
Molecular Signatures of Natural Killer Cells in CMV-Associated Anterior Uveitis, A New Type of CMV-Induced Disease in Immunocompetent Individuals

 ,
,
Abstract
:1. Introduction
2. Results
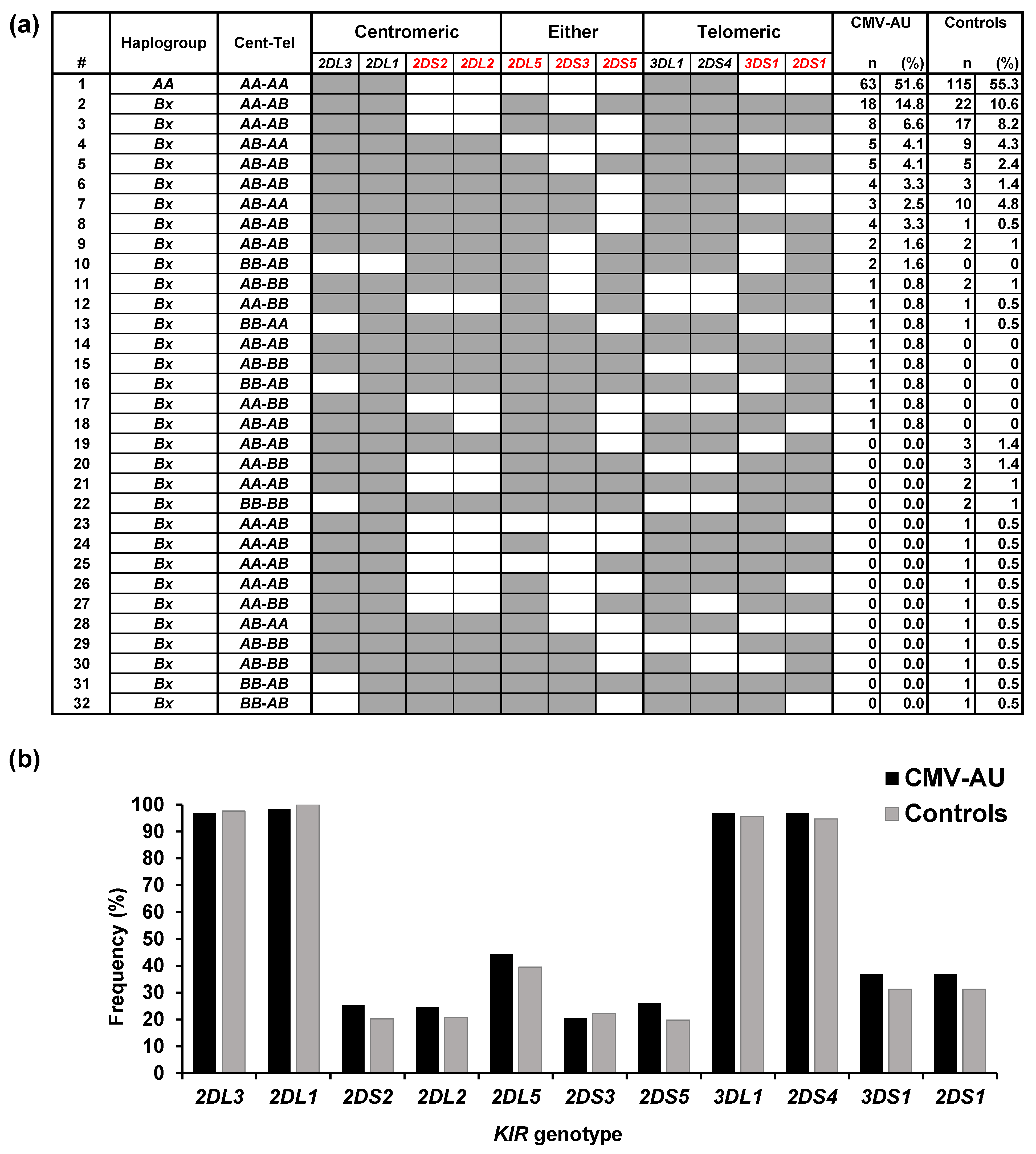
2.1. Polymorphisms of KIRs and Their Ligands Associated with CMV-AU
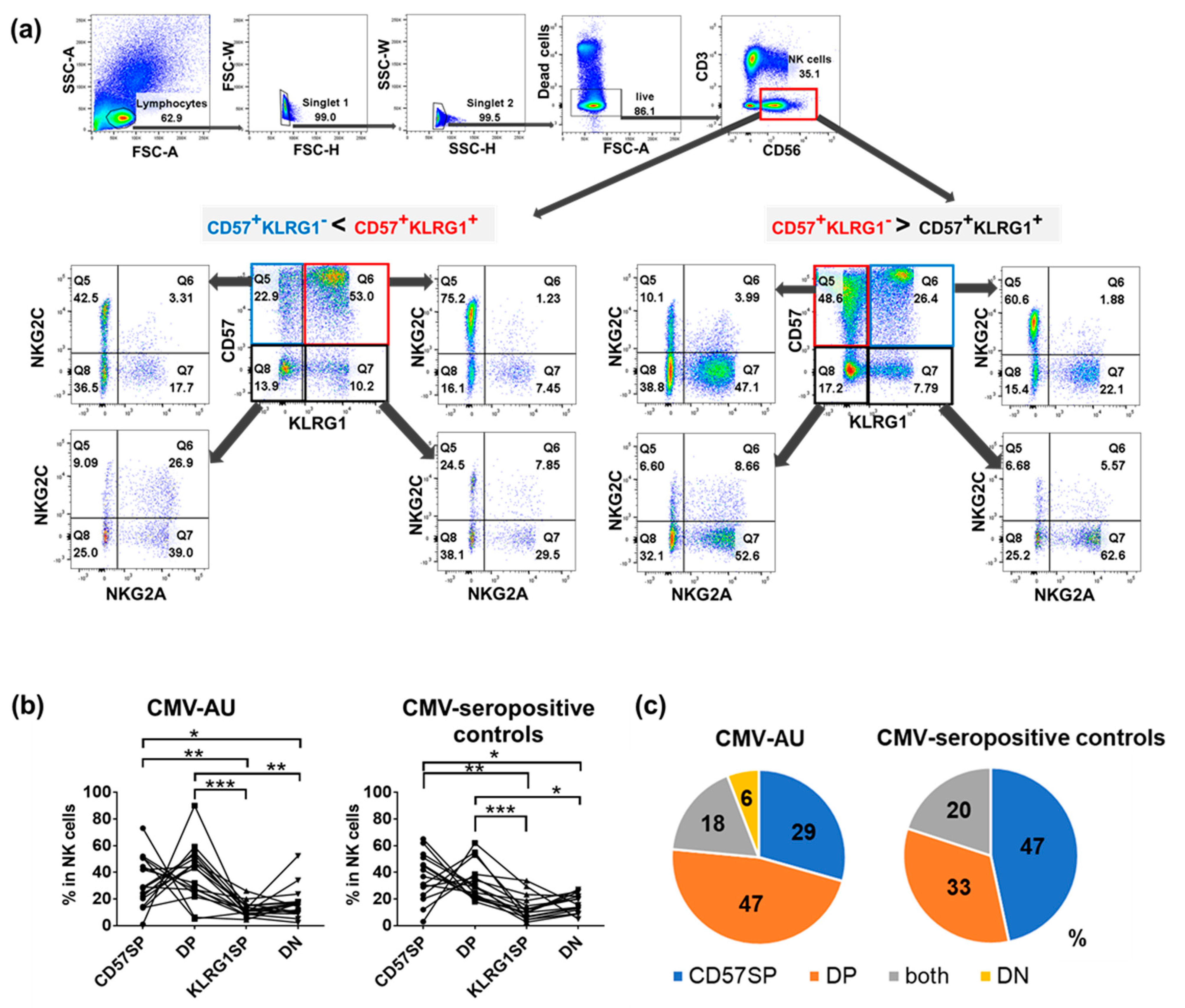
2.2. Identification of Four NK Cell Subsets Distinguished by Differential Expression of CD57 and KLRG1: Expansion of CD57+KLRG1− and CD57+KLRG1+ NK Cell Populations in CMV-Seropositive Individuals
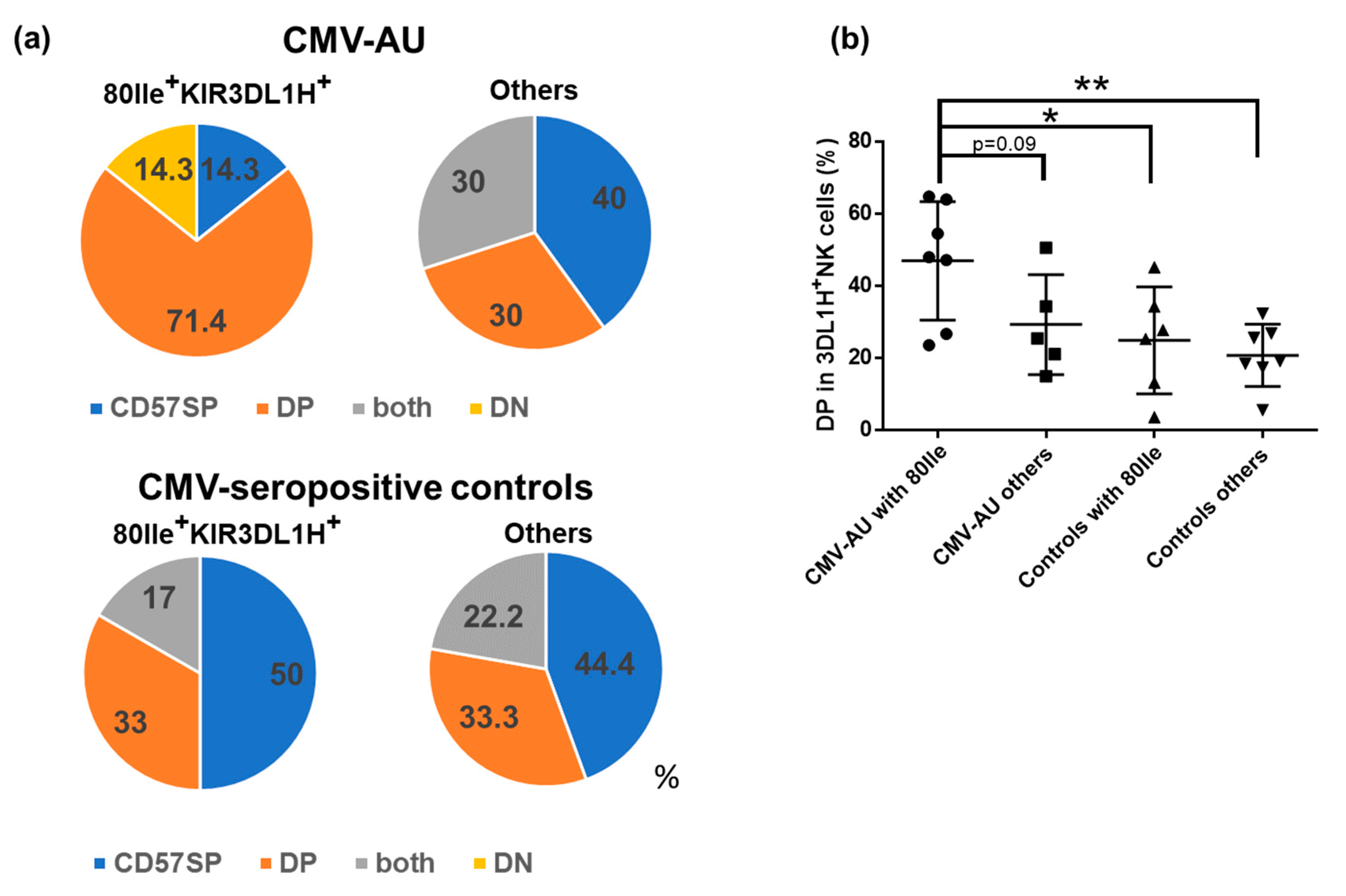
2.3. CD57/KLRG1 DP Subset Expansion in CMV-AU Individuals with HLA-B Bw4 80Ile and KIR3DL1H
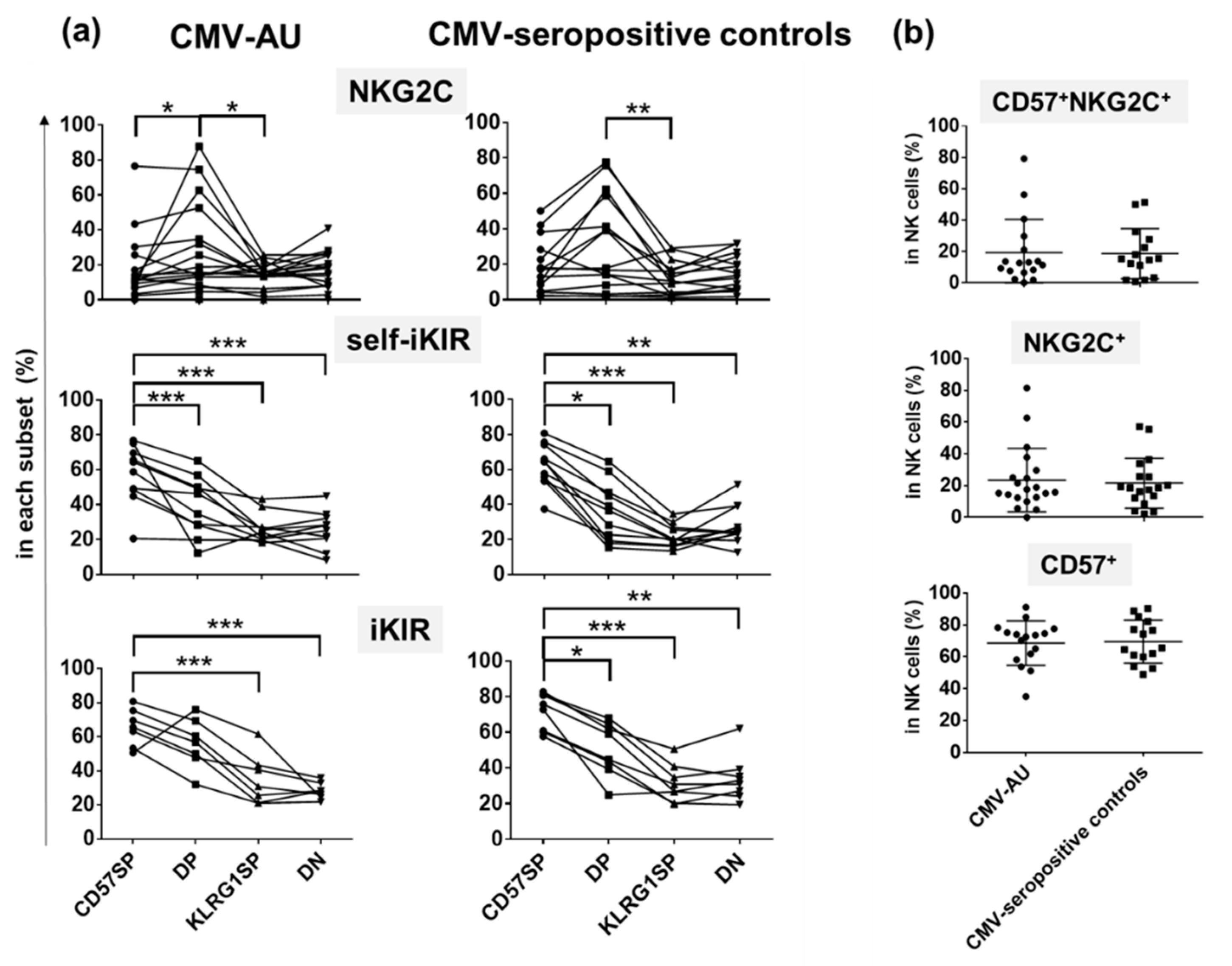
2.4. Differential Expression of NKG2C and iKIRs in CD57SP and DP NK Subsets
3. Discussion
4. Materials and Methods
4.1. Human Subjects
4.2. KIR Genotyping, KIR3DL1 Allele Typing and KIR Ligand Typing
4.3. Antibodies and Flow Cytometry
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CMV | Cytomegalovirus |
| CMV-AU | CMV-associated anterior uveitis |
| NK cells | natural killer cells |
| KIRs | Killer cell Immunoglobulin-like Receptors |
| PBMC | Peripheral blood mononuclear cells |
| PCR-SSP | PCR-based sequence-specific primer typing |
References
- Picarda, G.; Benedict, C.A. Cytomegalovirus: Shape-Shifting the Immune System. J. Immunol. 2018, 200, 3881–3889. [Google Scholar] [CrossRef] [Green Version]
- Berry, R.; Watson, G.M.; Jonjic, S.; Degli-Esposti, M.A.; Rossjohn, J. Modulation of innate and adaptive immunity by cytomegaloviruses. Nat. Rev. Immunol. 2020, 20, 113–127. [Google Scholar] [CrossRef]
- Bastidas-Legarda, L.Y.; Khakoo, S.I. Conserved and variable natural killer cell receptors: Diverse approaches to viral infections. Immunology 2019, 156, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Sugar, E.A.; Jabs, D.A.; Ahuja, A.; Thorne, J.E.; Danis, R.P.; Meinert, C.L.; Studies of the Ocular Complications of AIDS Research Group. Incidence of cytomegalovirus retinitis in the era of highly active antiretroviral therapy. Am. J. Ophthalmol. 2012, 153, 1016–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chee, S.P.; Bacsal, K.; Jap, A.; Se-Thoe, S.Y.; Cheng, C.L.; Tan, B.H. Clinical features of cytomegalovirus anterior uveitis in immunocompetent patients. Am. J. Ophthalmol. 2008, 145, 834–840. [Google Scholar] [CrossRef]
- Koizumi, N.; Suzuki, T.; Uno, T.; Chihara, H.; Shiraishi, A.; Hara, Y.; Inatomi, T.; Sotozono, C.; Kawasaki, S.; Yamasaki, K.; et al. Cytomegalovirus as an etiologic factor in corneal endotheliitis. Ophthalmology 2008, 115, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Chan, N.S.; Chee, S.P.; Caspers, L.; Bodaghi, B. Clinical Features of CMV-Associated Anterior Uveitis. Ocul. Immunol. Inflamm. 2018, 26, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, K.H.; Hasegawa, E.; Namba, K.; Okada, A.A.; Ohguro, N.; Goto, H.; Group, J.U.S.W. Epidemiology of uveitis in Japan: A 2016 retrospective nationwide survey. Jpn. J. Ophthalmol. 2021, 65, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Chee, S.P.; Jap, A. Cytomegalovirus anterior uveitis: Outcome of treatment. Br. J. Ophthalmol. 2010, 94, 1648–1652. [Google Scholar] [CrossRef]
- Chee, S.P.; Jap, A. Presumed fuchs heterochromic iridocyclitis and Posner-Schlossman syndrome: Comparison of cytomegalovirus-positive and negative eyes. Am. J. Ophthalmol. 2008, 146, 883–889. [Google Scholar] [CrossRef]
- Orange, J.S. Natural killer cell deficiency. J. Allergy Clin. Immunol. 2013, 132, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Biron, C.A.; Byron, K.S.; Sullivan, J.L. Severe herpesvirus infections in an adolescent without natural killer cells. N. Engl. J. Med. 1989, 320, 1731–1735. [Google Scholar] [CrossRef]
- Yawata, M.; Yawata, N.; Draghi, M.; Little, A.M.; Partheniou, F.; Parham, P. Roles for HLA and KIR polymorphisms in natural killer cell repertoire selection and modulation of effector function. J. Exp. Med. 2006, 203, 633–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karre, K.; Ljunggren, H.G.; Piontek, G.; Kiessling, R. Selective rejection of H-2-deficient lymphoma variants suggests alternative immune defence strategy. Nature 1986, 319, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Anfossi, N.; Andre, P.; Guia, S.; Falk, C.S.; Roetynck, S.; Stewart, C.A.; Breso, V.; Frassati, C.; Reviron, D.; Middleton, D.; et al. Human NK cell education by inhibitory receptors for MHC class I. Immunity 2006, 25, 331–342. [Google Scholar] [CrossRef]
- Kim, S.; Poursine-Laurent, J.; Truscott, S.M.; Lybarger, L.; Song, Y.J.; Yang, L.; French, A.R.; Sunwoo, J.B.; Lemieux, S.; Hansen, T.H.; et al. Licensing of natural killer cells by host major histocompatibility complex class I molecules. Nature 2005, 436, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Yawata, M.; Yawata, N.; Draghi, M.; Partheniou, F.; Little, A.M.; Parham, P. MHC class I-specific inhibitory receptors and their ligands structure diverse human NK-cell repertoires toward a balance of missing self-response. Blood 2008, 112, 2369–2380. [Google Scholar] [CrossRef] [Green Version]
- Parham, P. MHC class I molecules and KIRs in human history, health and survival. Nat. Rev. Immunol. 2005, 5, 201–214. [Google Scholar] [CrossRef]
- Boudreau, J.E.; Mulrooney, T.J.; le Luduec, J.B.; Barker, E.; Hsu, K.C. KIR3DL1 and HLA-B Density and Binding Calibrate NK Education and Response to HIV. J. Immunol. 2016, 196, 3398–3410. [Google Scholar] [CrossRef] [PubMed]
- Schust, D.J.; Tortorella, D.; Seebach, J.; Phan, C.; Ploegh, H.L. Trophoblast class I major histocompatibility complex (MHC) products are resistant to rapid degradation imposed by the human cytomegalovirus (HCMV) gene products US2 and US11. J. Exp. Med. 1998, 188, 497–503. [Google Scholar] [CrossRef] [Green Version]
- Tomasec, P.; Braud, V.M.; Rickards, C.; Powell, M.B.; McSharry, B.P.; Gadola, S.; Cerundolo, V.; Borysiewicz, L.K.; McMichael, A.J.; Wilkinson, G.W. Surface expression of HLA-E, an inhibitor of natural killer cells, enhanced by human cytomegalovirus gpUL40. Science 2000, 287, 1031–1033. [Google Scholar] [CrossRef]
- Heatley, S.L.; Pietra, G.; Lin, J.; Widjaja, J.M.; Harpur, C.M.; Lester, S.; Rossjohn, J.; Szer, J.; Schwarer, A.; Bradstock, K.; et al. Polymorphism in human cytomegalovirus UL40 impacts on recognition of human leukocyte antigen-E (HLA-E) by natural killer cells. J. Biol. Chem. 2013, 288, 8679–8690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guma, M.; Angulo, A.; Vilches, C.; Gomez-Lozano, N.; Malats, N.; Lopez-Botet, M. Imprint of human cytomegalovirus infection on the NK cell receptor repertoire. Blood 2004, 104, 3664–3671. [Google Scholar] [CrossRef] [Green Version]
- Rolle, A.; Brodin, P. Immune Adaptation to Environmental Influence: The Case of NK Cells and HCMV. Trends Immunol. 2016, 37, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Linn, Y.C.; Ren, E.C. KIR-HLA profiling shows presence of higher frequencies of strong inhibitory KIR-ligands among prognostically poor risk AML patients. Immunogenetics 2016, 68, 133–144. [Google Scholar] [CrossRef]
- Wong, A.; Tan, K.H.; Tee, C.S.; Yeo, G.S. Seroprevalence of cytomegalovirus, toxoplasma and parvovirus in pregnancy. Singap. Med. J. 2000, 41, 151–155. [Google Scholar]
- Yawata, M.; Yawata, N.; Abi-Rached, L.; Parham, P. Variation within the human killer cell immunoglobulin-like receptor (KIR) gene family. Crit. Rev. Immunol. 2002, 22, 463–482. [Google Scholar] [CrossRef]
- Hsu, K.C.; Chida, S.; Geraghty, D.E.; Dupont, B. The killer cell immunoglobulin-like receptor (KIR) genomic region: Gene-order, haplotypes and allelic polymorphism. Immunol. Rev. 2002, 190, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Yawata, M.; Yawata, N.; McQueen, K.L.; Cheng, N.W.; Guethlein, L.A.; Rajalingam, R.; Shilling, H.G.; Parham, P. Predominance of group A KIR haplotypes in Japanese associated with diverse NK cell repertoires of KIR expression. Immunogenetics 2002, 54, 543–550. [Google Scholar] [CrossRef]
- Jiang, K.; Zhu, F.M.; Lv, Q.F.; Yan, L.X. Distribution of killer cell immunoglobulin-like receptor genes in the Chinese Han population. Tissue Antigens 2005, 65, 556–563. [Google Scholar] [CrossRef]
- Cooley, S.; Weisdorf, D.J.; Guethlein, L.A.; Klein, J.P.; Wang, T.; Le, C.T.; Marsh, S.G.; Geraghty, D.; Spellman, S.; Haagenson, M.D.; et al. Donor selection for natural killer cell receptor genes leads to superior survival after unrelated transplantation for acute myelogenous leukemia. Blood 2010, 116, 2411–2419. [Google Scholar] [CrossRef] [Green Version]
- Carrington, M.; Martin, M.P.; van Bergen, J. KIR-HLA intercourse in HIV disease. Trends Microbiol. 2008, 16, 620–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, C.M.; Guethlein, L.A.; Shilling, H.G.; Pando, M.; Carr, W.H.; Rajalingam, R.; Vilches, C.; Parham, P. Different NK cell surface phenotypes defined by the DX9 antibody are due to KIR3DL1 gene polymorphism. J. Immunol. 2001, 166, 2992–3001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beziat, V.; Liu, L.L.; Malmberg, J.A.; Ivarsson, M.A.; Sohlberg, E.; Bjorklund, A.T.; Retiere, C.; Sverremark-Ekstrom, E.; Traherne, J.; Ljungman, P.; et al. NK cell responses to cytomegalovirus infection lead to stable imprints in the human KIR repertoire and involve activating KIRs. Blood 2013, 121, 2678–2688. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.C.; Beilke, J.N.; Lanier, L.L. Adaptive immune features of natural killer cells. Nature 2009, 457, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Guma, M.; Budt, M.; Saez, A.; Brckalo, T.; Hengel, H.; Angulo, A.; Lopez-Botet, M. Expansion of CD94/NKG2C+ NK cells in response to human cytomegalovirus-infected fibroblasts. Blood 2006, 107, 3624–3631. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Botet, M.; Muntasell, A.; Vilches, C. The CD94/NKG2C+ NK-cell subset on the edge of innate and adaptive immunity to human cytomegalovirus infection. Semin. Immunol. 2014, 26, 145–151. [Google Scholar] [CrossRef]
- Lopez-Verges, S.; Milush, J.M.; Schwartz, B.S.; Pando, M.J.; Jarjoura, J.; York, V.A.; Houchins, J.P.; Miller, S.; Kang, S.M.; Norris, P.J.; et al. Expansion of a unique CD57+ NKG2Chi natural killer cell subset during acute human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 14725–14732. [Google Scholar] [CrossRef] [Green Version]
- Manser, A.R.; Scherenschlich, N.; Thons, C.; Hengel, H.; Timm, J.; Uhrberg, M. KIR Polymorphism Modulates the Size of the Adaptive NK Cell Pool in Human Cytomegalovirus-Infected Individuals. J. Immunol. 2019, 203, 2301–2309. [Google Scholar] [CrossRef]
- Leffell, M.S.; Cherikh, W.S.; Land, G.; Zachary, A.A. Improved definition of human leukocyte antigen frequencies among minorities and applicability to estimates of transplant compatibility. Transplantation 2007, 83, 964–972. [Google Scholar] [CrossRef]
- Deng, Z.; Zhen, J.; Zhu, B.; Zhang, G.; Yu, Q.; Wang, D.; Xu, Y.; He, L.; Lu, L. Allelic diversity of KIR3DL1/3DS1 in a southern Chinese population. Hum. Immunol. 2015, 76, 663–666. [Google Scholar] [CrossRef]
- Norman, P.J.; Abi-Rached, L.; Gendzekhadze, K.; Korbel, D.; Gleimer, M.; Rowley, D.; Bruno, D.; Carrington, C.V.; Chandanayingyong, D.; Chang, Y.H.; et al. Unusual selection on the KIR3DL1/S1 natural killer cell receptor in Africans. Nat. Genet. 2007, 39, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Freud, A.G.; Caligiuri, M.A. Human natural killer cell development. Immunol. Rev. 2006, 214, 56–72. [Google Scholar] [CrossRef] [PubMed]
- di Bona, D.; Scafidi, V.; Plaia, A.; Colomba, C.; Nuzzo, D.; Occhino, C.; Tuttolomondo, A.; Giammanco, G.; de Grazia, S.; Montalto, G.; et al. HLA and killer cell immunoglobulin-like receptors influence the natural course of CMV infection. J. Infect. Dis. 2014, 210, 1083–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Duin, D.; Avery, R.K.; Hemachandra, S.; Yen-Lieberman, B.; Zhang, A.; Jain, A.; Butler, R.S.; Barnard, J.; Schold, J.D.; Fung, J.; et al. KIR and HLA interactions are associated with control of primary CMV infection in solid organ transplant recipients. Am. J. Transplant. Off. J. Am. Soc. Transplant. Am. Soc. Transplant. Surg. 2014, 14, 156–162. [Google Scholar] [CrossRef]
- Gonzalez, A.; Schmitter, K.; Hirsch, H.H.; Garzoni, C.; van Delden, C.; Boggian, K.; Mueller, N.J.; Berger, C.; Villard, J.; Manuel, O.; et al. KIR-associated protection from CMV replication requires pre-existing immunity: A prospective study in solid organ transplant recipients. Genes Immun. 2014, 15, 495–499. [Google Scholar] [CrossRef]
- Cook, M.; Briggs, D.; Craddock, C.; Mahendra, P.; Milligan, D.; Fegan, C.; Darbyshire, P.; Lawson, S.; Boxall, E.; Moss, P. Donor KIR genotype has a major influence on the rate of cytomegalovirus reactivation following T-cell replete stem cell transplantation. Blood 2006, 107, 1230–1232. [Google Scholar] [CrossRef] [Green Version]
- Savva, G.M.; Pachnio, A.; Kaul, B.; Morgan, K.; Huppert, F.A.; Brayne, C.; Moss, P.A.; The Medical Research Council Cognitive Function and Ageing Study. Cytomegalovirus infection is associated with increased mortality in the older population. Aging Cell 2013, 12, 381–387. [Google Scholar] [CrossRef]
- Vilches, C.; Castano, J.; Gomez-Lozano, N.; Estefania, E. Facilitation of KIR genotyping by a PCR-SSP method that amplifies short DNA fragments. Tissue Antigens 2007, 70, 415–422. [Google Scholar] [CrossRef]
- Yawata, N.; Selva, K.J.; Liu, Y.C.; Tan, K.P.; Lee, A.W.; Siak, J.; Lan, W.; Vania, M.; Arundhati, A.; Tong, L.; et al. Dynamic change in natural killer cell type in the human ocular mucosa in situ as means of immune evasion by adenovirus infection. Mucosal Immunol. 2016, 9, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Nelson, W.C.; Pyo, C.W.; Vogan, D.; Wang, R.; Pyon, Y.S.; Hennessey, C.; Smith, A.; Pereira, S.; Ishitani, A.; Geraghty, D.E. An integrated genotyping approach for HLA and other complex genetic systems. Hum. Immunol. 2015, 76, 928–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Haplotype Group | B Content | Motif | CMV-AU | Controls | |||
|---|---|---|---|---|---|---|---|
| Score | Cen | Tel | n | (%) | n | (%) | |
| AA | 0 | AA | AA | 63 | 51.6 | 115 | 55.3 |
| Bx | 1 | AA | AB | 26 | 21.3 | 45 | 21.6 |
| AB | AA | 8 | 6.6 | 20 | 9.6 | ||
| 2 | AA | BB | 2 | 1.6 | 5 | 2.4 | |
| AB | AB | 17 | 13.9 | 14 | 6.7 * | ||
| BB | AA | 1 | 0.8 | 1 | 0.5 | ||
| 3 | AB | BB | 2 | 1.6 | 4 | 1.9 | |
| BB | AB | 3 | 2.5 | 2 | 1 | ||
| 4 | BB | BB | 0 | 0 | 2 | 1 | |
| CMV-AU | Controls | P | |||||
| n | % | n | % | ||||
| C1+ | 118 | 99.2 | 206 | 99 | n.s. | ||
| C2+ | 27 | 22.7 | 45 | 21.6 | n.s. | ||
| CMV-AU | Controls | P | OR | 95% CI | |||
| n | % | n | % | ||||
| HLA-B Bw4+ | 69 | 59.5 | 122 | 58.7 | n.s. | ||
| 80Ile+ | 54 | 46.6 | 63 | 30.3 | 0.0058 | 1.94 | 1.23–3.1 |
| 80Thr+ | 17 | 14.7 | 70 | 33.7 | 0.0007 | 0.38 | 0.21–0.67 |
| ratio of 80Ile/80Thr | 3.1 | 0.9 | |||||
| KIR3DL1 | KIR3DL1 | Phenotypes | CMV-AU | CMV-IgG + Controls | ||
|---|---|---|---|---|---|---|
| (1st Allele) | (2nd Allele) | n | % | n | % | |
| 3DL1*001 | 3DL1*01502 | HH | 6 | 4.9 | 2 | 4.7 |
| 3DL1*01502 | 3DL1*01502 | HH | 21 | 17.2 | 15 | 34.9 * |
| 3DL1*01502 | 3DL1*020 | HH | 2 | 1.6 | 0 | 0 |
| 3DL1*01502 | 3DL1*029 | HH | 2 | 1.6 | 0 | 0 |
| 3DL1*020 | 3DL1*001 | HH | 0 | 0 | 1 | 2.3 |
| 3DL1*001 | 3DL1*00501 | HL | 3 | 2.5 | 2 | 4.7 |
| 3DL1*00501 | 3DL1*01502 | HL | 23 | 18.9 | 5 | 11.6 |
| 3DL1*00501 | 3DL1*02901 | HL | 2 | 1.6 | 0 | 0 |
| 3DL1*00701 | 3DL1*01502 | HL | 11 | 9 | 3 | 7 |
| 3DL1*00701 | 3DL1*020 | HL | 1 | 0.8 | 0 | 0 |
| 3DL1*01502 | 3DL1*008 | HL | 0 | 0 | 1 | 2.3 |
| 3DL1*01502 | N | H | 29 | 23.8 | 7 | 16.3 |
| 3DL1*020 | N | H | 2 | 1.6 | 1 | 2.3 |
| 3DL1*029 | N | H | 3 | 2.5 | 0 | 0 |
| 3DL1*00501 | 3DL1*00501 | LL | 2 | 1.6 | 0 | 0 |
| 3DL1*00501 | 3DL1*00701 | LL | 1 | 0.8 | 0 | 0 |
| 3DL1*00701 | 3DL1*00701 | LL | 1 | 0.8 | 2 | 4.7 |
| 3DL1*00501 | N | L | 7 | 5.7 | 3 | 7 |
| 3DL1*00701 | N | L | 1 | 0.8 | 0 | 0 |
| 3DL1*00501 | 3DL1*038 | L ? | 1 | 0.8 | 0 | 0 |
| N | N | 4 | 3.3 | 1 | 2.3 | |
| CMV-AU | CMV-IgG + Healthy Controls | |||
|---|---|---|---|---|
| n | % | n | % | |
| KIR3DL1H+ | 105 | 86.1 | 37 | 86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yawata, N.; Shirane, M.; Woon, K.; Lim, X.; Tanaka, H.; Kawano, Y.-I.; Yawata, M.; Chee, S.-P.; Siak, J.; Sonoda, K.-H. Molecular Signatures of Natural Killer Cells in CMV-Associated Anterior Uveitis, A New Type of CMV-Induced Disease in Immunocompetent Individuals. Int. J. Mol. Sci. 2021, 22, 3623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073623
Yawata N, Shirane M, Woon K, Lim X, Tanaka H, Kawano Y-I, Yawata M, Chee S-P, Siak J, Sonoda K-H. Molecular Signatures of Natural Killer Cells in CMV-Associated Anterior Uveitis, A New Type of CMV-Induced Disease in Immunocompetent Individuals. International Journal of Molecular Sciences. 2021; 22(7):3623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073623
Chicago/Turabian StyleYawata, Nobuyo, Mariko Shirane, Kaing Woon, Xinru Lim, Hidenori Tanaka, Yoh-Ichi Kawano, Makoto Yawata, Soon-Phaik Chee, Jay Siak, and Koh-Hei Sonoda. 2021. "Molecular Signatures of Natural Killer Cells in CMV-Associated Anterior Uveitis, A New Type of CMV-Induced Disease in Immunocompetent Individuals" International Journal of Molecular Sciences 22, no. 7: 3623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073623