
Newly Isolated Animal Pathogen Corynebacterium silvaticum Is Cytotoxic to Human Epithelial Cells


Abstract
:1. Introduction
2. Results
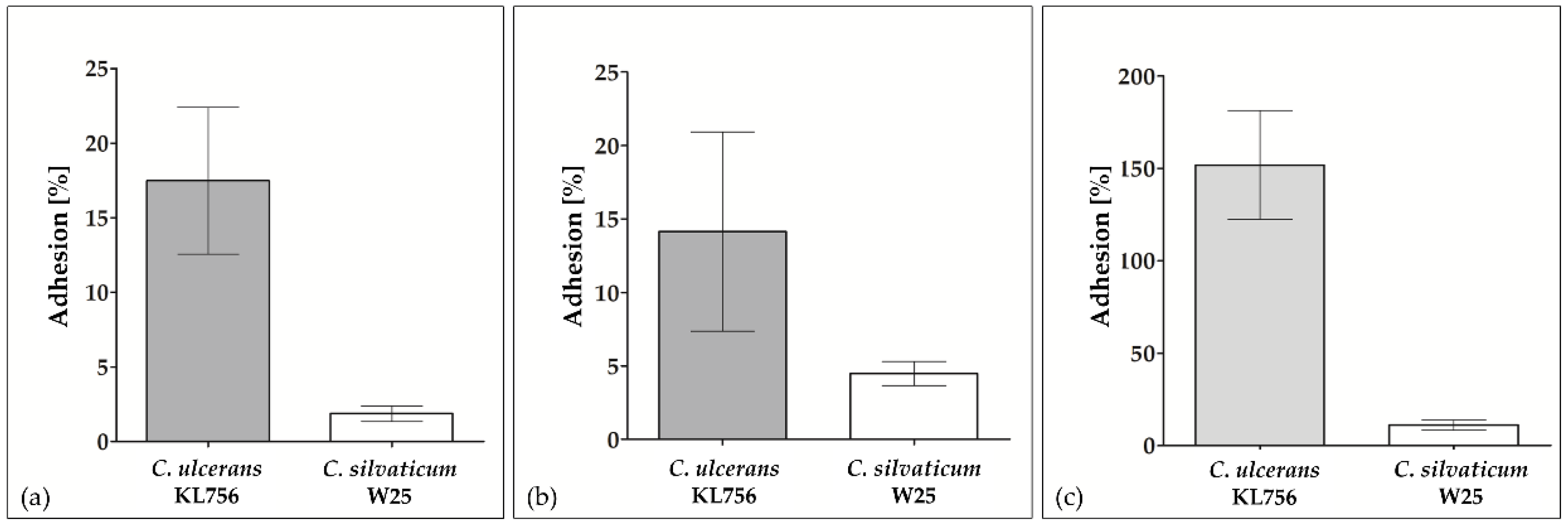
2.1. In Vitro Colonization of Epithelial Cells by W25
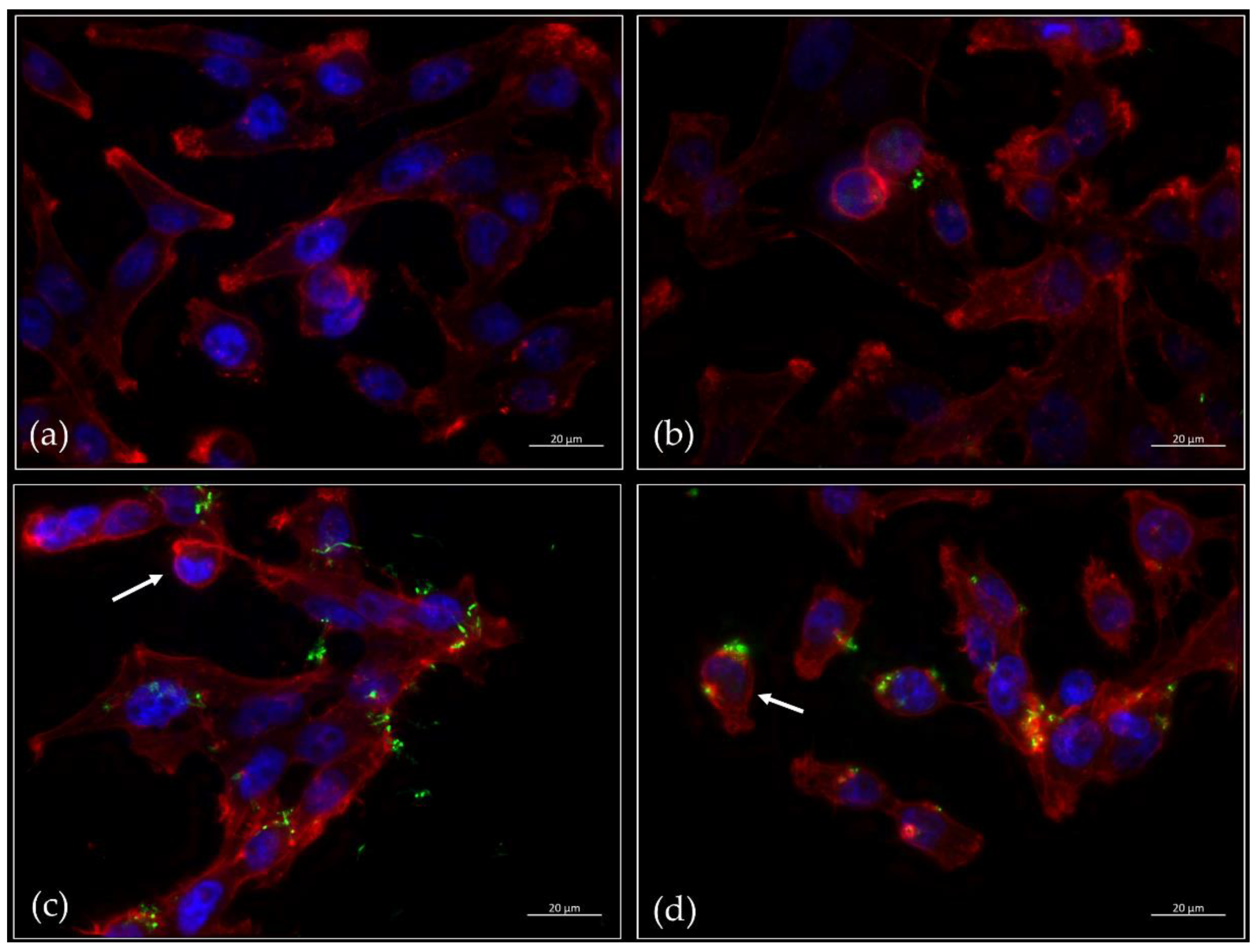
2.2. Microscopic Inspection of Infected Cells
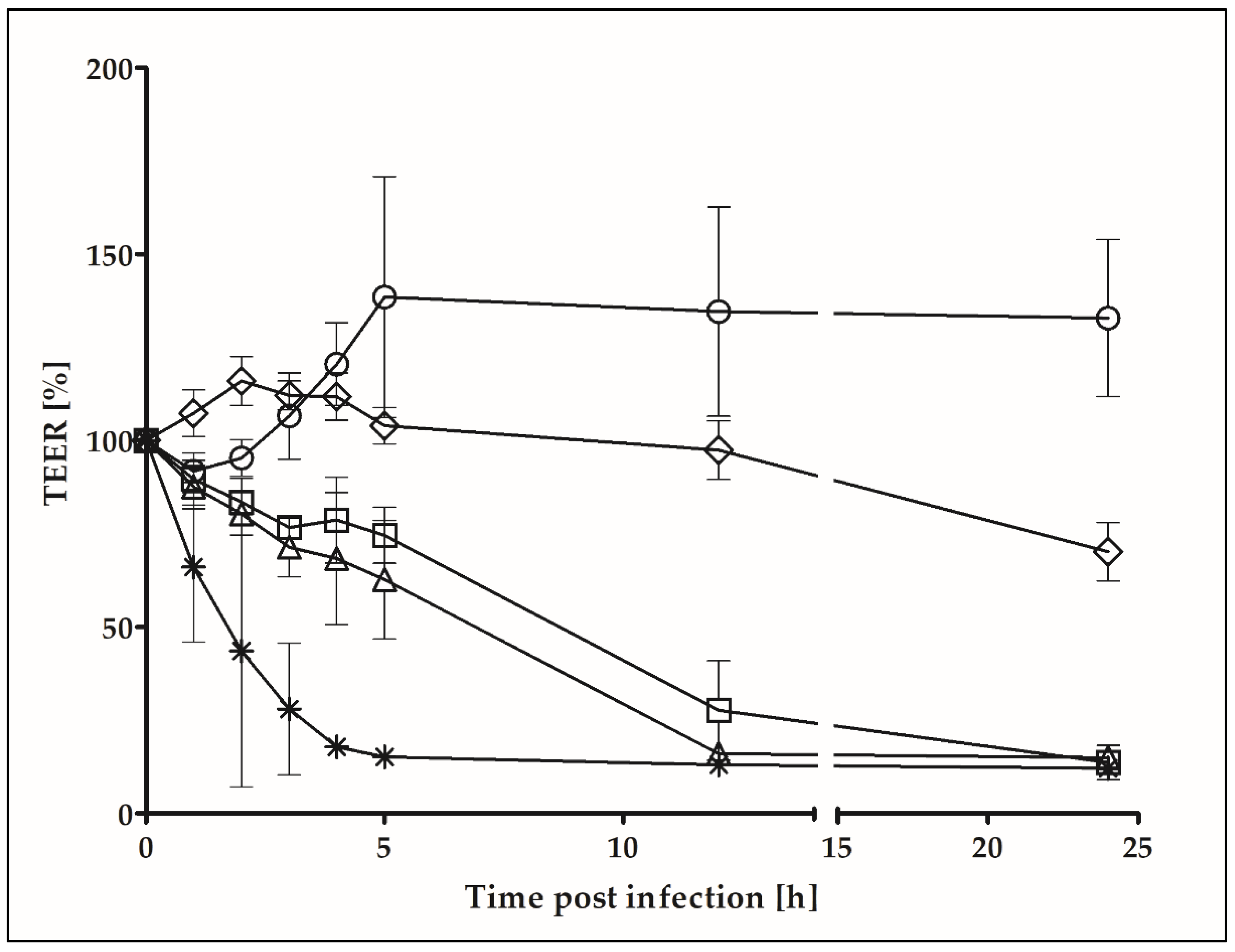
2.3. Transepithelial Electrical Resistance
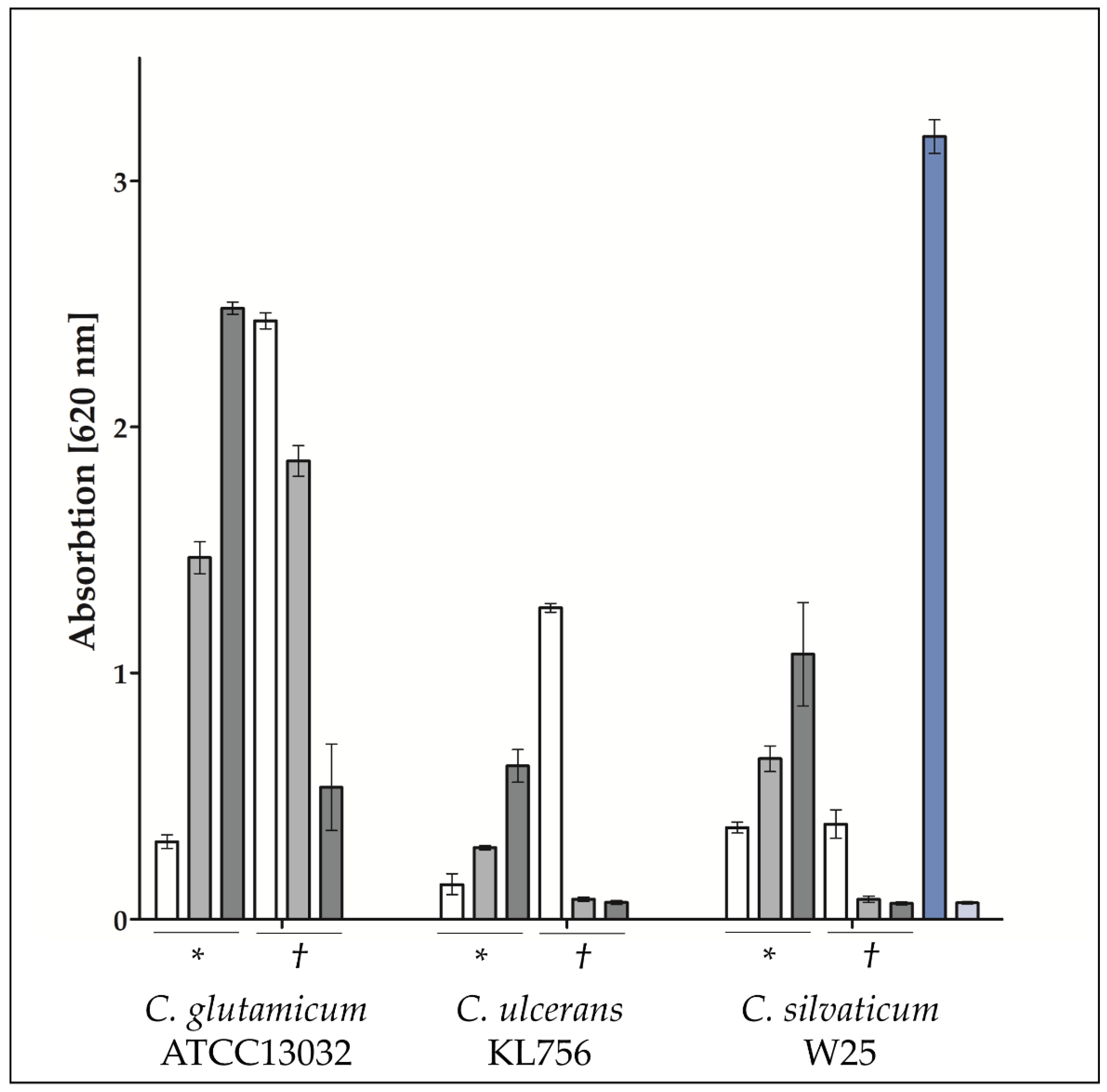
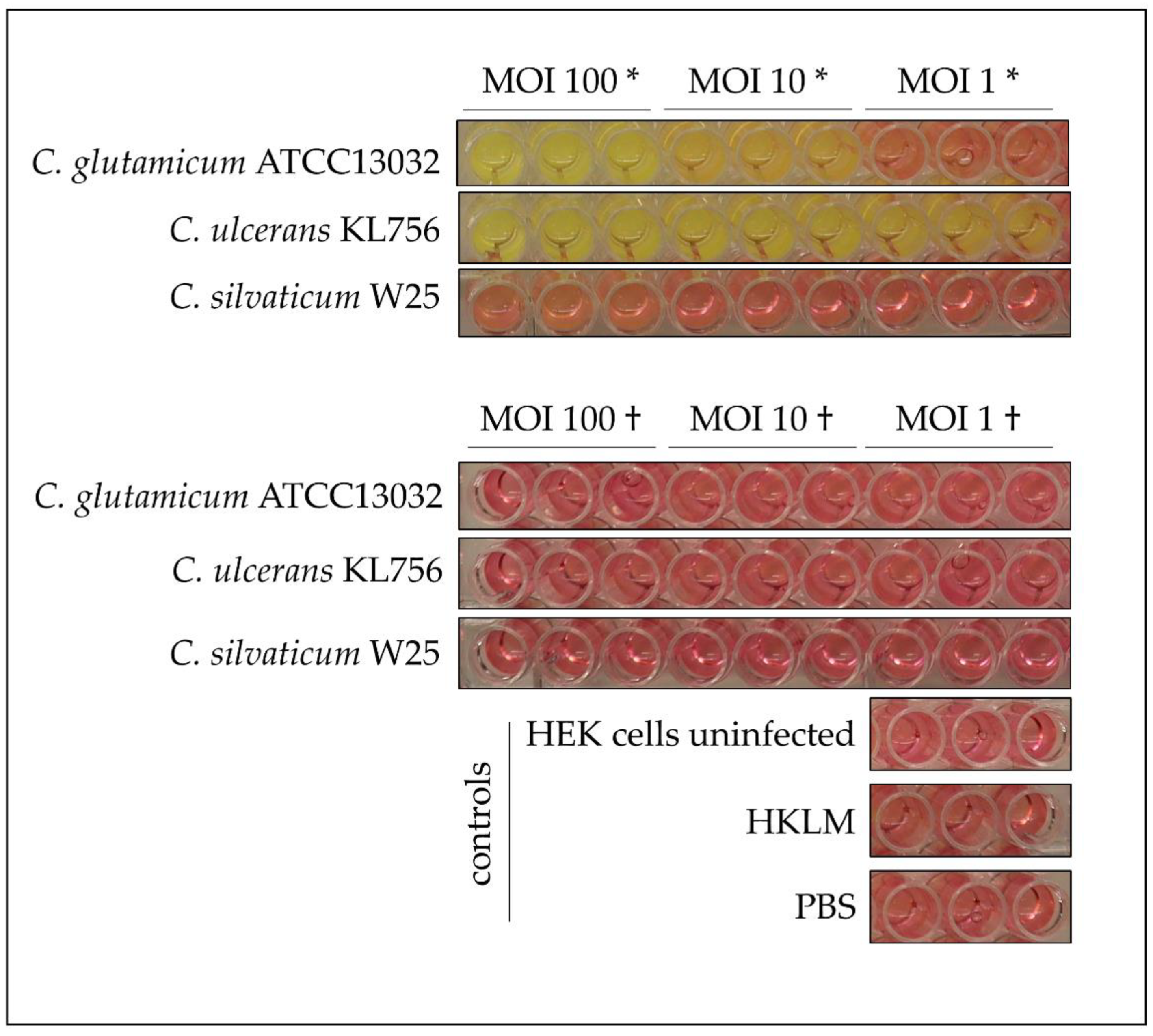
2.4. Recognition of Infection with Corynebacteria by Host Cells
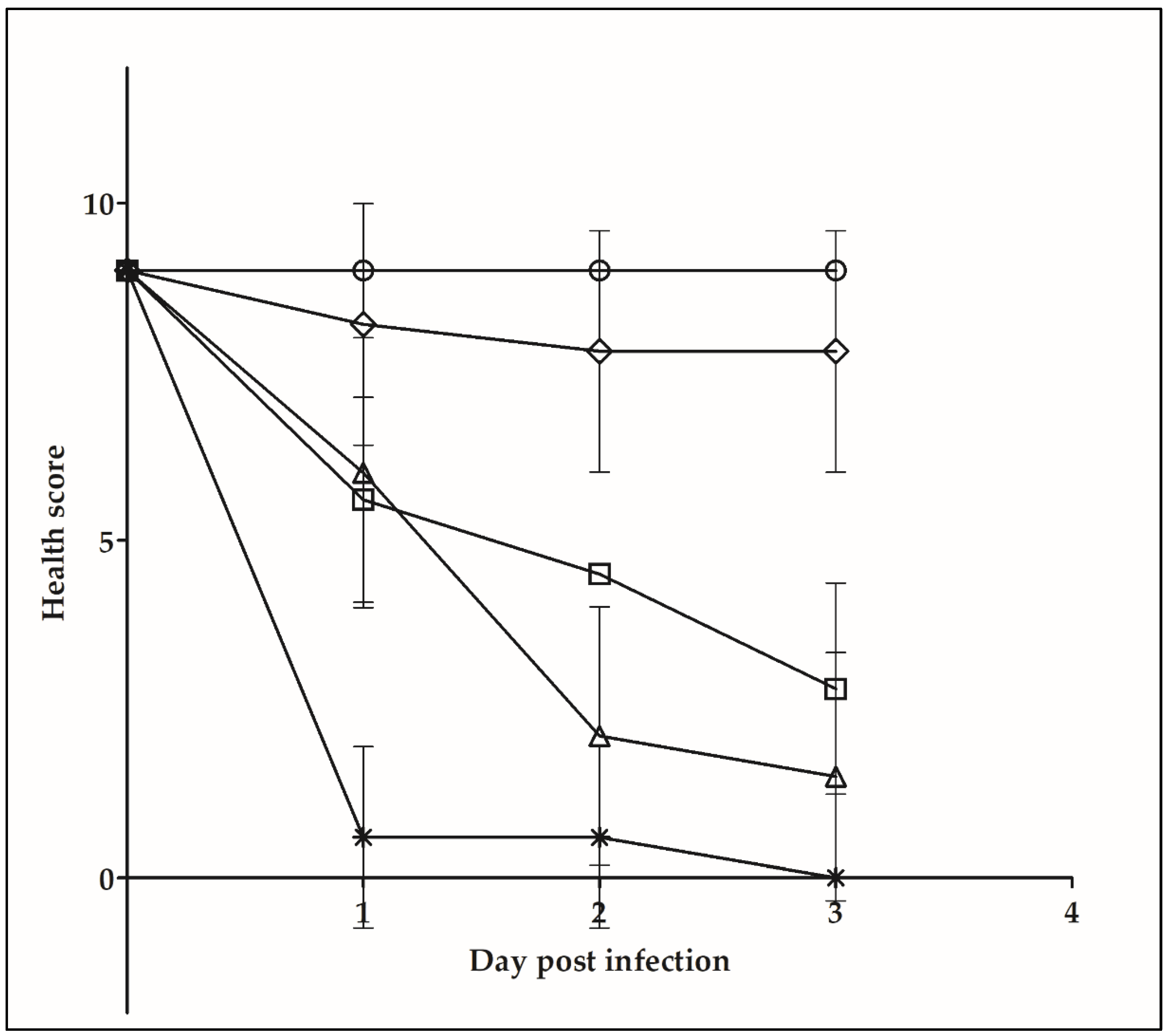
2.5. Galleria mellonella Infection Assay
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Cell Cultures
4.3. Adhesion and Invasion Assays
4.4. Transepithelial Electrical Resistance (TEER) Measurements
4.5. Reporter Assay for TLR Receptor Activation
4.6. Fluorescence Microscopy
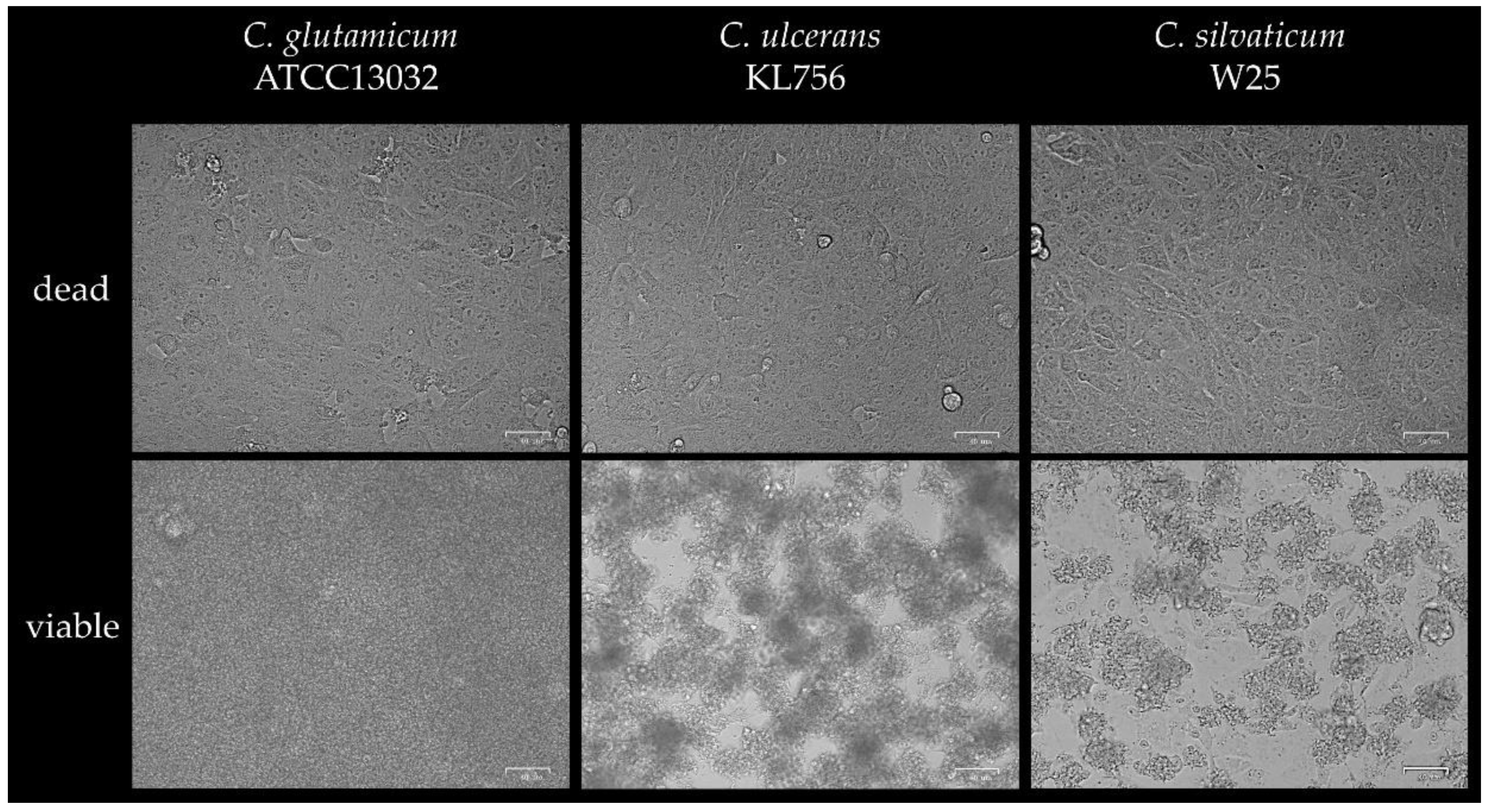
4.7. Cytotoxicity and LDH Activity Assay
4.8. Galleria Mellonella Infection
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CFU | Colony-forming units |
| DT | Diphtheria toxin |
| HEK | Human embryonal kidney |
| LD | Linear dichroism |
| LDH | Lactate dehydrogenase |
| NTTB | Non-toxigenic toxin-bearing |
| OD | Optical density |
| SEAP | Secreted alkaline phosphatase |
| TEER | Transepithelial electrical resistance |
| TLA | Three-letter acronym |
References
- LPSN.dsmz.de. Available online: www.bacterio.net/corynebacterium.html (accessed on 2 June 2020).
- Tauch, A.; Sandbote, J. The Family Corynebacteriaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackenbrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 239–277. ISBN 9783642301384. [Google Scholar]
- Sangal, V.; Hoskisson, P.A. Corynephages: Infections of the Infectors. In Corynebacterium diphtheriae and Related Toxigenic Species; Burkovski, A., Ed.; Springer: Dobrecht, Germany, 2014; pp. 67–81. ISBN 9789400776241. [Google Scholar]
- Riegel, P.; Ruimy, R.; De Brie, D.; Prkost, G.; Jehl, F.; Christen, R. Taxonomy of Corynebacterium diphtheriae and related taxa, with recognition of Corynebacterium ulcerans sp. nov. nom. rev. FEMS Microbiol. Lett. 1995, 126, 271–276. [Google Scholar] [CrossRef]
- Gilbert, R.; Stewart, F.C. Corynebacterium ulcerans: A pathogenic Microorganism Resembling Corynebacterium diphtheriae. J. Lab. Clin. Med. 1927, 12, 756–761. [Google Scholar]
- Burkovski, A. Pathogenesis of Corynebacterium diphtheriae and Corynebacterium ulcerans. Hum. Emerg. Re-Emerg. Infect. 2016, II, 699–709. [Google Scholar]
- Hacker, E.; Antunes, C.A.; Mattos-Guaraldi, A.L.; Burkovski, A.; Tauch, A. Corynebacterium ulcerans, an emerging human pathogen. Future Microbiol. 2016, 11, 1191–1208. [Google Scholar] [CrossRef]
- RKI Diphtherie: Erkrankung durch toxigene Corynebacterium ulcerans nach Katzenkontakt—Fallbericht. Epidemiol. Bull. 2011, 246–248.
- Lartigue, M.-F.; Monnet, X.; Le Fleche, A.; Grimont, P.A.D.; Benet, J.-J.; Durrbach, A.; Fabre, M.; Nordmann, P. Corynebacterium ulcerans in an immunocompromised patient with diphtheria and her dog. J. Clin. Microbiol. 2005, 43, 999–1001. [Google Scholar] [CrossRef] [Green Version]
- Hogg, R.A.; Wessels, J.; Hart, J.; Efstratiou, A.; De Zoysa, A.; Mann, G.; Allen, T.; Pritchard, G.C. Possible zoonotic transmission of toxigenic Corynebacterium ulcerans from companion animals in a human case of fatal diphtheria. Vet. Rec. 2009, 165, 691–692. [Google Scholar]
- Dias, A.A.S.O.; Silva, F.C.; Santos, L.S.; Ribeiro-Carvalho, M.M.; Sabbadini, P.S.; Santos, C.S.; Filardy, A.A.; Myioshi, A.; Azevedo, V.A.; Hirata, R.; et al. Strain-dependent arthritogenic potential of the zoonotic pathogen Corynebacterium ulcerans. Vet. Microbiol. 2011, 153, 323–331. [Google Scholar] [CrossRef]
- Berger, A.; Dangel, A.; Peters, M.; Mühldorfer, K.; Braune, S.; Eisenberg, T. Tox -positive Corynebacterium ulcerans in hedgehogs, Germany. Emerg. Microbes Infect. 2019, 8, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Berger, A.; Teusch, B.; Heinzinger, S.; Sing, A. Corynebacterium ulcerans—Ein Emerging Pathogen? Daten des Konsiliarlabors für Diphtherie 2011–2016. Epidemiol. Bull. 2018, 8, 83–90. [Google Scholar]
- Antunes, C.A.; Clark, L.; Wanuske, M.T.; Hacker, E.; Ott, L.; Simpson-Louredo, L.; de Luna, M.D.G.; Hirata, R.; Mattos-Guaraldi, A.L.; Hodgkin, J.; et al. Caenorhabditis elegans star formation and negative chemotaxis induced by infection with corynebacteria. Microbiology 2016, 162, 84–93. [Google Scholar] [CrossRef]
- Ott, L.; Mckenzie, A.; Baltazar, M.T.; Britting, S.; Bischof, A.; Burkovski, A.; Hoskisson, P.A. Evaluation of invertebrate infection models for pathogenic corynebacteria. FEMS Immunol. Med. Microbiol. 2012, 65, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Weerasekera, D.; Möller, J.; Kraner, M.E.; Antunes, C.A.; Mattos-Guaraldi, A.L.; Burkovski, A. Beyond diphtheria toxin: Cytotoxic proteins of Corynebacterium ulcerans and Corynebacterium diphtheriae. Microbiology 2019, 165, 1–15. [Google Scholar] [CrossRef]
- Peixoto, R.S.; Hacker, E.; Antunes, C.A.; Weerasekera, D.; Alves, A.; De Oliveira, D.S.; Martins, C.A.; Júnior, R.H.; Burkovski, A.; Mattos-Guaraldi, A.L. Pathogenic properties of a Corynebacterium diphtheriae strain isolated from a case of osteomyelitis. J. Med. Microbiol. 2016, 65, 1311–1321. [Google Scholar] [CrossRef]
- Weerasekera, D.; Stengel, F.; Sticht, H.; Mattos-Guaraldi, A.L.; Burkovski, A.; Antunes, C.A. The C-terminal coiled-coil domain of Corynebacterium diphtheriae DIP0733 is crucial for interaction with epithelial cells and pathogenicity in invertebrate animal model systems. BMC Microbiol. 2018, 18, 1–13. [Google Scholar] [CrossRef]
- Dorella, F.A.; Pacheco, L.G.C.; Oliveira, S.C.; Miyoshi, A.; Azevedo, V. Corynebacterium pseudotuberculosis: Microbiology, biochemical properties, pathogenesis and molecular studies of virulence. Vet. Res. 2006, 37, 201–2018. [Google Scholar] [CrossRef] [Green Version]
- Baird, G.J.; Fontaine, M.C. Corynebacterium pseudotuberculosis and its role in ovine caseous lymphadenitis. J. Comp. Pathol. 2007, 137, 179–201. [Google Scholar] [CrossRef]
- Contzen, M.; Sting, R.; Blazey, B.; Rau, J. Corynebacterium ulcerans from diseased wild boars. Zoonoses Public Health 2011, 58, 479–488. [Google Scholar] [CrossRef]
- Rau, J.; Blazey, B.; Contzen, M.; Sting, R. Corynebacterium ulcerans-Infektion bei einem Reh (Capreolus capreolus). Berl. Munch. Tierarztl. Wochenschr. 2012, 125, 159–162. [Google Scholar]
- Dangel, A.; Berger, A.; Rau, J.; Eisenberg, T.; Kämpfer, P.; Margos, G.; Contzen, M.; Busse, H.-J.; Konrad, R.; Peters, M.; et al. Corynebacterium silvaticum sp. nov., a unique group of NTTB corynebacteria in wild boar and roe deer. Int. J. Syst. Evol. Microbiol. 2020, 70, 3614–3624. [Google Scholar] [CrossRef]
- Rau, J.; Eisenberg, T.; Peters, M.; Berger, A.; Kutzer, P.; Lassnig, H.; Hotzel, H.; Sing, A.; Sting, R.; Contzen, M. Reliable differentiation of a non-toxigenic tox gene-bearing Corynebacterium ulcerans variant frequently isolated from game animals using MALDI-TOF MS. Vet. Microbiol. 2019, 237, 108399. [Google Scholar] [CrossRef]
- Viana, M.V.C.; Profeta, R.; da Silva, A.L.; Hurtado, R.; Cerqueira, J.C.; Ribeiro, B.F.S.; Almeida, M.O.; Morais-Rodrigues, F.; de Castro Soares, S.; Oliveira, M.; et al. Taxonomic classification of strain PO100/5 shows a broader geographic distribution and genetic markers of the recently described Corynebacterium silvaticum. PLoS ONE 2021, 15, 1–18. [Google Scholar]
- Möller, J.; Musella, L.; Melnikov, V.; Geißdörfer, W.; Burkovski, A.; Sangal, V. Phylogenomic characterisation of a novel corynebacterial species pathogenic to animals. Antonie Leeuwenhoek 2020, 113, 1225–1239. [Google Scholar] [CrossRef]
- Busch, A.; Möller, J.; Burkovski, A.; Hotzel, H. Genome sequence of a pathogenic Corynebacterium ulcerans strain isolated from a wild boar with necrotizing lymphadenitis. BMC Res. Notes 2019, 12, 10–12. [Google Scholar] [CrossRef] [Green Version]
- Tauch, A.; Burkovski, A. Molecular armory or niche factors: Virulence determinants of Corynebacterium species. FEMS Microbiol. Lett. 2015, 362, 1–6. [Google Scholar]
- Trost, E.; Al-dilaimi, A.; Papavasiliou, P.; Schneider, J.; Viehoever, P.; Burkovski, A.; Soares, S.C.; Almeida, S.S.; Dorella, F.A.; Miyoshi, A.; et al. Comparative analysis of two complete Corynebacterium ulcerans genomes and detection of candidate virulence factors. BMC Genom. 2011, 12, 383. [Google Scholar] [CrossRef] [Green Version]
- Hacker, E.; Ott, L.; Schulze-Luehrmann, J.; Lürmann, A.; Wiesmann, V.; Wittenberg, T.; Burkovski, A. The killing of macrophages by Corynebacterium ulcerans. Virulence 2016, 7, 45–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacker, E.; Ott, L.; Hasselt, K.; Mattos-Guaraldi, A.L.; Tauch, A.; Burkovski, A. Colonization of human epithelial cell lines by Corynebacterium ulcerans from human and animal sources. Microbiology 2015, 161, 1582–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittel, M.; Gastiger, S.; Amin, B.; Hofmann, J.; Burkovski, A. Surface and extracellular proteome of the emerging pathogen Corynebacterium ulcerans. Proteomes 2018, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Subedi, R.; Kolodkina, V.; Sutcliffe, I.C.; Simpson-Louredo, L.; Hirata, R.; Titov, L.; Mattos-Guaraldi, A.L.; Burkovski, A.; Sangal, V. Genomic analyses reveal two distinct lineages of Corynebacterium ulcerans strains. New Microbes New Infect. 2018, 25, 7–13. [Google Scholar] [CrossRef]
- Weerasekera, D.; Fastner, T.; Lang, R.; Burkovski, A.; Ott, L. Of mice and men: Interaction of Corynebacterium diphtheriae strains with murine and human phagocytes. Virulence 2019, 10, 414–428. [Google Scholar] [CrossRef] [Green Version]
- Möller, J.; Schorlemmer, S.; Hofmann, J.; Burkovski, A. Cellular and extracellular proteome of the animal pathogen Corynebacterium silvaticum, a close relative of zoonotic Corynebacterium ulcerans and Corynebacterium pseudotuberculosis. Proteomes 2020, 8, 19. [Google Scholar] [CrossRef]
- Ott, L.; Höller, M.; Rheinlaender, J.; Schäffer, T.E.; Hensel, M.; Burkovski, A. Strain-specific differences in pili formation and the interaction of Corynebacterium diphtheriae with host cells. BMC Microbiol. 2010, 10, 257. [Google Scholar] [CrossRef] [Green Version]
- Jander, G.; Rahme, L.; Ausubel, F.M. Positive Correlation between virulence of Pseudomonas aeruginosa mutants in mice and insects. J. Bacteriol. 2000, 182, 3843–3845. [Google Scholar] [CrossRef] [Green Version]
- Wand, M.E.; Müller, C.M.; Titball, R.W.; Michell, S.L. Macrophage and Galleria mellonella infection models reflect the virulence of naturally occurring isolates of B. pseudomallei, B. thailandensis and B. oklahomensis. BMC Microbiol. 2011, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.J.-Y.; Loh, J.M.S.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef] [Green Version]
- Mandlik, A.; Swierczynski, A.; Das, A.; Ton-that, H. Pili in Gram-positive bacteria: Assembly, involvement in colonization and biofilm development. Trend Microbiol. 2008, 16, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Mandlik, A.; Swierczynski, A.; Das, A.; Ton-that, H. Corynebacterium diphtheriae employs specific minor pilins to target human pharyngeal epithelial cells. Mol. Microbiol. 2007, 64, 111–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, L. Adhesion properties of toxigenic corynebacteria. AIMS Microbiol. 2018, 4, 85–103. [Google Scholar] [CrossRef]
- Fontaine, M.C.; Baird, G.J. Caseous lymphadenitis. Small Ruminand Res. 2008, 76, 42–48. [Google Scholar] [CrossRef]
- Windsor, P.A. Contol of caseous lymphadenitis. Vet. Clin. NA Food Anim. Pract. 2011, 27, 193–202. [Google Scholar] [CrossRef]
- Jorge, K.T.O.S.; Santos, T.M.; Tartaglia, N.R.; Aguiar, E.L.; Souza, R.F.S.; Mariutti, R.B.; Eberle, R.J.; Arni, R.K.; Portela, R.W.; Meyer, R. Putative virulence factors of Corynebacterium pseudotuberculosis FRC41: Vaccine potential and protein expression. Microb. Cell Fact. 2016, 15, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, A.D.S.; Borges, F.; Pauletti, R.B.; Seyffert, N.; Ribeiro, D.; Lage, A.P.; Heinemann, M.B.; Miyoshi, A.; Maria, A.; Gouveia, G. Caseous lymphadenitis: Epidemiology, diagnosis, and control. IIOAB J. 2011, 2, 33–43. [Google Scholar]
- Akira, S.; Hemmi, H. Recognition of pathogen-associated molecular patterns by TLR family. Immunol. Lett. 2003, 85, 85–95. [Google Scholar] [CrossRef]
- Oliveira-nascimento, L.; Massari, P.; Wetzler, L.M.; Lauvau, G.S.; Einstein, A.; Brinkmann, M.M. The role of TLR2 in infection and immunity. Front. Immunol. 2012, 3, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, C.M.; Mcelvaney, N.G. Toll-like receptor expression and function in airway epithelial cells. Arch. Immunolgiae Ther. Exp. 2005, 53, 418–427. [Google Scholar]
- West, A.P.; Koblansky, A.A. Recognition and Signaling by Toll-Like Receptors. Annu. Rev. Cell Dev. Biol. 2006, 22, 409–437. [Google Scholar] [CrossRef]
- Honda, K.; Ohba, Y.; Yanai, H.; Hegishi, H.; Mizutani, T.; Takaoka, A.; Taya, C.; Taniguchi, T. Spatiotemporal regulation of MyD88-IRF-7 signalling for robust type-I interferon induction. Nature 2005, 434, 1035–1040. [Google Scholar] [CrossRef]
- Costa-Mattioli, M.; Sonenberg, N. RAPping production of type I interferon in pDCs through mTOR. Nat. Immunol. 2008, 9, 1097–1099. [Google Scholar] [CrossRef]
- Honda, K.; Taniguchi, T. IRFs: Master regulators of signalling by Toll-like receptors and cytosolic pattern-recognition receptors. Nat. Rev. Immunol. 2006, 6, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Meylan, E.; Tschopp, J.; Karin, M. Intracellular pattern recognition receptors in the host response. Nature 2006, 442, 39–44. [Google Scholar] [CrossRef]
- Cutuli, M.A.; Petronio, G.P.; Venditti, N.; Di Marco, R.; Vergalito, F.; Magnifico, I.; Pietrangelo, L. Galleria mellonella as a consolidated in vivo model hosts: New developments in antibacterial strategies and novel drug testing. Virulence 2019, 10, 527–541. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Tang, H. Regulation and function of the melanization reaction in Drosophila. Fly 2009, 3, 105–111. [Google Scholar] [CrossRef]
- Abe, S.; Takayama, K.-I.; Kinoshita, S. Taxonomical studies on glutamic acid-producing bacteria. J. Gen. Appl. Microbiol. 1967, 13, 279–301. [Google Scholar] [CrossRef]
- Möller, J.; Kraner, M.; Sonnewald, U.; Sangal, V.; Tittlbach, H.; Winkler, J.; Winkler, T.H.; Melnikov, V.; Lang, R.; Sing, A.; et al. Proteomics of diphtheria toxoid vaccines reveals multiple proteins that are immunogenic and may contribute to protection of humans against Corynebacterium diphtheriae. Vaccine 2019, 37, 3061–3070. [Google Scholar] [CrossRef]
- Knoppová, M.; Phensaijai, M.; Veselý, M.; Zemanová, M.; Nešvera, J.; Pátek, M. Plasmid vectors for testing in vivo promoter activities in Corynebacterium glutamicum and Rhodococcus erythropolis. Curr. Microbiol. 2007, 55, 234–239. [Google Scholar] [CrossRef]
- Gey, G.O.; Coffman, W.D.; Kubicek, M.D. Tissue culture studies of the proliferative capacity of cervical carcinoma and normal epithelium. Cancer Res. 1952, 12, 264–265. [Google Scholar]
- Scherer, W.F.; Syverton, J.T.; Gey, G. Studies on the propagation in vitro of poliomyelitis viruses. IV. Viral multiplication in a stable strain of human malignant epithelial cells (strain HeLa) derived from an epidermoid carcinoma of the cervix. J. Exp. Med. 1953, 97, 695–710. [Google Scholar] [CrossRef] [Green Version]
- Yasumura, Y.; Kawakita, Y. Studies on SV40 in tissue culture: Preliminary step for cancer research In Vitro. Nihon Rinsho 1963, 21, 1201–1215. [Google Scholar]
- Peterson, W.D.; Stulberg, C.S.; Swanborg, N.K.; Robinson, A.R. Glucose-6-phosphate dehydrogenase isoenzymes in human cell cultutes determined by sucrose-agar gel and cellulose acetate zymograms. Proc. Soc. Exp. Biol. Med. 1968, 128, 772–776. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description/Source | Reference |
| C. glutamicum ATCC13032 | Type strain, non-pathogenic | [57] |
| C. ulcerans KL756 | Dog (tox+) | [58] |
| C. silvaticum W25 | Wild boar (tox+) | [27] |
| Salmonella enterica serovar Typhimurium NCTC 12023 | Wild-type (identical to ATCC 14028) | National Collection of TypeCultures (Colindale, UK) |
| Plasmids | Description | Reference |
| pERP1p45_gfp | gfpuv, KmR, rep, per, T1, T2 | [59] |
| Cell lines | Description | Reference |
| HeLa | Human cervical carcinoma cells | [60,61] |
| HEK-Blue 293 hTLR2 | Human TLR2/NFкB/SEAP reporter HEK293 cells | Invivogen |
| HEK-Blue 293 hTLR9 | Human TLR9/NFкB/SEAP reporter HEK293 cells | Invivogen |
| Vero | African green monkey kidney epithelial cells | [62] |
| Detroit 562 | Human hypopharyngeal carcinoma cells | [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Möller, J.; Busch, A.; Berens, C.; Hotzel, H.; Burkovski, A. Newly Isolated Animal Pathogen Corynebacterium silvaticum Is Cytotoxic to Human Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 3549. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073549
Möller J, Busch A, Berens C, Hotzel H, Burkovski A. Newly Isolated Animal Pathogen Corynebacterium silvaticum Is Cytotoxic to Human Epithelial Cells. International Journal of Molecular Sciences. 2021; 22(7):3549. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073549
Chicago/Turabian StyleMöller, Jens, Anne Busch, Christian Berens, Helmut Hotzel, and Andreas Burkovski. 2021. "Newly Isolated Animal Pathogen Corynebacterium silvaticum Is Cytotoxic to Human Epithelial Cells" International Journal of Molecular Sciences 22, no. 7: 3549. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073549