
Regulatory Role of Sugars on the Settlement Inducing Activity of a Conspecific Cue in Pacific Oyster Crassostrea gigas

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
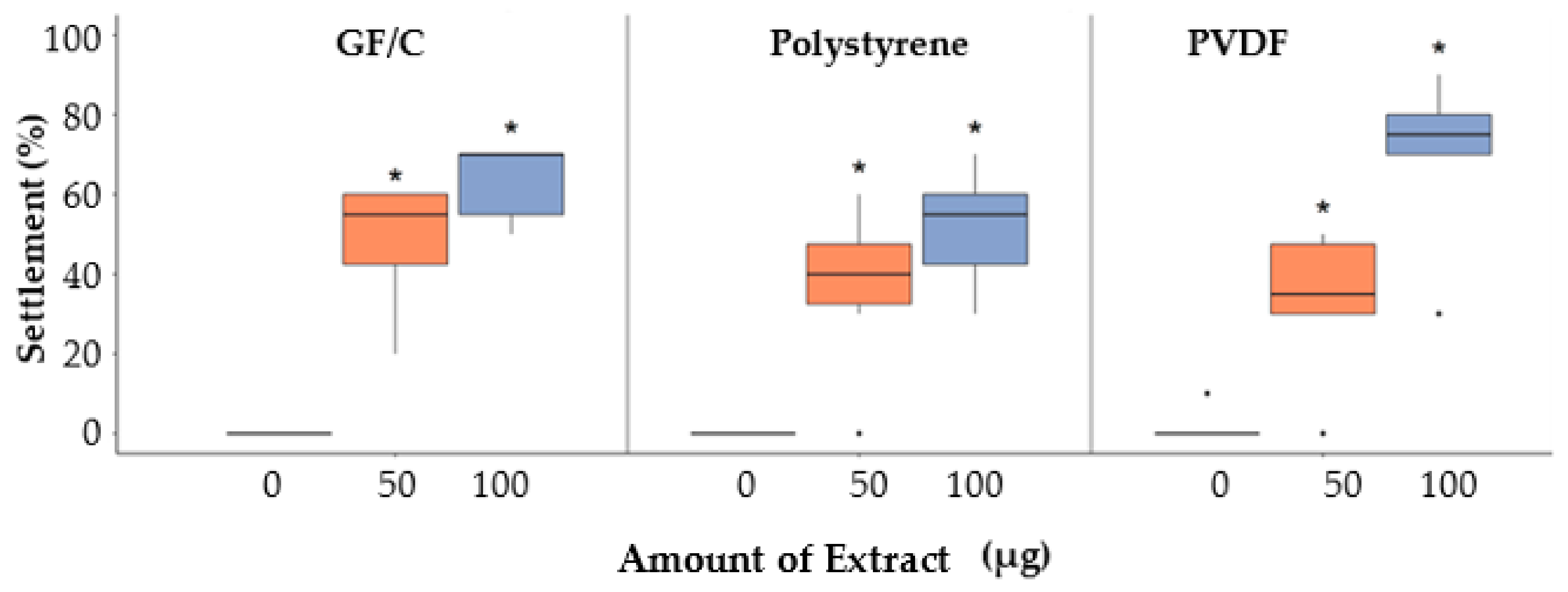
2.1. Settlement Inducing Activity of Crassostrea gigas Shell EDTA (CgSE) Extract and Larval Substrate Specificity
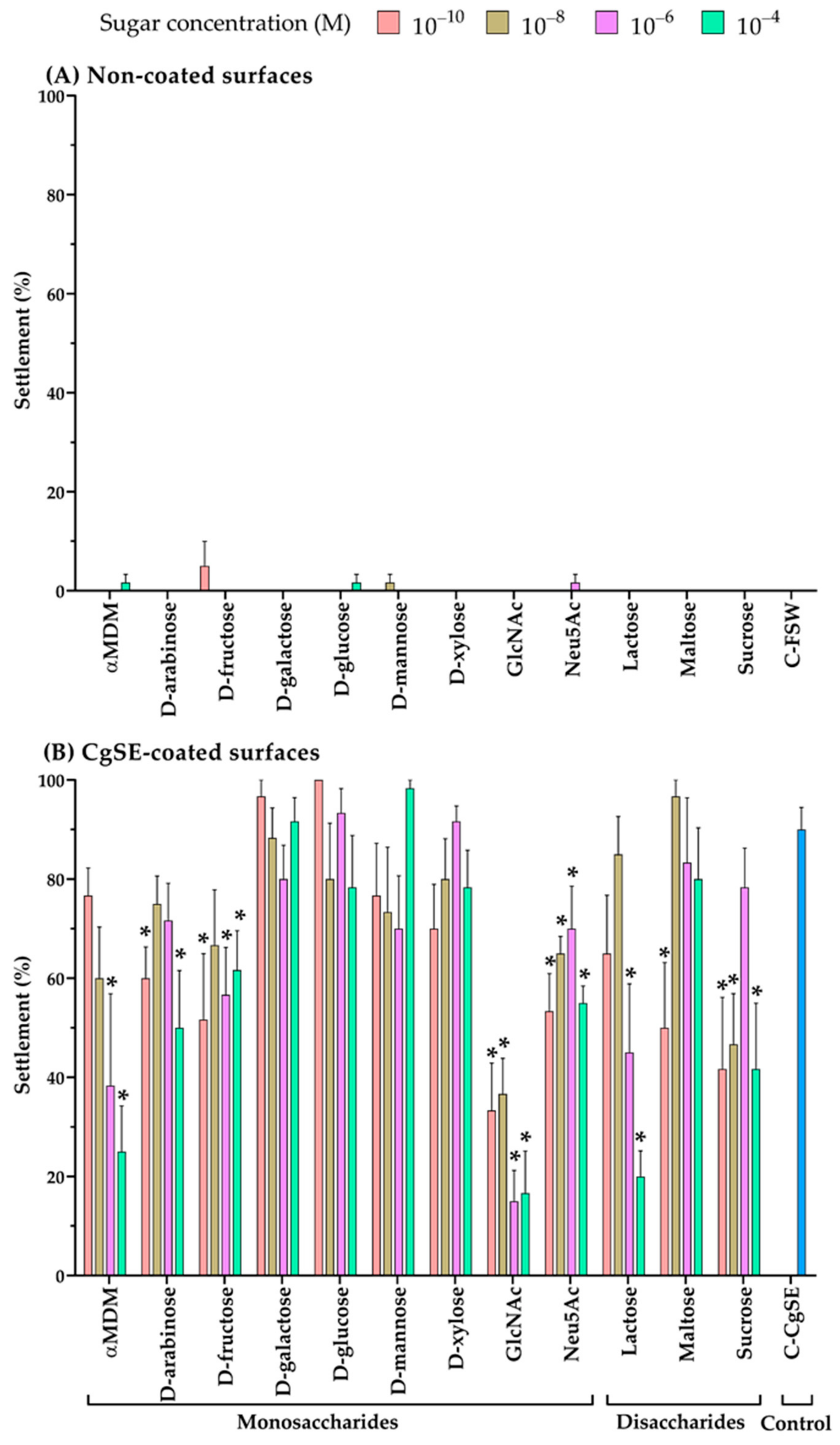
2.2. Sugars and CgSE Effects on the Settlement of Oyster Larvae
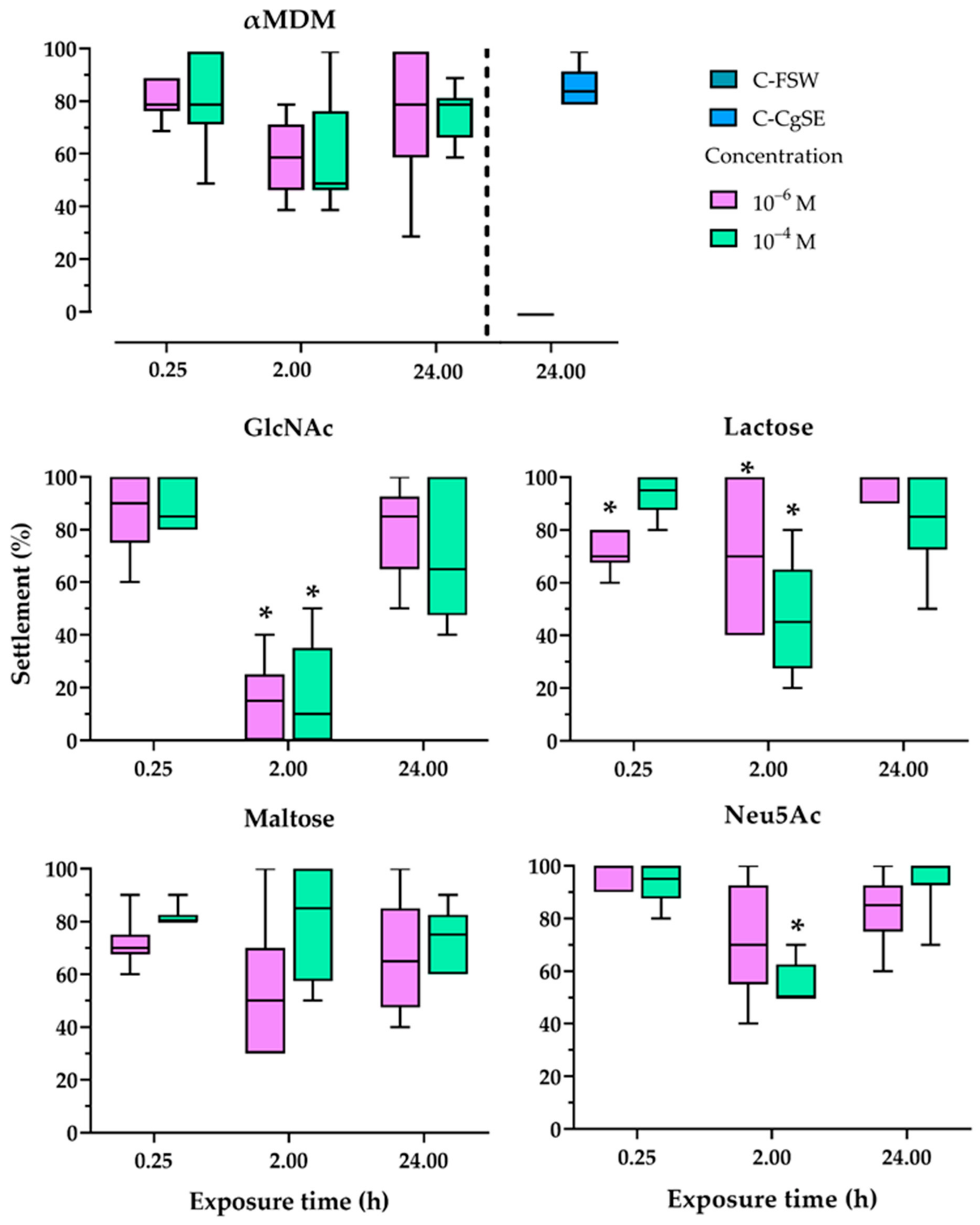
2.3. Effect of Exposure Time to Sugars and CgSE on Oyster Larval Settlement Response
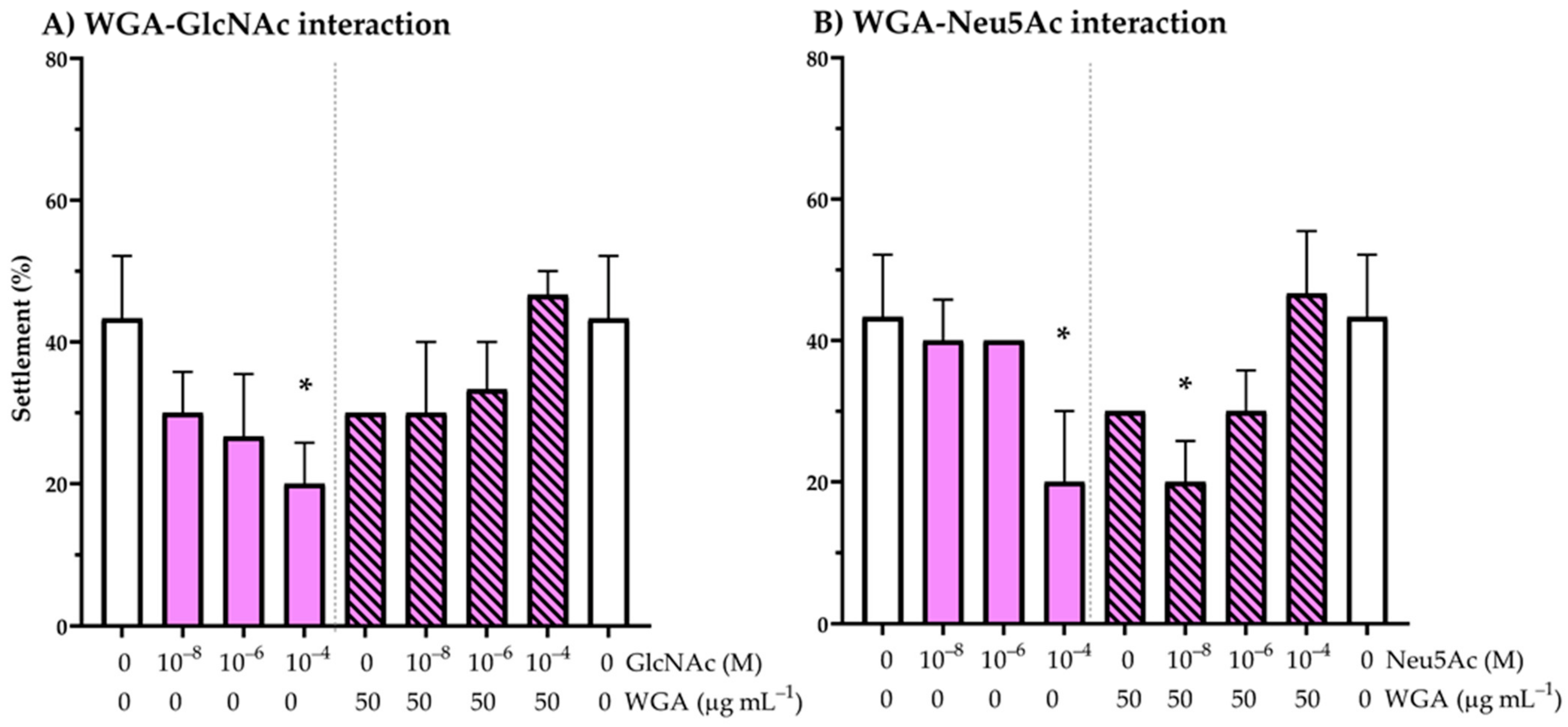
2.4. WGA Lectin Interaction with GlcNAc and Neu5Ac Binding: Effect on Oyster Larval Settlement
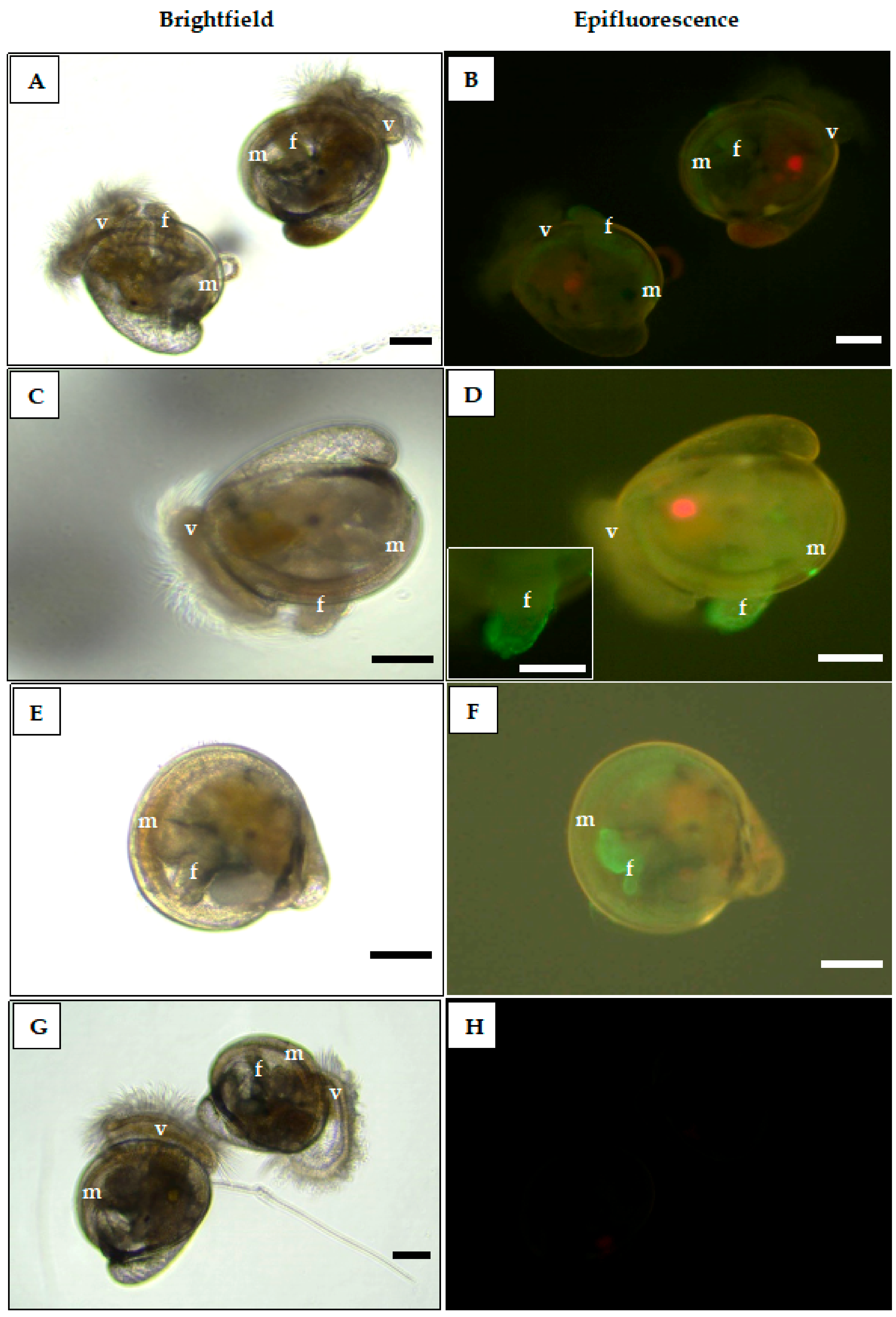
2.5. WGA-Binding Distribution on C. gigas Oyster Larval Tissues
3. Discussion
4. Materials and Methods
4.1. Spawning and Larval Culture of C. gigas
4.2. Shell Preparation and Matrix Extraction
4.3. Larval Substrate ‘Choice’ Settlement Assay
4.4. Treatment of Larvae with Sugars and Settlement Assay
4.5. Assay Protocol on the Effect of Exposure Time on Sugar and Conspecific Cues Induction of Settlement
4.6. Assay Protocol on WGA and Sugar (GlcNAc and Neu5Ac) Interaction
4.7. Treatment of Oyster Larvae with Fluorescein Isothiocyanate-Conjugated WGA
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hadfield, M.; Paul, V. Marine Chemical Ecology: Natural Chemical Cues for Settlement and Metamorphosis of Marine-Invertebrate Larvae; McClintock, J.B., Baker, B.J., Eds.; CRC Press: Boca Raton, FL, USA, 2001; ISBN 9780429124518. [Google Scholar]
- Prendergast, G.S. Settlement and Behaviour of Marine Fouling Organisms. Biofouling 2010, 30–59. [Google Scholar] [CrossRef]
- Hadfield, M.G. Biofilms and marine invertebrate larvae: What bacteria produce that larvae use to choose settlement sites. Ann. Rev. Mar. Sci. 2011, 3, 453–470. [Google Scholar] [CrossRef]
- Cranfield, H.J. Observations on the behaviour of the pediveliger of Ostrea edulis during attachment and cementing. Mar. Biol. 1973, 22, 203–209. [Google Scholar] [CrossRef]
- Coon, S.L.; Bonar, D.B.; Weiner, R.M. Induction of settlement and metamorphosis of the pacific oyster, Crassostrea gigas (Thunberg), by L-DOPA and catecholamines. J. Exp. Mar. Bio. Ecol. 1985, 94, 211–221. [Google Scholar] [CrossRef]
- Coon, S.L.; Walch, M.; Fitt, W.K.; Weiner, R.M.; Bonar, D.B. Ammonia Induces Settlement Behavior in Oyster Larvae. Biol. Bull. 1990, 179, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Joyce, A.; Vogeler, S. Molluscan bivalve settlement and metamorphosis: Neuroendocrine inducers and morphogenetic responses. Aquaculture 2018, 487, 64–82. [Google Scholar] [CrossRef]
- Vasquez, H.E.; Hashimoto, K.; Yoshida, A.; Hara, K.; Imai, C.C.; Kitamura, H.; Satuito, C.G. A glycoprotein in shells of conspecifics induces larval settlement of the Pacific oyster Crassostrea gigas. PLoS ONE 2013, 8, e82358. [Google Scholar] [CrossRef] [Green Version]
- Vasquez, H.E.; Hashimoto, K.; Kitamura, H.; Satuito, C.G. Wheat germ agglutinin-binding glycoprotein extract from shells of conspecifics induces settlement of larvae of the pacific oyster Crassostrea gigas (Thunberg). J. Shellfish Res. 2014, 33, 415–423. [Google Scholar] [CrossRef]
- Crisp, D.J. Chemical Factors Inducing Settlement in Crassostrea virginica (Gmelin). J. Anim. Ecol. 1967, 36, 329–335. [Google Scholar] [CrossRef]
- Poirier, L.A.; Clements, J.C.; Davidson, J.D.P.; Miron, G.; Davidson, J.; Comeau, L.A. Sink before you settle: Settlement behaviour of Eastern oyster (Crassostrea virginica) larvae on artificial spat collectors and natural substrate. Aquac. Rep. 2019, 13. [Google Scholar] [CrossRef]
- Tamburri, M.N.; Luckenbach, M.W.; Breitburg, D.L.; Bonniwell, S.M. Settlement of Crassostrea ariakensis larvae: Effects of substrate, biofilms, sediment and adult chemical cues. J. Shellfish Res. 2008, 27, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Bayne, B.L. The gregarious behaviour of the larvae of Ostrea edulis L. at settlement. J. Mar. Biol. Assoc. UK 1969, 49, 327–356. [Google Scholar] [CrossRef]
- Hirata, Y.; Tamura, Y.; Nagasawa, K. Influence of presoaking conditions of spat collectors in seawater containing adult pacific oysters (Crassostrea gigas) on their larval settlement. Nippon Suisan Gakkaishi 2008, 74, 1017–1023. [Google Scholar] [CrossRef]
- Zimmer-faust, R.K.; Tamburri, M. Chemical identity and ecological implications of a waterborne, larval settlement cue. Limnol. Oceanogr. 1994, 39, 1075–1087. [Google Scholar] [CrossRef]
- Turner, E.J.; Zimmer-Faust, R.K.; Palmer, M.A.; Luckenbach, M.; Pentchef, N.D. Settlement of oyster (Crassostrea virginica) larvae: Effects of water flow and a water-soluble chemical cue. Limnol. Oceanogr. 1994, 39, 1579–1593. [Google Scholar] [CrossRef] [Green Version]
- Carroll, J.M.; Riddle, K.; Woods, K.E.; Finelli, C.M. Recruitment of the eastern oyster, Crassostrea virginica, in response to settlement cues and predation in North Carolina. J. Exp. Mar. Biol. Ecol. 2015, 463, 1–7. [Google Scholar] [CrossRef]
- Pascual, M.S.; Zampatti, E.A. Evidence of a chemically mediated adult-larval interaction triggering settlement in Ostrea puelchana: Applications in hatchery production. Aquaculture 1995, 133, 33–44. [Google Scholar] [CrossRef]
- Veitch, F.P.; Hidu, H. Gregarious setting in the American oyster Crassostrea virginica Gmelin: I. Properties of a partially purified “Setting factor”. Chesap. Sci 1971, 12, 173–178. [Google Scholar] [CrossRef]
- Fitt, W.K.; Labare, M.P.; Fuqua, W.C.; Walch, M.; Coon, S.L.; Bonar, D.B.; Colwell, R.R.; Weiner, R.M. Factors influencing bacterial production of inducers of settlement behavior of larvae of the oyster Crassostrea Gigas. Microb. Ecol. 1989, 17, 287–298. [Google Scholar] [CrossRef]
- Fitt, W.K.; Coon, S.L.; Walch, M.; Weiner, R.M.; Colwell, R.R.; Bonar, D.B. Settlement behavior and metamorphosis of oyster larvae (Crassostrea gigas) in response to bacterial supernatants. Mar. Biol. 1990, 106, 389–394. [Google Scholar] [CrossRef]
- Tamburri, M.N.; Zimmer-Faust, R.K.; Tamplin, M.L. Natural Sources and Properties of Chemical Inducers Mediating Settlement of Oyster Larvae: A Re-examination. Biol. Bull. 1992, 183, 327–338. [Google Scholar] [CrossRef]
- Hashimoto, K.; Vasquez, H.E.; Kitamura, H.; Satuito, C.G. Larval settlement of the Pacific Oyster Crassostrea gigas in response to marine microbial films and monospecies bacterial films. Sess. Org. 2017, 34, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.E. The role of extracellular polysaccharides in biofilms. J. Biotechnol. 1989, 10, 181–202. [Google Scholar] [CrossRef]
- Neu, T.R.; Marshall, K.C. Bacterial Polymers: Physicochemical Aspects of Their Interactions at Interfaces. J. Biomater. Appl. 1990, 5, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Weiner, R.; Langille, S.; Quintero, E. Structure, function and immunochemistry of bacterial exopolysaccharides. J. Ind. Microbiol. 1995, 15, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Maki, J.S.; Mitchell, R. Involvement of Lectins in the Settlement and Metamorphosis of Marine Invertebrate Larvae. Bull. Mar. Sci. 1985, 37, 675–683. [Google Scholar]
- Sharon, N.; Lis, H. Lectins; Springer: Dordrecht, The Netherlands, 2007; ISBN 978-1-4020-6605-4. [Google Scholar]
- Neal, A.L.; Yule, A.B. The link between cypris temporary adhesion and settlement of Balanus balanoides (L). Biofouling 1992, 6, 33–38. [Google Scholar] [CrossRef]
- Khandeparker, L.; Anil, A.C.; Raghukumar, S. Exploration and metamorphosis in Balanus amphitrite Darwin (Cirripedia; Thoracica) cyprids: Significance of sugars and adult extract. J. Exp. Mar. Biol. Ecol. 2002, 281, 77–88. [Google Scholar] [CrossRef]
- Khandeparker, L.; Anil, A.C. Role of conspecific cues and sugars in the settlement of cyprids of the barnacle, Balanus amphitrite. J. Zool. 2011, 284, 206–214. [Google Scholar] [CrossRef]
- Finelli, C.M.; Wethey, D.S. Behavior of oyster (Crassostrea virginica) larvae in flume boundary layer flows. Mar. Biol. 2003, 143, 703–711. [Google Scholar] [CrossRef]
- Kirchman, D.; Graham, S.; Reish, D.; Mitchell, R. Lectins may mediate in the settlement and metamorphosis of Janua (Dexiospira) brasiliensis Grube (Polychaeta: Spirorbidae). Mar. Biol. Lett. 1982, 3, 131. [Google Scholar]
- Matsumura, K.; Nagano, M.; Fusetani, N. Purification of a larval settlement-inducing protein complex (SIPC) of the Barnacle, Balanus amphitrite. J. Exp. Zool. 1998, 281, 12–20. [Google Scholar] [CrossRef]
- Matsumura, K.; Mori, S.; Nagano, M.; Fusetani, N. Lentil lectin inhibits adult extract-induced settlement of the barnacle, Balanus amphitrite. J. Exp. Zool. 1998, 280, 213–219. [Google Scholar] [CrossRef]
- Mitchell, R. Colonization by Higher Organisms. In Microbial Adhesion and Aggregation; Springer: Berlin/Heidelberg, Germany, 1984. [Google Scholar]
- Tommasone, S.; Allabush, F.; Tagger, Y.K.; Norman, J.; Köpf, M.; Tucker, J.H.R.; Mendes, P.M. The challenges of glycan recognition with natural and artificial receptors. Chem. Soc. Rev. 2019, 48, 5488–5504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.E.; Drickamer, K.; Schnaar, R.L.; Etzler, M.E.; Varki, A. Discovery and Classification of Glycan-Binding Proteins. In Essentials of Glycobiology; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2015; ISBN 9780879697709. [Google Scholar]
- Lis, H. Lectins as Molecules and as Tools. Annu. Rev. Biochem. 1986, 55, 35–67. [Google Scholar] [CrossRef] [PubMed]
- Neal, A.L.; Yule, A.B. The effects of dissolved sugars upon the temporary adhesion of barnacle cyprids. J. Mar. Biol. Assoc. UK 1996, 76, 649–655. [Google Scholar] [CrossRef]
- Alberts, B.; Johnson, A.; Lewis, J.; Morgan, D.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell; W. W. Norton & Company: New York, NY, USA, 2017. [Google Scholar]
- Charifi, M.; Sow, M.; Ciret, P.; Benomar, S.; Massabuau, J.C. The sense of hearing in the Pacific oyster, Magallana gigas. PLoS ONE 2017, 12, e0185353. [Google Scholar] [CrossRef] [Green Version]
- Pales Espinosa, E.; Koller, A.; Allam, B. Proteomic characterization of mucosal secretions in the eastern oyster, Crassostrea virginica. J. Proteom. 2016, 132, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Foulon, V.; Boudry, P.; Artigaud, S.; Guérard, F.; Hellio, C. In silico analysis of Pacific oyster (Crassostrea gigas) transcriptome over developmental stages reveals candidate genes for larval settlement. Int. J. Mol. Sci. 2019, 20, 197. [Google Scholar] [CrossRef] [Green Version]
- Monsigny, M.; Sene, C.; Obrenovitch, A.; Roche, A.-C.; Delmotte, F.; Boschetti, E. Properties of Succinylated Wheat-Germ Agglutinin. Eur. J. Biochem. 1979, 98, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Petraccioli, A.; Maio, N.; Guarino, F.M.; Scillitani, G. Seasonal variation in glycoconjugates of the pedal glandular system of the rayed Mediterranean limpet, Patella caerulea (Gastropoda: Patellidae). Zoology 2013, 116, 186–196. [Google Scholar] [CrossRef]
- Matsumura, K.; Nagano, M.; Kato-Yoshinaga, Y.; Yamazaki, M.; Clare, A.S.; Fusetani, N. Immunological studies on the settlement-inducing protein complex (SIPC) of the barnacle Balanus amphitrite and its possible involvement in larva-larva interactions. Proc. R. Soc. B Biol. Sci. 1998, 265, 1825–1830. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, E.P.; Perrigault, M.; Ward, J.E.; Shumway, S.E.; Allam, B. Microalgal cell surface carbohydrates as recognition sites for particle sorting in suspension-feeding bivalves. Biol. Bull. 2010, 218, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foulon, V.; Artigaud, S.; Buscaglia, M.; Bernay, B.; Fabioux, C.; Petton, B.; Elies, P.; Boukerma, K.; Hellio, C.; Guérard, F.; et al. Proteinaceous secretion of bioadhesive produced during crawling and settlement of Crassostrea gigas larvae. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cranfield, H.J. A study of the morphology, ultrastructure, and histochemistry of the food of the pediveliger of Ostrea edulis. Mar. Biol. 1973, 22, 187–202. [Google Scholar] [CrossRef]
- Cranfield, H.J. Observations on the function of the glands of the foot of the pediveliger of Ostrea edulis during settlement. Mar. Biol. 1973, 22, 211–223. [Google Scholar] [CrossRef]
- Johnstone, M.B.; Ellis, S.; Mount, A.S. Visualization of shell matrix proteins in hemocytes and tissues of the eastern oyster, Crassostrea virginica. J. Exp. Zool. Part. B Mol. Dev. Evol. 2008, 310, 227–239. [Google Scholar] [CrossRef]
- Johnstone, M.B.; Wheeler, A.P.; Falwell, E.P.; Staton, M.E.; Saski, C.A.; Mount, A.S. Folian-cv1 is a member of a highly acidic phosphoprotein class derived from the foliated layer of the eastern oyster (Crassostrea virginica) shell and identified in hemocytes and mantle. Front. Mar. Sci. 2019, 6, 1–21. [Google Scholar] [CrossRef]
- Davies, M.S.; Hawkins, S.J. Mucus from Marine Molluscs. Adv. Mar. Biol. 1998, 34, 1–71. [Google Scholar]
- Jing, X.; Espinosa, E.P.; Perrigault, M.; Allam, B. Identification, molecular characterization and expression analysis of a mucosal C-type lectin in the eastern oyster, Crassostrea virginica. Fish. Shellfish Immunol. 2011, 30, 851–858. [Google Scholar] [CrossRef]
- Wang, X.; Li, L.; Zhu, Y.; Du, Y.; Song, X.; Chen, Y.; Huang, R.; Que, H.; Fang, X.; Zhang, G. Oyster Shell Proteins Originate from Multiple Organs and Their Probable Transport Pathway to the Shell Formation Front. PLoS ONE 2013, 8, e66522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cranfield, H.J. Observations on the morphology of the mantle folds of the pediveliger of Ostrea edulis L. and their function during settlement. J. Mar. Biol. Assoc. UK 1974, 54, 1–12. [Google Scholar] [CrossRef]
- Yamaguchi, K. Shell structure and behaviour related to cementation in oysters. Mar. Biol. 1994, 118, 89–100. [Google Scholar] [CrossRef]
- Liu, C.; Li, S.; Kong, J.; Liu, Y.; Wang, T.; Xie, L.; Zhang, R. In-depth proteomic analysis of shell matrix proteins of Pinctada fucata. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messenger, J.B.; Nixon, M.; Ryan, K.P. Magnesium Chloride as an anaesthetic for cephalopods. Comp. Biochem. Physiol. Part C Comp. 1985, 82, 203–205. [Google Scholar] [CrossRef]
- Draper, N.R.; Smith, H. Applied Regression Analysis, 3rd ed.; Wiley: Hoboken, NJ, USA, 1998; ISBN 9780471170822. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sedanza, M.G.; Kim, H.-J.; Seposo, X.; Yoshida, A.; Yamaguchi, K.; Satuito, C.G. Regulatory Role of Sugars on the Settlement Inducing Activity of a Conspecific Cue in Pacific Oyster Crassostrea gigas. Int. J. Mol. Sci. 2021, 22, 3273. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063273
Sedanza MG, Kim H-J, Seposo X, Yoshida A, Yamaguchi K, Satuito CG. Regulatory Role of Sugars on the Settlement Inducing Activity of a Conspecific Cue in Pacific Oyster Crassostrea gigas. International Journal of Molecular Sciences. 2021; 22(6):3273. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063273
Chicago/Turabian StyleSedanza, Mary Grace, Hee-Jin Kim, Xerxes Seposo, Asami Yoshida, Kenichi Yamaguchi, and Cyril Glenn Satuito. 2021. "Regulatory Role of Sugars on the Settlement Inducing Activity of a Conspecific Cue in Pacific Oyster Crassostrea gigas" International Journal of Molecular Sciences 22, no. 6: 3273. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063273