.; Yi, H. Genome-wide identification of GH3 genes in Brassica oleracea and identification of a promoter region for anther-specific expression of a GH3 gene. BMC Genom. 2021, 22, 22. [Google Scholar] [CrossRef] [PubMed]
- Kirungu, J.N.; Magwanga, R.O.; Lu, P.; Cai, X.Y.; Liu, F. Functional characterization of Gh_A08G1120 (GH3.5) gene reveal their significant role in enhancing drought and salt stress tolerance in cotton. BMC Genet. 2019, 20, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, K.; Chasman, D.; Roy, S.; Ané, J.M. Physiological Responses and Gene Co-Expression Network of Mycorrhizal Roots under K+ Deprivation. Plant Physiol. 2017, 173, 1811–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iliev, E.A.; Xu, W.; Polisensky, D.H.; Oh, M.H.; Torisky, R.S.; Clouse, S.D.; Braam, J. Transcriptional and posttranscriptional regulation of Arabidopsis TCH4 expression by diverse stimuli. Roles of cis regions and brassinosteroids. Plant Physiol. 2002, 130, 770–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.Q.; Li, J.; Chen, Y.; Lu, Y.F.; Lu, L.M. De novo transcriptome analysis of tobacco seedlings and identification of the early response gene network under low-potassium stress. Genet. Mol. Res. 2016, 15, 15. [Google Scholar] [CrossRef]
- Ruan, L.; Zhang, J.; Xin, X.; Zhang, C.; Ma, D.; Chen, L.; Zhao, B. Comparative analysis of potassium deficiency-responsive transcriptomes in low potassium susceptible and tolerant wheat (Triticum aestivum L.). Sci. Rep. 2015, 5, 10090. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.Y.; Feng, H.M.; Zhang, S.; Xiao, H.J.; Hu, Q.D.; Chen, G.; Xuan, W.; Moran, N.; Murphy, A.; Yu, L.; et al. Potassium transporter OsHAK5 alters rice architecture via ATP-dependent transmembrane auxin fluxes. Plant Commun. 2020, 5, 100052. [Google Scholar] [CrossRef]
- Zhang, M.; Huang, P.; Ji, Y.; Wang, S.; Wang, S.; Li, Z.; Guo, Y.; Ding, Z.; Wu, W.; Wang, Y. KUP9 maintains root meristem activity by regulating K+ and auxin homeostasis in response to low K. Embo Rep. 2020, 21, e50164. [Google Scholar] [CrossRef]
- Hu, W.; Di, Q.; Zhang, J.; Liu, J.; Shi, X.J. Response of grafting tobacco to low potassium stress. BMC Plant Biol. 2020, 20, 286. [Google Scholar] [CrossRef]
- Du, X.Q.; Wang, F.L.; Li, H.; Jing, S.; Yu, M.; Li, J.; Wu, W.H.; Kudla, J.; Wang, Y. The Transcription Factor MYB59 Regulates K+/NO3− Translocation in the Arabidopsis Response to Low K+ Stress. Plant Cell 2019, 31, 699–714. [Google Scholar] [CrossRef] [Green Version]
- Ruffel, S. Nutrient-Related Long-Distance Signals: Common Players and Possible Cross-Talk. Plant Cell Physiol. 2018, 59, 1723–1732. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yu, M.; Du, X.Q.; Wang, Z.F.; Wu, W.H.; Quintero, F.J.; Jin, X.H.; Li, H.D.; Wang, Y. NRT1.5/NPF7.3 Functions as a Proton-Coupled H+/K+ Antiporter for K+ Loading into the Xylem in Arabidopsis. Plant Cell 2017, 29, 2016–2026. [Google Scholar] [CrossRef] [Green Version]
- Bouguyon, E.; Perrine-Walker, F.; Pervent, M.; Rochette, J.; Cuesta, C.; Benkova, E.; Martinière, A.; Bach, L.; Krouk, G.; Gojon, A.; et al. Nitrate Controls Root Development through Posttranscriptional Regulation of the NRT1.1/NPF6.3 Transporter/Sensor. Plant Physiol. 2016, 172, 1237–1248. [Google Scholar] [PubMed] [Green Version]
- Balzergue, C.; Dartevelle, T.; Godon, C.; Laugier, E.; Meisrimler, C.; Teulon, J.; Creff, A.; Bissler, M.; Brouchoud, C.; Hagège, A.; et al. Low phosphate activates STOP1-ALMT1 to rapidly inhibit root cell elongation. Nat. Commun. 2017, 8, 15300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.J.; Nagy, R.; Chen, H.Y.; Pfrunder, S.; Yu, Y.C.; Santelia, D.; Frommer, W.B.; Martinoia, E. SWEET17, a facilitative transporter, mediates fructose transport across the tonoplast of Arabidopsis roots and leaves. Plant Physiol. 2014, 164, 777–789. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.; Huh, J.H.; Yu, Y.C.; Ho, L.H.; Chen, L.Q.; Tholl, D.; Frommer, W.B.; Guo, W.J. The Arabidopsis vacuolar sugar transporter SWEET2 limits carbon sequestration from roots and restricts Pythium infection. Plant J. Cell Mol. Biol. 2015, 83, 1046–1058. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, S.K.; Pandit, E.; Nayak, D.K.; Behera, L.; Mohapatra, T. Genes, pathways and transcription factors involved in seedling stage chilling stress tolerance in indica rice through RNA-Seq analysis. BMC Plant Biol. 2019, 19, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raddatz, N.; Morales de Los Rios, L.; Lindahl, M.; Quintero, F.J.; Pardo, J.M. Coordinated Transport of Nitrate, Potassium, and Sodium. Front. Plant Sci. 2020, 11, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Liu, F.; Crawford, N.; Wang, Y. Molecular Regulation of Nitrate Responses in Plants. Int. J. Mol. Sci. 2018, 19, 2039. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.P.; Takeshi, Y.; Kondou, Y.; Schachtman, D.P.; Matsui, M.; Shin, R. Identification and characterization of transcription factors regulating Arabidopsis HAK5. Plant Cell Physiol. 2013, 54, 1478–1490. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Jing, W.; Xiao, L.Y.; Jin, Y.K.; Shen, L.K.; Zhang, W.H. The Rice High-Affinity Potassium Transporter1;1 Is Involved in Salt Tolerance and Regulated by an MYB-Type Transcription Factor. Plant Physiol. 2015, 168, 1076–1090. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.Y.; Hu, L.Q.; Chen, X.Y.; Zhang, R.Z.; Cheng, D.G.; Li, H.S.; Xu, Z.S.; Li, L.C.; Zhou, Y.B.; Liu, A.F.; et al. Genome-wide analysis and identification of the low potassium stress responsive gene SiMYB3 in foxtail millet (Setariaitalica L.). BMC Genom. 2019, 20, 136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Q.; Jiang, H.; Wang, H.; Cui, J.; Wang, J.H.; Hu, J.; Guo, L.B.; Qian, Q.; Xue, D.W. Transcriptome Analysis of Rice Seedling Roots in Response to Potassium Deficiency. Sci. Rep. 2017, 7, 5523. [Google Scholar] [CrossRef] [Green Version]
- Tu, X.Y.; Mejía-Guerra, M.K.; Valdes Franco, J.A.; Tzeng, D.; Chu, P.Y.; Shen, W.; Wei, Y.Y.; Dai, X.R.; Li, P.H.; Buckler, E.S.; et al. Reconstructing the maize leaf regulatory network using ChIP-seq data of 104 transcription factors. Nat. Commun. 2020, 11, 5089. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, Y.; Zhang, Z.; Wang, B.; Eneji, A.E.; Duan, L.; Li, Z.; Tian, X. Cotton shoot plays a major role in mediating senescence induced by potassium deficiency. J. Plant Physiol. 2012, 169, 327–335. [Google Scholar] [CrossRef]
- Tang, D.; Qian, H.; Zhao, L.; Huang, D.; Tang, K. Transgenic tobacco plants expressing BoRS1 gene from Brassica oleracea var. acephala show enhanced tolerance to water stress. J. Biosci. 2005, 30, 647–655. [Google Scholar] [PubMed]
- Xu, J.; Tian, X.L.; Egrinya, E.A.; Li, Z.H. Functional characterization of GhAKT1, a novel Shaker-like K⁺ channel gene involved in K⁺ uptake from cotton (Gossypium hirsutum). Gene 2014, 545, 61–71. [Google Scholar] [CrossRef]
- Siren, J.; Valimaki, N.; Makinen, V. Indexing Graphs for Path Queries with Applications in Genome Research. IEEE/ACM Trans. Comput. Biol. Bioinform. 2014, 11, 375–388. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, D.; Li, F.; Yi, F.; Eneji, A.E.; Tian, X.; Li, Z. Transcriptome Analysis Unravels Key Factors Involved in Response to Potassium Deficiency and Feedback Regulation of K+ Uptake in Cotton Roots. Int. J. Mol. Sci. 2021, 22, 3133. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063133
Yang D, Li F, Yi F, Eneji AE, Tian X, Li Z. Transcriptome Analysis Unravels Key Factors Involved in Response to Potassium Deficiency and Feedback Regulation of K+ Uptake in Cotton Roots. International Journal of Molecular Sciences. 2021; 22(6):3133. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063133
Chicago/Turabian StyleYang, Doudou, Fangjun Li, Fei Yi, A. Egrinya Eneji, Xiaoli Tian, and Zhaohu Li. 2021. "Transcriptome Analysis Unravels Key Factors Involved in Response to Potassium Deficiency and Feedback Regulation of K+ Uptake in Cotton Roots" International Journal of Molecular Sciences 22, no. 6: 3133. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063133