
Understanding Plant Social Networking System: Avoiding Deleterious Microbiota but Calling Beneficials


1
Biotechnology Research Institute, College of Natural Sciences, Chungbuk National University, Cheongju 28644, Korea
2
Molecular Phytobacteriology Laboratory, Infection Disease Research Center, KRIBB, Daejeon 34141, Korea
3
Biosystem and Bioengineering Program, University of Science and Technology (UST) KRIBB School, Daejeon 34141, Korea
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(7), 3319; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073319
Submission received: 21 January 2021
/
Revised: 9 March 2021
/
Accepted: 19 March 2021
/
Published: 24 March 2021
(This article belongs to the Special Issue Plant Microbe Interaction 4.0)
Abstract
:Plant association with microorganisms elicits dramatic effects on the local phytobiome and often causes systemic and transgenerational modulation on plant immunity against insect pests and microbial pathogens. Previously, we introduced the concept of the plant social networking system (pSNS) to highlight the active involvement of plants in the recruitment of potentially beneficial microbiota upon exposure to insects and pathogens. Microbial association stimulates the physiological responses of plants and induces the development of their immune mechanisms while interacting with multiple enemies. Thus, beneficial microbes serve as important mediators of interactions among multiple members of the multitrophic, microscopic and macroscopic communities. In this review, we classify the steps of pSNS such as elicitation, signaling, secreting root exudates, and plant protection; summarize, with evidence, how plants and beneficial microbes communicate with each other; and also discuss how the molecular mechanisms underlying this communication are induced in plants exposed to natural enemies. Collectively, the pSNS modulates robustness of plant physiology and immunity and promotes survival potential by helping plants to overcome the environmental and biological challenges.
1. Introduction
Phytobiome refers to the ecosystem within and surrounding a plant, which comprises diverse organisms including viruses, bacteria, archaea, fungi, oomycetes, other plants and even animals. Since their first appearance in the ecosystem, plants have evolved alongside a variety of beneficial organisms, pathogens and insects. In other words, being sessile organisms, plants have evolved strategies to survive under biotic- and abiotic stresses [1]. The physiology of aboveground plant organs such as leaf and stem differs from that of belowground structures such as the root system, and the difference between two types of plant structures facilitates interaction and communication between biotic stresses [2]. In response to external stimuli such as pathogen and insect attacks, plants have developed a systemic translocational signaling system, in addition to local modulation on immunity. Depending on the types of interactions between biotic communities, the biochemical and physiological fitness of plants has been modified synergistically, antagonistically or neutrally [1].
Interaction with a certain biotic stimulus stimulates the development of defense strategies at a local (infection site) area in plants [3]. Subsequently, defense signals derived from a local region of infected plants are transferred to systemic sites [3]. In addition to the intracellular signals and molecules, the plant-derived signals and molecules act as inter-compartment or inter-plant regulators [4]. To compromise the stress factors, plants orchestrate sophisticated machineries to cooperate with the soil microbiome [5,6,7]. In the rhizosphere, a number of microbes interact with plant roots, which can stimulate plant growth and immunity but also acquire some key nutrients in return [5,8]. Thus, plants and mutualistic microbes communicate with each other, and beneficial interactions between these partners can facilitate defense against invading enemies [9,10].
Multitrophic interactions between plants and microbes (i.e., detrimental insect–plant interactions and beneficial microbe–plant interactions) can be utilized to attenuate plant disease occurrence. For instance, in pepper (Capsicum annuum), foliar aphid feeding recruited rhizosphere bacteria and stimulated plant immunity against the leaf spot pathogen, Xanthomonas axonopodis [11]. In tobacco (Nicotiana benthamiana), whitefly infestation modulated plant immunity to produce endogenous salicylic acid (SA), resulting in the attenuation of Agrobacterium tumefaciens-induced gall formation [12]. More recently, activation of plant immunity in tomato (Solanum lycopersicum) by four Gram-positive bacteria controlled the incidence of bacterial wilt disease caused by Ralstonia solanacearum [13]. In addition, root-associated bacteria triggered the release of plant volatile organic compounds (VOCs), referred to as microbe-induced plant volatiles, and affected the rhizosphere microbiota of neighboring plants [14]. These results indicate that plants interact with beneficial microbes under certain stress conditions to control the response to third-party organisms. In this review, we describe the steps that constitute the plant social networking system (pSNS), show how plants and beneficial microbes communicate with each other, and highlight the strategies and underlying mechanisms of the pSNS. With field applications in mind, we also summarize the technological limitations of pSNS and how these could be surmounted.
2. The Plant SNS Hypothesis
Recently, the role of plant-associated microbial community has been reviewed in plant–plant communications as wired- and wireless components [15]. In current review, we have focused on the role of plant in modulating SNS against multitrophic stimuli. Therefore, the definition of plant SNS should be clarified first as compared to indirect defense and induced systemic resistance (ISR). Plant defense mechanisms can be classified into two categories: preformed defense and induced defense [16]. Induced defense is normally when they have turned off their defensive strategies to regulate the fitness cost of plants under natural conditions, but plant immunity can be induced by the perception of invading pathogens and insects [16]. Subsequently, during plant–insect interaction, plant defense mechanisms can be similarly divided into two categories: direct defense and indirect defense. In direct defense, plants produce toxic compounds to directly control the population of invading insects. However, in indirect defense, plants attract carnivores that feed on the invading insects, thus indirectly controlling the insect population (Figure 1A [17,18]). The feeding of insects on plant leaves activates the indirect defense of plants, resulting in the activation of elicitors (fatty-acid conjugates, enzymes, cell wall fragments, peptides and esters), plant hormones (SA, jasmonic acid [JA] and ethylene [ET]) and plant volatiles (terpenes, aldehydes, ketones, esters, alcohols and nitrogen compounds) (Figure 1A [19]). These serial events attract natural enemies (predators, parasitic wasps and omnivores), which suppress the insect population.
Induced systemic resistance (ISR) is a form of induced plant defense initiated by beneficial microbes (endophytes, mycorrhizal fungi and plant growth-promoting rhizobacteria) against invading pathogens and insects (Figure 1B). The majority of beneficial microbes are localized in the plant rhizosphere, and several of these microbes promote plant growth and stimulate biotic and abiotic stress resistance [20]. The beneficial microbe-triggered ISR in plants generally suppresses a variety of pathogens and insects [21,22,23].
The induced and indirect defense mechanisms have been thoroughly investigated in plants, both ecologically and biochemically. The most well-established area of research is the role of phytohormones in plant immune signaling. SA, JA and ET function as the main regulators of plant defense responses against pathogens and insects. SA-dependent pathways are activated by biotrophic pathogens and sucking insects, whereas JA- and ET-dependent pathways are induced by necrotrophic pathogens and chewing insects [24,25]. In addition, JA- and ET-related pathways show extensive crosstalk during plant defense responses [24]. In the ISR pathway, while increasing evidence suggests that JA and ET play a pivotal role in suppressing diseases and insects [23,26], more recent studies show that antagonism between SA and JA/ET has been broken down in some cases [27,28].
Here, we propose the plant SNS hypothesis, which is unlike plant indirect defense and ISR (Figure 1C). When plants are attacked by insects or infected by pathogens, certain signals or molecules released by aboveground tissues are transmitted to the rhizosphere, resulting in recruitment of beneficial microbes. These microbes then activate plant defense responses against spatially separated and systemically localized pathogens and insects. The major difference of plant SNS compared with ISR is plant self-modulation against enemy’s attacks through recruiting beneficial microbe-mediated systemic signaling. Indirect defense is not stimulated against plant microbial pathogens, but it can be operated against insect pests. Hereafter, we focus on how plants build the SNS in a step-by-step manner, including the elicitation in a local area, activation and transduction of systemic signals, secretion of bioactive root exudates and chemicals into the rhizosphere, the establishment of a favorable environment (by recruiting beneficial microbes and avoiding plant pathogens), and molecular mechanisms underlying the plant SNS.
3. Building Blocks and Molecular Mechanisms of the Plant SNS
A stepwise procedure for building the plant SNS (Figure 2) is described below. Here, we describe the steps that constitute the plant social networking system (pSNS); summarize, with evidence, how plants and beneficial microbes communicate with each other in each step; and finally, highlight the strategies and underlying mechanisms of the pSNS.
3.1. Elicitation: Plant Incuction in a Local Area by Insect and Pathogen Attacks
Plants are constantly exposed to diverse pathogens and insects, and unlike animals, plants have developed effective immune strategies to defend themselves against these biotic stressors by recognizing common and species-specific determinants on pathogens and insects. Interestingly, plants have developed an innate immune response that resembles the animal immune system. Plants can detect common foreign signals such as pathogen/microbe/herbivore-associated molecular patterns (PAMPs/MAMPs/HAMPs) via pattern-recognizing receptors (PRRs) [29,30]. The well-characterized plant PAMPs include flg22 (the N-terminus of bacterial flagellin), elf18/elf26 (the N-terminus of bacterial elongation factor Tu), peptidoglycans (a component of bacterial cell walls), and chitin (a component of fungal cell walls) [29,30,31,32,33]. In addition, plants can also perceive herbivore-derived precursors to form fatty acid–amino acid conjugates as HAMPs including volicitin, inceptins, caeliferin and bruchin [34,35,36,37].
The perception of MAMPs and HAMPs by plant cells induces an influx of extracellular calcium ions (Ca2+), production of reactive oxygen species (ROS) [38,39] and activation of mitogen-activated protein kinase (MAPK) cascades that play an important role in the regulation of downstream signaling [40,41,42]. Phytohormone signaling pathways represent a critical component of plant defense responses against pathogens and insects, as discussed above. Among various plant hormones, SA, JA and ET are the key regulators of plant defense [24,25]. These plant hormones are also utilized as signaling molecules to activate plant systemic immune responses.
3.2. Signaling: Activation and Transduction of Systemic Signaling Molecules
After recognizing a microbial pathogen or an insect pest’s determinants, plants generate signal molecules that translocate from the attack site (local site) to systemic organs. Here we summarize the signaling molecules that play an important role in this system. The signaling molecules include SA, MeSA, JA, JA derivatives, volatile organic compounds, and lipid-derived molecules.
3.2.1. SA and Methyl Salicylate
SA and its volatile derivative, methyl salicylate (MeSA), are strong systemic signal candidates. The first reports strongly support that SA is normally required for activating systemic resistance in tobacco and cucumber (Cucumis sativus) [43,44,45]. Because the first several reports supported the potential role of SA as a systemic signal, subsequent studies revealed its role in systemic signal transduction using mutant lines [46,47,48,49,50,51]. Further investigation showed that glycosylated SA compounds, such as SA-glycosyd (SAG), and MeSA are accumulated by plant cells upon pathogen infection [52,53,54]. An early study showed that MeSA is transmitted from Tobacco mosaic virus (TMV)-infected donor (emitter) plants to healthy (receiver) plants, resulting in a 30% decrease in TMV lesion size [55]. Several grafting experiments revealed that SA is converted into MeSA in infected leaves, which is subsequently translocated to distal systemic leaves, where it is converted back to SA to activate systemic resistance against biotrophic pathogens and sucking insects [56,57,58,59,60,61]. SA-dependent systemic signaling is mainly activated by sucking (phloem-feeding) aphids and spider mites [62].
3.2.2. JA and Its Derivatives
Unlike SA, JA and its derivatives, methyl jasmonate (MeJA) and jasmonoyl isoleucine (JA-Ile), function as signaling molecules in plant defense against necrotrophic pathogens and chewing insects [24,63,64]. JA is synthesized in vascular bundles, which also host prosystemin biosynthesis, and JA, systemin and prosystemin are related to each other in a double feedback manner [65,66]. Upon the attack of insect herbivores, polyunsaturated fatty acids (PUFAs) are released from the plant membrane lipids by lipases such as defective in anther dehiscence 1 (DAD1) and dongle (DGL), while PUFAs such as linolenic acid (C18:3) and linoleic acid (C18:2) are oxygenated by lipoxygenases (LOXs), which catalyze the synthesis of a large variety of oxylipins, including JA and green leafy volatiles (GLVs), through 6–7 independent pathways [67,68,69,70]. In addition to JA, conjugates of JA also act as systemic signals [71]. For example, JA-Ile acts as an active signal for the regulation of defense responses rather than JA itself [72,73], and MeJA is critically involved in diverse interactions as a key signal [74].
3.2.3. Gaseous Signals VOCs
Besides the volatile forms of SA and JA, other VOCs are also released by plants in response to attack by a variety of insect herbivores [75,76,77,78]. More than 1000 VOCs have been identified in plant tissues including flowers, and vegetative organs [79,80]. Plant-derived VOCs are the main signals involved in multitrophic interactions. Among these VOCs, the main volatiles (C6-alcohols and C6-aldehydes) are derived from GLVs by the action of hydroperoxide lyase (HPL) through the octadecanoic-derived LOX pathway, and some of the GLVs are required for direct defense, which explains why the deletion of the HPL gene in transgenic potato (Solanum tuberosum) decreased the resistance to aphids [81]. Cis-3-hexen-1-ol is emitted upon herbivore attack to attract a generalist predator under field conditions [76]. Trans-2-hexenal is also released upon herbivore-induced wounding, and its emission influences the accumulation of sesquiterpenes in wounded Arabidopsis thaliana and cotton (Gossypium hirsutum) plants, suggesting that GLVs are involved in intra- and inter-plant defense signaling pathways in response to diverse insects [82,83]. VOCs emitted upon insect infestation are referred to as herbivore-induced plant volatiles (HIPVs). Other groups of VOCs, including monoterpenes (limonene, linalool and ocimene), sesquiterpenes (bergamotene, carphyllene and farnesene), MeSA and MeJA, are released from the wound site within 24 h of herbivore attack [79,80,84,85].
3.2.4. Lipid-Derived Signals
Unlike the plant VOCs induced by herbivore attack, plants generate lipid-based signal molecules that translocate from one tissue to another. The defective in induced resistance 1 (DIR1) gene encodes a 7 kDa lipid transfer protein, which is localized in petiole exudates of pathogen-infected leaves either as an oligomer or in a complex with other proteins [86,87]. DIR1 interacts with other lipid-derived signaling compounds such as azelaic acid (AzA), dehydroabietic acid (DA), glycerol-3-phosphate (G3P) and MeSA [86,88,89,90].
G3P functions as an important signaling molecule in systemic resistance, and localizes to the cytosol and chloroplast. Several mutant-based studies have proven the function of G3P as a systemic signal. In Arabidopsis, disruption of the suppressor of fatty acid deficiency 1 (SFD1) gene, which encodes dihydroxy acetone phosphate (DHAP), elucidated the role of G3P as a systemic signal [91]. The sfd1 mutant showed low levels of SA accumulation and pathogenesis-related 1 (PR1) expression in distal tissues [91,92]. In other studies, mutation of SFD2, fatty acid desaturase 7 (FAD7), monogalactosyl synthase 1 (MGD1) and glycerol insensitive 1/nonhost 1 (GLI1/NHO1) abolished systemic responses [88,93].
AzA, a nine-carbon dicarboxylic acid, is the final product of lipid peroxidation under biotic stress [94]. The notion that AzA functions as a signaling molecule in systemic resistance was first verified by Jung and colleagues [89]; the authors showed that AzA acts as a priming molecule, and SA levels are elevated in distal leaves after infection by Pseudomonas syringae pv. maculicola strain PmaDG3 or upon the application of exogenous AzA, loss of systemic resistance of AZI1 induced by AzA is observed by gene disruption analysis [89]. In contrast to the results of Jung and colleagues [89], other studies could not elucidate the priming effect of AzA on SA accumulation in systemic leaves [95]. The possible routes of AzA biosynthesis in plants under biotic stress conditions are complex and controversial, indicating that the bacteria-derived nine-carbon product (i.e., AzA) of plants is potentially synthesized by non-enzymatic pathways, although LOXs are mainly required for the accumulation of lipid derivatives [94]. Recent studies show that galactolipids produce AzA via the ROS-mediated pathway [94,96]. The aboveground elicitation by pathogen and insect attacks leads to the activation of plant signaling transduction pathways, resulting in the secretion of root exudates that modulate rhizosphere microbiota and plant immunity.
3.3. Secreting Root Exudates: Plant Secretion of Bioactive Root Exudates and Chemicals into the Rhizosphere
The chemical signals of plants infected with pathogens and insects can be translocated to the root and affect compositions and contents of root exudates in the rhizosphere. The concept of plant–microbe interactions in the rhizosphere is not novel; however, the exact roles and composition of plant-derived root exudates remain unclear. Rhizodeposits released into the rhizosphere by plant roots include root exudates, mucilage, lysates released from wound sites and senescing cells that contain mono-, polysaccharides, organic acids, phenolic compounds, amino acids, and proteins [97,98]. Root exudates generated from the meristematic zone of root tips are the major components of rhizodeposits, and contain sugars, amino acids, organic acids, phenolics, alcohols, polypeptides and proteins [97,99]. Chemical compounds in root exudates play a pivotal role in acquiring mineral nutrients, attracting beneficial microbes and suppressing deleterious pathogens in the rhizosphere [5,100,101,102,103]. Here, we highlight the diverse compounds in root exudates released in response to nutrient limitations, pathogen infection and insect infestation, thus attracting microbes and other biotic stimuli.
3.3.1. Secretion of Strigolactones (SLs), Flavonoids and Coumarins under Nutrient Limitation Conditions
Under nutrient-deficient conditions, plants secrete large amounts of SLs, flavonoids and coumarins into the rhizosphere. The recently discovered plant hormone, SL, is synthesized from a carotenoid precursor and is secreted by plants under phosphate-deficient conditions to recruit arbuscular mycorrhizal (AM) fungi, resulting in phosphate uptake [104,105]. SLs affect the interactions between the host plant and AM fungi; abolishing SL biosynthesis decreases the colonization efficiency of Gigaspora rosea [106]. Flavonoids are a group of secondary metabolites secreted into the rhizosphere under phosphate- and nitrogen-limiting conditions. Flavonoids participate in the interaction between legumes and Rhizobium spp. and between actinorhizal plants and Frankia spp.; plants utilize nitrogen fixed by the bacteria, and in turn bacteria obtain carbon sources from the plant [107]. Coumarins are low-molecular-weight (LMW) secondary metabolites similar to flavonoids and are involved in plant–pathogen interactions as antimicrobial compounds [108]. However, recent studies revealed the function of coumarins as components of root exudates released by plants under nutrient-deficient conditions. Arabidopsis roots secrete diverse coumarins, including scopoletin, esculetin, fraxetin and sideretin, under iron limitation conditions. Additionally, genetic analyses revealed that Arabidopsis mutant lines grown in iron-deficient soils lack the ability to secrete or synthesize coumarins [109,110,111,112]. Roots of the annual grass Avena barbata secrete tryptophan and sucrose into the rhizosphere, and the tryptophan residue located close to the lateral roots potentially interacts with indole-3-acetic acid (IAA) to modulate lateral root initiation [113].
3.3.2. Secretion of Malic Acid and Phenolic/Organic Acid Compounds upon Pathogen Infection
The infection of Arabidopsis leaves by P. syringae pv. tomato (Pto) DC3000 facilitates the secretion of malic acid into the rhizosphere, which attracts Bacillus subtilis FB17 [5]. In addition, Pto infection increases the secretion of long-chain organic acids (pentadecanoic acid, hexadecanoic acid, palmitoleic acid, octadecanoic acid and arachidic acid) and amino acids (isoleucine, leucine, methionine, proline, tryptophan and ornithine) [114]. Infection by soil-borne pathogens modifies the root exudates of host plants and alters the microbial composition of the rhizosphere [20,115]. Infection of soybean (Glycine max) roots by Pythium ultimum increased the concentrations of phenolic and organic acids such as vanillic acid, p-coumaric acid and fumaric acid by 4-fold [116]. Additionally, antimicrobial compounds such as caffeic acid ester and rosmarinic acid are released by sweet basil (Ocimum basilicum) in response to infection by Pythium spp. [117]. Fusarium spp. induce the secretion of antifungal phenolics from barley (Hordeum vulgare) roots [118]. Infection of tobacco roots by the root-knot nematode Meloidogyne incognita leads to the accumulation of the defense-related compound nicotine in aboveground tissues, which is effectively utilized to attenuate foliar herbivores [119]. In potato, the powdery scab pathogen Spongospora subterranea facilitates the secretion of root exudates containing 24 different kinds of LMW compounds such as amino acids, sugars and organic acids, among others [120]. In tobacco, infection by bacterial wilt and black shank pathogens increases the secretion of amines, alcohols, lipids, sugars and esters in root exudates, and these compounds modulate the pathogen-antagonizing microbes [121].
3.3.3. Secretion of Benzoxazinoids and SA upon Insect Infestation
Insect infestation of aboveground and belowground plant tissues stimulates the secretion of root exudates into the rhizosphere to recruit microbes or suppress disease. Maize (Zea mays) roots secrete benzoxazinoids (BXs) into the rhizosphere [122,123]. However, the role of BXs in the context of exudates and their effect on soil microbial composition has not yet been investigated. Infestation of maize leaves by Spodoptera frugiperda induces the secretion of BXs into the rhizosphere, which affects the soil microbiota [124]. Insect infestation also leads to the production of certain volatiles in root exudates. Western corn rootworm feeding induces the accumulation of (E)-β-caryophyllene in belowground tissues, which attracts entomopathogenic nematodes [125]. In Citrus spp., four terpene volatiles were detected in root exudates after infestation by root weevil (Diaprepes abbreviatus) [126].
Whitefly infestation of aboveground tissues elicits the SA-dependent pathway in belowground tissues to suppress the soil-borne pathogen R. solanacearum in pepper [127] and A. tumefaciens-induced crown gall formation in tobacco [12]. Similarly, the attack of aboveground tissues by aphids modulates defense responses in belowground tissues via SA- and JA-dependent pathways to control the population of foliar bacterial pathogens and soil-borne pathogens [11]. Moreover, in potato, aphid (Myzus persicae) infestation of aboveground tissues induces the secretion of root exudates into the rhizosphere, showing that root exudates from aphid feeding on aboveground into the belowground and Neprilysin-1 of cyst nematode (Globodera pallida) are highly linked [128]. Moth-induced defoliation of the aboveground plant parts of the subarctic mountain birch (Betula pubescens) results in the release of carbon-rich compounds into the rhizosphere to modulate ectomycorrhizal fungi [129].
3.4. Plant Protection
Since root exudates containing diverse compounds and molecules are secreted into the rhizosphere, they can potentially generate signals that increase plant protection. In this part, we introduce three scenarios: recruitment of beneficial microbes by root exudates, antibiosis and antimicrobial compounds, and induced systemic resistance (ISR).
3.4.1. Recruitment of Beneficial Microbes by Root Exudates
One of the main functions of plant root exudates is the recruitment of beneficial microbes, which will protect plants under biotic and abiotic stress conditions. The composition of root exudates generally varies with soil nutrient status, disease incidence and abiotic stresses [130], and affects the soil microbial composition [99,131]. Studies show that biotic stresses can cause the secretion of chemicals into root exudates, thus attracting other microbes. For example, Fusarium oxysporum-infected tomato plants recruit Proteobacteria, Actinobacteria and Firmicutes [132], and Botrytis cinerea infection induces the accumulation of Trichoderma harzianum in the rhizosphere of tomato and cucumber plants [133]. In Arabidopsis, infection of leaves by the downy mildew pathogen Hyaloperonospora arabidopsidis leads to the recruitment of beneficial microbes such as genus of Xanthomonas, Stenotrophomonas, and Microbacterium [5,134,135,136], while infection by Pto DC3000 facilitates the attraction of B. subtilis FB17 [5,136] and leads to the assembly of a beneficial rhizosphere microbiome [114]. In sugar beet (Beta vulgaris), infection by Rhizoctonia solani alters the microbiome composition and attracts bacteria belonging to the families Oxalobacteraceae, Burkholderiaceae, Sphingobacteriaceae and Sphigomonadaceae [137].
In addition to pathogen infection, insect infestation has also been shown to attract beneficial microbes. In pepper, aboveground aphid feeding recruits beneficial microbes in the rhizosphere; thus, the population density of Bacillus subtilis GB03 was significantly higher in aphid feeding plants than in control plants, but the population density of Pseudomonas protegens Pf-5 was not affected by aphid feeding [11]. Similarly, whitefly infestation of pepper plants stimulates the recruitment of Gram-positive bacteria and fungi in the rhizosphere [127]. In maize, chewing by S. frugiperda alters the soil microbiota [124]. In Brassica napus, infestation of belowground tissues by cabbage root fly (Delia radicum) attracts four bacterial genera (Bacillus, Paenibacillus, Psedomonas and Stenotrophomonas) in the rhizosphere [138]. Root herbivory of bentgrass (Agrostis spp.) and clover (Trifolium spp.) plants by Tipula paludosa can utilize Pseudomonas compared with non-infested plants [139].
3.4.2. Antibiosis and Antimicrobial Compounds
Beneficial microbes recruited by the secretion of plant root exudates produce antibiotics, which can be utilized as biological control agents against harmful pathogens via a phenomenon known as antibiosis [140]. The major antibiotics produced by bacteria include hydrogen cyanide (HCN) [141], phenazine-1-carboxylic acid [142], phenazine-1-carboxyamide [142], 2,4-diacetyl phloroglucinol (Phl) [143], pyoluteorin [144] and pyrrolnitrin [145]. Phenazine-1-carboxylic acid produced by Pseudomonas fluorescens 2–79 in the wheat rhizosphere attenuates the disease-causing fungus Gaeumannomyces graminis var. tritici, and mutant analysis of phenazine-1-carboxylic acid showed that its mutation partially contributes to the alleviation of disease symptoms [146]. HCN, a volatile antibiotic produced by P. fluorescens CHA0, negatively regulates the fungal pathogen of black root rot, Thielaviopsis basicola, in tobacco [147].
In addition, lipopeptides such as fengycin, surfactin and iturin are LMW compounds produced by B. subtilis strains. These lipopeptides can directly suppress pathogenic fungi under pre- and post-harvest conditions [148]. Iturin A derived from B. subtilis strains PCL1608 and PCL1612 directly controls two fungal pathogens, Fusarium oxysporum and Rosellinia necatrix [149]. Surfactin produced by B. subtilis strain 6051 acts as a biocontrol agent in response to pathogenic bacteria [150].
Moreover, some studies have shown that root exudates secreted by plants exposed to pathogens exhibit direct antimicrobial activity. Two types of LMW antimicrobial compounds, phytoanticipins and phytoalexins, are involved in the direct suppression of pathogens [151,152]. The distinction between these two types of compounds is, however, difficult because pathogen infections sometimes induce the accumulation of phytoalexins as well as phytoanticipins [151]. Root exudates of barley plants infected by Fusarium graminearum contain aromatic compounds including five phenylpropanoids, such as t-cinnamic acid, p-coumaric acid, ferulic acid, syringic acid and vanillic acid, which exhibit direct antifungal activity and inhibit the germination of F. graminearum macroconidia [153]. In addition, scopoletin, a coumarin from plants, directly inhibits F. oxysporum and Verticillium dahliae [154]. Root exudates of pine tree (Pinus resinosa) associated with the ectomycorrhizal fungus Paxillus involutus contain ethanol-soluble compounds that act as antifungal molecules, suppressing the sporulation of F. oxysporum by 80% [155].
3.4.3. Induced Systemic Resistance
Beneficial microbes, including bacteria and fungi, can activate ISR, and its initiation signals translocate the whole plant to suppress the invading pathogens/insects (Figure 1B). Pseudomonas spp., Bacillus spp., Trichoderma spp. and mycorrhizae have been shown to enhance plant immunity [3,156,157,158]. Here, we describe how beneficial microbes induce the plant defense response against pathogens and insects in aboveground tissues. This phenomenon was first proven by some research groups in 1991: in carnation (Dianthus caryophyllus) plants, colonization by P. fluorescens WCS417r elicited resistance, which led to the accumulation of antimicrobial phytoalexins in response to the aboveground infection by the fungal pathogen F. oxysporum [159]; in cucumber plants, root colonization of Pseudomonas putida and Serratia marcescens 90–166 suppressed the symptoms of anthracnose caused by the fungal pathogen Colletotrichum orbiculare [160,161]. These two strains also stimulate ISR against several pathogens including F. oxysporum (Fusarium wilt pathogen; [162]), P. syringae (bacterial angular leaf spot pathogen; [163]), Cucumber mosaic cucumovirus (CMV) [164] and Erwinia tracheiphila (cucurbit wilt pathogen; [165]). The activation of ISR by rhizobacteria suppresses the disease caused by P. syringae pv. tabaci [166]. Some bacteria activate ISR in tobacco to suppress blue mold disease caused by Peronospora tabacina [167].
In addition to activating ISR against pathogens, mycorrhizal fungi also exert negative effects of herbivore performance [168,169]. In tomato, B. subtilis BEB-DN promotes the activation of ISR in aboveground tissues against Bemisia tabaci infestation [170], and pre-inoculation of tomato plants with B. subtilis triggers resistance against Bemisia tabaci under greenhouse conditions [171]. A field study showed that the population size of cucumber beetles is significantly decreased by several bacteria such as P. putida strain 89B-61, S. marcescens strain 90–166, Flavimonas oryzihabitans strain INR-5 and Bacillus pumilus strain INR-7 [172]. However, insect performance is positively regulated by P. fluorescens WCS417r in Arabidopsis [173]. The molecular mechanism of ISR has been well established in Arabidopsis, for example, using P. fluorescens WCS417r; systemic resistance triggered by P. fluorescens WCS417r activates the deposition of callose and up-regulates the expression of JA/ET-related genes, PDF1.2 and VSP2 [174,175,176]. These data suggest that JA- and ET-mediated pathways promote ISR, especially in Arabidopsis and tomato [177,178]. On the other hand, some studies rely on the activation of SA-dependent pathways triggered by certain rhizobacteria [179,180]. The summary of plant SNS steps is listed in Table 1.
3.5. Molecular Mechanisms Underlying the Plant SNS
In this part, the molecular mechanisms of plant SNS have been introduced in several recent studies. In pepper, whitefly infestation of aboveground plant parts elicits SA and JA signaling both in above- and below-ground tissues, and alters the microbiome assembly, leading to the attenuation of Xanthomonas axonopodis pv. vesicatoria (Xav) and R. solanacearum SL1931 [127]. Interestingly, although insect feeding does not cause any physiological changes in aboveground tissues, the root biomass is augmented, indicating that certain molecules and/or signals in aboveground organs are potentially transferred to belowground tissues [127]. Increase in root biomass has been consistently observed in several studies [12,181,182].
Whitefly infestation stimulates SA-mediated signaling attracting beneficial microbes, which would reduce the incidence of A. tumefaciens-induced gall formation in both above- and belowground tissues. Song and colleagues monitored the level of endogenous SA in whitefly-infested tobacco plants, and genetic analysis supported the phytohormone assays showing that the SA biosynthetic gene, ICS1, plays an important role in the accumulation of SA [12]. SA directly suppresses the expression of pathogenicity and virulence-related genes in Agrobacterium. Moreover, the content of IAA in whitefly-infested tobacco plants increased upon Agrobacterium inoculation [12], suggesting that IAA plays a pivotal role in the positive regulation of root biomass. This result was supported by the induction of auxin response genes and nutrient transporter genes in whitely infested roots [181].
Similar to whitefly infestation, aboveground aphid feeding elicits SA and JA signaling pathways, recruits B. subtilis GB03 and prevents the soil-borne bacterial pathogen R. solanacearum SL1931, thus priming pepper immunity in response to the pathogenic bacterium Xav and a non-pathogenic bacterium X. axonopodis pv. glycines (Xag). In a recent study, the severity level of disease caused by Xav was reduced by approximately 4-fold in aphid feeding plants compared with control plants, and the hypersensitive response index against Xag was significantly delayed in aphid feeding plants [11].
In addition to insect-induced plant SNS, pathogen-triggered plant SNS has been recently investigated. B. cinerea, a foliar fungal pathogen, secretes peroxidases and oxylipins as chemoattractant molecules into the roots of tomato plants for attracting Trichoderma harzianum T22 and inhibiting F. oxysporum [133]. Inoculation of Arabidopsis leaves with the pathogenic bacterium Pto DC3000 stimulates intra-plant long-distance signaling, activates the malate transporter AtALMT1 and leads to the secretion of malic acid. Malic acid in root exudates attracts B. subtilis FB17, which colonizes plant roots. These sequential signaling events enhance ISR to modulate defense responses against Pto DC3000 [136].
In sugar beet, inoculation of roots with the fungal pathogen R. solani induces the plants to respond to oxalic acid. Ribosomal RNA-based analyses revealed that R. solani infection significantly increased the population densities of Oxalobacteraceae, Burkholderiaceae, Sphingobacteriaceae and Sphingomonadaceae in the rhizosphere, and up-regulated the expression of genes involved in the regulation of bacterial stress responses, resulting in protection against R. solani [137].
4. Technological Limitations, Fundamental Issues, and Potential Troubleshooting Approaches
Although the proposed plant SNS represents a good strategy for stimulating plant growth and immunity, certain technical and experimental limitations and unanswered questions need to be addressed. Firstly, building multitrophic interactions is quite complicated and therefore must be adapted for very narrow studies. To investigate the interactions among host plants, microbes and pathogens/insects, a comprehensive analysis strategy is needed. This requires multidisciplinary omics-based tools in diverse fields, such as plant biochemistry, plant genetics, microbiology, genomics, transcriptomics, metabolomics, metagenomics and bioinformatics, for the elucidation of specific compounds in root exudates of pathogen-infected plants [114,124,183].
Secondly, to isolate and characterize composition of root exudates, artificially designed experimental procedures based on well-established systems are needed under laboratory and greenhouse conditions. For example, when whiteflies feed on plant leaves, the collection of root exudates from the rhizosphere soil is not easy. To fill the gap in the current situation, an in vitro bioassay has been alternatively invented [12,136]. Additionally, root exudate profiles under each condition need to be evaluated for connecting the plant phenotypes. Furthermore, after the isolation and characterization of compounds and molecules in root exudates, the functions of these compounds must be validated under natural conditions.
Thirdly, a single strain of a pathogen is sometimes not sufficient for manipulating the potential pathogens and insects. Accumulating evidence suggests that a certain bacterial strain, which has no effect on controlling plant defense responses, is effective when inoculated with other bacteria [5,184]. This is supported by the finding that inoculation of multiple strains is more effective in enhancing pathogen resistance than inoculation of a single strain [5,185,186]. Moreover, a recent study showed that microbial synthetic communities produce ISR-promoting substances that can be used as inoculants [186]. Diverse synthetic communities effectively suppress Fusarium wilt disease in tomato [186]. To extend the usage of synthetic community approaches, a variety of combinations of microbes have been evaluated for resistance to pathogens and insects. The elucidation of keystone taxa in complex synthetic microbial communities is very important for effective control of pathogens and insects.
Finally, the proposed plant SNS is essential for conclusively understanding the beneficial microbe-driven systemic resistance in plants exposed to pathogens and insects, as well as for the isolation and characterization of specific genes, traits, compounds/chemicals and microbial strains; however, whether studies on the plant SNS will generate reproducible results under field conditions remains unclear. For example, a single bacterial strain as well as synthetic communities of bacteria elicits negative effects on pathogen in vitro; however, the results may be different under field conditions [187]. Therefore, the reproducibility of plant SNS is a major issue for manipulating plant systemic defense responses against pathogens and insects. This suggests that plant SNS-derived ISR should be sustainably investigated and monitored in a variety of multitrophic interactions to improve agriculture in the future.
5. Future Perspectives
Plants serve as the major food source for humans, and an increase in plant yield is needed in the near future to meet the needs of the growing world population [188]. Plants are constantly exposed to diverse biotic and abiotic stresses, and therefore have developed sophisticated strategies for overcoming unfavorable conditions. Although many approaches have been used to maintain and produce more edible plants, increasing the plant yield remains a challenge because of natural disasters and invasion by insects and pathogens [189]. Approximately 10–16% of crop plants have been devastated by pathogens and insects, and the usage of chemicals has also increased substantially in the last few decades [190]. One of the proposed ecofriendly methods for increasing plant yield, despite the negative impact of pathogens and insects, is the plant SNS. To decrease the amount of chemical agents needed to control pathogens and insects, it is important to isolate and characterize certain genes, evaluate plant defense traits and beneficial microbe-related traits, and investigate the microbial community in a variety of plant species. If the plant SNS shows reproducible results in the field, this strategy will enhance our understanding of the ecological, economical and industrial aspects of agriculture.
Author Contributions
Conceptualization, C.-M.R.; writing—original draft preparation, Y.-S.P.; writing—review and editing, Y.-S.P. and C.-M.R.; visualization, Y.-S.P. and C.-M.R.; supervision, C.-M.R.; project administration, Y.-S.P. and C.-M.R.; funding acquisition Y.-S.P. and C.-M.R. Both authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the KRIBB initiative program, by the Strategic Initiative for Microbiomes in Agriculture and Food, Ministry of Agriculture, Food and Rural Affairs, Republic of Korea (as part of the multi-ministerial Genome Technology to Business Translation Program 918017-4), and by the National Research Foundation (NRF) of Korea funded by the Ministry of Education (NRF-2018R1D1A3B07049986 and 2020M3E9A1111636).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hauser, T.P.; Christensen, S.; Heimes, C.; Kiaer, L.P. Combined effects of arthropod herbivores and phytopathogens on plant performance. Funct. Ecol. 2013, 27, 623–632. [Google Scholar] [CrossRef]
- Soler, R.; Harvey, J.A.; Bezemer, T.M.; Stuefer, J.F. Plants as green phones: Novel insights into plant-mediated communication between below- and above-ground insects. Plant Signal. Behav. 2008, 3, 519–520. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Geem, M.; Gols, R.; van Dam, N.M.; van der Putten, W.H.; Fortuna, T.; Harvey, J.A. The importance of aboveground-belowground interactions on the evolution and maintenance of variation in plant defense traits. Front. Plant Sci. 2013, 4, 431. [Google Scholar]
- Berendsen, R.L.; Vismans, G.; Yu, K.; Song, Y.; de Jonge, R.; Burgman, W.P.; Burmølle, M.; Herschend, J.; Bakker, P.A.H.M.; Pieterse, C.M.J. Disease-induced assemblage of a plant-beneficial bacterial consortium. ISME J. 2018, 12, 1496–1507. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Hacquard, S.; Garrido-Oter, R.; González, A.; Spaepen, S.; Ackermann, G.; Lebeis, S.; McHardy, A.C.; Dangl, J.L.; Knight, R.; Ley, R.; et al. Microbiota and host nutrition across plant and animal kingdoms. Cell Host Microbe 2015, 17, 603–616. [Google Scholar] [CrossRef] [Green Version]
- Bakker, P.A.H.M.; Pieterse, C.M.J.; de Jonge, R.; Berendsen, R.L. The soil-borne legacy. Cell 2018, 172, 1178–1180. [Google Scholar] [CrossRef] [Green Version]
- Durán, P.; Thiergart, T.; Garrido-Oter, R.; Agler, M.; Kemen, E.; Schulze-Lefert, P.; Hacquard, S. Microbial interkingdom interactions in roots promote Arabidopsis survival. Cell 2018, 175, 973–983. [Google Scholar] [CrossRef]
- Lee, B.; Lee, S.; Ryu, C.M. Foliar aphid feeding recruits rhizosphere bacteria and primes plant immunity against pathogenic and non-pathogenic bacteria in pepper. Ann. Bot. 2012, 110, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Song, G.C.; Lee, S.; Hong, J.; Choi, H.K.; Hong, G.H.; Bae, D.W.; Mysore, K.S.; Park, Y.S.; Ryu, C.M. Aboveground insect infestation attenuates belowground Agrobacterium-mediated genetic transformation. New Phytol. 2015, 207, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kong, H.G.; Song, G.C.; Ryu, C.M. Disruption of Firmicutes and Actinobacteria abundance in tomato rhizosphere causes the incidence of bacterial wilt disease. ISME J. 2021, 15, 330–347. [Google Scholar] [CrossRef]
- Kong, H.G.; Song, G.C.; Sim, H.J.; Ryu, C.M. Achieving similar root microbiota composition in neighbouring plants through airborne signalling. ISME J. 2021, 15, 397–408. [Google Scholar] [CrossRef]
- Sharifi, R.; Ryu, C.M. Social networking in crop plants: Wired and wireless cross-plant communications. Plant Cell Environ. 2020. [Google Scholar] [CrossRef]
- Dicke, M.; van Poecke, R.M.P. Signalling in plant-insect interactions: Signal transduction in direct and indirect plant defence. In Plant Signal Transduction; Scheel, D., Wasternack, C., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 289–316. [Google Scholar]
- Hagenbucher, S.; Olson, D.M.; Ruberson, J.R.; Wäckers, F.L.; Romeis, J. Resistance mechanisms against arthropod herbivores in cotton and their interactions with natural enemies. Crit. Rev. Plant Sci. 2013, 32, 458–482. [Google Scholar] [CrossRef]
- Xu, S.; Zhou, W.; Pottinger, S.; Baldwin, I.T. Herbivore associated elicitor-induced defences are highly specific among closely related Nicotiana species. BMC Plant Biol. 2015, 15, 2. [Google Scholar] [CrossRef] [Green Version]
- Aljbory, Z.; Chen, M.S. Indirect plant defense against insect herbivores: A review. Insect Sci. 2018, 25, 2–23. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Pineda, A.; Zheng, S.J.; van Loon, J.J.; Pieterse, C.M.; Dicke, M. Helping plants to deal with insects: The role of beneficial soil-borne microbes. Trends Plant Sci. 2010, 15, 507–514. [Google Scholar] [CrossRef]
- Partida-Martínez, L.P.; Heil, M. The microbe-free plant: Fact or artifact? Front. Plant Sci. 2011, 2, 100. [Google Scholar] [CrossRef] [Green Version]
- Zamioudis, C.; Pieterse, C.M. Modulation of host immunity by beneficial microbes. Mol. Plant Microbe Interact. 2012, 25, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Ent, S.; Van Wees, S.C.; Pieterse, C.M. Jasmonate signaling in plant interactions with resistance-inducing beneficial microbes. Phytochemistry 2009, 70, 1581–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenk, S.T.; Schikora, A. AHL-priming functions via oxylipin and salicylic acid. Front. Plant Sci. 2014, 5, 784. [Google Scholar] [CrossRef]
- Vos, C.M.; De Cremer, K.; Cammue, B.P.; De Coninck, B. The toolbox of Trichoderma spp. in the biocontrol of Botrytis cinerea disease. Mol. Plant Pathol. 2015, 16, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Boller, T.; Felix, G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Steinbrenner, A.D.; Muñoz-Amatriaín, M.; Chaparro, A.F.; Aguilar-Venegas, J.M.; Lo, S.; Okuda, S.; Glauser, G.; Dongiovanni, J.; Shi, D.; Hall, M.; et al. A receptor-like protein mediates plant immune responses to herbivore-associated molecular patterns. Proc. Natl. Acad. Sci. USA 2020, 117, 31510–31518. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Gómez, L.; Boller, T. Flagellin perception: A paradigm for innate immunity. Trends Plant Sci. 2002, 7, 251–256. [Google Scholar] [CrossRef]
- Zipfel, C.; Kunze, G.; Chinchilla, D.; Caniard, A.; Jones, J.D.; Boller, T.; Felix, G. Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated transformation. Cell 2006, 125, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Alborn, H.T.; Hansen, T.V.; Jones, T.H.; Bennett, D.C.; Tumlinson, J.H.; Schmelz, E.A.; Teal, P.E.A. Disulfooxy fatty acids from the American bird grasshopper Schistocerca americana, elicitors of plant volatiles. Proc. Natl. Acad. Sci. USA 2007, 104, 12976–12981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doss, R.P.; Oliver, J.E.; Proebsting, W.M.; Potter, S.W.; Kuy, S.R.; Clement, S.L.; Williamson, R.T.; Carney, J.R.; DeVilbiss, E.D. Bruchins: Insect-derived plant regulators that stimulate neoplasm formation. Proc. Natl. Acad. Sci. USA 2000, 97, 6218–6223. [Google Scholar] [CrossRef] [Green Version]
- Schmelz, E.A.; LeClere, S.; Carroll, M.J.; Alborn, H.T.; Teal, P.E.A. Cowpea chloroplastic ATP synthase is the source of multiple plant defense elicitors during insect herbivory. Plant Physiol. 2007, 144, 793–805. [Google Scholar] [CrossRef] [Green Version]
- Truitt, C.L.; Wei, H.X.; Paré, P.W. A plasma membrane protein from Zea mays binds with the herbivore elicitor volicitin. Plant Cell 2004, 16, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Jeworutzki, E.; Roelfsema, M.R.; Anschütz, U.; Krol, E.; Elzenga, J.T.; Felix, G.; Boller, T.; Hedrich, R.; Becker, D. Early signaling through the Arabidopsis pattern recognition receptors FLS2 and EFR involves Ca-associated opening of plasma membrane anion channels. Plant J. 2010, 62, 367–378. [Google Scholar] [CrossRef]
- Ranf, S.; Eschen-Lippold, L.; Pecher, P.; Lee, J.; Scheel, D. Interplay between calcium signalling and early signalling elements during defence responses to microbe- or damage-associated molecular patterns. Plant J. 2011, 68, 100–113. [Google Scholar] [CrossRef]
- Colcombet, J.; Hirt, H. Arabidopsis MAPKs: A complex signalling network involved in multiple biological processes. Biochem. J. 2008, 413, 217–226. [Google Scholar] [CrossRef]
- Ryan, C.A. The systemin signaling pathway: Differential activation of plant defensive genes. Biochim. Biophys. Acta 2000, 1477, 112–121. [Google Scholar] [CrossRef]
- Seo, S.; Okamoto, M.; Seto, H.; Ishizuka, K.; Sano, H.; Ohashi, Y. Tobacco MAP kinase: A possible mediator in wound signal transduction pathways. Science 1995, 270, 1988–1992. [Google Scholar] [CrossRef]
- Gaffney, T.; Friedrich, L.; Vernooij, B.; Negrotto, D.; Nye, G.; Uknes, S.; Ward, E.; Kessmann, H.; Ryals, J. Requirement of salicylic acid for the induction of systemic acquired resistance. Science 1993, 261, 754–756. [Google Scholar] [CrossRef]
- Malamy, J.; Carr, J.P.; Klessig, D.F.; Raskin, I. Salicylic acid: A likely endogenous signal in the resistance response of tobacco to viral infection. Science 1990, 250, 1002–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Métraux, J.P.; Signer, H.; Ryals, J.; Ward, E.; Wyss-Benz, M.; Gaudin, J.; Raschdorf, K.; Schmid, E.; Blum, W.; Inverardi, B. Increase in salicylic acid at the onset of systemic acquired resistance in cucumber. Science 1990, 250, 1004–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaney, T.P.; Friedrich, L.; Ryals, J.A. Arabidopsis signal transduction mutant defective in chemically and biologically induced disease resistance. Proc. Natl. Acad. Sci. USA 1995, 92, 6602–6606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawrath, C.; Métraux, J.P. Salicylic acid induction-deficient mutants of Arabidopsis express PR-2 and PR-5 and accumulate high levels of camalexin after pathogen inoculation. Plant Cell 1999, 11, 1393–1404. [Google Scholar]
- Nawrath, C.; Heck, S.; Parinthawong, N.; Métraux, J.P. EDS5, an essential component of salicylic acid-dependent signaling for disease resistance in Arabidopsis, is a member of the MATE transporter family. Plant Cell 2002, 14, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Tsuda, K.; Truman, W.; Sato, M.; Nguyen, L.V.; Katagiri, F.; Glazebrook, J. CBP60g and SARD1 play partially redundant critical roles in salicylic acid signaling. Plant J. 2011, 67, 1029–1041. [Google Scholar] [CrossRef]
- Wildermuth, M.C.; Dewdney, J.; Wu, G.; Ausubel, F.M. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 2001, 414, 562–565. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, S.; Ding, P.; Wang, D.; Cheng, Y.T.; He, J.; Gao, M.; Xu, F.; Li, Y.; Zhu, Z.; et al. Control of salicylic acid synthesis and systemic acquired resistance by two members of a plant-specific family of transcription factors. Proc. Natl. Acad. Sci. USA 2010, 107, 18220–18225. [Google Scholar] [CrossRef] [Green Version]
- Attaran, E.; Zeier, T.E.; Griebel, T.; Zeier, J. Methyl salicylate production and jasmonate signaling are not essential for systemic acquired resistance in Arabidopsis. Plant Cell 2009, 21, 954–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.I.; Raskin, I. Purification, cloning, and expression of a pathogen inducible UDP-glucose:salicylic acid glucosyltransferase from tobacco. J. Biol. Chem. 1999, 274, 36637–36642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malamy, J.; Hennig, J.; Klessig, D.F. Temperature-dependent induction of salicylic acid and its conjugates during the resistance response to tobacco mosaic-virus infection. Plant Cell 1992, 4, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Shulaev, V.; Silverman, P.; Raskin, I. Airborne signaling by methyl salicylate in plant pathogen resistance. Nature 1997, 385, 718–721. [Google Scholar] [CrossRef]
- Dempsey, D.A.; Klessig, D.F. SOS-too many signals for systemic acquired resistance? Trends Plant Sci. 2012, 17, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Kaiyomo, E.; Kumar, D.; Mosher, S.L.; Klessig, D.F. Methyl salicylate is a critical mobile signal for plant systemic acquired resistance. Science 2007, 318, 113–116. [Google Scholar] [CrossRef]
- Park, S.W.; Liu, P.P.; Forouhar, F.; Vlot, A.C.; Tong, L.; Tietjen, K.; Klessig, D.F. Use of a synthetic salicylic acid analog to investigate the roles of methyl salicylate and its esterases in plant disease resistance. J. Biol. Chem. 2009, 284, 7307–7317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic Acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [Green Version]
- Van Butselaar, T.; Van den Ackerveken, G. Salicylic acid steers the growth-immunity tradeoff. Trends Plant Sci. 2020, 25, 566–576. [Google Scholar] [CrossRef]
- Chen, J.; Clinton, M.; Qi, G.; Wang, D.; Liu, F.; Fu, Z.Q. Reprogramming and remodeling: Transcriptional and epigenetic regulation of salicylic acid-mediated plant defense. J. Exp. Bot. 2020, 71, 5256–5268. [Google Scholar] [CrossRef]
- Arimura, G.; Matsui, K.; Takabayashi, J. Chemical and molecular ecology of herbivore-induced plant volatiles: Proximate factors and their ultimate functions. Plant Cell Physiol. 2009, 50, 911–923. [Google Scholar] [CrossRef] [Green Version]
- Koo, A.J.; Howe, G.A. The wound hormone jasmonate. Phytochemistry 2009, 70, 1571–1580. [Google Scholar] [CrossRef] [Green Version]
- Gorman, Z.; Christensen, S.A.; Yan, Y.; He, Y.; Borrego, E.; Kolomiets, M.V. Green leaf volatiles and jasmonic acid enhance susceptibility to anthracnose diseases caused by Colletotrichum graminicola in maize. Mol. Plant Pathol. 2020, 21, 702–715. [Google Scholar] [CrossRef] [Green Version]
- Hause, B.; Stenzel, I.; Miersch, O.; Maucher, H.; Kramell, R.; Ziegler, J.; Wasternack, C. Tissue-specific oxylipin signature of tomato flowers–allene oxide cyclase is highly expressed in distinct flower organs and vascular bundles. Plant J. 2000, 24, 113–126. [Google Scholar] [CrossRef]
- Jacinto, T.; McGurl, B.; Francheschi, V.; Delano-Freier, J.; Ryan, C.A. Tomato prosystemin promoter confers wound-inducible, vascular bundle-specific expression of the β-glucuronidase gene in transgenic tomato plants. Planta 1997, 203, 406–412. [Google Scholar] [CrossRef]
- Blée, E. Impact of phyto-oxylipins in plant defense. Trends Plant Sci. 2002, 7, 315–322. [Google Scholar] [CrossRef]
- Dave, A.; Graham, I.A. Oxylipin signaling: A distinct role for the jasmonic acid precursor cis-(+)-12-oxo-phytodienoic acid (cis-OPDA). Front. Plant Sci. 2012, 3, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feussner, I.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Narvaez-Vasquez, J.; Florin-Christensen, J.; Ryan, C.A. Positional specificity of a phospholipase-A activity induced by wounding, systemin, and oligosaccharide elicitors in tomato leaves. Plant Cell 1999, 11, 2249–2260. [Google Scholar] [CrossRef] [Green Version]
- Farmer, E.E. Plant biology: Jasmonate perception machines. Nature 2007, 448, 659–660. [Google Scholar] [CrossRef]
- Kang, J.H.; Wang, L.; Giri, A.; Baldwin, I.T. Silencing threonine deaminase and JAR4 in Nicotiana attenuata impairs jasmonic acid-isoleucine-mediated defenses against Manduca sexta. Plant Cell 2006, 18, 3303–3320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staswick, P.E.; Tiryaki, I. The oxylipin signal jasmonic acid is activated by an enzyme that conjugates it to isoleucine in Arabidopsis. Plant Cell 2004, 16, 2117–2127. [Google Scholar] [CrossRef] [Green Version]
- Cheong, J.J.; Choi, Y.D. Methyl jasmonate as a vital substance in plants. Trends Genet. 2003, 19, 409–413. [Google Scholar] [CrossRef]
- Dicke, M.; van Loon, J.J.A. Multitrophic effects of herbivore-induced plant volatiles in an evolutionary context. Entomol. Exp. Appl. 2000, 97, 237–249. [Google Scholar] [CrossRef] [Green Version]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef] [PubMed]
- Meiners, T.; Hilker, M. Induction of plant synomones by oviposition of a phytophagous insect. J. Chem. Ecol. 2000, 26, 221–232. [Google Scholar] [CrossRef]
- Li, T.; Holst, T.; Michelsen, A.; Rinnan, R. Amplification of plant volatile defence against insect herbivory in a warming Arctic tundra. Nat. Plants 2019, 5, 568–574. [Google Scholar] [CrossRef]
- Dudareva, N.; Cseke, L.; Blanc, V.M.; Pichersky, E. Evolution of floral scent in Clarkia: Novel patterns of S-linalool synthase gene expression in the C. breweri flower. Plant Cell 1996, 8, 1137–1148. [Google Scholar]
- Pichersky, E.; Noel, J.P.; Dudareva, N. Biosynthesis of plant volatiles: Nature’s diversity and ingenuity. Science 2006, 311, 808–811. [Google Scholar] [CrossRef] [Green Version]
- Vancanneyt, G.; Sanz, C.; Farmaki, T.; Paneque, M.; Ortego, F.; Castañera, P.; Sánchez-Serrano, J.J. Hydroperoxide lyase depletion in transgenic potato plants leads to an increase in aphid performance. Proc. Natl. Acad. Sci. USA 2001, 98, 8139–8144. [Google Scholar] [CrossRef] [Green Version]
- Bate, N.J.; Rothstein, S.J. C-6-volatiles derived from the lipoxygenase pathway induce a subset of defense-related genes. Plant J. 1998, 16, 561–569. [Google Scholar] [CrossRef]
- Farmer, E.E. Surface-to-air signals. Nature 2001, 411, 854–856. [Google Scholar] [CrossRef]
- Paré, P.W.; Tumlinson, J.H. De novo biosynthesis of volatiles induced by insect herbivory in cotton plants. Plant Physiol. 1997, 114, 1161–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turlings, T.C.J.; Lengwiler, U.B.; Bernasconi, M.L.; Wechsler, D. Timing of induced volatile emissions in maize seedlings. Planta 1998, 207, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, R.; Venables, B.; Petros, R.A.; Nalam, V.; Li, M.; Wang, X.; Takemoto, L.J.; Shah, J. An abietane diterpenoid is a potent activator of systemic acquired resistance. Plant J. 2012, 71, 161–172. [Google Scholar] [CrossRef]
- Maldonado, A.M.; Doerner, P.; Dixon, R.A.; Lamb, C.J.; Cameron, R.K. A putative lipid transfer protein involved in systemic resistance signaling in Arabidopsis. Nature 2002, 419, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Chanda, B.; Xia, Y.; Mandal, M.K.; Yu, K.; Sekine, K.T.; Gao, Q.M.; Selote, D.; Hu, Y.; Stromberg, A.; Navarre, D.; et al. Glycerol-3-phosphate is a critical mobile inducer of systemic immunity in plants. Nat. Genet. 2011, 43, 421–427. [Google Scholar] [CrossRef]
- Jung, H.W.; Tschaplinkski, T.J.; Wang, L.; Glazebrook, J.; Greenberg, J.T. Priming in systemic plant immunity. Science 2009, 324, 89–91. [Google Scholar] [CrossRef]
- Liu, P.P.; von Dahl, C.C.; Park, S.W.; Klessig, D.F. Interconnection between methyl salicylate and lipid-based long-distance signaling during the development of systemic acquired resistance in Arabidopsis and tobacco. Plant Physiol. 2011, 155, 1762–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandi, A.; Welti, R.; Shah, J. The Arabidopsis thaliana dihydroxyacetone phosphate reductase gene suppressor of fatty acid desaturase deficiency1 is required for glycerolipid metabolism and for the activation of systemic acquired resistance. Plant Cell 2004, 16, 465–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenc-Kukula, K.; Chaturvedi, R.; Roth, M.; Welti, R.; Shah, J. Biochemical and molecular-genetic characterization of the Arabidopsis thaliana SFD1-encoded dihydroxyacetone phosphate reductase. Front. Plant Sci. 2012, 3, 26. [Google Scholar] [PubMed] [Green Version]
- Chaturvedi, R.; Krothapalli, K.; Makandar, R.; Nandi, A.; Sparks, A.A.; Roth, M.R.; Welti, R.; Shah, J. Plastid ω-3 desaturase-dependent accumulation of a systemic acquired resistance inducing activity in petiole exudates of Arabidopsis thaliana is independent of jasmonic acid. Plant J. 2008, 54, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Zoeller, M.; Stingl, N.; Krischke, M.; Fekete, A.; Waller, F.; Berger, S.; Mueller, M.J. Lipid profiling of the Arabidopsis hypersensitive response reveals specific lipid peroxidation and fragmentation processes: Biogenesis of pimelic and azelaic acid. Plant Physiol. 2012, 160, 365–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, K.; Soares, J.M.; Mandal, M.K.; Wang, C.; Chanda, B.; Gifford, A.N.; Fowler, J.S.; Navarre, D.; Kachroo, A.; Kachroo, P. A feedback regulatory loop between G3P and lipid transfer proteins DIR1 and AZI1 mediates azelaic-acid-induced systemic immunity. Cell Rep. 2013, 3, 1266–1278. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; El-Shetehy, M.; Shine, M.B.; Yu, K.; Navarre, D.; Wendehenne, D.; Kachroo, A.; Kachroo, P. Free radicals mediate systemic acquired resistance. Cell Rep. 2014, 7, 348–355. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [Green Version]
- Dennis, P.G.; Miller, A.J.; Hirsch, P.R. Are root exudates more important than other sources of rhizodeposits in structuring rhizosphere bacterial communities? FEMS Microbiol. Ecol. 2010, 72, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the root microbiome by plant molecules: The basis for targeted disease suppression and plant growth promotion. Front. Plant Sci. 2020, 10, 1741. [Google Scholar] [CrossRef]
- Rolfe, S.A.; Griffiths, J.; Ton, J. Crying out for help with root exudates: Adaptive mechanisms by which stressed plants assemble health-promoting soil microbiomes. Curr. Opin. Microbiol. 2019, 49, 73–82. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Verbon, E.H.; Trapet, P.L.; Stringlis, I.A.; Kruijs, S.; Bakker, P.A.H.M.; Pieterse, C.M.J. Iron and Immunity. Annu. Rev. Phytopathol. 2017, 55, 355–375. [Google Scholar] [CrossRef] [PubMed]
- Al-Babili, S.; Bouwmeester, H.J. Strigolactones, a novel carotenoid-derived plant hormone. Annu. Rev. Plant Biol. 2015, 66, 161–186. [Google Scholar] [CrossRef]
- Yoneyama, K.; Yoneyama, K.; Takeuchi, Y.; Sekimoto, H. Phosphorus deficiency in red clover promotes exudation of orobanchol, the signal for mycorrhizal symbionts and germination stimulant for root parasites. Planta 2007, 225, 1031–1038. [Google Scholar] [CrossRef]
- Gomez-Roldan, V.; Girard, D.; Bécard, G.; Puech, V. Strigolactones: Promising plant signals. Plant Signal. Behav. 2007, 2, 163–164. [Google Scholar] [CrossRef]
- Abdel-Lateif, K.; Bogusz, D.; Hocher, V. The role of flavonoids in the establishment of plant roots endosymbioses with arbuscular mycorrhiza fungi, rhizobia and Frankia bacteria. Plant Signal. Behav. 2012, 7, 636–641. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.H.; Schmidt, W. Mobilization of iron by plant-borne coumarins. Trends Plant Sci. 2017, 22, 538–548. [Google Scholar] [CrossRef]
- Fourcroy, P.; Sisó-Terraza, P.; Sudre, D.; Savirón, M.; Reyt, G.; Gaymard, F.; Abadía, A.; Abadia, J.; Alvarez-Fernández, A.; Briat, J.F. Involvement of the ABCG37 transporter in secretion of scopoletin and derivatives by Arabidopsis roots in response to iron deficiency. New Phytol. 2014, 201, 155–167. [Google Scholar] [CrossRef]
- Rajniak, J.; Giehl, R.F.H.; Chang, E.; Murgia, I.; von Wirén, N.; Sattely, E.S. Biosynthesis of redox-active metabolites in response to iron deficiency in plants. Nat. Chem. Biol. 2018, 14, 442–450. [Google Scholar] [CrossRef]
- Schmidt, H.; Günther, C.; Weber, M.; Spörlein, C.; Loscher, S.; Böttcher, C.; Schobert, R.; Clemens, S. Metabolome analysis of Arabidopsis thaliana roots identifies a key metabolic pathway for iron acquisition. PLoS ONE 2014, 9, e102444. [Google Scholar] [CrossRef] [Green Version]
- Siwinska, J.; Siatkowska, K.; Olry, A.; Grosjean, J.; Hehn, A.; Bourgaud, F.; Meharg, A.A.; Carey, M.; Lojkowska, E.; Ihnatowicz, A. Scopoletin 8-hydroxylase: A novel enzyme involved in coumarin biosynthesis and iron-deficiency responses in Arabidopsis. J. Exp. Bot. 2018, 69, 1735–1748. [Google Scholar] [CrossRef] [Green Version]
- Jaeger, C.H., III; Lindow, S.E.; Miller, W.; Clark, E.; Firestone, M.K. Mapping of sugar and amino acid availability in soil around roots with bacterial sensors of sucrose and tryptophan. Appl. Environ. Microbiol. 1999, 65, 2685–2690. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Zhao, J.; Wen, T.; Zhao, M.; Li, R.; Goossens, P.; Huang, Q.; Bai, Y.; Vivanco, J.M.; Kowalchuk, G.A.; et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome 2018, 6, 156. [Google Scholar] [CrossRef] [Green Version]
- Bezemer, T.M.; van Dam, N.M. Linking aboveground and belowground interactions via induced plant defenses. Trends. Ecol. Evol. 2005, 20, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Jousset, A.; Rochat, L.; Lanoue, A.; Bonkowski, M.; Keel, C.; Scheu, S. Plants respond to pathogen infection by enhancing the antifungal gene expression of root-associated bacteria. Mol. Plant. Microbe Interact. 2011, 24, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Bais, H.P.; Walker, T.S.; Schweizer, H.P.; Vivanco, J.M. Root specific elicitation and antimicrobial activity of rosmarinic acid in hairy root cultures of sweet basil (Ocimum basilicum L.). Plant. Physiol. Biochem. 2002, 40, 983–995. [Google Scholar] [CrossRef]
- Lanoue, A.; Burlat, V.; Henkes, G.J.; Koch, I.; Schurr, U.; Röse, U.S. De novo biosynthesis of defense root exudates in response to Fusarium attack in barley. New Phytol. 2010, 185, 577–588. [Google Scholar] [CrossRef]
- Kaplan, I.; Halitschke, R.; Kessler, A.; Sardanelli, S.; Denno, R.F. Effects of plant vascular architecture on aboveground–belowground-induced responses to foliar and root herbivores on Nicotiana tabacum. J. Chem. Ecol. 2008, 34, 1349–1359. [Google Scholar] [CrossRef]
- Balendres, M.A.; Nichols, D.S.; Tegg, R.S.; Wilson, C.R. Metabolomes of potato root exudates: Compounds that stimulate resting spore germination of the soil-borne pathogen Spongospora subterranea. J. Agric. Food Chem. 2016, 64, 7466–7674. [Google Scholar] [CrossRef]
- Li, C.; Yu, J.; Gan, L.; Sun, J. Effects of tobacco pathogens and their antagonistic bacteria on tobacco root exudates. Open J. Appl. Sci. 2018, 8, 518–531. [Google Scholar] [CrossRef] [Green Version]
- Pétriacq, P.; Williams, A.; Cotton, A.; McFarlane, A.E.; Rolfe, S.A.; Ton, J. Metabolite profiling of non-sterile rhizosphere soil. Plant J. 2017, 92, 147–162. [Google Scholar] [CrossRef]
- Robert, C.A.; Veyrat, N.; Glauser, G.; Marti, G.; Doyen, G.R.; Villard, N.; Gaillard, M.D.; Köllner, T.G.; Giron, D.; Body, M.; et al. A specialist root herbivore exploits defensive metabolites to locate nutritious tissues. Ecol. Lett. 2012, 15, 55–64. [Google Scholar] [CrossRef]
- Hu, L.; Robert, C.A.M.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.; Manzo, D.; Chervet, N.; Steinger, T.; van der Heijden, M.G.A.; et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 2738. [Google Scholar] [CrossRef] [Green Version]
- Rasmann, S.; Köllner, T.G.; Degenhardt, J.; Hiltpold, I.; Toepfer, S.; Kuhlmann, U.; Gershenzon, J.; Turlings, T.C. Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 2005, 434, 732–737. [Google Scholar] [CrossRef]
- Ali, J.G.; Alborn, H.T.; Stelinski, L.L. Subterranean herbivore-induced volatiles released by citrus roots upon feeding by Diaprepes abbreviatus recruit entomopathogenic nematodes. J. Chem. Ecol. 2010, 36, 361–368. [Google Scholar] [CrossRef]
- Yang, J.W.; Yi, H.S.; Kim, H.; Lee, B.; Lee, S.; Ghim, S.Y.; Ryu, C.M. Whitefly infestation of pepper plants elicits defence responses against bacterial pathogens in leaves and roots and changes the below-ground microflora. J. Ecol. 2011, 99, 46–56. [Google Scholar] [CrossRef]
- Hoysted, G.A.; Bell, C.A.; Lilley, C.J.; Urwin, P.E. Aphid colonization affects potato root exudate composition and the hatching of a soil borne pathogen. Front. Plant Sci. 2018, 9, 1278. [Google Scholar] [CrossRef]
- Saravesi, K.; Aikio, S.; Wäli, P.R.; Ruotsalainen, A.L.; Kaukonen, M.; Huusko, K.; Suokas, M.; Brown, S.P.; Jumpponen, A.; Tuomi, J.; et al. Moth outbreaks alter root-associated fungal communities in subarctic mountain birch forests. Microb. Ecol. 2015, 69, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Lankau, R.A.; Wheeler, E.; Bennett, A.E.; Strauss, S.Y. Plant–soil feedbacks contribute to an intransitive competitive network that promotes both genetic and species diversity. J. Ecol. 2011, 99, 176–185. [Google Scholar] [CrossRef]
- Neal, A.L.; Ahmad, S.; Gordon-Weeks, R.; Ton, J. Benzoxazinoids in root exudates of maize attract Pseudomonas putida to the rhizosphere. PLoS ONE 2012, 7, e35498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniou, A.; Tsolakidou, M.D.; Stringlis, I.A.; Pantelides, I.S. Rhizosphere microbiome recruited from a suppressive compost improves plant fitness and increases protection against vascular wilt pathogens of tomato. Front. Plant Sci. 2017, 8, 2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardi, N.; Vitale, S.; Turrà, D.; Reverberi, M.; Fanelli, C.; Vinale, F.; Marra, R.; Ruocco, M.; Pascale, A.; d’Errico, G.; et al. Root exudates of stressed plants stimulate and attract Trichoderma soil fungi. Mol. Plant. Microbe Interact. 2018, 31, 982–994. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, D.S.; Teixeira, P.J.P.L. Root-exuded coumarin shapes the root microbiome. Proc. Natl. Acad. Sci. USA 2018, 115, 5629–5631. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; Mazzola, M. ECOLOGY. Soil immune responses. Science 2016, 352, 1392–1393. [Google Scholar] [CrossRef] [PubMed]
- Rudrappa, T.; Czymmek, K.J.; Paré, P.W.; Bais, H.P. Root-secreted malic acid recruits beneficial soil bacteria. Plant Physiol. 2008, 148, 1547–1556. [Google Scholar] [CrossRef] [Green Version]
- Chapelle, E.; Mendes, R.; Bakker, P.A.; Raaijmakers, J.M. Fungal invasion of the rhizosphere microbiome. ISME J. 2016, 10, 265–268. [Google Scholar] [CrossRef] [Green Version]
- Ourry, M.; Lebreton, L.; Chaminade, V.; Guillerm-Erckelboudt, A.Y.; Hervé, M.; Linglin, J.; Marnet, N.; Ourry, A.; Paty, C.; Poinsot, D.; et al. Influence of belowground herbivory on the dynamics of root and rhizosphere microbial communities. Front. Ecol. Evol. 2018, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Grayston, J.S.; Dawson, L.A.; Treonis, A.M.; Murray, P.J.; Ross, J.; Reid, E.J.; MacDougall, R. Impact of root herbivory by insect larvae on soil microbial communities. Eur. J. Soil Biol. 2001, 37, 277–280. [Google Scholar] [CrossRef]
- Chin-A-Woeng, T.F.; Bloemberg, G.V.; Mulders, I.H.; Dekkers, L.C.; Lugtenberg, B.J. Root colonization by phenazine-1-carboxamide-producing bacterium Pseudomonas chlororaphis PCL1391 is essential for biocontrol of tomato foot and root rot. Mol. Plant Microbe Interact. 2000, 13, 1340–1345. [Google Scholar] [CrossRef] [Green Version]
- Haas, D.; Keel, C. Regulation of antibiotic production in root-colonizing Pseudomonas spp. and relevance for biological control of plant disease. Annu. Rev. Phytopathol. 2003, 41, 117–153. [Google Scholar] [CrossRef]
- Mavrodi, D.V.; Blankenfeldt, W.; Thomashow, L.S. Phenazine compounds in fluorescent pseudomonas spp. biosynthesis and regulation. Annu. Rev. Phytopathol. 2006, 44, 417–445. [Google Scholar] [CrossRef] [PubMed]
- Thomashow, L.S.; Weller, D.M. Current concepts in the use of introduced bacteria for biological disease control: Mechanisms and antifungal metabolites. In Plant-Microbe Interactions; Stacey, G., Keen, N., Eds.; Chapman & Hall: New York, NY, USA, 1996; Volume 1, pp. 187–235. [Google Scholar]
- Nowak-Thompson, B.; Chaney, N.; Wing, J.S.; Gould, S.J.; Loper, J.E. Characterization of the pyoluteorin biosynthetic gene cluster of Pseudomonas fluorescens Pf-5. J. Bacteriol. 1999, 181, 2166–2174. [Google Scholar] [CrossRef] [Green Version]
- Kirner, S.; Hammer, P.E.; Hill, D.S.; Altmann, A.; Fischer, I.; Weislo, L.J.; Lanahan, M.; van Pée, K.H.; Ligon, J.M. Functions encoded by pyrrolnitrin biosynthetic genes from Pseudomonas fluorescens. J. Bacteriol. 1998, 180, 1939–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomashow, L.S.; Weller, D.M. Role of a phenazine antibiotic from Pseudomonas fluorescens in biological control of Gaeumannomyces graminis var. tritici. J. Bacteriol. 1988, 170, 3499–3508. [Google Scholar] [CrossRef] [Green Version]
- Voisard, C.; Keel, C.; Haas, D.; Dèfago, G. Cyanide production by Pseudomonas fluorescens helps suppress black root rot of tobacco under gnotobiotic conditions. EMBO J. 1989, 8, 351–358. [Google Scholar] [CrossRef]
- Romero, D.; de Vicente, A.; Olmos, J.L.; Dávila, J.C.; Pérez-García, A. Effect of lipopeptides of antagonistic strains of Bacillus subtilis on the morphology and ultrastructure of the cucurbit fungal pathogen Podosphaera fusca. J. Appl. Microbiol. 2007, 103, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Cazorla, F.M.; Romero, D.; Pérez-García, A.; Lugtenberg, B.J.; de Vicente, A.; Bloemberg, G. Isolation and characterization of antagonistic Bacillus subtilis strains from the avocado rhizoplane displaying biocontrol activity. J. Appl. Microbiol. 2007, 103, 1950–1959. [Google Scholar] [CrossRef]
- Bais, H.P.; Fall, R.; Vivanco, J.M. Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef] [Green Version]
- Baetz, U.; Martinoia, E. Root exudates: The hidden part of plant defense. Trends Plant Sci. 2014, 19, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanEtten, H.D.; Mansfield, J.W.; Bailey, J.A.; Farmer, E.E. Two classes of plant antibiotics: Phytoalexins versus ‘phytoanticipins’. Plant Cell 1994, 6, 1191–1192. [Google Scholar] [CrossRef] [PubMed]
- Lanoue, A.; Burlat, V.; Schurr, U.; Röse, U.S. Induced root-secreted phenolic compounds as a belowground plant defense. Plant Signal. Behav. 2010, 5, 1037–1038. [Google Scholar] [CrossRef] [PubMed]
- Stringlis, I.A.; Yu, K.; Feussner, K.; de Jonge, R.; Van Bentum, S.; Van Verk, M.C.; Berendsen, R.L.; Bakker, P.A.H.M.; Feussner, I.; Pieterse, C.M.J. MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health. Proc. Natl. Acad. Sci. USA 2018, 115, E5213–E5222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duchesne, L.C.; Peterson, R.L.; Ellis, B.E. Interaction between the ectomycorrhizal fungus Paxillus involutus and Pinus resinosa induces resistance to Fusarium oxysporum. Can. J. Bot. 1988, 66, 558–562. [Google Scholar] [CrossRef]
- Romera, F.J.; García, M.J.; Lucena, C.; Martínez-Medina, A.; Aparicio, M.A.; Ramos, J.; Alcántara, E.; Angulo, M.; Pérez-Vicente, R. Induced systemic resistance (ISR) and Fe deficiency responses in dicot plants. Front. Plant Sci. 2019, 10, 287. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Trotel-Aziz, P.; Villaume, S.; Rabenoelina, F.; Schwarzenberg, A.; Nguema-Ona, E.; Clément, C.; Baillieul, F.; Aziz, A. Bacillus subtilis and Pseudomonas fluorescens trigger common and distinct systemic immune responses in Arabidopsis thaliana depending on the pathogen lifestyle. Vaccines 2020, 8, 503. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.D.; Borrego, E.J.; Kenerley, C.M.; Kolomiets, M.V. Oxylipins other than jasmonic acid are xylem-resident signals regulating systemic resistance induced by Trichoderma virens in maize. Plant Cell 2020, 32, 166–185. [Google Scholar] [CrossRef]
- Van Peer, R.; Niemann, G.J.; Schippers, B. Induced resistance and phytoalexin accumulation in biological control of Fusarium wilt of carnation by Pseudomonas spp. strain WCS417r. Phytopathology 1991, 81, 1508–1512. [Google Scholar]
- Wei, G.; Kloepper, J.W.; Tuzun, S. Induction of systemic resistance of cucumber to Colletotrichum orbiculare by select strains of plant growth-promoting rhizobacteria. Phytopathology 1991, 81, 1508–1512. [Google Scholar] [CrossRef]
- Wei, G.; Kloepper, J.W.; Tuzun, S. Induction of systemic resistance to cucumber disease and increased plant growth by plant growth-promoting rhizobacteria under field conditions. Phytopathology 1996, 86, 221–224. [Google Scholar] [CrossRef]
- Liu, L.; Kloepper, J.W.; Tuzun, S. Induction of systemic resistance in cucumber against Fusarium wilt by plant growth-promoting rhizobacteria. Phytopathology 1995, 85, 695–698. [Google Scholar] [CrossRef]
- Liu, L.; Kloepper, J.W.; Tuzun, S. Induction of systemic resistance in cucumber against bacterial angular leaf spot by plant growth-promoting rhizobacteria. Phytopathology 1995, 85, 843–847. [Google Scholar] [CrossRef]
- Raupach, G.S.; Liu, L.; Murphy, J.F.; Tuzun, S.; Kloepper, J.W. Induced systemic resistance in cucumber and tomato against cucumber mosaic cucumovirus using plant growth-promoting rhizobacteria (PGPR). Plant Dis. 1996, 80, 891–894. [Google Scholar] [CrossRef]
- Kloepper, J.W. Plant growth-promoting rhizobacteria as biological control agents. In Soil Microbial Ecology—Applications in Agricultural and Environmental Management; Metting, F.B., Jr., Ed.; Marcel Dekker: New York, NY, USA, 1993; pp. 255–274. [Google Scholar]
- Park, K.S.; Kloepper, J.W. Activation of PR-1a promoter by rhizobacteria that induce systemic resistance in tobacco against Pseudomonas syringae pv. tabaci. Biol. Control 2000, 18, 2–9. [Google Scholar] [CrossRef]
- Zhang, S.; Reddy, M.; Kloepper, J. Development of assays for assessing induced systemic resistance by plant growth-promoting rhizobacteria against Blue Mold of Tobacco. Biol. Control 2002, 23, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Gehring, C.; Bennett, A. Mycorrhizal fungal-plant-insect interactions: The importance of community approach. Environ. Entomol. 2009, 38, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Koricheva, J.; Gange, A.C.; Jones, T. Effects of mycorrhizal fungi on insect herbivores: A meta-analysis. Ecology 2009, 90, 2088–2097. [Google Scholar] [CrossRef]
- Valenzuela-Soto, J.H.; Estrada-Hernández, M.G.; Ibarra-Laclette, E.; Délano-Frier, J.P. Inoculation of tomato plants (Solanum lycopersicum) with growth-promoting Bacillus subtilis retards whitefly Bemisia tabaci development. Planta 2010, 231, 397–410. [Google Scholar] [CrossRef]
- Hanafi, A.; Traoré, M.; Schnitzler, W.; Woitke, M. Induced resistance of tomato to whiteflies and pythium with the PGPR Bacillus subtilis in a soilless crop grown under greenhouse conditions. Acta. Hortic. 2007, 747, 315–323. [Google Scholar] [CrossRef]
- Zehnder, G.; Kloepper, J.; Yao, C.; Wei, G. Induction of systemic resistance in cucumber against cucumber beetles (Coleoptera: Chrysomelidae) by plant growth-promoting rhizobacteria. J. Econ. Entomol. 1997, 90, 391–396. [Google Scholar] [CrossRef]
- Pineda, A.; Zheng, S.J.; Van Loon, J.J.A.; Dicke, M. Rhizobacteria modify plant–aphid interactions: A case of induced systemic susceptibility. Plant Biol. 2012, 14, 83–90. [Google Scholar] [CrossRef]
- Ahn, I.P.; Lee, S.W.; Suh, S.C. Rhizobacteria-induced priming in Arabidopsis is dependent on ethylene, jasmonic acid, and NPR1. Mol. Plant Microbe Interact. 2007, 20, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.J.; Van Wees, S.C.M.; Hoffland, E.; Van Pelt, J.A.; Van Loon, L.C. Systemic resistance in Arabidopsis induced by biocontrol bacteria is independent of salicylic acid accumulation and pathogenesis-related gene expression. Plant Cell 1996, 8, 1225–1237. [Google Scholar]
- Van Oosten, V.R.; Bodenhausen, N.; Reymond, P.; Van Pelt, J.A.; Van Loon, L.C.; Dicke, M.; Pieterse, C.M. Differential effectiveness of microbially induced resistance against herbivorous insects in Arabidopsis. Mol. Plant Microbe Interact. 2008, 21, 919–930. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.; van Wees, S.C.; van Pelt, J.A.; Knoester, M.; Laan, R.; Gerrits, H.; Weisbeek, P.J.; van Loon, L.C. A novel signaling pathway controlling induced systemic resistance in Arabidopsis. Plant Cell 1998, 10, 1571–1580. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Audenaert, K.; Pattery, T.; Cornelis, P.; Höfte, M. Induction of systemic resistance to Botrytis cinerea in tomato by Pseudomonas aeruginosa 7NSK2: Role of salicylic acid, pyochelin, and pyocyanin. Mol. Plant Microbe Interact. 2002, 15, 1147–1156. [Google Scholar] [CrossRef] [Green Version]
- De Meyer, G.; Capieau, K.; Audenaert, K.; Buchala, A.; Métraux, J.P.; Höfte, M. Nanogram amounts of salicylic acid produced by the rhizobacterium Pseudomonas aeruginosa 7NSK2 activate the systemic acquired resistance pathway in bean. Mol. Plant Microbe Interact. 1999, 12, 450–458. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.S.; Ryu, C.M. Understanding cross-communication between aboveground and belowground tissues via transcriptome analysis of a sucking insect whitefly-infested pepper plants. Biochem. Biophys. Res. Commun. 2014, 443, 272–277. [Google Scholar] [CrossRef]
- Park, Y.S.; Bae, D.W.; Ryu, C.M. Aboveground whitefly infestation modulates transcriptional levels of anthocyanin biosynthesis and jasmonic acid signaling-related genes and augments the cope with drought stress of maize. PLoS ONE 2015, 10, e0143879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, A.C.; Jiang, T.; Liu, Y.X.; Bai, Y.C.; Reed, J.; Qu, B.; Goossens, A.; Nützmann, H.W.; Bai, Y.; Osbourn, A. A specialized metabolic network selectively modulates Arabidopsis root microbiota. Science 2019, 364, eaau6389. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Weller, D.M. Natural plant protection by 2,4-diacetylphloroglucinol-producing Pseudomonas spp. in take-all decline soils. Mol. Plant Microbe Interact. 1998, 11, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F.; Gerbino, R.; et al. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 2015, 17, 316–331. [Google Scholar] [CrossRef] [PubMed]
- Tsolakidou, M.D.; Stringlis, I.A.; Fanega-Sleziak, N.; Papageorgiou, S.; Tsalakou, A.; Pantelides, I.S. Rhizosphere-enriched microbes as a pool to design synthetic communities for reproducible beneficial outputs. FEMS Microbiol. Ecol. 2019, 95, fiz138. [Google Scholar] [CrossRef] [PubMed]
- Vannier, N.; Agler, M.; Hacquard, S. Microbiota-mediated disease resistance in plants. PLoS Pathog. 2019, 15, e1007740. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Newton, A.C. Climate change, plant diseases and food security: An overview. Plant. Pathol. 2011, 60, 2–14. [Google Scholar] [CrossRef]
- Doornbos, R.F.; van Loon, L.C.; Bakker, P.A.H. Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere. Agron. Sustain. Dev. 2012, 32, 227–243. [Google Scholar] [CrossRef]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and bacterial plant diseases. Front. Microbiol. 2017, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Schematic representation of the plant defense response. (A) Indirect defense. Whitefly infestation elicits the plant defense response, which indirectly suppresses the performance of insects attracted by natural volatiles. (B) Induced systemic resistance (ISR). Beneficial microbes in the rhizosphere induce systemic signals and systemic-acquired resistance against pathogens and insects. (C) Plant social networking system (pSNS). Insect infestation and pathogen infection of aboveground tissues induces the secretion of root exudates into the rhizosphere, which leads to the recruitment of beneficial microbes, thus activating systemic resistance against pathogens and insects.
Figure 1.
Schematic representation of the plant defense response. (A) Indirect defense. Whitefly infestation elicits the plant defense response, which indirectly suppresses the performance of insects attracted by natural volatiles. (B) Induced systemic resistance (ISR). Beneficial microbes in the rhizosphere induce systemic signals and systemic-acquired resistance against pathogens and insects. (C) Plant social networking system (pSNS). Insect infestation and pathogen infection of aboveground tissues induces the secretion of root exudates into the rhizosphere, which leads to the recruitment of beneficial microbes, thus activating systemic resistance against pathogens and insects.
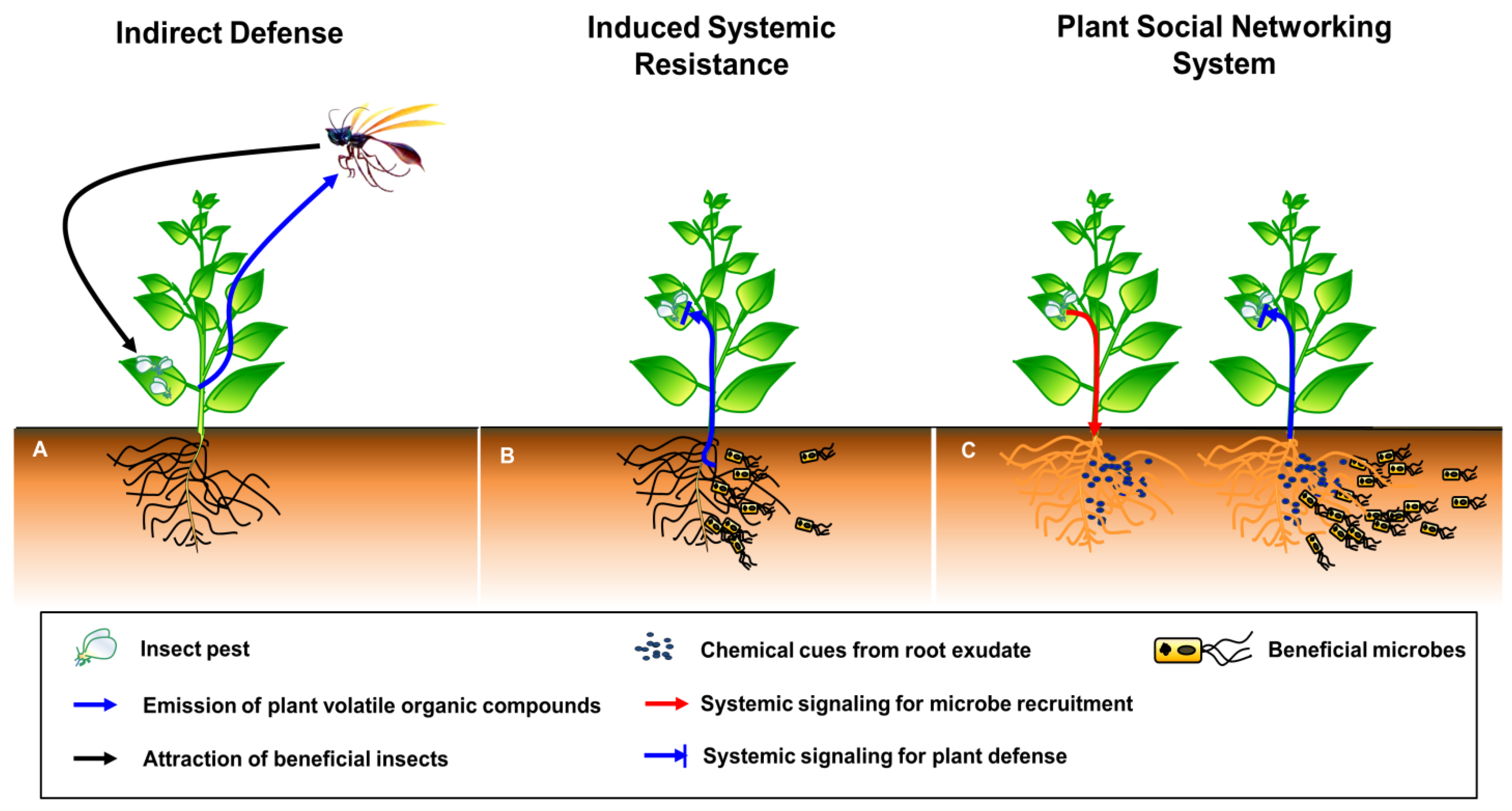
Figure 2.
Schematic representation of the plant SNS. The events at each step of multitrophic interactions include (1) elicitation; (2) systemic signaling; (3) root exudate secretion; and (4) plant protection.
Figure 2.
Schematic representation of the plant SNS. The events at each step of multitrophic interactions include (1) elicitation; (2) systemic signaling; (3) root exudate secretion; and (4) plant protection.
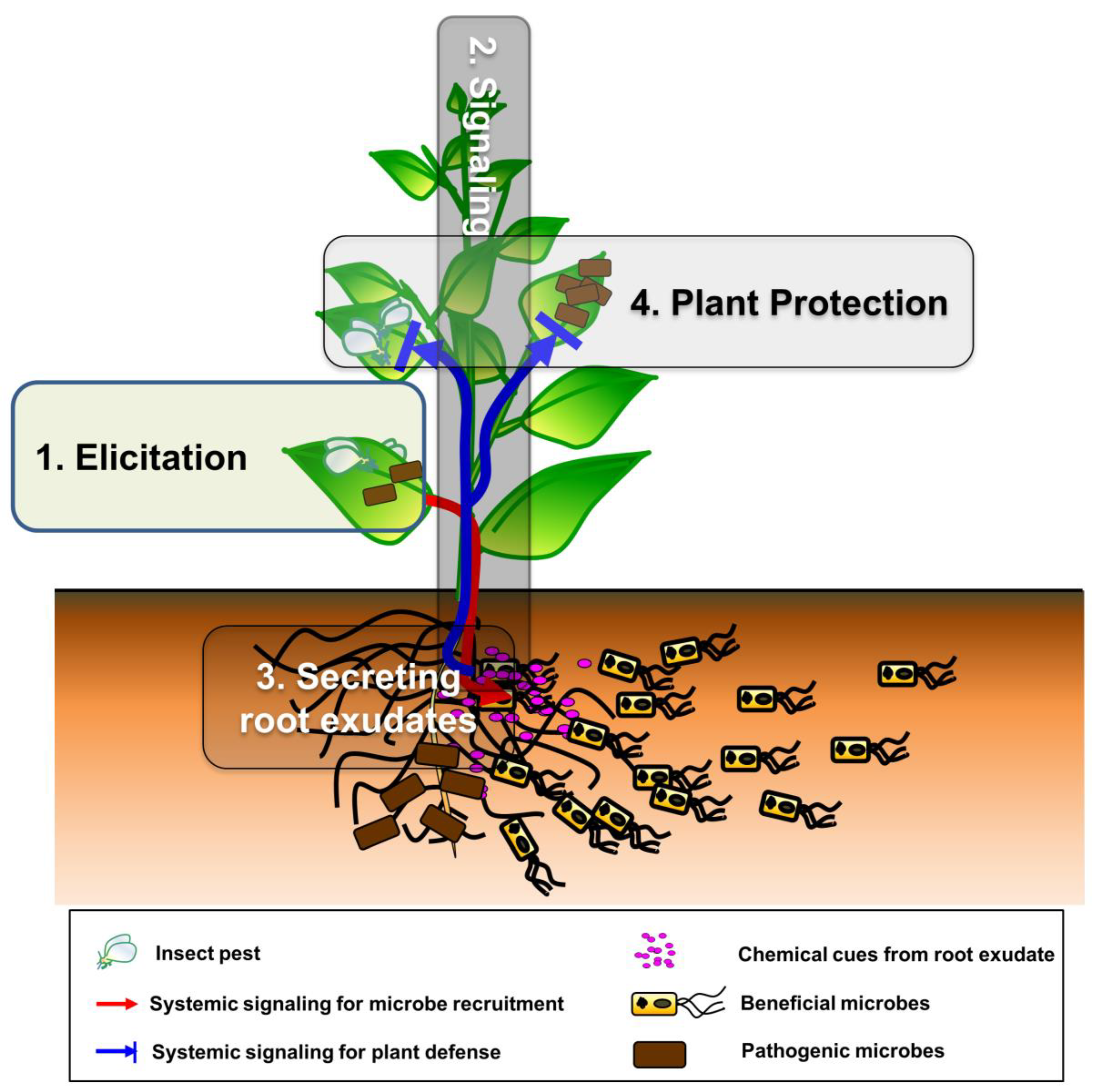

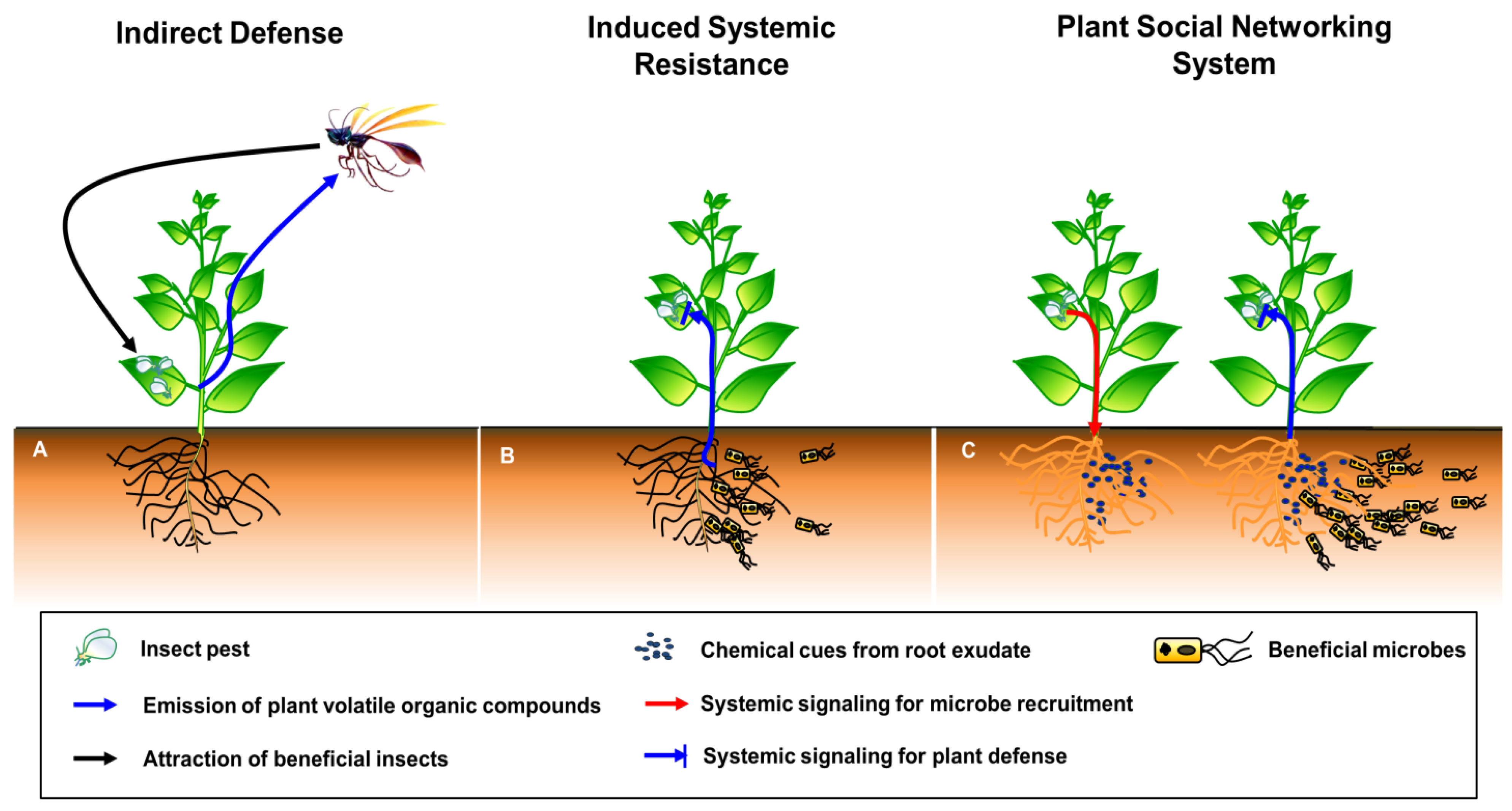{kind=link}
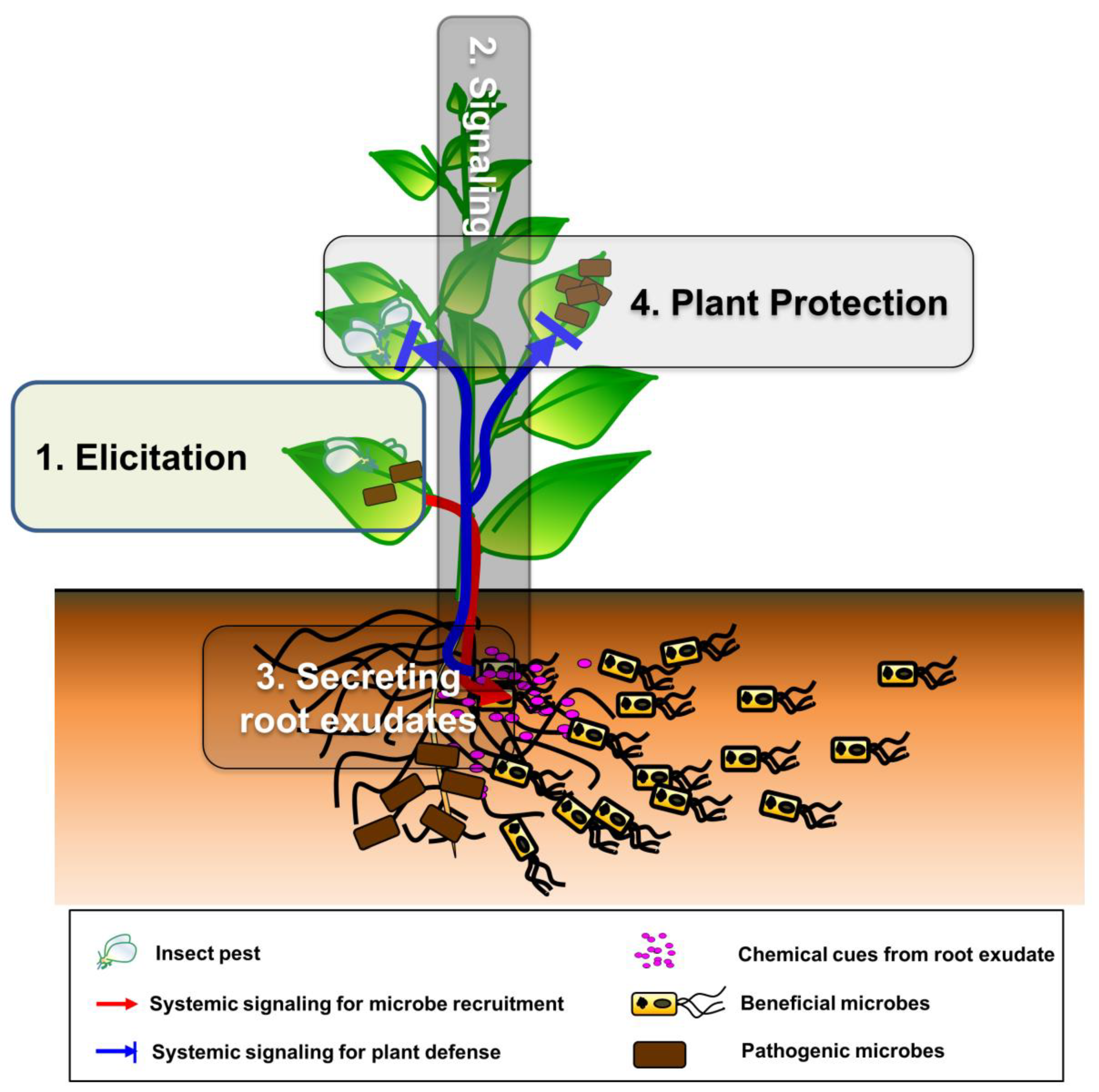{kind=link}
Table 1.
Summary of plant SNS steps described in this review.
| Steps of Plant SNS | Triggers/Determinants | Effect/Mechanisms on Plant | References |
|---|---|---|---|
| PAMPs/MAMPs/HAMPs: flg22, elf18/elf26, peptidoglycans, chitin, volicitin, inceptins, caeliferin, and bruchin | Plant pattern receptors perceive PAMPs/MAMPs/HAMPs | [29,30,31,32,33,34,35,36,37] |
| Ca2+, ROS, MAP Kinase cascades, and phytohormones | Regulation of plant defense responses | [38,39,40,41,42] | |
| SA and Me-SA | Activating systemic resistance against biotrophic pathogens and sucking insects | [43,44,45,56,57,58,59,60,61,62] |
| JA, MeJA, and JA-Ile | Defensive signal against necrotrophic pathogens and chewing insects | [63,71,72,73,74] | |
| Volatile organic compounds (VOCs): C6-alcohol, C6-aldehydes, cis-3-hexen-1-ol trans-2-hexenal, monoterpenes (limonene, linalool, ocimene), and sesquiterpenes (bergamotene, carphyllene and farnesene) | Released by plants in response to a variety of insects | [75,76,77,78,81,82,83,84,85] | |
| Lipid-derived signals: DIR1, G3P, and AzA | Signaling molecules to activate systemic defense responses to pathogens | [86,87,89,91,92,93,94,95] | |
| Stringolactones, flavonoids, and coumains | Secretion under phosphate- and nitrogen-deficient conditions. Effect on the interaction between plant and AM fungi | [104,105,106,107,108,109,110,111,112] |
| Malic acid, phenolic compounds, and organic acids | Secretion after infection with bacterial and fungal pathogens and nematodes | [5,114,116,117,118,119,120,121] | |
| Benzoxazinoids and SA | Secretion upon insect infestation | [11,12,122,123,124,125,126,127,128,129] | |
| Beneficial microbes by root exudates | Recruitment of beneficial microbes from plants infected with pathogens and insects | [5,11,114,124,127,132,133,134,135,136,137,138,139] |
| Antibiosis and antimicrobial compounds: hydrogen cyanide, phenazine-1-carboxylic acid, phenazine-1-carboxyamide, 2,4-diacetyl phloroglucinol, pyoluteorin, pyrrolnitrin, phenazine-1-carboxylic acid, t-cinnamic acid, p-coumaric acid, ferulic acid, syringic acid, vanillic acid, scopoletin, and ethanol-soluble compounds | Direct suppression of pathogens and insects by antibiotics, lipopeptides, phenylpropanoids and ethanol-soluble compounds | [140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155] | |
| Microbes elicit induced systemic resistance | Activation of broad spectrum plant immunity against pathogens and insects | [159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175,176,177,178,179,180] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, Y.-S.; Ryu, C.-M. Understanding Plant Social Networking System: Avoiding Deleterious Microbiota but Calling Beneficials. Int. J. Mol. Sci. 2021, 22, 3319. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073319
AMA Style
Park Y-S, Ryu C-M. Understanding Plant Social Networking System: Avoiding Deleterious Microbiota but Calling Beneficials. International Journal of Molecular Sciences. 2021; 22(7):3319. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073319
Chicago/Turabian StylePark, Yong-Soon, and Choong-Min Ryu. 2021. "Understanding Plant Social Networking System: Avoiding Deleterious Microbiota but Calling Beneficials" International Journal of Molecular Sciences 22, no. 7: 3319. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073319
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.