
Study on Demethoxycurcumin as a Promising Approach to Reverse Methicillin-Resistance of Staphylococcus aureus


Abstract
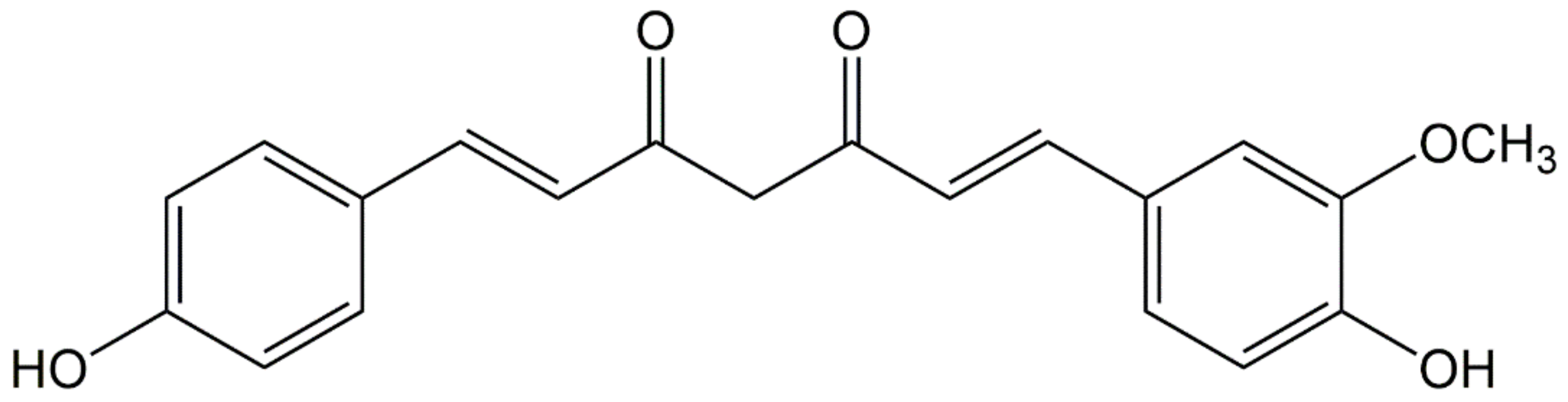
:1. Introduction
2. Results
2.1. The Minimum Inhibitory Concentration (MIC) of DMC and β-Lactam Antibiotics
2.2. Synergistic Effects of DMC and Antibiotics Based on Fractional Inhibitory Concentration Index (FICI)
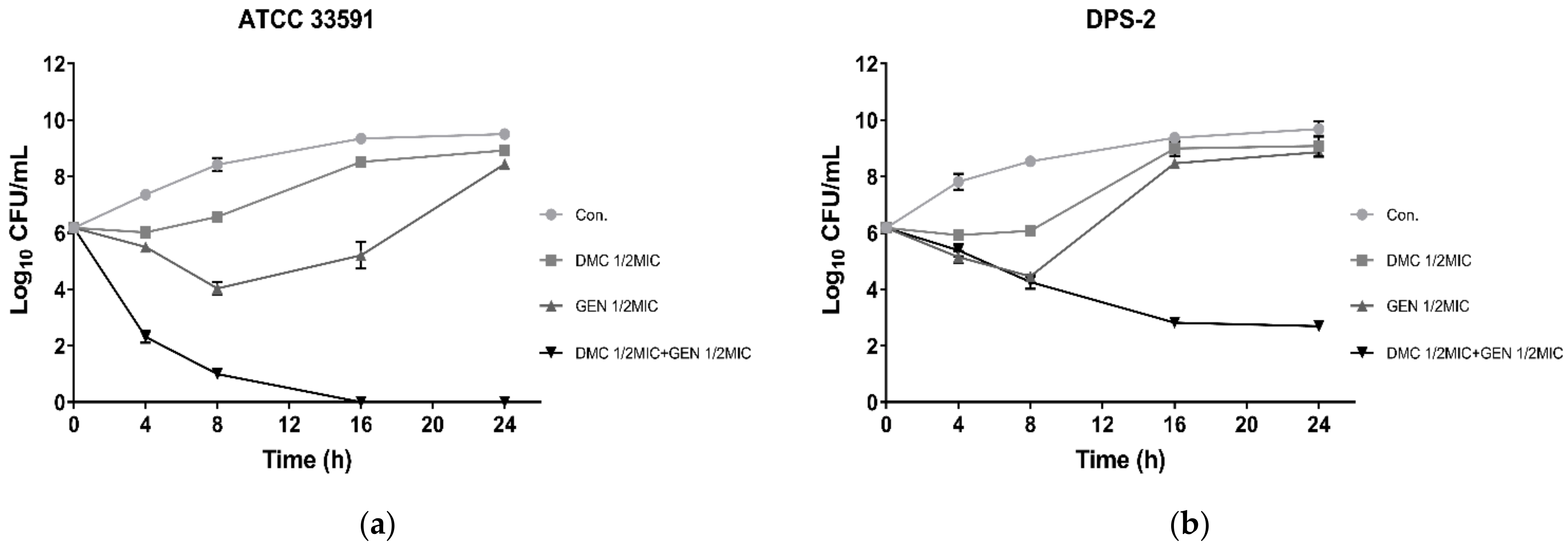
2.3. Time–Kill Assay
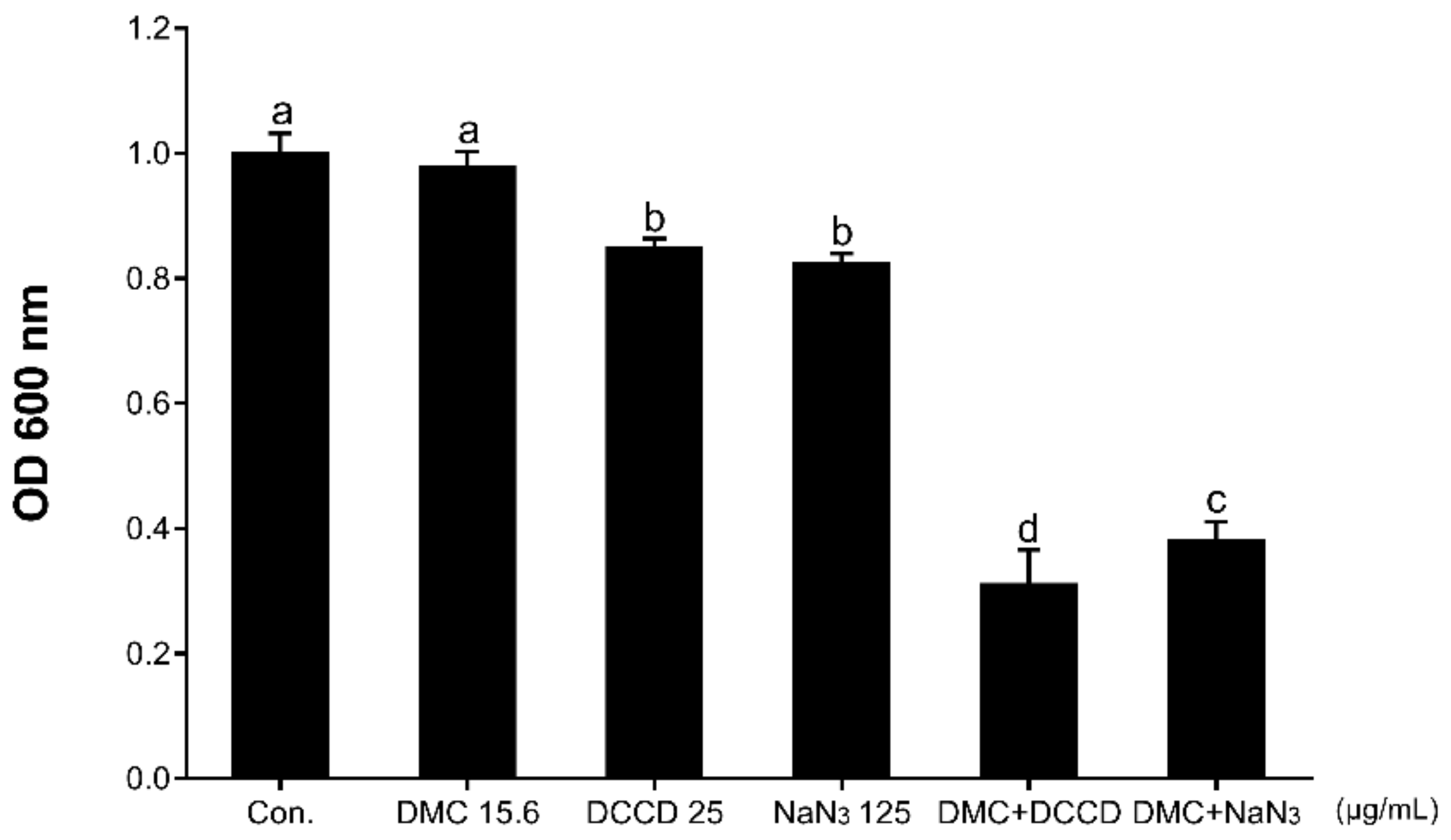
2.4. Antibacterial Activity with Membrane Permeability or ATP Synthase Inhibitor
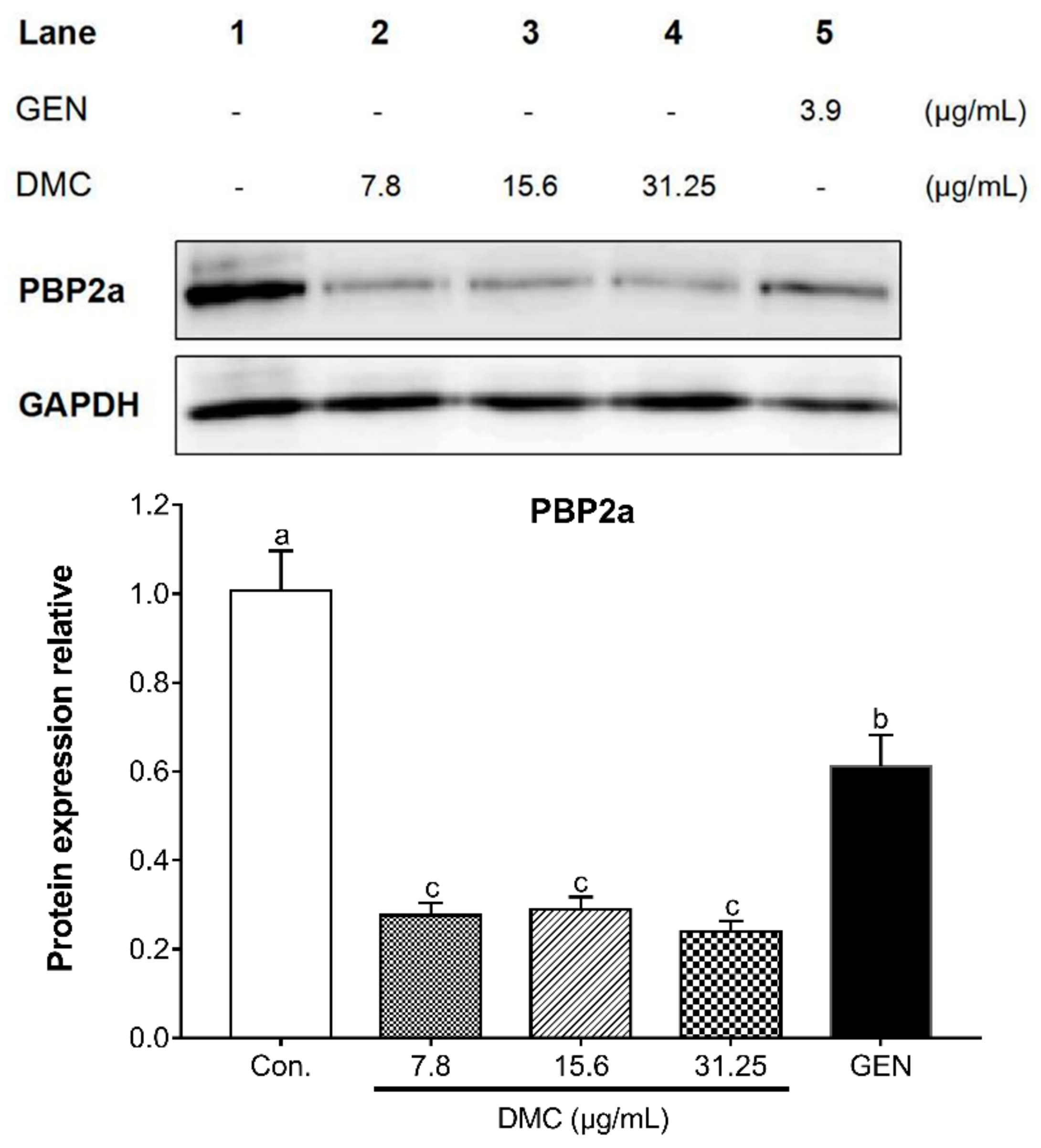
2.5. The Inhibitory Effect of DMC on the Expression of PBP2a Protein and Related Gene in MRSA
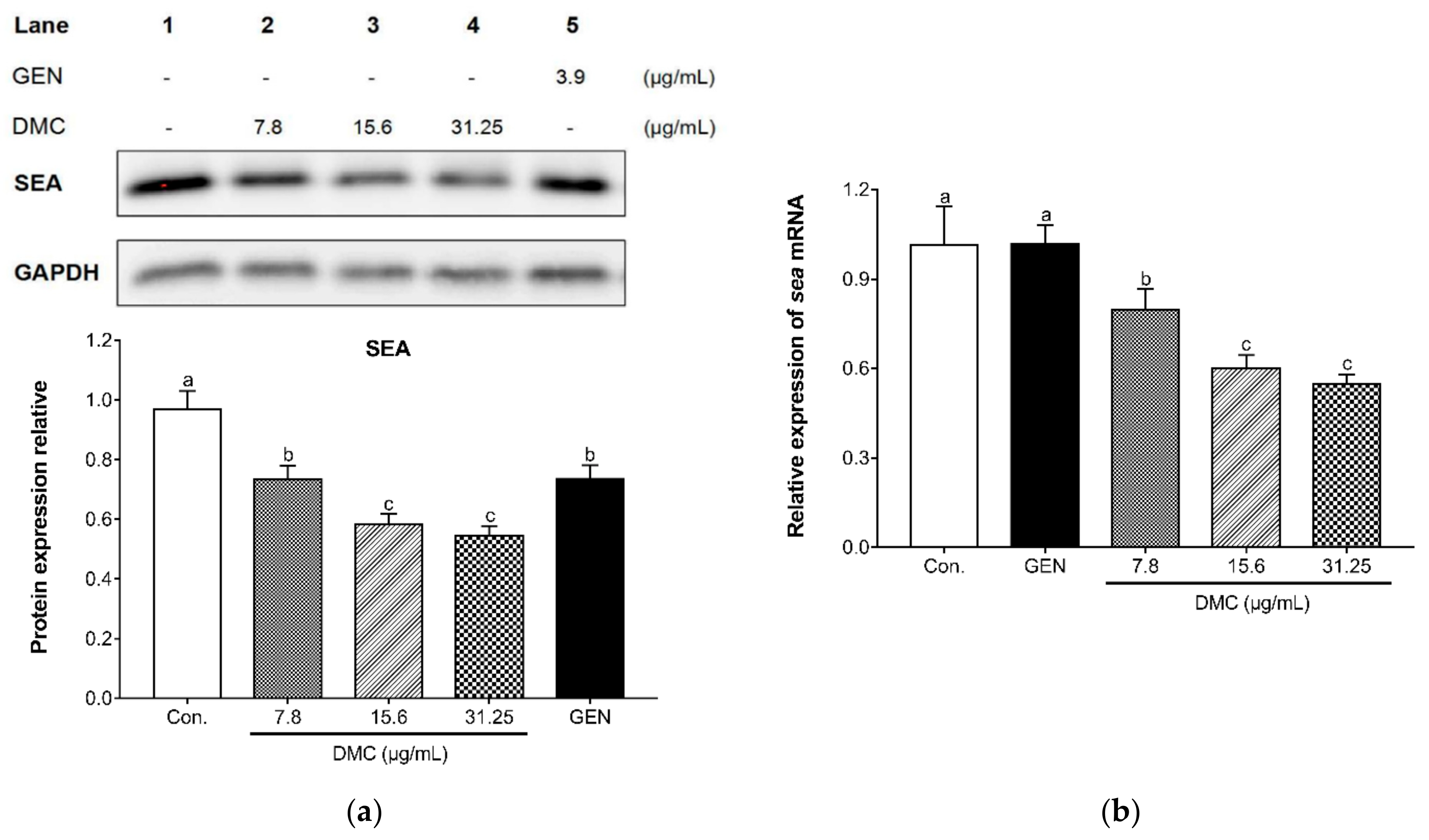
2.6. The Inhibitory Effect of DMC on the Expression of SEA and Related Gene in MRSA
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Reagents and Instruments
4.3. Determination of Minimal Inhibitory Concentration
4.4. Determination of the In Vitro Effects of Combinations of DMC and Antibiotics
4.5. Time–Kill Assay
4.6. Determination of the In Vitro Effects of DMC on Membrane-Permeabilizing Agents and ATP Synthase Inhibitors
4.7. Western Blot Analysis
4.8. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liang, Y.; Tu, C.; Tan, C.; El-Sayed Ahmed, M.A.E.; Dai, M.; Xia, Y.; Liu, Y.; Zhong, L.L.; Shen, C.; Chen, G.; et al. Antimicrobial resistance, virulence genes profiling and molecular relatedness of methicillin-resistant Staphylococcus aureus strains isolated from hospitalized patients in Guangdong Province, China. Infect. Drug Resist. 2019, 12, 447–459. [Google Scholar] [CrossRef] [Green Version]
- Tommasi, R.; Brown, D.G.; Walkup, G.K.; Manchester, J.I.; Miller, A.A. ESKAPEing the labyrinth of antibacterial discovery. Nat. Rev. Drug Discov. 2015, 14, 529–542. [Google Scholar] [CrossRef]
- Gajdács, M. The Continuing Threat of Methicillin-Resistant Staphylococcus aureus. Antibiotics 2019, 8, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savoldi, A.; Azzini, A.M.; Baur, D.; Tacconelli, E. Is there still a role for vancomycin in skin and soft-tissue infections? Curr. Opin. Infect. Dis. 2018, 31, 120–130. [Google Scholar] [CrossRef]
- Xue, L.; Chen, Y.Y.; Yan, Z.; Lu, W.; Wan, D.; Zhu, H. Staphyloxanthin: A potential target for antivirulence therapy. Infect. Drug Resist. 2019, 12, 2151–2160. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Mou, Q.; Xu, X.; Qi, S.; Leung, P.H. Synergistic antibacterial activity between penicillenols and antibiotics against methicillin-resistant Staphylococcus aureus. R. Soc. Open Sci. 2018, 5, 172466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Mokhtar, M.A.; Hetta, H.F. Ambulance vehicles as a source of multidrug-resistant infections: A multicenter study in Assiut City, Egypt. Infect. Drug Resist. 2018, 11, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Astley, R.; Miller, F.C.; Mursalin, M.H.; Coburn, P.S.; Callegan, M.C. An Eye on Staphylococcus aureus Toxins: Roles in Ocular Damage and Inflammation. Toxins 2019, 11, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stryjewski, M.E.; Chambers, H.F. Skin and soft-tissue infections caused by community-acquired methicillin-resistant Staphylococcus aureus. Clin. Infect. Dis. 2008, 46 (Suppl. S5), S368–S377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belluzo, B.S.; Abriata, L.A.; Giannini, E.; Mihovilcevic, D.; Dal Peraro, M.; Llarrull, L.I. An experiment-informed signal transduction model for the role of the Staphylococcus aureus MecR1 protein in beta-lactam resistance. Sci. Rep. 2019, 9, 19558. [Google Scholar] [CrossRef]
- Khan, A.U.; Ali, A.; Danishuddin; Srivastava, G.; Sharma, A. Potential inhibitors designed against NDM-1 type metallo-beta-lactamases: An attempt to enhance efficacies of antibiotics against multi-drug-resistant bacteria. Sci. Rep. 2017, 7, 9207. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.C.; Chen, C.C.; Lu, Y.C.; Lin, T.P.; Chuang, Y.C.; Tang, H.J. Appropriate composites of cefoperazone-sulbactam against multidrug-resistant organisms. Infect. Drug Resist. 2018, 11, 1441–1445. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Ma, G.; Zhu, Y.; Zeng, L.; Ahmad, A.; Wang, C.; Pang, B.; Fang, H.; Zhao, L.; Hao, Q. Active-Site Conformational Fluctuations Promote the Enzymatic Activity of NDM-1. Antimicrob. Agents Chemother. 2018, 62, e01518–e01579. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.H.; Chen, P.Y.; Hu, R.M.; Lin, Y.T.; Li, L.H.; Yang, T.C. Impacts of L1 Promoter Variation and L2 Clavulanate Susceptibility on Ticarcillin-Clavulanate Susceptibility of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2018, 62, e01218–e01222. [Google Scholar] [CrossRef] [Green Version]
- Cabeen, M.T.; Jacobs-Wagner, C. Bacterial cell shape. Nat. Rev. Microbiol. 2005, 3, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Tseng, Y.H.; Deng, F.S.; Lin, J.W.; Tseng, Y.H.; Weng, S.F. Contribution of Phe-7 to Tat-dependent export of beta-lactamase in Xanthomonas campestris. Antimicrob. Agents Chemother. 2012, 56, 3597–3602. [Google Scholar] [CrossRef] [Green Version]
- Aslam, A.; Gajdács, M.; Zin, C.S.; Ab Rahman, N.S.; Ahmed, S.I.; Zafar, M.Z.; Jamshed, S. Evidence of the Practice of Self-Medication with Antibiotics among the Lay Public in Low- and Middle-Income Countries: A Scoping Review. Antibiotics 2020, 9, 597. [Google Scholar] [CrossRef]
- Cheong, W.L.; Tsang, M.S.; So, P.K.; Chung, W.H.; Leung, Y.C.; Chan, P.H. Fluorescent TEM-1 beta-lactamase with wild-type activity as a rapid drug sensor for in vitro drug screening. Biosci. Rep. 2014, 34, e00136. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.L.; Papp-Wallace, K.M.; Bonomo, R.A. Activity of ceftazidime/avibactam against isogenic strains of Escherichia coli containing KPC and SHV beta-lactamases with single amino acid substitutions in the Omega-loop. J. Antimicrob. Chemother. 2015, 70, 2279–2286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandavasi, V.G.; Langan, P.S.; Weiss, K.L.; Parks, J.M.; Cooper, J.B.; Ginell, S.L.; Coates, L. Active-Site Protonation States in an Acyl-Enzyme Intermediate of a Class A beta-Lactamase with a Monobactam Substrate. Antimicrob. Agents Chemother. 2017, 61, e01616–e01636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro, J.M.; Covas, G.; Rausch, D.; Filipe, S.R.; Schneider, T.; Sahl, H.G.; Pinho, M.G. The pentaglycine bridges of Staphylococcus aureus peptidoglycan are essential for cell integrity. Sci. Rep. 2019, 9, 5010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekizuka, T.; Niwa, H.; Kinoshita, Y.; Uchida-Fujii, E.; Inamine, Y.; Hashino, M.; Kuroda, M. Identification of a mecA/mecC-positive MRSA ST1-t127 isolate from a racehorse in Japan. J. Antimicrob. Chemother. 2020, 75, 292–295. [Google Scholar] [CrossRef]
- Tavares, D.A.; Sa-Leao, R.; Miragaia, M.; de Lencastre, H. Large screening of CA-MRSA among Staphylococcus aureus colonizing healthy young children living in two areas (urban and rural) of Portugal. BMC Infect. Dis. 2010, 10, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, J.; Andersen, P.S.; Winstel, V.; Peschel, A. Staphylococcus aureus CC395 harbours a novel composite staphylococcal cassette chromosome mec element. J. Antimicrob. Chemother. 2017, 72, 1002–1005. [Google Scholar] [PubMed] [Green Version]
- Ba, X.; Harrison, E.M.; Lovering, A.L.; Gleadall, N.; Zadoks, R.; Parkhill, J.; Peacock, S.J.; Holden, M.T.; Paterson, G.K.; Holmes, M.A. Old Drugs To Treat Resistant Bugs: Methicillin-Resistant Staphylococcus aureus Isolates with mecC Are Susceptible to a Combination of Penicillin and Clavulanic Acid. Antimicrob. Agents Chemother. 2015, 59, 7396–7404. [Google Scholar] [CrossRef] [Green Version]
- Thalso-Madsen, I.; Torrubia, F.R.; Xu, L.; Petersen, A.; Jensen, C.; Frees, D. The Sle1 Cell Wall Amidase Is Essential for beta-Lactam Resistance in Community-Acquired Methicillin-Resistant Staphylococcus aureus USA300. Antimicrob. Agents Chemother. 2019, 64, e01919–e01931. [Google Scholar] [CrossRef] [Green Version]
- Mwangi, M.M.; Kim, C.; Chung, M.; Tsai, J.; Vijayadamodar, G.; Benitez, M.; Jarvie, T.P.; Du, L.; Tomasz, A. Whole-genome sequencing reveals a link between beta-lactam resistance and synthetases of the alarmone (p)ppGpp in Staphylococcus aureus. Microb. Drug Resist. 2013, 19, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Yoon, E.J.; Kim, D.; Kim, J.W.; Lee, K.J.; Kim, H.S.; Kim, Y.R.; Shin, J.H.; Shin, J.H.; Shin, K.S.; et al. Ceftaroline Resistance by Clone-Specific Polymorphism in Penicillin-Binding Protein 2a of Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2018, 62, e00418–e00485. [Google Scholar] [CrossRef] [Green Version]
- Hou, Z.; Zhou, Y.; Wang, H.; Bai, H.; Meng, J.; Xue, X.; Luo, X. Co-blockade of mecR1/blaR1 signal pathway to restore antibiotic susceptibility in clinical isolates of methicillin-resistant Staphylococcus aureus. Arch. Med. Sci. 2011, 7, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Zhang, J.; Yan, Y.; Yang, M.; Li, C.; Li, J.; Zhong, L.; Gong, Q.; Yu, H. Network Pharmacology-Based Strategy to Investigate the Pharmacologic Mechanisms of Atractylodes macrocephala Koidz. for the Treatment of Chronic Gastritis. Front. Pharmacol. 2019, 10, 1629. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Ban, I.; Lee, H.; Baik, M.Y.; Kim, W. Puffing as a Novel Process to Enhance the Antioxidant and Anti-Inflammatory Properties of Curcuma longa L. (Turmeric). Antioxidants 2019, 8, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mapoung, S.; Suzuki, S.; Fuji, S.; Naiki-Ito, A.; Kato, H.; Yodkeeree, S.; Ovatlarnporn, C.; Takahashi, S.; Limtrakul Dejkriengkraikul, P. Cyclohexanone curcumin analogs inhibit the progression of castration-resistant prostate cancer in vitro and in vivo. Cancer Sci. 2019, 110, 596–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, E.; Lucariello, A.; Contieri, M.; Esposito, T.; De Luca, A.; Guerra, G.; Perna, A. Therapeutic effects of turmeric in several diseases: An overview. Chemico-Biol. Interact. 2019, 310, 108729. [Google Scholar] [CrossRef] [PubMed]
- Chien, M.H.; Yang, W.E.; Yang, Y.C.; Ku, C.C.; Lee, W.J.; Tsai, M.Y.; Lin, C.W.; Yang, S.F. Dual Targeting of the p38 MAPK-HO-1 Axis and cIAP1/XIAP by Demethoxycurcumin Triggers Caspase-Mediated Apoptotic Cell Death in Oral Squamous Cell Carcinoma Cells. Cancers 2020, 12, 703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatamipour, M.; Ramezani, M.; Tabassi, S.A.S.; Johnston, T.P.; Ramezani, M.; Sahebkar, A. Demethoxycurcumin: A naturally occurring curcumin analogue with antitumor properties. J. Cell Physiol. 2018, 233, 9247–9260. [Google Scholar] [CrossRef]
- Hatamipour, M.; Ramezani, M.; Tabassi, S.A.S.; Johnston, T.P.; Sahebkar, A. Demethoxycurcumin: A naturally occurring curcumin analogue for treating non-cancerous diseases. J. Cell Physiol. 2019, 234, 19320–19330. [Google Scholar] [CrossRef]
- Gajdács, M.; Ábrók, M.; Lázár, A.; Burián, K. Increasing relevance of Gram-positive cocci in urinary tract infections: A 10-year analysis of their prevalence and resistance trends. Sci. Rep. 2020, 10, 17658. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, M.; Yu, Y.; Liu, B.; Liu, Y. In Vitro Activity of Ceftaroline And Comparators Against Staphylococcus aureus Isolates: Results From 6 Years Of The ATLAS Program (2012 To 2017). Infect. Drug Resist. 2019, 12, 3349–3358. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Da, F.; Ma, X.; Wang, N.; Wang, Y.; Zhang, H.; Li, M.; Zhou, Y.; Xue, X.; Hou, Z.; et al. Antisense growth inhibition of methicillin-resistant Staphylococcus aureus by locked nucleic acid conjugated with cell-penetrating peptide as a novel FtsZ inhibitor. Antimicrob. Agents Chemother. 2015, 59, 914–922. [Google Scholar] [CrossRef] [Green Version]
- Mun, S.H.; Kang, O.H.; Joung, D.K.; Kim, S.B.; Choi, J.G.; Shin, D.W.; Kwon, D.Y. In vitro anti-MRSA activity of carvone with gentamicin. Exp. Ther. Med. 2014, 7, 891–896. [Google Scholar] [CrossRef]
- Zhou, T.; Li, Z.; Kang, O.H.; Mun, S.H.; Seo, Y.S.; Kong, R.; Shin, D.W.; Liu, X.Q.; Kwon, D.Y. Antimicrobial activity and synergism of ursolic acid 3-O-alpha-L-arabinopyranoside with oxacillin against methicillin-resistant Staphylococcus aureus. Int. J. Mol. Med. 2017, 40, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toei, M.; Noji, H. Single-molecule analysis of F0F1-ATP synthase inhibited by N,N-dicyclohexylcarbodiimide. J. Biol. Chem. 2013, 288, 25717–25726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safo, M.K.; Zhao, Q.; Ko, T.P.; Musayev, F.N.; Robinson, H.; Scarsdale, N.; Wang, A.H.; Archer, G.L. Crystal structures of the BlaI repressor from Staphylococcus aureus and its complex with DNA: Insights into transcriptional regulation of the bla and mec operons. J. Bacteriol. 2005, 187, 1833–1844. [Google Scholar] [CrossRef] [Green Version]
- Llarrull, L.I.; Toth, M.; Champion, M.M.; Mobashery, S. Activation of BlaR1 protein of methicillin-resistant Staphylococcus aureus, its proteolytic processing, and recovery from induction of resistance. J. Biol. Chem. 2011, 286, 38148–38158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, Y.; Itaya, H.; Kitahara, Y.; Theresia, N.M.; Kutukova, E.A.; Yomantas, Y.A.; Date, M.; Kikuchi, Y.; Wachi, M. Double mutation of cell wall proteins CspB and PBP1a increases secretion of the antibody Fab fragment from Corynebacterium glutamicum. Microb. Cell Fact. 2014, 13, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dar, J.A.; Thoker, M.A.; Khan, J.A.; Ali, A.; Khan, M.A.; Rizwan, M.; Bhat, K.H.; Dar, M.J.; Ahmed, N.; Ahmad, S. Molecular epidemiology of clinical and carrier strains of methicillin resistant Staphylococcus aureus (MRSA) in the hospital settings of north India. Ann. Clin. Microbiol. Antimicrob. 2006, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milheirico, C.; de Lencastre, H.; Tomasz, A. Full-Genome Sequencing Identifies in the Genetic Background Several Determinants That Modulate the Resistance Phenotype in Methicillin-Resistant Staphylococcus aureus Strains Carrying the Novel mecC Gene. Antimicrob. Agents Chemother. 2017, 61, e02500–e02516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiramatsu, K. Molecular evolution of MRSA. Microbiol. Immunol. 1995, 39, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, E.; Kuwahara-Arai, K.; Richardson, J.F.; Hiramatsu, K. Distribution of mec regulator genes in methicillin-resistant Staphylococcus clinical strains. Antimicrob. Agents Chemother. 1993, 37, 1219–1226. [Google Scholar] [CrossRef] [Green Version]
- Rosato, A.E.; Kreiswirth, B.N.; Craig, W.A.; Eisner, W.; Climo, M.W.; Archer, G.L. mecA-blaZ corepressors in clinical Staphylococcus aureus isolates. Antimicrob. Agents Chemother. 2003, 47, 1460–1463. [Google Scholar] [CrossRef] [Green Version]
- Black, C.C.; Eberlein, L.C.; Solyman, S.M.; Wilkes, R.P.; Hartmann, F.A.; Rohrbach, B.W.; Bemis, D.A.; Kania, S.A. The role of mecA and blaZ regulatory elements in mecA expression by regional clones of methicillin-resistant Staphylococcus pseudintermedius. Vet. Microbiol. 2011, 151, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Speri, E.; Fishovitz, J.; Mobashery, S. Structure-activity relationship of the cinnamamide family of antibiotic potentiators for methicillin-resistant Staphylococcus aureus (MRSA). MedChemComm 2018, 9, 2008–2016. [Google Scholar] [CrossRef] [PubMed]
- Klitgaard, J.K.; Skov, M.N.; Kallipolitis, B.H.; Kolmos, H.J. Reversal of methicillin resistance in Staphylococcus aureus by thioridazine. J. Antimicrob. Chemother. 2008, 62, 1215–1221. [Google Scholar] [CrossRef]
- Huang, T.M.; Chou, C.C. Methicillin-sensitive and methicillin-resistant Staphylococcus aureus strains and their toxin genes in the nostrils of dogs and workers at an animal shelter. J. Appl. Microbiol. 2019, 126, 1899–1909. [Google Scholar] [CrossRef] [PubMed]
- Koziel, J.; Chmiest, D.; Bryzek, D.; Kmiecik, K.; Mizgalska, D.; Maciag-Gudowska, A.; Shaw, L.N.; Potempa, J. The Janus face of alpha-toxin: A potent mediator of cytoprotection in staphylococci-infected macrophages. J. Innate Immun. 2015, 7, 187–198. [Google Scholar] [CrossRef]
- Pinchuk, I.V.; Beswick, E.J.; Reyes, V.E. Staphylococcal enterotoxins. Toxins 2010, 2, 2177–2197. [Google Scholar] [CrossRef] [Green Version]
- Luna, B.M.; Ershova, K.; Yan, J.; Ulhaq, A.; Nielsen, T.B.; Hsieh, S.; Pantapalangkoor, P.; Vanscoy, B.; Ambrose, P.; Rudin, S.; et al. Adjunctive transferrin to reduce the emergence of antibiotic resistance in Gram-negative bacteria. J. Antimicrob. Chemother. 2019, 74, 2631–2639. [Google Scholar] [CrossRef]
- Lee, Y.S.; Han, S.H.; Lee, S.H.; Kim, Y.G.; Park, C.B.; Kang, O.H.; Keum, J.H.; Kim, S.B.; Mun, S.H.; Seo, Y.S.; et al. The mechanism of antibacterial activity of tetrandrine against Staphylococcus aureus. Foodborne Pathog. Dis. 2012, 9, 686–691. [Google Scholar] [CrossRef]
- Joung, D.K.; Lee, Y.S.; Han, S.H.; Lee, S.W.; Cha, S.W.; Mun, S.H.; Kong, R.; Kang, O.H.; Song, H.J.; Shin, D.W.; et al. Potentiating activity of luteolin on membrane permeabilizing agent and ATPase inhibitor against methicillin-resistant Staphylococcus aureus. Asian Pac. J. Trop. Med. 2016, 9, 19–22. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC (μg/mL) | |||
|---|---|---|---|---|
| OXA | AMP | GEN | DMC | |
| ATCC 25923 | 0.9 | 0.9 | 0.9 | 62.5 |
| ATCC 33591 | 125 | 125 | 7.8 | 62.5 |
| DPS-1 | 62.5 | 62.5 | 125 | 62.5 |
| DPS-2 | 0.9 | 1.9 | 125 | 62.5 |
| CCARM 3090 | 250 | 31.25 | 125 | 62.5 |
| CCARM 3091 | 1000 | 62.5 | 2000 | 62.5 |
| CCARM 3095 | 500 | 31.25 | 250 | 62.5 |
| CCARM 3102 | 250 | 62.5 | 500 | 62.5 |
| Strains | Agents | MIC (μg/mL) | FICI | Outcome | |
|---|---|---|---|---|---|
| Alone | Combination | ||||
| ATCC33591 | DMC | 62.5 | 3.9 | 0.306 | synergy |
| GEN | 7.8 | 1.9 | |||
| DPS-1 | DMC | 62.5 | 7.8 | 0.375 | synergy |
| GEN | 125 | 31.25 | |||
| DPS-2 | DMC | 62.5 | 7.8 | 0.250 | synergy |
| GEN | 125 | 15.6 | |||
| CCARM 3090 | DMC | 62.5 | 7.8 | 0.375 | synergy |
| GEN | 125 | 31.25 | |||
| CCARM 3091 | DMC | 62.5 | 7.8 | 0.375 | synergy |
| GEN | 2000 | 500 | |||
| CCARM 3095 | DMC | 62.5 | 15.6 | 0.375 | synergy |
| GEN | 250 | 31.25 | |||
| CCARM 3102 | DMC | 62.5 | 3.9 | 0.312 | synergy |
| GEN | 500 | 125 | |||
| Gene | Primer Sequence |
|---|---|
| mecA | |
| Forward | 5′-GCAATCGCTAAAGAACTAAG-3′ |
| Reverse | 5′-AATGGGACCAACATAACCTA-3′ |
| mecR1 | |
| Forward | 5′-ACACGACTTCTTCGGTTAG-3′ |
| Reverse | 5′-GTACAATTTGGGATTTCACT-3′ |
| blaZ | |
| Forward | 5′-AGAGATTTGCCTATGCTTCA-3′ |
| Reverse | 5′-AGTATCTCCGCTTTTATTATTT-3′ |
| blaR1 | |
| Forward | 5′-ACAATGAAGTAGAAGCCGATAGAT-3′ |
| Reverse | 5′-GTCGGTCAAGTCCAAACA-3′ |
| sea | |
| Forward | 5′-ATGGTGCTTATTATGGTTATC-3′ |
| Reverse | 5′-CGTTTCCAAAGGTACTGTATT-3′ |
| 16S | |
| Forward | 5′-ACTCCTACGGGAGGCAGCAG-3′ |
| Reverse | 5′-ATTACCGCGGCTGCTGG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.-Q.; Kang, O.-H.; Kwon, D.-Y. Study on Demethoxycurcumin as a Promising Approach to Reverse Methicillin-Resistance of Staphylococcus aureus. Int. J. Mol. Sci. 2021, 22, 3778. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073778
Li Q-Q, Kang O-H, Kwon D-Y. Study on Demethoxycurcumin as a Promising Approach to Reverse Methicillin-Resistance of Staphylococcus aureus. International Journal of Molecular Sciences. 2021; 22(7):3778. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073778
Chicago/Turabian StyleLi, Qian-Qian, Ok-Hwa Kang, and Dong-Yeul Kwon. 2021. "Study on Demethoxycurcumin as a Promising Approach to Reverse Methicillin-Resistance of Staphylococcus aureus" International Journal of Molecular Sciences 22, no. 7: 3778. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073778