
Sphingosine-1-Phosphate Receptor Type 4 (S1P4) Is Differentially Regulated in Peritoneal B1 B Cells upon TLR4 Stimulation and Facilitates the Egress of Peritoneal B1a B Cells and Subsequent Accumulation of Splenic IRA B Cells under Inflammatory Conditions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. S1P Receptor Expression Is Differentially Regulated during Peritoneal B Cell Activation In Vitro
2.2. Peritoneal B Cell Activation via TLR4 Leads to Reduced Chemotactic Response to S1P In Vitro
2.3. Impact of S1P4 Deficiency on the Trafficking of Adoptively Transferred Peritoneal B Cells in Sterile LPS-Induced Peritonitis
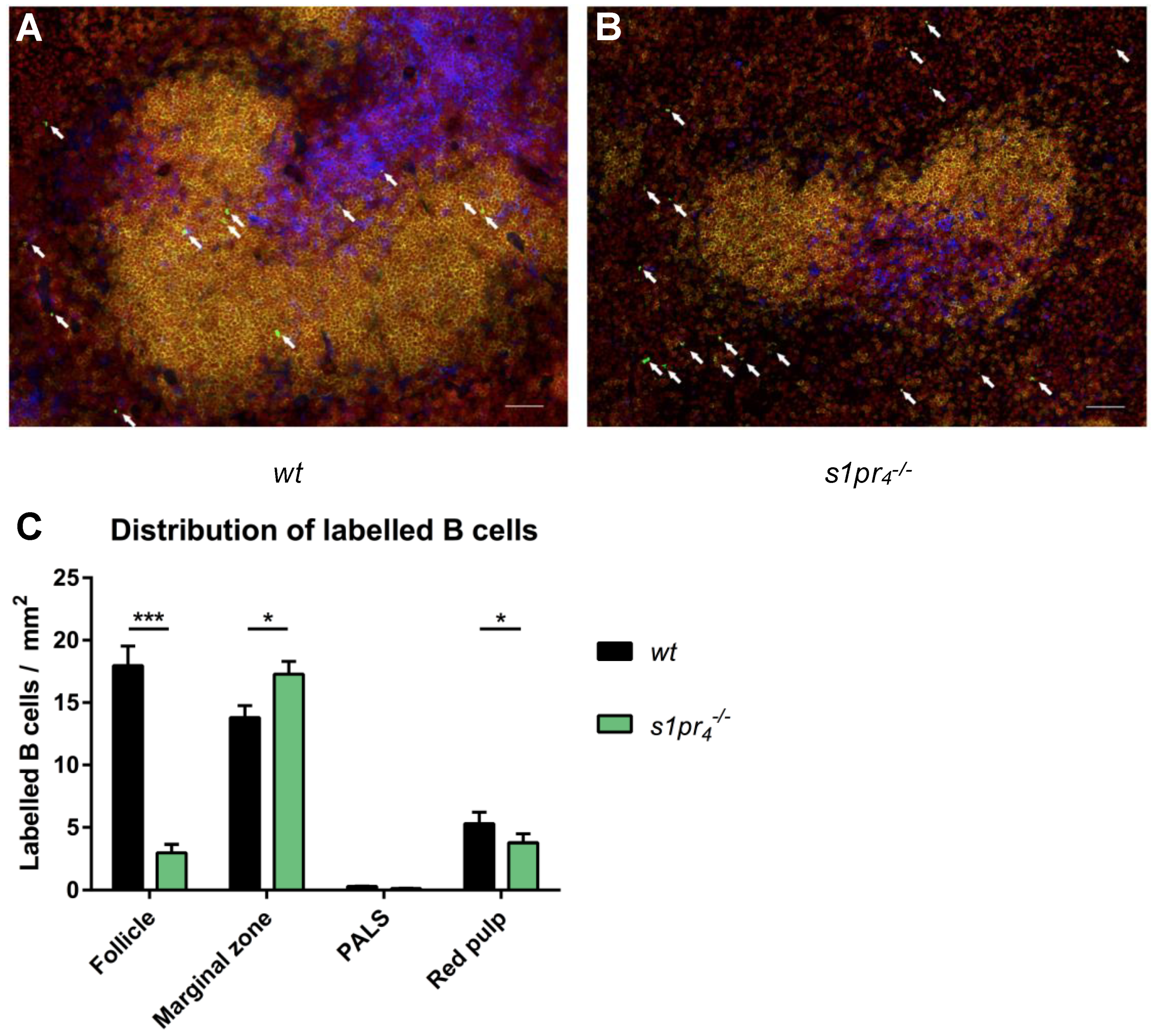
2.4. Impact of S1P4 on the Localization of Migrating Peritoneal B Cells within the Spleen, Lymph Nodes, and Peyer’s Patches after LPS Stimulation
2.5. Impact of S1P4 Expression on the Number of Splenic IRA B Cells after Peritoneal LPS Stimulation
2.6. Influence of S1P4 Expression on the Composition of Peritoneal B Cell Populations and Cytokine Levels in A Murine Model of Polymicrobial Abdominal Sepsis
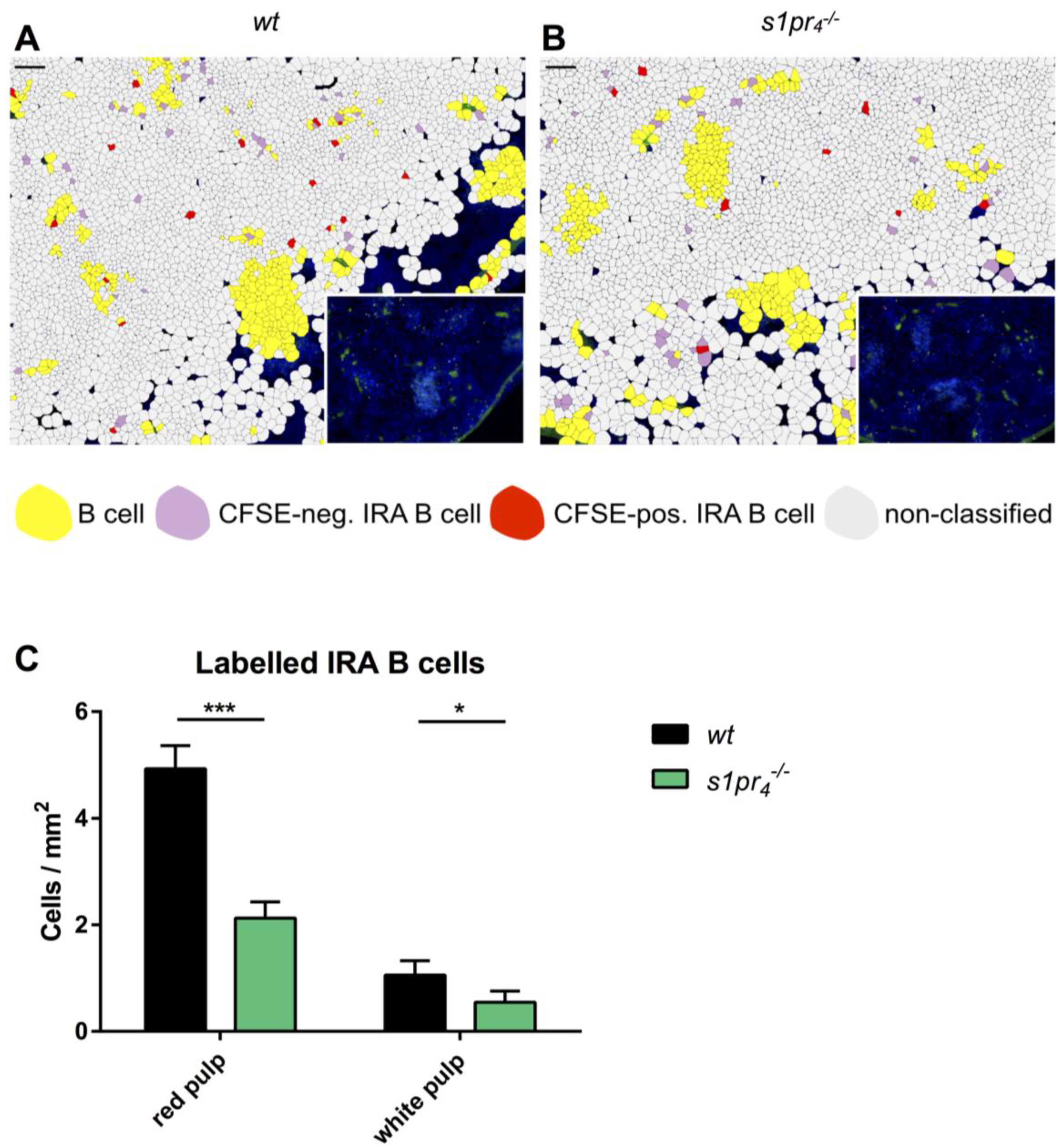
2.7. S1P4 Deficiency Results in a Significant Reduction of IRA B Cells in the Spleens of CASP Mice
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Cell Isolation from the Peritoneal Cavity
4.3. Flow Cytometry
4.4. In Vitro Activation of Peritoneal B Cell Subpopulations
4.5. RT-PCR of S1P Receptors in Peritoneal B Cells after In Vitro Stimulation
4.6. Transwell Migration Assay of Peritoneal B Cells after LPS Stimulation
4.7. Adoptive Cell Transfer under LPS Stimulation
4.8. Colon Ascendens Stent Peritonitis (CASP)
4.9. Cytokine Analysis
4.10. Fluorescence Microscopy of Splenic Tissue Sections
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CASP | colon ascendens stent peritonitis |
| CFSE | carboxyfluorescein-succinimidyl-ester |
| DAPI | 4′,6-diamidino-2-phenylindole |
| FACS | fluorescence-activated cell sorting |
| i.p. | intraperitoneal |
| IRA B cell | innate response activator B cell |
| LPS | lipopolysaccharide |
| PALS | periarteriolar lymphoid sheaths |
| PerC | peritoneal cavity |
| PMA | phorbol-12-myristat-13-acetat |
| PP | Peyer’s patch |
| RT-PCR | real-time polymerase chain reaction |
| SED | subepithelial dome |
| SEM | standard error of mean |
| SLO | secondary lymphoid organs |
| S1P | sphingosine-1-phosphate |
| S1PR | S1P receptor |
| s1pr4−/− | mouse homozygous deficient in S1P4 |
| TLR | toll-like receptor |
| wt | wild-type (C57BL/6J) mouse |
References
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.L.; Sakr, Y.; Singer, M.; Martin-Loeches, I.; Machado, F.R.; Marshall, J.C.; Finfer, S.; Pelosi, P.; Brazzi, L.; Aditianingsih, D.; et al. Prevalence and Outcomes of Infection Among Patients in Intensive Care Units in 2017. JAMA 2020, 323, 1478–1487. [Google Scholar] [CrossRef] [PubMed]
- Blot, S.; Antonelli, M.; Arvaniti, K.; Blot, K.; Creagh-Brown, B.; de Lange, D.; De Waele, J.; Deschepper, M.; Dikmen, Y.; Dimopoulos, G.; et al. Epidemiology of intra-abdominal infection and sepsis in critically ill patients: "AbSeS", a multinational observational cohort study and ESICM Trials Group Project. Intensive Care Med. 2019, 45, 1703–1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurin, L.P.; Brissette, M.J.; Lepage, S.; Cailhier, J.F. Regulation of experimental peritonitis: A complex orchestration. Nephron. Exp. Nephrol. 2012, 120, e41–e46. [Google Scholar] [CrossRef] [PubMed]
- Enderes, J.; van der Linde, J.; Muller, J.; Tran, B.T.; von Bernstorff, W.; Heidecke, C.D.; Schulze, T. FTY720-Induced Lymphopenia Does Not Aggravate Mortality in a Murine Model of Polymicrobial Abdominal Sepsis. Shock 2017, 47, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Baumgarth, N.; Waffarn, E.E.; Nguyen, T.T. Natural and induced B-1 cell immunity to infections raises questions of nature versus nurture. Ann. N. Y. Acad. Sci. 2015, 2015, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Barbeiro, D.F.; Barbeiro, H.V.; Faintuch, J.; Ariga, S.K.; Mariano, M.; Popi, A.F.; de Souza, H.P.; Velasco, I.T. Soriano FG. B-1 cells temper endotoxemic inflammatory responses. Immunobiology 2011, 216, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Minguet, S.; Dopfer, E.P.; Pollmer, C.; Freudenberg, M.A.; Galanos, C.; Reth, M.; Huber, M.; Schamel, W.W. Enhanced B-cell activation mediated by signals through Toll-like receptors. J. Exp. Med. 2006, 203, 2541–2550. [Google Scholar]
- Ha, S.A.; Tsuji, M.; Suzuki, K.; Meek, B.; Yasuda, N.; Kaisho, T.; Fagarasan, S. Regulation of B1 cell migration by signals through Toll-like receptors. J. Exp. Med. 2006, 203, 2541–2550. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Ohdan, H.; Zhao, G.; Yang, Y.G.; Sykes, M. Peritoneal cavity B cells are precursors of splenic IgM natural antibody-producing cells. J. Immunol. 2003, 171, 5406–5414. [Google Scholar] [CrossRef]
- Yang, Y.; Tung, J.W.; Ghosn, E.E.; Herzenberg, L.A. Division and differentiation of natural antibody-producing cells in mouse spleen. Proc. Natl. Acad. Sci. USA 2007, 104, 4542–4546. [Google Scholar] [CrossRef] [Green Version]
- Rauch, P.J.; Chudnovskiy, A.; Robbins, C.S.; Weber, G.F.; Etzrodt, M.; Hilgendorf, I.; Tiglao, E.; Figueiredo, J.L.; Iwamoto, Y.; Theurl, I.; et al. Innate response activator B cells protect against microbial sepsis. Science 2012, 335, 597–601. [Google Scholar] [CrossRef] [Green Version]
- Weber, G.F.; Chousterman, B.G.; He, S.; Fenn, A.M.; Nairz, M.; Anzai, A.; Brenner, T.; Uhle, F.; Iwamoto, Y.; Robbins, C.S.; et al. Interleukin-3 amplifies acute inflammation and is a potential therapeutic target in sepsis. Science 2015, 347, 1260–1265. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.; Lee, J.G.; Shin, S.H.; Kim, T.J. LPS-induced migration of peritoneal B-1 cells is associated with upregulation of CXCR4 and increased migratory sensitivity to CXCL12. J. Korean Med. Sci. 2012, 27, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Höpken, U.E.; Winter, S.; Achtman, A.H.; Krüger, K.; Lipp, M. CCR7 regulates lymphocyte egress and recirculation through body cavities. J. Leukoc. Biol. 2010, 87, 671–682. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Blaho, V.A.; Hla, T. An update on the biology of sphingosine 1-phosphate receptors. J. Lipid. Res. 2014, 55, 1596–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, T.K.; Bian, J.; Gill, D.L. Sphingosine 1-phosphate generated in the endoplasmic reticulum membrane activates release of stored calcium. J. Biol. Chem. 1994, 269, 22628–22635. [Google Scholar] [CrossRef]
- Cuvillier, O.; Pirianov, G.; Kleuser, B.; Vanek, P.G.; Coso, O.A.; Gutkind, S.; Spiegel, S. Suppression of ceramide-mediated programmed cell death by sphingosine-1-phosphate. Nature 1996, 381, 800–803. [Google Scholar] [CrossRef]
- Chun, J.; Hla, T.; Lynch, K.R.; Spiegel, S.; Moolenaar, W.H. International Union of Basic and Clinical Pharmacology. LXXVIII. Lysophospholipid receptor nomenclature. Pharmacol. Rev. 2010, 62, 579–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cinamon, G.; Matloubian, M.; Lesneski, M.J.; Xu, Y.; Low, C.; Lu, T.; Proia, R.L.; Cyster, J.G. Sphingosine 1-phosphate receptor 1 promotes B cell localization in the splenic marginal zone. Nat. Immunol. 2004, 5, 713–720. [Google Scholar] [CrossRef]
- Kabashima, K.; Haynes, N.M.; Xu, Y.; Nutt, S.L.; Allende, M.L.; Proia, R.L.; Cyster, J.G. Plasma cell S1P1 expression determines secondary lymphoid organ retention versus bone marrow tropism. J. Exp. Med. 2006, 203, 2683–2690. [Google Scholar] [CrossRef]
- Kleinwort, A.; Luhrs, F.; Heidecke, C.D.; Lipp, M.; Schulze, T. S1P Signalling Differentially Affects Migration of Peritoneal B Cell Populations In Vitro and Influences the Production of Intestinal IgA In Vivo. Int. J. Mol. Sci. 2018, 19, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunisawa, J.; Kurashima, Y.; Gohda, M.; Higuchi, M.; Ishikawa, I.; Miura, F.; Ogahara, I.; Kiyono, H. Sphingosine 1-phosphate regulates peritoneal B-cell trafficking for subsequent intestinal IgA production. Blood. 2007, 109, 3749–3756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graeler, M.; Goetzl, J. Activation-regulated expression and chemotctic function of shingosine 1-phosphate receptors in mouse splenic T cells. FASEB J. 2002, 16, 1874–1878. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; von Bernstorff, W.; Heidecke, C.D.; Schulze, T. Differential S1P Receptor Profiles on M1- and M2-Polarized Macrophages Affect Macrophage Cytokine Production and Migration. Biomed. Res. Int. 2017, 2017, 7584621. [Google Scholar] [CrossRef] [Green Version]
- Wittebole, X.; Castanares-Zapatero, D.; Laterre, P.F. Toll-like receptor 4 modulation as a strategy to treat sepsis. Mediat. Inflamm. 2010, 2010, 14. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B.; Rietschel, E.T. Innate immune sensing and its roots: The story of endotoxin. Nat. Rev. Immunol. 2003, 3, 169–176. [Google Scholar] [CrossRef]
- Butcher, E.C.; Picker, L.J. Lymphocyte homing and homeostasis. Science 1996, 272, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Ansel, K.M.; Harris, R.B.; Cyster, J.G. CXCL13 is required for B1 cell homing, natural antibody production, and body cavity immunity. Immunity 2002, 16, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Cyster, J.G. Chemokines, sphingosine-1-phosphate, and cell migration in secondary lymphoid organs. Annu. Rev. Immunol. 2005, 23, 127–159. [Google Scholar] [CrossRef]
- Matloubian, M.; Lo, C.G.; Cinamon, G.; Lesneski, M.J.; Xu, Y.; Brinkmann, V.; Allende, M.L.; Proia, R.L.; Cyster, J.G. Lymphocyte egress from thymus and peripheral lymphoid organs is dependent on S1P receptor 1. Nature 2004, 427, 355–360. [Google Scholar] [CrossRef]
- Gohda, M.; Kunisawa, J.; Miura, F.; Kagiyama, Y.; Kurashima, Y.; Higuchi, M.; Ishikawa, I.; Ogahara, I.; Kiyono, H. Sphingosine 1-Phosphate Regulates the Egress of IgA Plasmablasts from Peyer’s Patches for Intestinal IgA Responses. J. Immunol. 2008, 180, 5335–5343. [Google Scholar] [CrossRef] [Green Version]
- Martin, F.; Kearney, J.F. Marginal-zone B cells. Nat. Rev. Immunol. 2002, 2, 323–335. [Google Scholar] [CrossRef]
- Berberich, S.; Förster, R.; Pabst, O. The peritoneal micromilieu commits B cells to home to body cavities and the small intestine. Blood 2007, 109, 4627–4634. [Google Scholar] [CrossRef] [Green Version]
- Schulze, T.; Golfier, S.; Tabeling, C.; Rabel, K.; Graler, M.H.; Witzenrath, M.; Lipp, M. Sphingosine-1-phospate receptor 4 (S1P4) deficiency profoundly affects dendritic cell function and TH17-cell differentiation in a murine model. FASEB J. 2011, 25, 4024–4036. [Google Scholar] [CrossRef]
- Aziz, M.; Holodick, N.E.; Rothstein, T.L.; Wang, P. B-1a Cells Protect Mice from Sepsis: Critical Role of CREB. J. Immunol. 2017, 199, 750–760. [Google Scholar] [CrossRef]
- Weber, G.F.; Chousterman, B.G.; Hilgendorf, I.; Robbins, C.S.; Theurl, I.; Gerhardt, L.M.; Iwamoto, Y.; Quach, T.D.; Ali, M.; Chen, J.W. Pleural innate response activator B cells protect against pneumonia via a GM-CSF-IgM axis. J. Exp. Med. 2014, 211, 1243–1256. [Google Scholar] [CrossRef] [PubMed]
- Cyster, J.G.; Ngo, V.N.; Ekland, E.H.; Gunn, M.D.; Sedgwick, J.D.; Ansel, K.M. Chemokines and B-cell homing to follicles. Curr Top. Microbiol. Immunol. 1999, 246, 87–92. [Google Scholar] [PubMed]
- Cinamon, G.; Zachariah, M.A.; Lam, O.M.; Foss, F.W., Jr.; Cyster, J.G. Follicular shuttling of marginal zone B cells facilitates antigen transport. Nat. Immunol. 2008, 9, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabroe, I.; Jones, E.C.; Whyte, M.K.; Dower, S.K. Regulation of human neutrophil chemokine receptor expression and function by activation of Toll-like receptors 2 and 4. Immunology 2005, 115, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3-New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Maier, S.; Traeger, T.; Entleutner, M.; Westerholt, A.; Kleist, B.; Huser, N.; Holzmann, B.; Stier, A.; Pfeffer, K.; Heidecke, C.D. Cecal ligation and puncture versus colon ascendens stent peritonitis: Two distinct animal models for polymicrobial sepsis. Shock 2004, 21, 505–511. [Google Scholar] [CrossRef]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riese, J.; Gromann, A.; Lührs, F.; Kleinwort, A.; Schulze, T. Sphingosine-1-Phosphate Receptor Type 4 (S1P4) Is Differentially Regulated in Peritoneal B1 B Cells upon TLR4 Stimulation and Facilitates the Egress of Peritoneal B1a B Cells and Subsequent Accumulation of Splenic IRA B Cells under Inflammatory Conditions. Int. J. Mol. Sci. 2021, 22, 3465. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073465
Riese J, Gromann A, Lührs F, Kleinwort A, Schulze T. Sphingosine-1-Phosphate Receptor Type 4 (S1P4) Is Differentially Regulated in Peritoneal B1 B Cells upon TLR4 Stimulation and Facilitates the Egress of Peritoneal B1a B Cells and Subsequent Accumulation of Splenic IRA B Cells under Inflammatory Conditions. International Journal of Molecular Sciences. 2021; 22(7):3465. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073465
Chicago/Turabian StyleRiese, Janik, Alina Gromann, Felix Lührs, Annabel Kleinwort, and Tobias Schulze. 2021. "Sphingosine-1-Phosphate Receptor Type 4 (S1P4) Is Differentially Regulated in Peritoneal B1 B Cells upon TLR4 Stimulation and Facilitates the Egress of Peritoneal B1a B Cells and Subsequent Accumulation of Splenic IRA B Cells under Inflammatory Conditions" International Journal of Molecular Sciences 22, no. 7: 3465. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073465