
Epigenetic Consequences of in Utero Exposure to Rosuvastatin: Alteration of Histone Methylation Patterns in Newborn Rat Brains


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
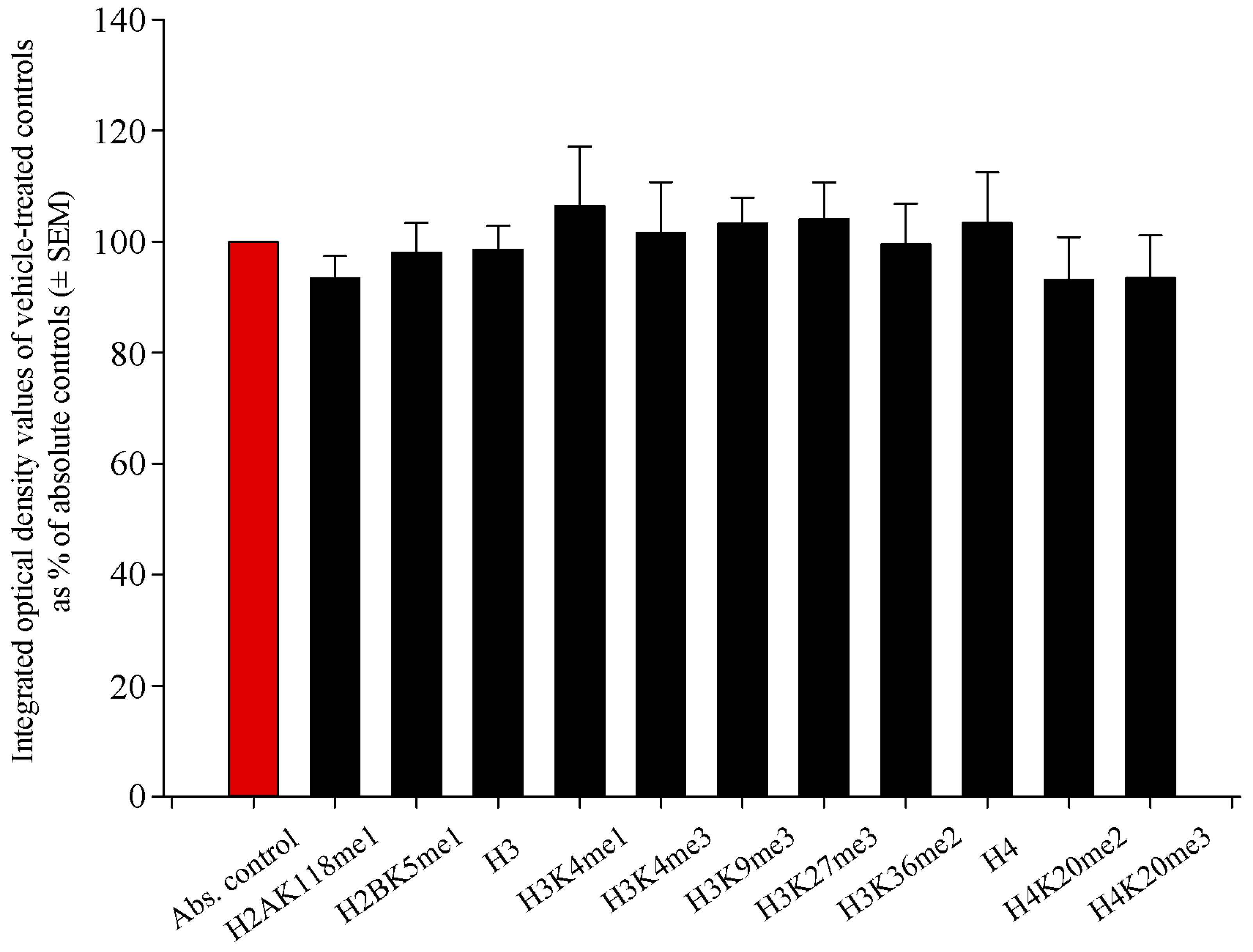
2.1. Histone Methylation Patterns of Absolute Controls and Vehicle-Treated Controls Did Not Differ Significantly
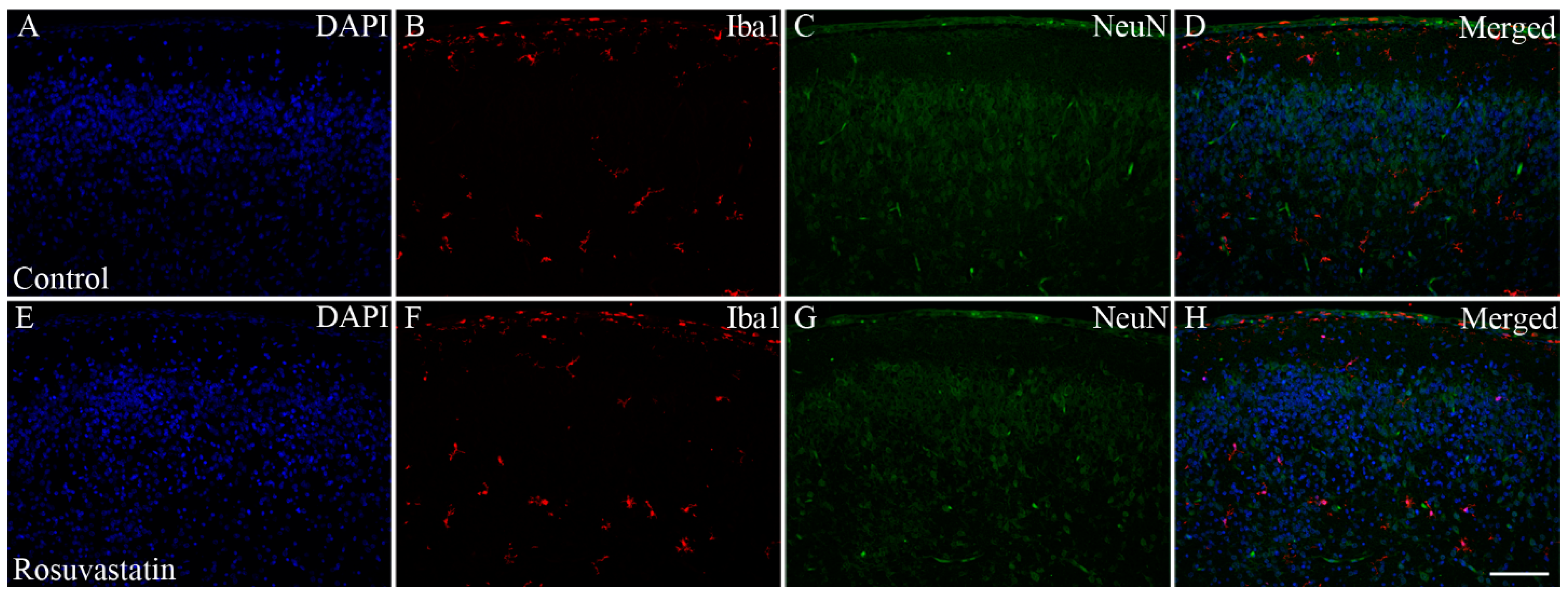
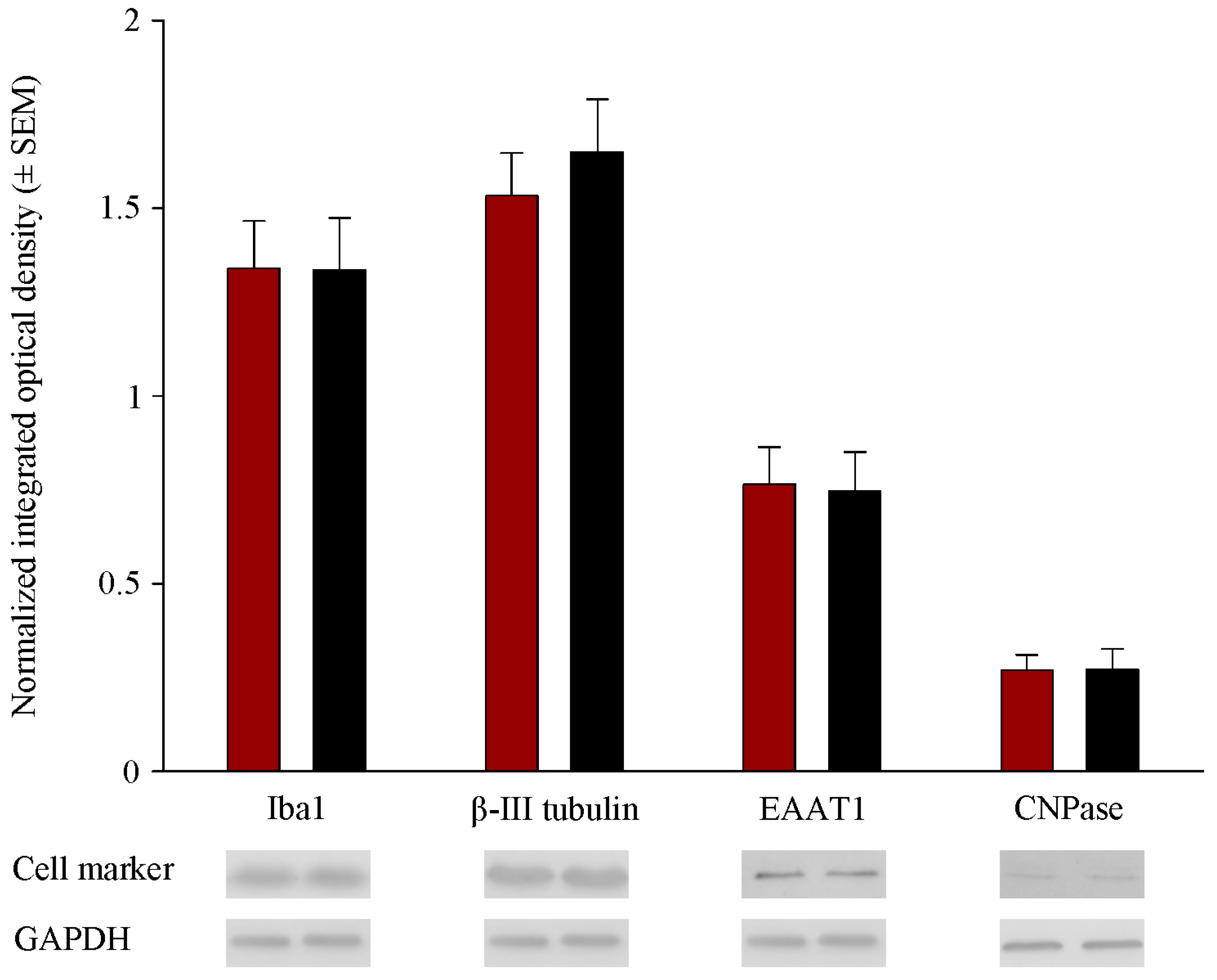
2.2. Prenatal RST Exposure Does Not Affect Cell Composition in the Newborn Brain
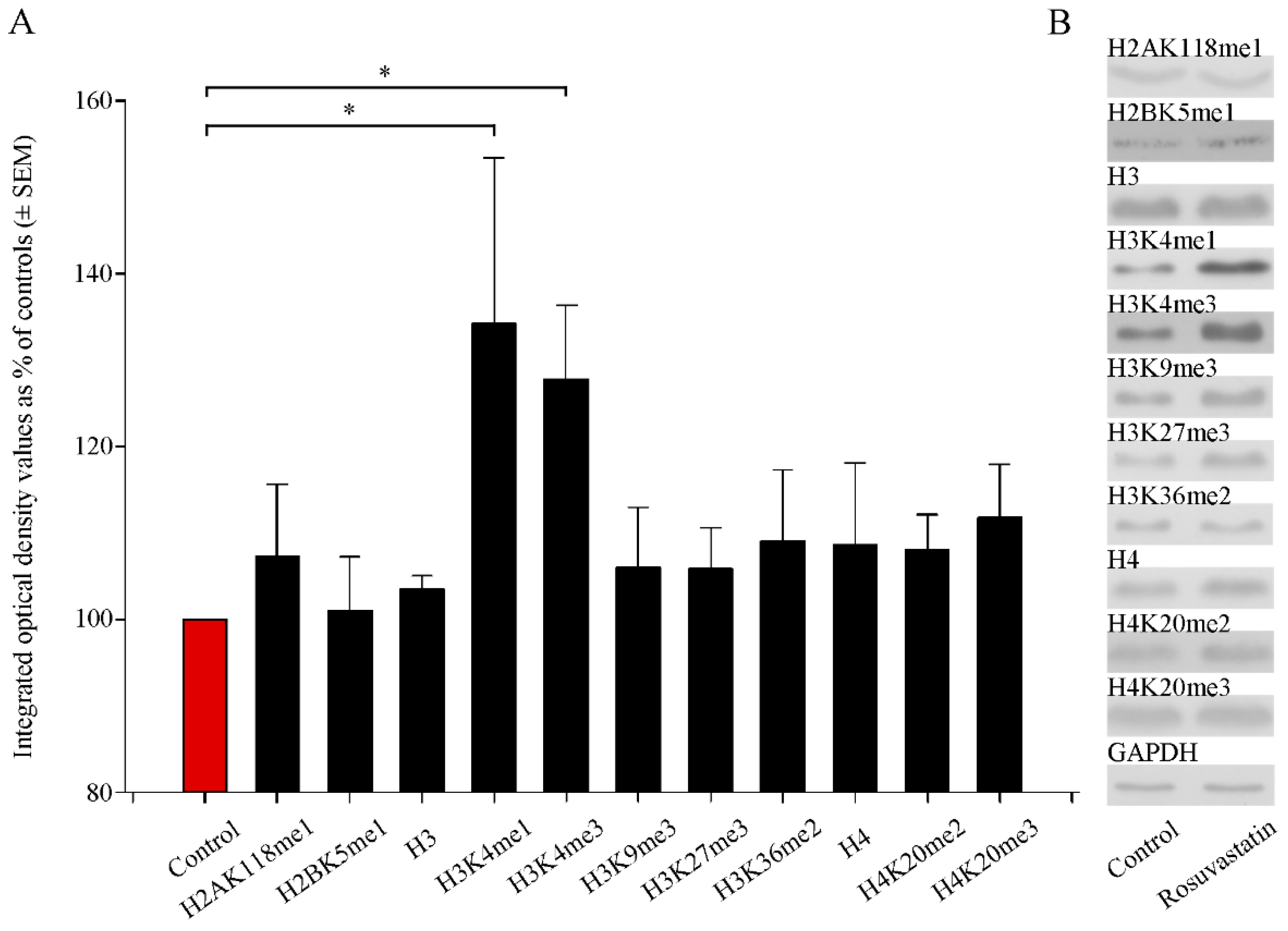
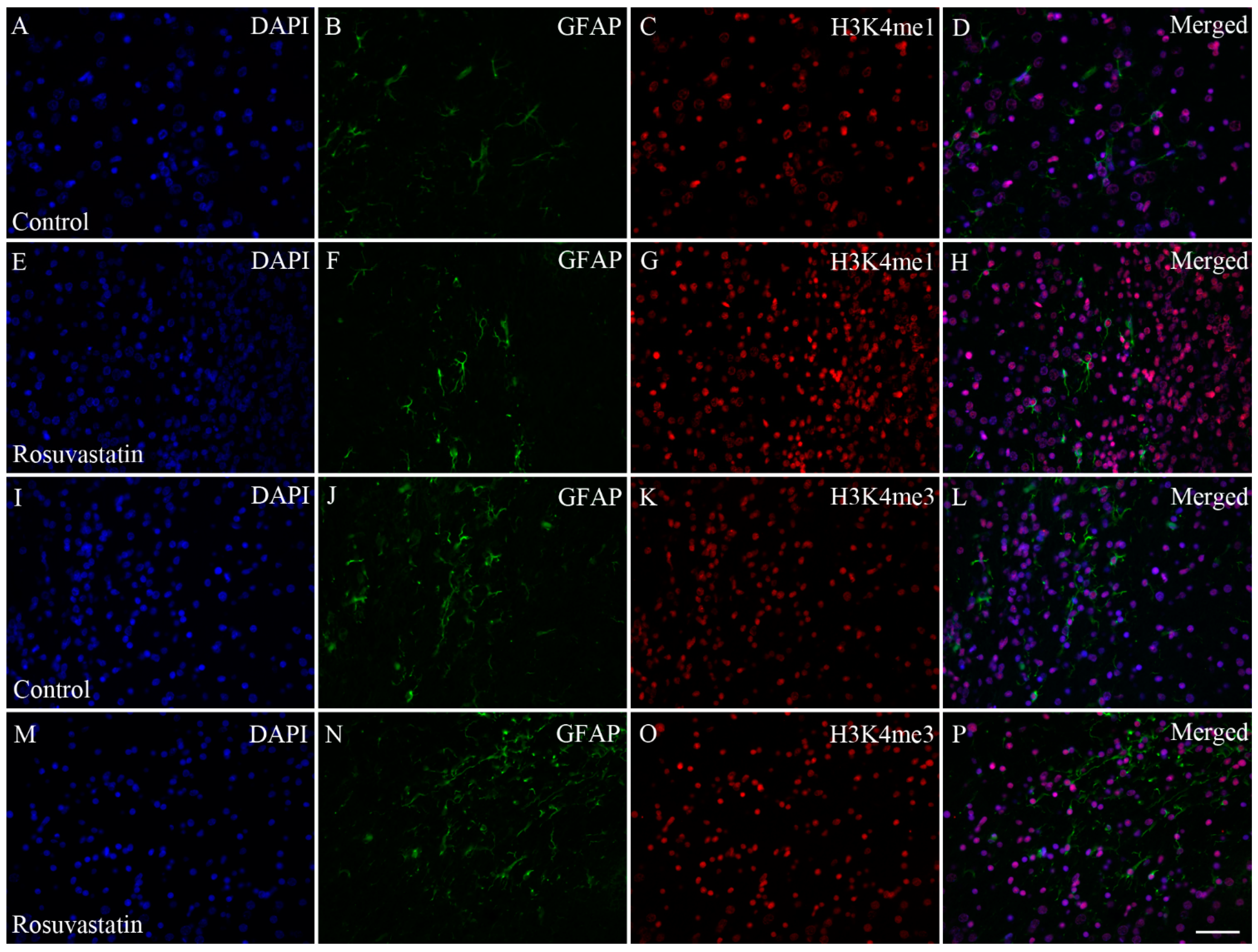
2.3. In Utero RST Exposure Alters Histone Methylation Patterns in the Newborn Brain
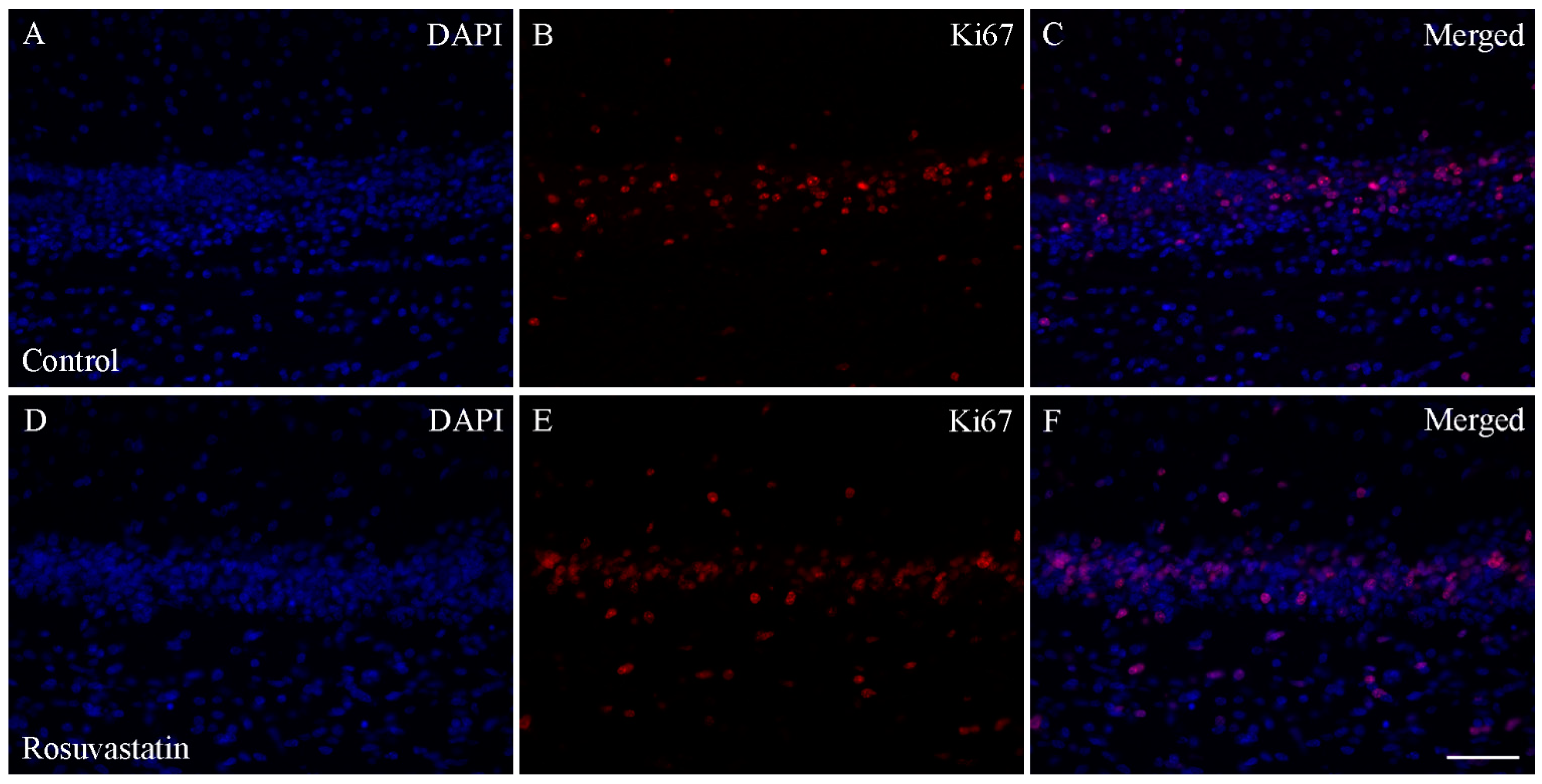
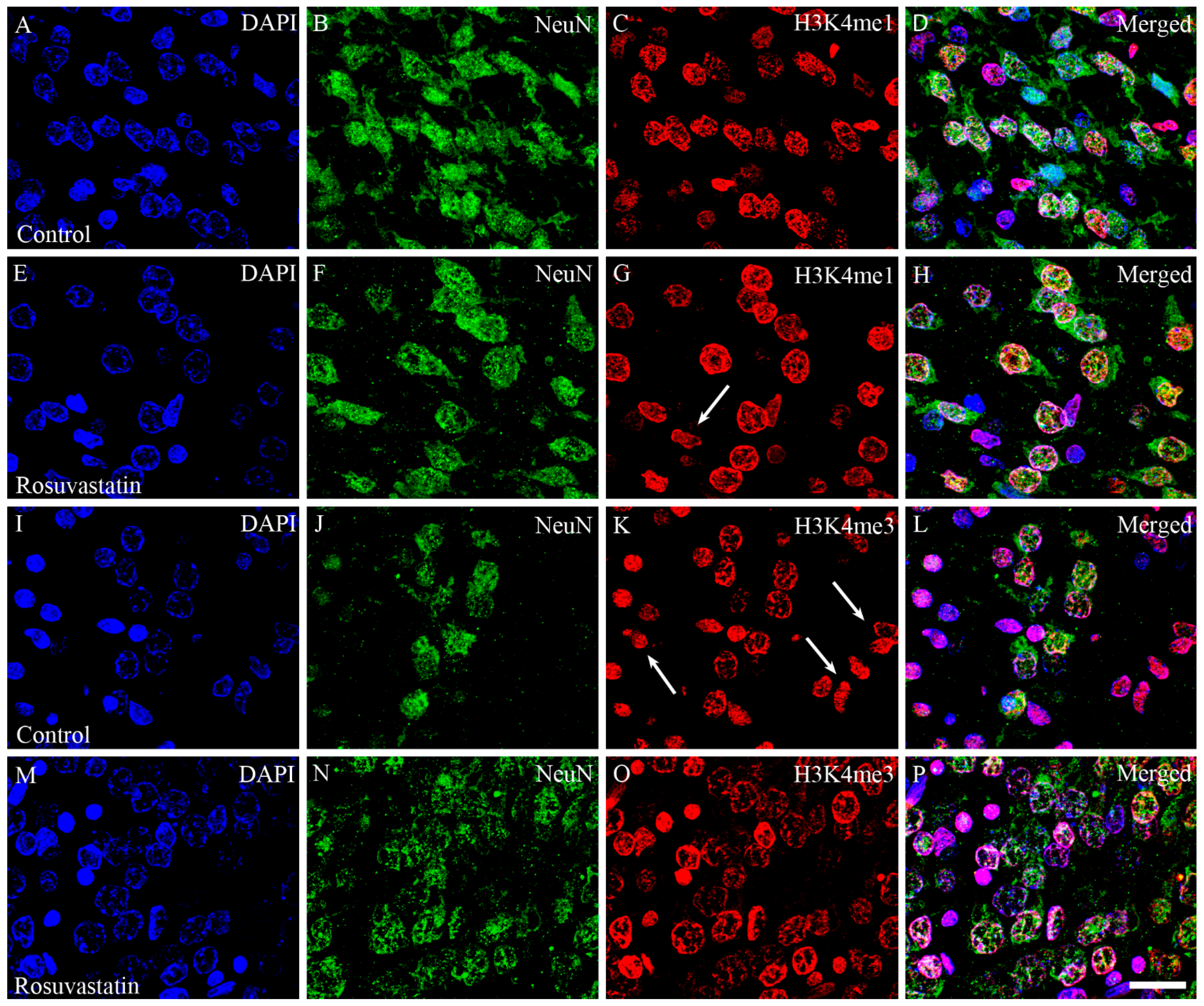
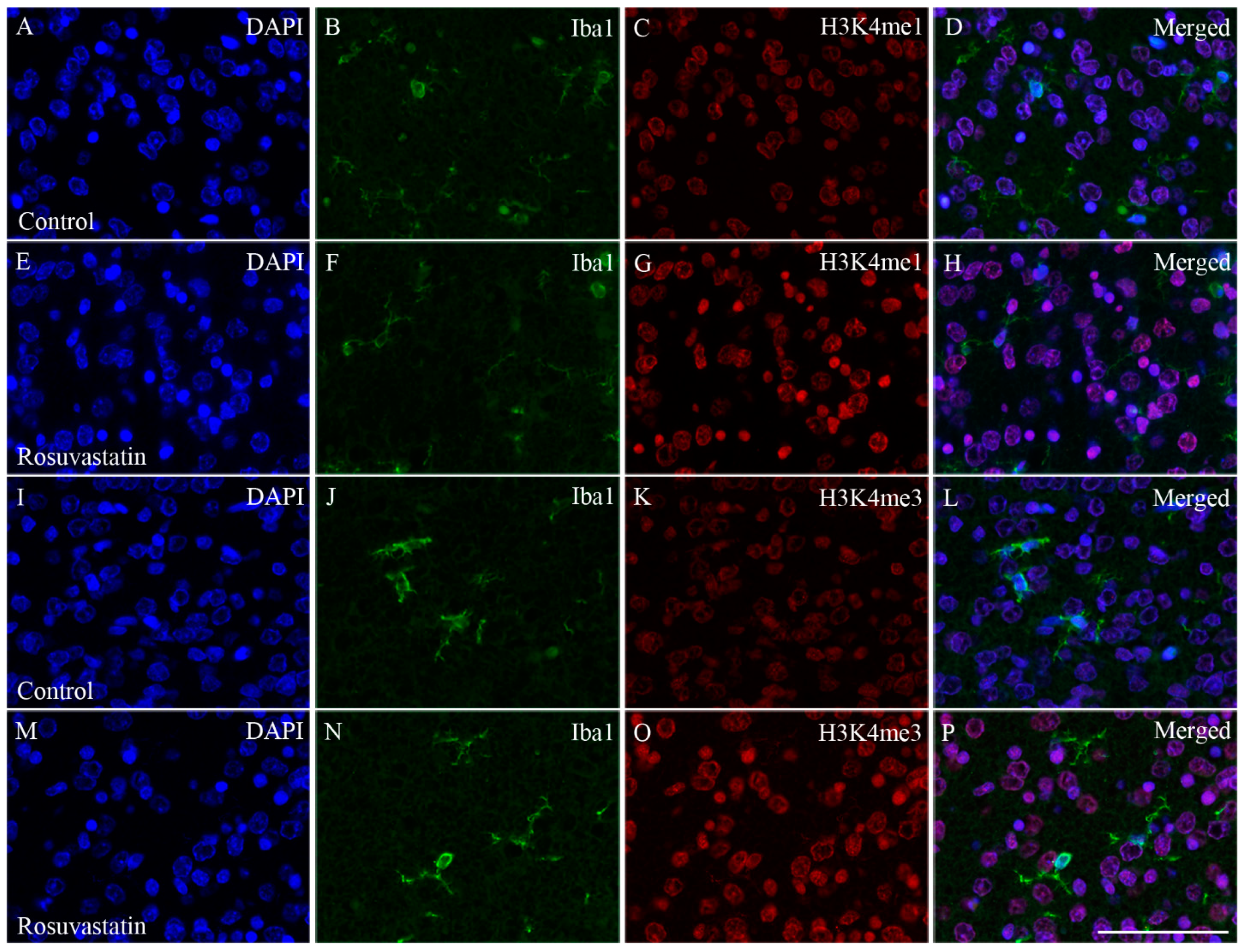
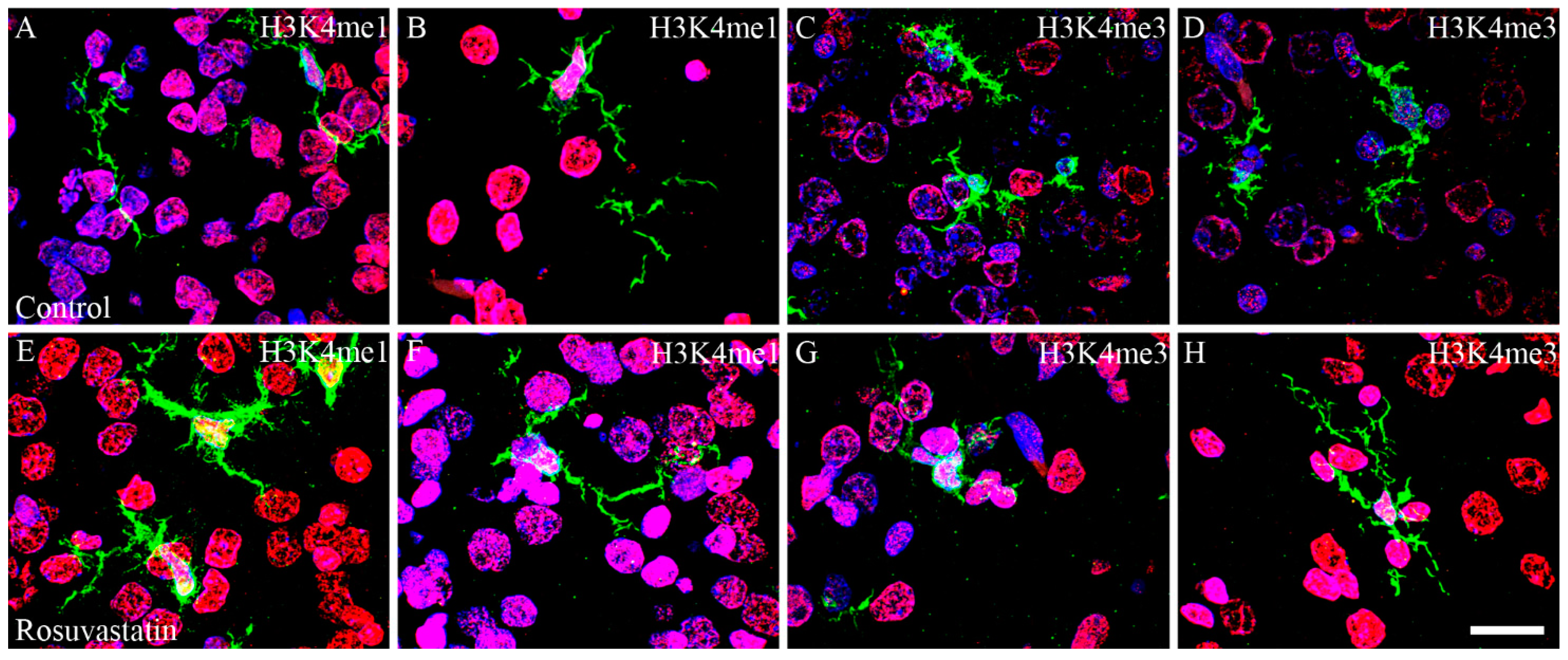
2.4. The Increase in H3K4me1 and H3K4me3 Is Localized Mainly to Neuronal Cell Nuclei
3. Discussion
4. Materials and Methods
4.1. Animal Handling and Treatment
4.2. Antibodies
4.3. Histology
4.4. Fluorescent and Confocal Immunohistochemistry
4.5. Western Blot Analyses
4.6. Imaging and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| CNS | central nervous system |
| CNPase | 2′,3′-cyclic nucleotide 3′-phosphodiesterase |
| DAPI | 2-[4-(aminoiminomethyl)phenyl]-1H-indole-6-carboximidamide hydrochloride |
| EAAT1 | excitatory amino acid transporter 1 (also known as glutamate aspartate transporter 1 (GLAST-1)) |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase (EC 1.2.1.12) |
| GFAP | glial fibrillary acidic protein |
| H | histone |
| HGM-CoA | 3-hydroxy-3-methylglutaryl coenzyme A |
| H&E | hematoxylin and eosin |
| Iba1 | ionized calcium-binding adaptor molecule 1 |
| IgG | immunoglobulin G |
| Ki67 | proliferation marker antigen identified by the monoclonal antibody Ki67 |
| NeuN | anti-neuronal nuclei protein (hexaribonucleotide-binding protein-3) |
| PBS | phosphate-buffered saline |
| RST | rosuvastatin |
| RT | room temperature |
| SDS | sodium dodecyl sulfate |
| SEM | standard error of the mean |
| TBS | Tris-buffered saline |
References
- Taylor, F.; Huffman, M.D.; Macedo, A.F.; Moore, T.H.; Burke, M.; Davey Smith, G.; Ward, K.; Ebrahim, S. Statins for the primary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2013, 2013, CD004816. [Google Scholar] [CrossRef]
- Burg, J.S.; Espenshade, P.J. Regulation of HMG-CoA reductase in mammals and yeast. Progr. Lipid Res. 2011, 50, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicki, A.S.; Poston, R.; Ferro, A. The lipid and non-lipid effects of statins. Pharmacol. Therap. 2003, 99, 95–112. [Google Scholar] [CrossRef]
- Famer, D.; Wahlund, L.O.; Crisby, M. Rosuvastatin reduces microglia in the brain of wild type and ApoE knockout mice on a high cholesterol diet; implications for prevention of stroke and AD. Biochem. Biophys. Res. Commun. 2010, 402, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Van der Most, P.J.; Dolga, A.M.; Nijholt, I.M.; Luiten, P.G.M.; Eisel, U.L. Statins: Mechanisms of neuroprotection. Progr. Neurobiol. 2009, 88, 64–75. [Google Scholar] [CrossRef]
- Zipp, F.; Waiczies, S.; Aktas, O.; Neuhaus, O.; Hemmer, B.; Schraven, B.; Nitsch, R.; Hartung, H.P. Impact of HMG-CoA reductase inhibition on brain pathology. Trends Pharmacol. Sci. 2007, 28, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Kata, D.; Földesi, I.; Feher, L.Z.; Hackler, L., Jr.; Puskas, L.G.; Gulya, K. Rosuvastatin enhances anti-inflammatory and inhibits pro-inflammatory functions in cultured microglial cells. Neuroscience 2016, 314, 47–63. [Google Scholar] [CrossRef]
- Kurata, T.; Miyazaki, K.; Kozuki, M.; Morimoto, N.; Ohta, Y.; Ikeda, Y.; Abe, K. Atorvastatin and pitavastatin reduce senile plaques and inflammatory responses in a mouse model of Alzheimer’s disease. Neurol. Res. 2012, 34, 601–610. [Google Scholar] [CrossRef]
- Zelcer, N.; Khanlou, N.; Clare, R.; Jiang, Q.; Reed-Geaghan, E.G.; Landreth, G.E.; Vinters, H.V.; Tontonoz, P. Attenuation of neuroinflammation and Alzheimer’s disease pathology by liver x receptors. Proc. Natl. Acad. Sci. USA 2007, 104, 10601–10606. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.D.; Couldwell, W.T.; Hinton, D.R.; Su, Y.; He, S.; Anker, L.; Law, R.E. Lovastatin induces growth inhibition and apoptosis in human malignant glioma cells. Biochem. Biophys. Res. Commun. 1994, 205, 1681–1687. [Google Scholar] [CrossRef]
- Karlic, H.; Haider, F.; Thaler, R.; Spitzer, S.; Klaushofer, K.; Varga, F. Statin and bisphosphonate induce starvation in fast-growing cancer cell lines. Int. J. Mol. Sci. 2017, 18, 1982. [Google Scholar] [CrossRef] [PubMed]
- Afshari, A.R.; Mollazadeh, H.; Henney, N.C.; Jamialahmad, T.; Sahebkar, A. Effects of statins on brain tumors: A review. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Gachpazan, M.; Kashani, H.; Khazaei, M.; Hassanian, S.M.; Rezayi, M.; Asgharzadeh, F.; Ghayour-Mobarhan, M.; Ferns, G.A.; Avan, A. The impact of statin therapy on the survival of patients with gastrointestinal cancer. Curr. Drug Targets 2019, 20, 738–747. [Google Scholar] [CrossRef]
- Kostapanos, M.S.; Milionis, H.J.; Elisaf, M.S. Rosuvastatin-associated adverse effects and drug-drug interactions in the clinical setting of dyslipidemia. Am. J. Cardiovasc. Drugs 2010, 10, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Thorogood, M.; Seed, M.; De Mott, K. Guideline Development Group. Management of fertility in women with familial hypercholesterolaemia: Summary of NICE guidance. BJOG 2009, 116, 478–479. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, L.M.; Erramouspe, J.; Cleveland, K.W. Teratogenic risk of statins in pregnancy. Ann. Pharmacother. 2012, 46, 1419–1424. [Google Scholar] [CrossRef]
- Lecarpentier, E.; Morel, O.; Fournier, T.; Elefant, E.; Chavatte-Palmer, P.; Tsatsaris, V. Statins and pregnancy: Between supposed risks and theoretical benefits. Drugs 2012, 72, 773–788. [Google Scholar] [CrossRef]
- Morton, S.; Thangaratinam, S. Statins in pregnancy. Curr. Opin. Obstet. Gynecol. 2013, 25, 433–440. [Google Scholar] [CrossRef]
- Taguchi, N.; Rubin, E.T.; Hosokawa, A.; Choi, J.; Ying, A.Y.; Moretti, M.E.; Koren, G.; Ito, S. Prenatal exposure to HMG-CoA reductase inhibitors: Effects on fetal and neonatal outcomes. Reprod. Toxicol. 2008, 26, 175–177. [Google Scholar] [CrossRef]
- Beverly, B.E.J.; Furr, J.R.; Lambright, C.S.; Wilson, V.S.; McIntyre, B.S.; Foster, P.M.D.; Travlos, G.; Earl Gray, L., Jr. In utero exposure to simvastatin reduces postnatal survival and permanently alters reproductive tract development in the Crl:CD(SD) male rat. Toxicol. Appl. Pharmacol. 2019, 365, 112–123. [Google Scholar] [CrossRef]
- Emami, F.; Asl, B.M.; Seydi, E.; Zargar, M.; Naserzadeh, P.; Pourahmad, J. Embryotoxic effects of atorvastatin on mouse fetus. Iran. J. Pharm. Sci. 2013, 9, 13–23. [Google Scholar]
- Henck, J.W.; Craft, W.R.; Black, A.; Colgin, J.; Anderson, J.A. Pre- and postnatal toxicity of the HMG-CoA reductase inhibitor atorvastatin in rats. Toxicol. Sci. 1998, 41, 88–99. [Google Scholar] [CrossRef] [PubMed]
- McTaggart, F.; Buckett, L.; Davidson, R.; Holdgate, G.; McCormick, A.; Schneck, D.; Smith, G.; Warwick, M. Preclinical and clinical pharmacology of rosuvastatin, a new 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitor. Am. J. Cardiol. 2001, 87, 28B–32B. [Google Scholar] [CrossRef]
- Asztalos, B.F.; Le Maulf, F.; Dallal, G.E.; Stein, E.; Jones, P.H.; Horvath, K.V.; McTaggart, F.; Schaefer, E.J. Comparison of the effects of high doses of rosuvastatin versus atorvastatin on the subpopulations of high-density lipoproteins. Am. J. Cardiol. 2007, 99, 681–685. [Google Scholar] [CrossRef]
- Lwin, E.M.P.; Leggett, C.; Ritchie, U.; Gerber, C.; Song, Y.; Hague, W.; Turner, S.; Upton, R.; Garg, S. Transfer of rosuvastatin into breast milk: Liquid chromatography-mass spectrometry methodology and clinical recommendations. Drug Des. Dev. Ther. 2018, 12, 3645–3651. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.; Li, B.; Wang, W.; Liu, X.; Liu, X.; Xia, Y.; Zhang, C.; Zhang, Y.; Zhang, M.; An, F. Rosuvastatin alleviates diabetic cardiomyopathy by inhibiting NLRP3 inflammasome and MAPK pathways in a type 2 diabetes rat model. Cardiovasc. Drugs Ther. 2014, 28, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Satoh, M.; Tabuchi, T.; Itoh, T.; Nakamura, M. NLRP3 inflammasome activation in coronary artery disease: Results from prospective and randomized study of treatment with atorvastatin or rosuvastatin. Clin. Sci. (Lond.) 2014, 126, 233–241. [Google Scholar] [CrossRef]
- Biswas, S.; Rao, C.M. Epigenetic tools (The Writers, The Readers and The Erasers) and their implications in cancer therapy. Eur. J. Pharmacol. 2018, 837, 8–24. [Google Scholar] [CrossRef] [PubMed]
- Hyun, K.; Jeon, J.; Park, K.; Kim, J. Writing, erasing and reading histone lysine methylations. Exp. Mol. Med. 2017, 49, e324. [Google Scholar] [CrossRef]
- Klein, B.J.; Krajewski, K.; Restrepo, S.; Lewis, P.W.; Strahl, B.D.; Kutateladze, T.G. Recognition of cancer mutations in histone H3K36 by epigenetic writers and readers. Epigenetics 2018, 13, 683–692. [Google Scholar] [CrossRef]
- Venkatesh, S.; Workman, J. Histone exchange, chromatin structure and the regulation of transcription. Nat. Rev. Mol. Cell. Biol. 2015, 16, 178–189. [Google Scholar] [CrossRef]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Trojer, P.; Reinberg, D. Histone lysine demethylases and their impact on epigenetics. Cell 2006, 125, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, M.; Eberl, H.C.; Matarese, F.; Marks, H.; Denissov, S.; Butter, F.; Lee, K.K.; Olsen, J.V.; Hyman, A.A.; Stunnenberg, H.G.; et al. Quantitative interaction proteomics and genome-wide profiling of epigenetic histone marks and their readers. Cell 2010, 142, 967–980. [Google Scholar] [CrossRef] [PubMed]
- Danielewicz, H.; Gurgul, A.; Dębińska, A.; Myszczyszyn, G.; Szmatoła, T.; Myszkal, A.; Jasielczuk, I.; Drabik-Chamerska, A.; Hirnle, L.; Boznański, A. Maternal atopy and offspring epigenome-wide methylation signature. Epigenetics 2020, 9, 1–13. [Google Scholar] [CrossRef]
- Monk, D.; Mackay, D.J.G.; Eggermann, T.; Maher, E.R.; Riccio, A. Genomic imprinting disorders: Lessons on how genome, epigenome and environment interact. Nat. Rev. Genet. 2019, 20, 235–248. [Google Scholar] [CrossRef]
- Elahi, M.M.; Cagampang, F.R.; Anthony, F.W.; Curzen, N.; Ohri, S.K.; Hanson, M.A. Statin treatment in hypercholesterolemic pregnant mice reduces cardiovascular risk factors in their offspring. Hypertension 2008, 51, 939–944. [Google Scholar] [CrossRef]
- Smith, D.D.; Costantine, M.M. The role of statins in the prevention of preeclampsia. Am. J. Obstet. Gynecol. 2020. [Google Scholar] [CrossRef]
- Faqi, A.S.; Prohaska, D.; Lopez, R.; McIntyre, G. Developmental toxicity of the HMG-CoA reductase inhibitor (PPD10558) in rats and rabbits. Birth Defects Res. B Dev. Reprod. Toxicol. 2012, 95, 23–37. [Google Scholar] [CrossRef]
- Morrow, T.; Song, M.R.; Ghosh, A. Sequential specification of neurons and glia by developmentally regulated extracellular factors. Development 2001, 128, 3585–3594. [Google Scholar] [PubMed]
- Bandeira, F.; Lent, R.; Herculano-Houzel, S. Changing numbers of neuronal and non-neuronal cells underlie postnatal brain growth in the rat. Proc. Natl. Acad. Sci. USA 2009, 106, 14108–14113. [Google Scholar] [CrossRef] [PubMed]
- Reemst, K.; Noctor, S.C.; Lucassen, P.J.; Hol, E.M. The indispensable roles of microglia and astrocytes during brain development. Front. Hum. Neurosci. 2016, 10, 566. [Google Scholar] [CrossRef] [PubMed]
- Sauvageot, C.M.; Stiles, C.D. Molecular mechanisms controlling cortical gliogenesis. Curr. Opin. Neurobiol. 2002, 12, 244–249. [Google Scholar] [CrossRef]
- Li, Y. Epigenetic mechanisms link maternal diets and gut microbiome to obesity in the offspring. Front. Genet. 2018, 9, 342. [Google Scholar] [CrossRef] [PubMed]
- Johnson, I.T. The cancer risk related to meat and meat products. Br. Med. Bull. 2017, 121, 73–81. [Google Scholar] [CrossRef]
- Khambadkone, S.G.; Cordner, Z.A.; Dickerson, F.; Severance, E.G.; Prandovszky, E.; Pletnikov, M.; Xiao, J.; Li, Y.; Boersma, G.J.; Talbot, C.C., Jr.; et al. Nitrated meat products are associated with mania in humans and altered behavior and brain gene expression in rats. Mol. Psychiatry 2020, 25, 560–571. [Google Scholar] [CrossRef]
- Shen, E.; Shulha, H.; Weng, Z.; Akbarian, S. Regulation of histone H3K4 methylation in brain development and disease. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130514. [Google Scholar] [CrossRef]
- Batista, I.A.A.; Helguero, L.A. Biological processes and signal transduction pathways regulated by the protein methyltransferase SETD7 and their significance in cancer. Signal Transduct. Target Ther. 2018, 3, 19. [Google Scholar] [CrossRef]
- Malik, S.; Bhaumik, S.R. Mixed lineage leukemia: Histone H3 lysine 4 methyltransferases from yeast to human. FEBS J. 2010, 277, 1805–1821. [Google Scholar] [CrossRef] [PubMed]
- Guenther, M.G.; Jenner, R.G.; Chevalier, B.; Nakamura, T.; Croce, C.M.; Canaani, E.; Young, R.A. Global and Hox-specific roles for the MLL1 methyltransferase. Proc. Natl. Acad. Sci. USA 2005, 102, 8603–8608. [Google Scholar] [CrossRef]
- Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.Y.; Schones, D.E.; Wang, Z.; Wei, G.; Chepelev, I.; Zhao, K. High-resolution profiling of histone methylations in the human genome. Cell 2007, 129, 823–837. [Google Scholar] [CrossRef]
- Jones, W.D.; Dafou, D.; McEntagart, M.; Woollard, W.J.; Elmslie, F.V.; Holder-Espinasse, M.; Irving, M.; Saggar, A.K.; Smithson, S.; Trembath, R.C.; et al. De novo mutations in MLL cause Wiedemann-Steiner syndrome. Am. J. Hum. Genet. 2012, 91, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Najmabadi, H.; Hu, H.; Garshasbi, M.; Zemojtel, T.; Abedini, S.S.; Chen, W.; Hosseini, M.; Behjati, F.; Haas, S.; Jamali, P.; et al. Deep sequencing reveals 50 novel genes for recessive cognitive disorders. Nature 2011, 478, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Kim, S.Y.; Artis, S.; Molfese, D.L.; Schumacher, A.; Sweatt, J.D.; Paylor, R.E.; Lubin, F.D. Histone methylation regulates memory formation. J. Neurosci. 2010, 30, 3589–3599. [Google Scholar] [CrossRef]
- Connor, C.M.; Dincer, A.; Straubhaar, J.; Galler, J.R.; Houston, I.B.; Akbarian, S. Maternal immune activation alters behavior in adult offspring, with subtle changes in the cortical transcriptome and epigenome. Schizophr. Res. 2012, 140, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Maćkowiak, M.; Bator, E.; Latusz, J.; Mordalska, P.; Wędzony, K. Prenatal MAM administration affects histone H3 methylation in postnatal life in the rat medial prefrontal cortex. Eur. Neuropsychopharmacol. 2014, 24, 271–289. [Google Scholar] [CrossRef]
- Jie, L.; Ahn, J.H.; Wang, G.G. Understanding histone H3 lysine 36 methylation and its deregulation in disease. Cell. Mol. Life Sci. 2019, 76, 2899–2916. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, Y.; Jia, J.; Fang, Y.; Tang, Y.; Wu, H.; Fang, D. H3K36me3, message from chromatin to DNA damage repair. Cell. Biosci. 2020, 10, 9. [Google Scholar] [CrossRef]
- Maleszewska, M.; Steranka, A.; Smiech, M.; Kaza, B.; Pilanc, P.; Dabrowski, M.; Kaminska, B. Sequential changes in histone modifications shape transcriptional responses underlying microglia polarization by glioma. Glia 2021, 69, 109–123. [Google Scholar] [CrossRef]
- Bannister, A.J.; Zegerman, P.; Partridge, J.F.; Miska, E.A.; Thomas, J.O.; Allshire, R.C.; Kouzarides, T. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature 2001, 410, 120–124. [Google Scholar] [CrossRef]
- Saksouk, N.; Simboeck, E.; Déjardin, J. Constitutive heterochromatin formation and transcription in mammals. Epigenetics Chromatin 2015, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Bridgeman, S.; Northrop, W.; Ellison, G.; Sabapathy, T.; Melton, P.E.; Newsholme, P.; Mamotte, C.D.S. Statins do not directly inhibit the activity of major epigenetic modifying enzymes. Cancers 2019, 11, 516. [Google Scholar] [CrossRef]
- Koren, G.; Pastuszak, A.; Ito, S. Drugs in pregnancy. N. Engl. J. Med. 1998, 338, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Cuylen, S.; Blaukopf, C.; Politi, A.Z.; Müller-Reichert, T.; Neumann, B.; Poser, I.; Ellenberg, J.; Hyman, A.A.; Gerlich, D.W. Ki-67 acts as a biological surfactant to disperse mitotic chromosomes. Nature 2016, 535, 308–312. [Google Scholar] [CrossRef]
- Szabo, M.; Gulya, K. Development of the microglial phenotype in culture. Neuroscience 2013, 241, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dulka, K.; Szabo, M.; Lajkó, N.; Belecz, I.; Hoyk, Z.; Gulya, K. Epigenetic Consequences of in Utero Exposure to Rosuvastatin: Alteration of Histone Methylation Patterns in Newborn Rat Brains. Int. J. Mol. Sci. 2021, 22, 3412. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073412
Dulka K, Szabo M, Lajkó N, Belecz I, Hoyk Z, Gulya K. Epigenetic Consequences of in Utero Exposure to Rosuvastatin: Alteration of Histone Methylation Patterns in Newborn Rat Brains. International Journal of Molecular Sciences. 2021; 22(7):3412. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073412
Chicago/Turabian StyleDulka, Karolina, Melinda Szabo, Noémi Lajkó, István Belecz, Zsófia Hoyk, and Karoly Gulya. 2021. "Epigenetic Consequences of in Utero Exposure to Rosuvastatin: Alteration of Histone Methylation Patterns in Newborn Rat Brains" International Journal of Molecular Sciences 22, no. 7: 3412. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073412