
The Fibronectin Expression Determines the Distinct Progressions of Malignant Gliomas via Transforming Growth Factor-Beta Pathway

Abstract
:1. Introduction
2. Results
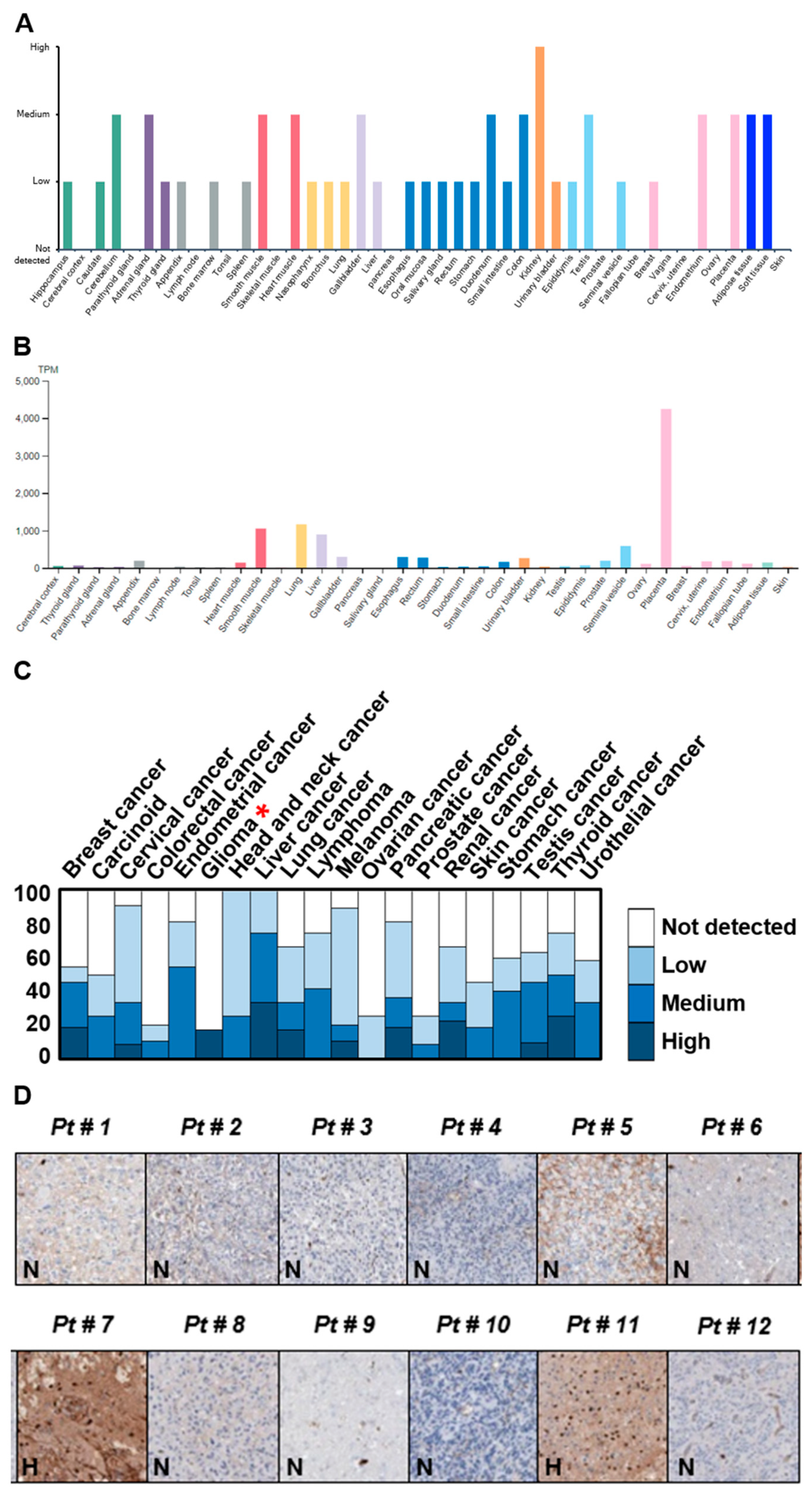
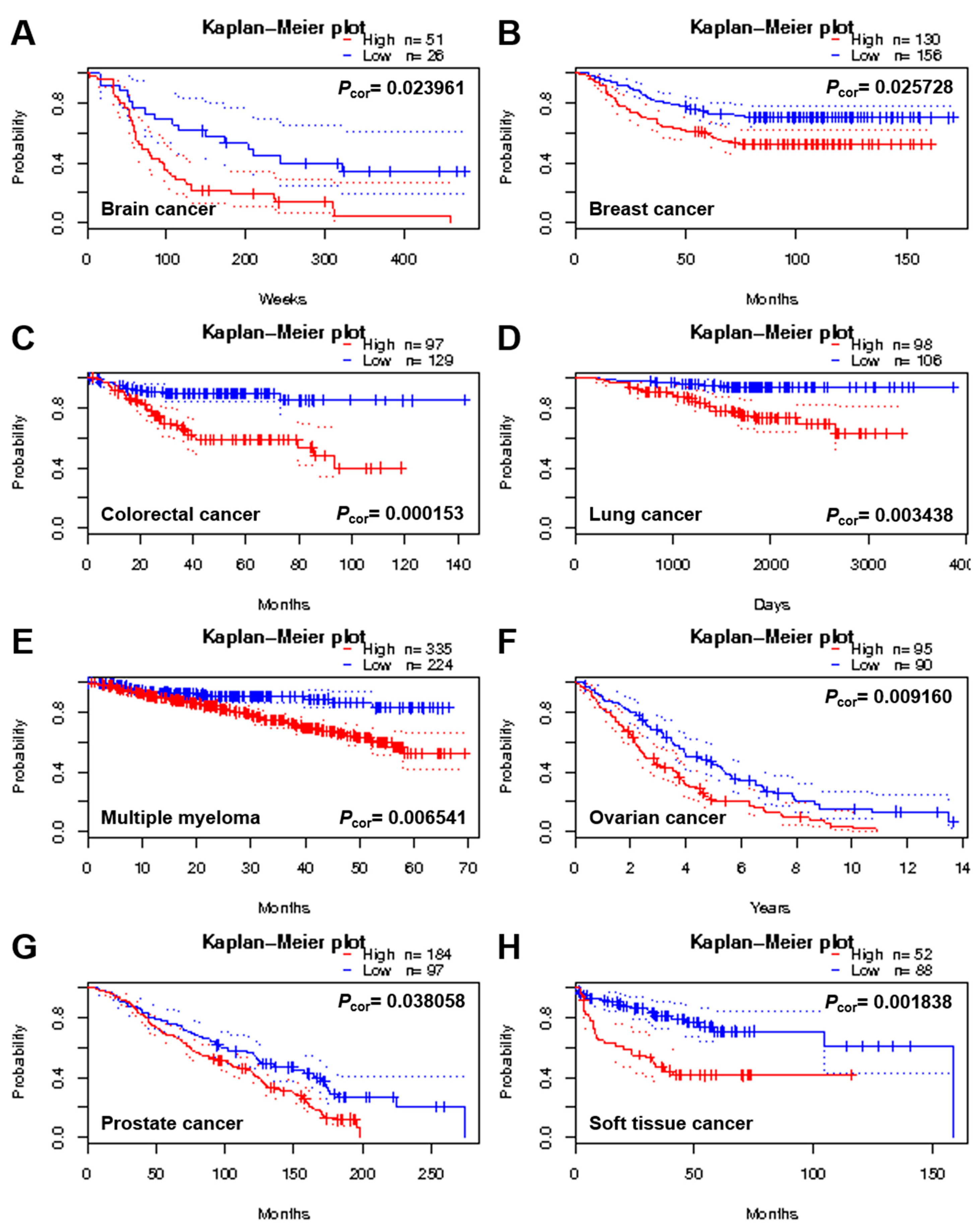
2.1. Identification of Potential Protein Biomarker for GBM
2.2. Recruitment of Specific Brain Tumor Glioma Specimens
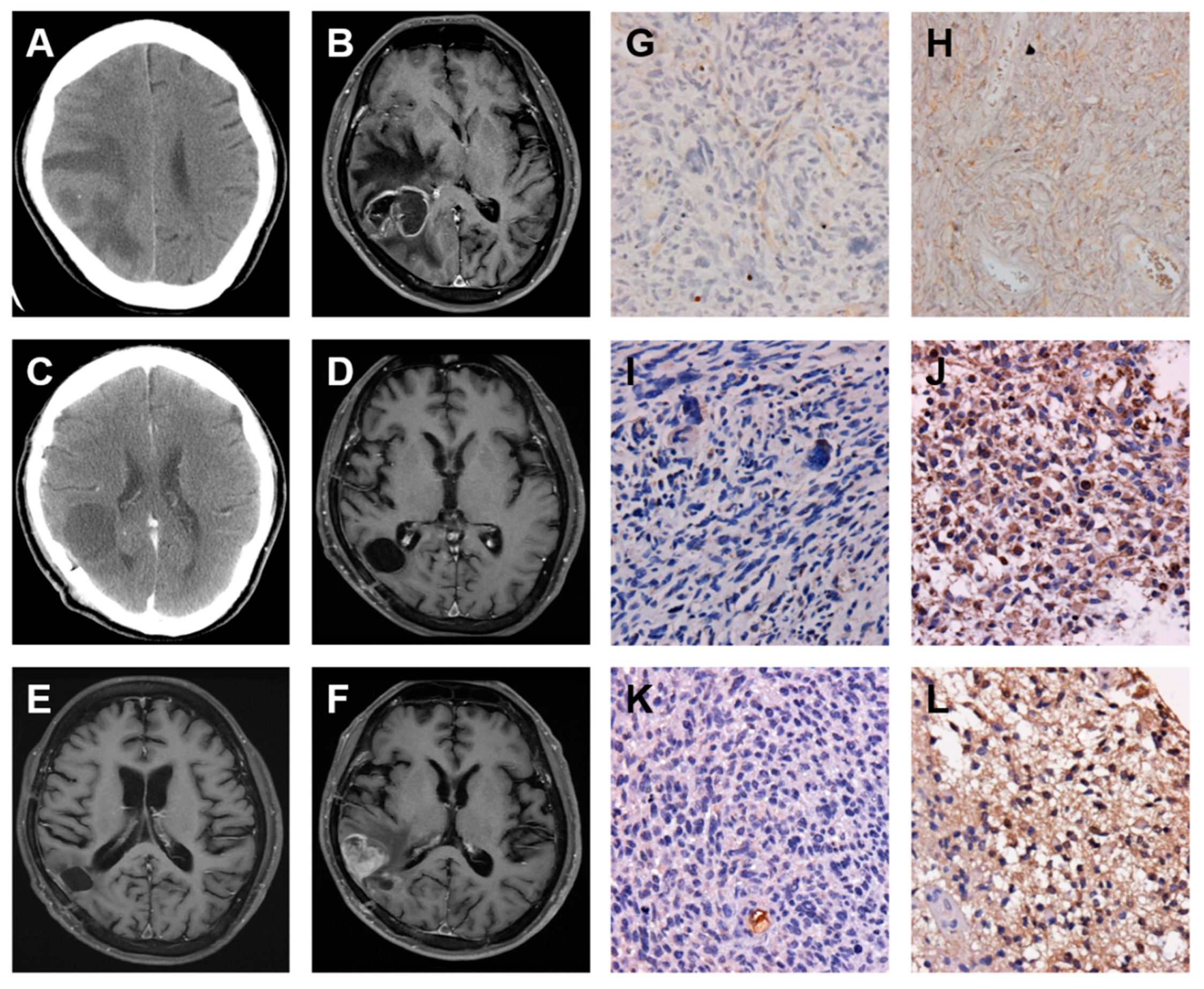
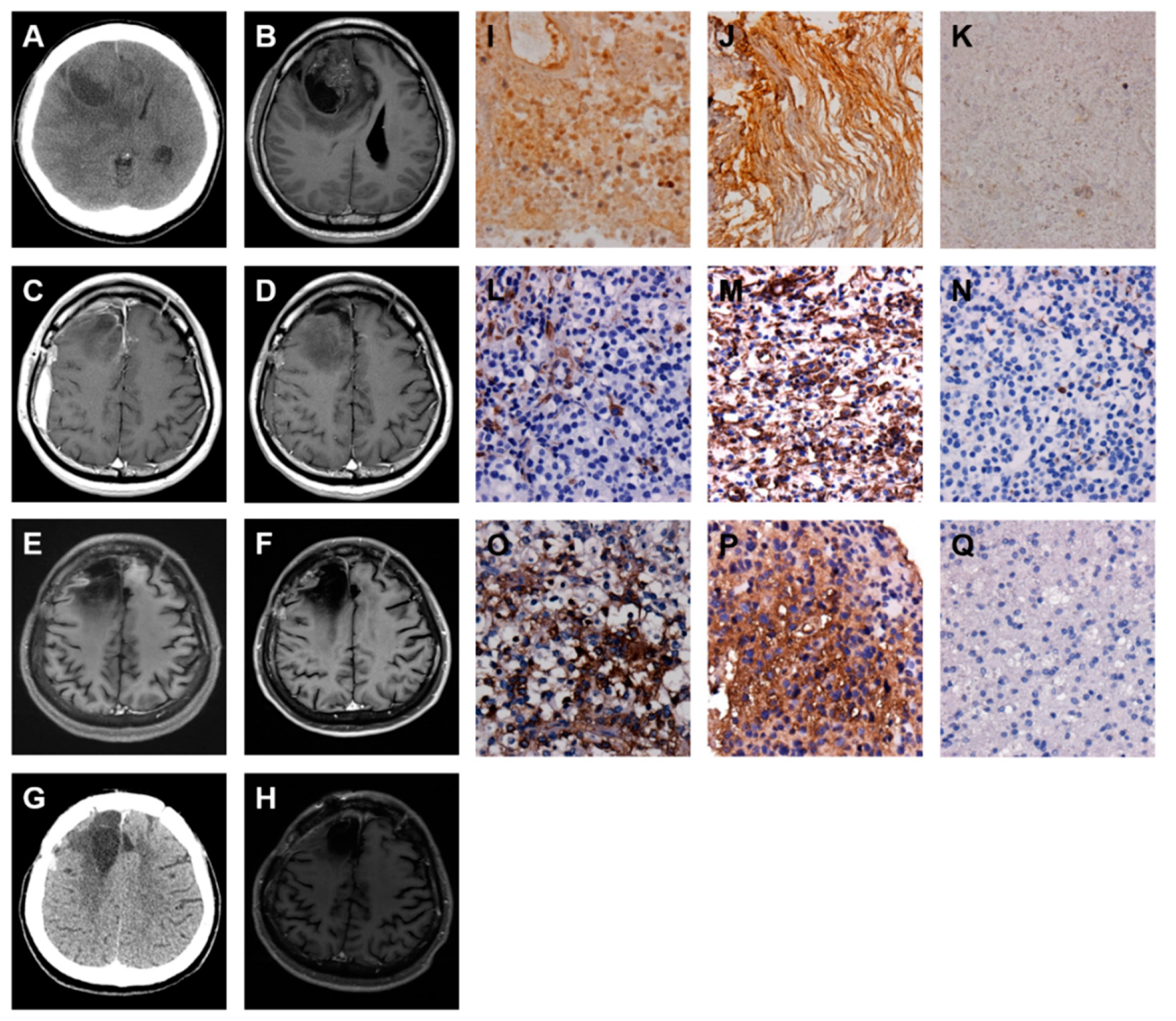
2.3. A Low-Grade Astrocytoma with Local GBM Transformation
2.4. A GBM with Local Recurrence
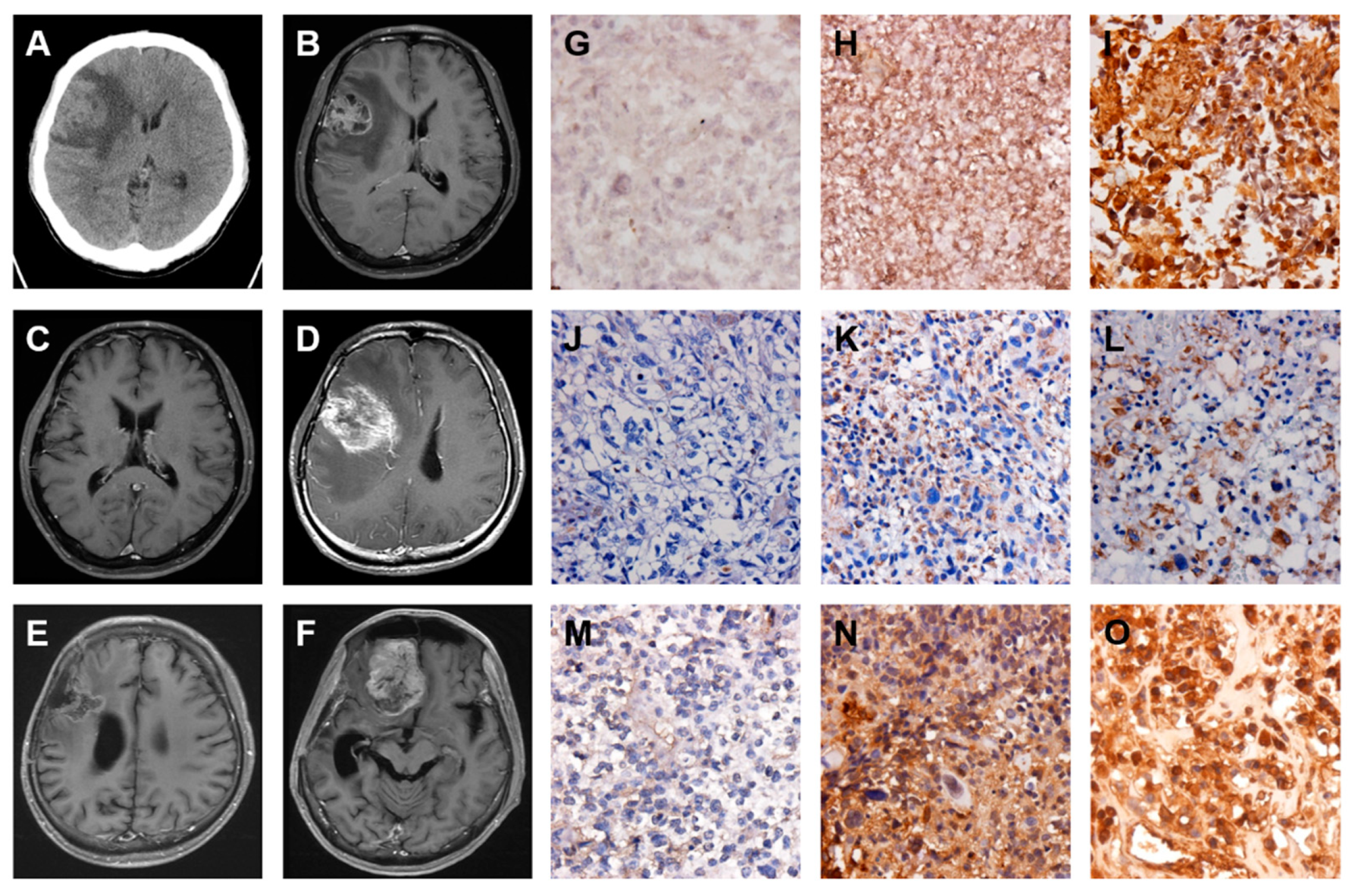
2.5. A GBM with Local Recurrence and Then Remote Brain Metastasis
2.6. A GBM with Spinal Metastasis
2.7. A GBM with Local Recurrence and Then Low-Grade Astrocytoma Progression
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Web-Based Open Megadata Minings
5.2. Patients and Specimens
5.3. Immunohistochemistry (IHC) Staining
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| α-SMA | smooth muscle α-actin |
| CCRT | concomitant medication of chemo-radio-therapy |
| C-IV | type IV collagen |
| CMFMC | Chi Mei Foundation Medical Center |
| CNS | central nervous system |
| CT | computerized tomography |
| ECM | extracellular matrix |
| EMT | epithelial–mesenchymal transition |
| FAK | focal adhesion kinase |
| FN | fibronectin |
| GBM | glioblastoma multiforme |
| GFAP | glial fibrillary acidic protein |
| GMT | glial to mesenchymal transition |
| IHC | immunohistochemical |
| LM | laminin |
| MGMT | O-6-methylguanine-DNA methyltransferase |
| MET | mesenchymal–epithelial transition |
| MMP | matrix metalloprotease |
| MRI | magnetic resonance imaging |
| RT | radio-therapy |
| RT-PCR | reverse transcription–polymerase chain reaction |
| SFK | Src family kinases |
| TGF-β | transforming growth factor-beta |
| TMZ | temozolamide |
| TN-C | tenascin-C |
| VIM | vimentin |
References
- Ostrom, Q.T.; Gittleman, H.; Fulop, J.; Liu, M.; Blanda, R.; Kromer, C.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS statistical report: Primary brain and central nervous system tumors diagnosed in the United States in 2008–2012. Neuro. Oncol. 2015, 17, iv1–iv62. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, Q.T.; de Blank, P.M.; Kruchko, C.; Petersen, C.M.; Liao, P.; Finlay, J.L.; Stearns, D.S.; Wolff, J.E.; Wolinsky, Y.; Letterio, J.; et al. Alex’s Lemonade stand foundation infant and childhood primary brain and central nervous system tumors diagnosed in the United States in 2007–2011. Neuro. Oncol. 2015, 16, x1–x36. [Google Scholar] [CrossRef] [PubMed]
- Affronti, M.L.; Heery, C.R.; Herndon, J.E., 2nd; Rich, J.N.; Reardon, D.A.; Desjardins, A.; Vredenburgh, J.J.; Friedman, A.H.; Bigner, D.D.; Friedman, H.S. Overall survival of newly diagnosed glioblastoma patients receiving carmustine wafers followed by radiation and concurrent temozolomide plus rotational multiagent chemotherapy. Cancer 2009, 115, 3501–3511. [Google Scholar] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef]
- Ammirati, M.; Galicich, J.H.; Arbit, E.; Liao, Y. Reoperation in the treatment of recurrent intracranial malignant gliomas. Neurosurgery 1987, 21, 607–614. [Google Scholar] [CrossRef]
- Choucair, A.K.; Levin, V.A.; Gutin, P.H.; Davis, R.L.; Silver, P.; Edwards, M.S.B.; Wilson, C.B. Development of multiple lesions during radiation therapy and chemotherapy in patients with gliomas. J. Neurosurg. 1986, 65, 654–658. [Google Scholar] [CrossRef]
- Gaspar, L.E.; Fisher, B.J.; Macdonald, D.R.; LeBer, D.V.; Halperin, E.C.; Schold, S.C., Jr.; Cairncross, J.G. Supratentorial malignant glioma: Patterns of recurrence and implications for external beam local treatment. Int. J. Radiat. Oncol. Biol. Phys. 1992, 24, 55–57. [Google Scholar] [CrossRef]
- Halperin, E.C.; Burger, P.C.; Bullard, D.E. The fallacy of the localized supratentorial malignant glioma. Int. J. Radiat. Oncol. Biol. Phys. 1988, 15, 505–509. [Google Scholar] [CrossRef]
- Lee, S.W.; Fraass, B.A.; Marsh, L.H.; Herbort, K.; Gebarski, S.S.; Martel, M.K.; Radany, E.H.; Lichter, A.S.; Sandler, H.M. Patterns of failure following high-dose 3-D conformal radiotherapy for high-grade astrocytomas: A quantitative dosimetric study. Int. J. Radiat. Oncol. Biol. Phys. 1999, 43, 79–88. [Google Scholar] [CrossRef]
- Lun, M.; Lok, E.; Gautam, S.; Wu, E.; Wong, E.T. The natural history of extracranial metastasis from glioblastoma multiforme. J. Neurooncol. 2011, 105, 261–273. [Google Scholar] [CrossRef]
- Goldbrunner, R.H.; Bernstein, J.J.; Tonn, J.C. Cell-extracellular matrix interaction in glioma invasion. Acta Neurochir. 1999, 141, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Perris, R. The extracellular matrix in neural crest-cell migration. Trends Neurosci. 1997, 20, 23–30. [Google Scholar] [CrossRef]
- Lin, T.C.; Yang, C.H.; Chang, W.T.; Lin, Y.R.; Cheng, H.C. Fibronectin in cancer: Friend or foe. Cells 2020, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Giese, A.; Westphal, M. Glioma invasion in the central nervous system. Neurosurgery 1996, 39, 235–252. [Google Scholar] [CrossRef]
- Huang, L.T.; Tsai, C.L.; Huang, S.H.; Chang, M.M.; Chang, W.T.; Cheng, L.H.; Cheng, H.C. Depleting rhoA/stress fiber-organized fibronectin matrices on tumor cells non-autonomously aggravates fibroblast-driven tumor cell growth. Int. J. Mol. Sci. 2020, 21, 8272. [Google Scholar] [CrossRef]
- Sheppard, A.M.; Brunstrom, J.E.; Thornton, T.N.; Gerfen, R.W.; Broekelmann, T.J.; McDonald, J.A.; Pearlman, A.L. Neuronal production of fibronectin in the cerebral during migration and layer formation is unique to specific cortical domains. Dev. Biol. 1995, 172, 504–518. [Google Scholar] [CrossRef] [Green Version]
- Kochi, N.; Tani, E.; Morimura, T.; Nagaki, T. Immunohistochemical study of fibronectin in human glioma and meningioma. Acta Neuropathol. 1983, 59, 119–126. [Google Scholar] [CrossRef]
- Morris, C.S.; Esiri, M.M. Immunocytochemical study of macrophages and microglia cells and extracellular matrix components in human CNS disease. J. Neurol. Sci. 1991, 101, 47–58. [Google Scholar] [CrossRef]
- Chintala, S.K.; Sawaya, R.; Gokaslan, Z.L.; Fuller, G.; Rao, J.S. Immunohistochemical localization of extracellular matrix proteins in human glioma, both in vivo and in vitro. Cancer Lett. 1996, 101, 107–114. [Google Scholar] [CrossRef]
- Ohnishi, T.; Arita, N.; Hiraga, S.; Higuchi, M.; Hayakawa, T. Human malignant glioma cells migrate to fibronectin and laminin: Role of extracellular matrix components in glioma cell invasion. In Biological Aspects of Brain Tumors; Tabuchi, K., Ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA; Tokyo, Japan, 1991; pp. 408–415. [Google Scholar]
- Friedlander, D.R.; Zagzag, D.; Shiff, B.; Cohen, H.; Allen, J.C.; Kelly, P.J.; Grumet, M. Migration of brain tumor cells on extracellular matrix proteins in vitro correlates with tumor type and grade and involves αV and ß1 integrins. Cancer Res. 1996, 56, 1939–1947. [Google Scholar] [PubMed]
- Enam, S.A.; Rosenblum, M.L.; Edvardsen, K. Role of extracellular matrix in tumor invasion: Migration of glioma cells along fibronectin-positive mesenchymal cell process. Neurosurgery 1998, 42, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Weber, K. Intermediate filaments: Structure, dynamics, function, and disease. Annu. Rev. Biochem. 1994, 63, 345–382. [Google Scholar] [CrossRef]
- Steinert, P.M.; Roop, D.R. Molecular and cellular biology of intermediate filaments. Annu. Rev. Biochem. 1988, 57, 593–625. [Google Scholar] [CrossRef]
- Green, K.J.; Bohringer, M.; Gocken, T.; Jones, J.C. Intermediate filament associated proteins. Adv. Protein Chem. 2005, 70, 143–202. [Google Scholar] [PubMed]
- Chen, Y.R.; Juan, H.F.; Huang, H.C.; Huang, H.H.; Lee, Y.J.; Liao, M.Y.; Tseng, C.W.; Lin, L.L.; Chen, J.Y.; Wang, M.J. Quantitative proteomic and genomic profiling reveals metastasis-related protein expression patterns in gastric cancer cells. J. Proteome Res. 2006, 5, 2727–2742. [Google Scholar] [CrossRef]
- Chang, H.Y.; Lee, C.H.; Li, Y.S.; Huang, J.T.; Lan, S.H.; Wang, Y.F.; Lai, W.W.; Wang, Y.C.; Lin, Y.-J.; Liu, H.-S.; et al. MicroRNA-146a suppresses tumor malignancy via targeting vimentin in esophageal squamous cell carcinoma cells with lower fibronectin membrane assembly. J. Biomed. Sci. 2020, 27, 102. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, M.; Weksler, B.; Tsuruta, D.; Goldman, R.D.; Yoon, K.J.; Hopkinson, S.B.; Flitney, F.W.; Jones, J.C.R. Structure and function of a vimentin-associated matrix adhesion in endothelial cells. Mol. Biol. Cell 2001, 12, 85–100. [Google Scholar] [CrossRef] [Green Version]
- Gilles, C.; Polette, M.; Zahm, J.M.; Tournier, J.M.; Volders, L.; Foidart, J.M.; Birembaut, P. Vimentin contributes to human mammary epithelial cell migration. J. Cell Sci. 1999, 112, 4615–4625. [Google Scholar]
- Hirano, H.; Maeda, H.; Takeuchi, Y.; Susaki, Y.; Kobayashi, R.; Hayashi, A.; Ose, N.; Yamaguchi, T.; Yokota, S.; Mori, M. Lymphatic invasion of micropapillary cancer cells in associated with a poor prognosis of pathological stage IA lung adenocarcinomas. Oncol. Lett. 2014, 8, 1107–1111. [Google Scholar] [CrossRef]
- Dauphin, M.; Barbe, C.; Lemaire, S.; Nawrocki-Raby, B.; Lagonotte, E.; Delepine, G.; Birembaut, P.; Gilles, C.; Polette, M. Vimentin expression predicts the occurrence of metastases in non-small cell lung carcinomas. Lung Cancer 2013, 81, 117–122. [Google Scholar] [CrossRef]
- Al-Saad, S.; Al-Shibli, K.; Donnem, T.; Persson, M.; Bremnes, R.M.; Busund, L.T. The prognostic impact of NF-kappaB p105, vimentin, E-cadherin and Par6 expression in epithelial and stromal compartment in non-small-cell lung cancer. Br. J. Cancer 2008, 99, 1476–1483. [Google Scholar] [CrossRef] [Green Version]
- Islam, S.; Kim, J.B.; Trendel, J.; Wheelock, M.J.; Johnson, K.R. Vimentin expression in human squamous carcinoma cells: Relationship with phenotypic changes and cadherin-based cell adhesion. J. Cell Biochem. 2000, 78, 141–150. [Google Scholar] [CrossRef]
- Liu, L.K.; Jiang, X.Y.; Zhou, X.X.; Wang, D.M.; Song, X.L.; Jiang, H.B. Upregulation of vimentin and aberrant expression of E-cadherin/b-catenin complex in oral squamous cell carcinomas: Correlation with the clinicopathological features and patient outcome. Mod. Pathol. 2010, 23, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Satelli, A.; Li, S. Vimentin in cancer and its potential as a molecular target for cancer therapy. Cell Mol. Life Sci. 2011, 68, 3033–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R. EMT: When epithelial cells decide to become mesenchymal-like cells. J. Clin. Investig. 2009, 119, 1417–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moustakas, A.; Heldin, C.H. Signaling networks guiding epithelial-mesenchymal transitions during embryogenesis and cancer progression. Cancer Sci. 2007, 98, 1512–1520. [Google Scholar] [CrossRef]
- Miettinen, P.J.; Ebner, R.; Lopez, A.R.; Derynck, R. TGF-β-induced transdifferentiation of mammary epithelial cells to mesenchymal cells: Involvement of type I receptors. J. Cell Biol. 1994, 127, 2021–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavadil, J.; Böttinger, E.P. TGF-β and epithelial-to-mesenchymal transitions. Oncogene 2005, 24, 5764–5774. [Google Scholar] [CrossRef] [Green Version]
- Derynck, R.; Akhurst, R.J.; Balmain, A. TGF-β signaling in tumor suppression and cancer progression. Nat. Genet. 2001, 29, 117–129. [Google Scholar] [CrossRef]
- Iwadate, Y.; Matsutani, T.; Hirono, S.; Shinozaki, N.; Saeki, N. Transforming growth factor-β and stem cell markers are highly expressed around necrotic areas in glioblastoma. J. Neurooncol. 2016, 129, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Schwarzbauer, J. Mammary epithelial cell interactions with fibronectin stimulate epithelial-mesenchymal transition. Oncogene 2014, 33, 1649–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakae, S.; Kato, T.; Murayama, K.; Sasaki, H.; Abe, M.; Kumon, M.; Kumai, T.; Yamashiro, K.; Inamasu, J.; Hasegawa, M. Remote intracranial recurrence of IDH mutant gliomas is associated with TP53 mutations and an 8q gain. Oncotarget 2017, 8, 84729–84742. [Google Scholar] [CrossRef] [Green Version]
- Kyritsis, A.P.; Levin, V.A.; Yung, W.K.; Leeds, N.E. Imaging patterns of multifocal gliomas. Eur. J. Radiol. 1993, 16, 163–170. [Google Scholar] [CrossRef]
- Dean, B.L.; Drayer, B.P.; Bird, C.R.; Flom, R.A.; Hodak, J.A.; Coons, S.W.; Carey, R.G. Gliomas: Classification with MR imaging. Radiology 1990, 174, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Shimura, T.; Hirano, A.; Nakasu, S.; Llena, J.F.; Leeds, N.E. Multifocal glioma of the brain—an autopsy case. No Shinkei Geka 1986, 14, 97–101. [Google Scholar]
- Ralte, A.M.; Sharma, M.C.; Karak, A.K.; Mehta, V.S.; Sarkar, C. Clinicopathological features, MIB-1 labeling index and apoptotic index in recurrent astrocytic tumors. Pathol. Oncol. Res. 2001, 7, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, C.; Ralte, A.M.; Sharma, M.C.; Mehta, V.S. Recurrent astrocytic tumours—A study of p53 immunoreactivity and malignant progression. Br. J. Neurosurg. 2002, 16, 335–342. [Google Scholar] [CrossRef]
- Li, R.; Chen, X.; You, Y.; Wang, X.; Liu, Y.; Hu, Q.; Yan, W. Comprehensive portrait of recurrent glioblastoma multiforme in molecular and clinical characteristics. Oncotarget 2015, 6, 30968–30974. [Google Scholar] [CrossRef] [Green Version]
- Cloughesy, T.F.; Cavenee, W.K.; Mischel, P.S. Glioblastoma: From molecular pathology to targeted treatment. Annu. Rev. Pathol. 2014, 9, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Hillan, K.J.; Novotny, W. Bevacizumab (Avastin), a humanized anti-VEGF monoclonal antibody for cancer therapy. Biochem. Biophys. Res. Commun. 2005, 333, 328–335. [Google Scholar] [CrossRef]
- Litak, J.; Grochowski, C.; Litak, J.; Osuchowska, I.; Gosik, K.; Radzikowska, E.; Kamieniak, P.; Rolinski, J. TLR-4 signaling vs. immune checkpoints, miRNAs molecules, cancer stem cells, and wingless-signaling interplay in glioblastoma multiforme—future perspectives. Int. J. Mol. Sci. 2020, 21, 3114. [Google Scholar] [CrossRef] [PubMed]
- Germanò, A.; Galatioto, S.; Caffo, M.; Caruso, G.; La Torre, D.; Cardia, E.; Tomasello, F. Immunohistochemical tenascin-C expression in paediatric supratentorial glioblastoma multiforme. Child’s Nerv. Syst. 2000, 16, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Lee, S.H.; Liao, I.C.; Huang, S.H.; Cheng, H.C.; Liao, P.C. Secretomic analysis identifies alpha-1 antitrypsin (A1AT) as a required protein in cancer cell migration, invasion, and pericellular fibronectin assembly for facilitating lung colonization of lung adenocarcinoma cells. Mol. Cell Proteom. 2012, 11, 1320–1339. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Carraher, C.; Schwarzbauer, J.E. Assembly of fibronectin extracellular matrix. Annu. Rev. Cell Dev. Biol. 2010, 26, 397–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harburger, D.S.; Calderwood, D.A. Integrin signalling at a glance. J. Cell Sci. 2009, 122, 159–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.J.; Lin, J.F.; Cheng, L.H.; Chang, W.T.; Kao, Y.H.; Chang, M.M.; Wang, B., Jr.; Cheng, H.C. Pterostilbene prevents AKT-ERK axis-mediated polymerization of surface fibronectin on suspended lung cancer cells independently of apoptosis and suppresses metastasis. J. Hematol. Oncol. 2017, 10, 72. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.C.; Liao, Y.C.; Chang, W.T.; Yang, C.H.; Cheng, L.H.; Cheng, M.; Cheng, H.C. The establishment of a lung colonization assay for circulating tumor cell visualization in lung tissues. J. Vis. Exp. 2018, 136, e56761. [Google Scholar]
- Ruoslahti, E. Fibronectin and its integrin receptors in cancer. Adv. Cancer Res. 1999, 76, 1–20. [Google Scholar]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [Green Version]
- Rozario, T.; DeSimone, D.W. The extracellular matrix in development and morphogenesis: A dynamic view. Dev. Biol. 2010, 341, 126–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnishi, T.; Hiraga, S.; Izumoto, S.; Matsumura, H.; Kanemura, Y.; Arita, N.; Hayakawa, T. Role of fibronectin-stimulated tumor cell migration in glioma invasion in vivo: Clinical significance of fibronectin and fibronectin receptor expressed in human glioma tissues. Clin. Exp. Metastasis 1998, 16, 729–741. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, S.S.; Quah, M.Y.; Nielsen, S.; Atkins, J.; Au, G.G.; Cairns, M.J.; Nahar, P.; Lombard, J.M.; Tanwar, P.S. Inhibition of extracellular matrix mediated TGF-β signalling suppresses endometrial cancer metastasis. Oncotarget 2017, 8, 71400–71417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griggs, L.A.; Hassan, N.T.; Malik, R.S.; Griffin, B.P.; Martinez, B.A.; Elmore, L.W.; Lemmon, C.A. Fibronectin fibrils regulate TGF-β1-induced Epithelial-Mesenchymal Transition. Matrix Biol. 2017, 60–61, 157–175. [Google Scholar] [CrossRef]
- Kubelt, C.; Hattermann, K.; Sebens, S.; Mehdorn, H.M.; Held-Feindt, J. Epithelial-to-mesenchymal transition in paired human primary and recurrent glioblastomas. Int. J. Oncol. 2015, 46, 2515–2525. [Google Scholar] [CrossRef] [Green Version]
- Kahlert, U.D.; Maciaczyk, D.; Doostkam, S.; Orr, B.A.; Simons, B.; Bogiel, T.; Reithmeier, T.; Prinz, M.; Schubert, J.; Niedermann, G.; et al. Activation of canonical WNT/β-catenin signaling enhances in vitro motility of glioblastoma cells by activation of ZEB1 and other activators of epithelial-to-mesenchymal transition. Cancer Lett. 2012, 325, 42–53. [Google Scholar] [CrossRef]
- Brabletz, T. To differentiate or not-routes towards metastasis. Nat. Rev. Cancer 2012, 12, 425–436. [Google Scholar] [CrossRef]
- Mahabir, R.; Tanino, M.; Elmansuri, A.; Wang, L.; Kimura, T.; Itoh, T.; Ohba, Y.; Nishihara, H.; Shirato, H.; Tsuda, M.; et al. Sustained elevation of Snail promotes glial-mesenchymal transition after irradiation in malignant glioma. Neuro Oncol. 2014, 16, 671–685. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Age/Sex | 1st Surgery | 2nd Surgery | 3rd Surgery | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Path | Date | Location | Path | Date | Location | Path | Date | Location | ||
| I | 40/F | LGA | October 2010 | R temporal | GBM | May 2013 | R temporal | |||
| II | 52/M | GBM | July 2009 | R parietal | GBM | December 2010 | R parietal | |||
| III | 44/M | GBM | August 2010 | R frontal | GBM | July 2012 | R frontal | GBM | June 2013 | R frontal |
| IV | 35/M | GBM | May 2013 | L temporal | GBM | August 2014 | T spine | |||
| V | 50/M | GBM | December 2009 | R frontal | GBM | January 2016 | R frontal | LGA | June 2016 | R frontal |
| Type | ST | 1st Surgery | 2nd Surgery | 3rd Surgery | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Path | FN * | VIM * | TGF-β | Path | FN * | VIM * | TGF-β | Path | FN * | VIM * | TGF-β | ||
| I | 36 | LGA | 1 | 2 | 2 | GBM | 2 | 3 | 3 | ||||
| II | 20 | GBM | 1 | 1 | 1 | GBM | 2 | 2 | 2 | ||||
| III | 41 | GBM | 1 | 0 | 1 | GBM | 2 | 1 | 2 | GBM | 3 | 2 | 3 |
| IV | 15 | GBM | 2 | 2 | 1 | GBM | 3 | 2 | 3 | ||||
| V | alive | GBM | 2 | 1 | 2 | GBM | 3 | 3 | 3 | LGA | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.-W.; Yang, C.-H.; Lin, Y.-H.; Hou, Y.-C.; Cheng, T.-J.; Chang, S.-T.; Huang, Y.-H.; Chung, S.-T.; Chio, C.-C.; Shan, Y.-S.; et al. The Fibronectin Expression Determines the Distinct Progressions of Malignant Gliomas via Transforming Growth Factor-Beta Pathway. Int. J. Mol. Sci. 2021, 22, 3782. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073782
Chen C-W, Yang C-H, Lin Y-H, Hou Y-C, Cheng T-J, Chang S-T, Huang Y-H, Chung S-T, Chio C-C, Shan Y-S, et al. The Fibronectin Expression Determines the Distinct Progressions of Malignant Gliomas via Transforming Growth Factor-Beta Pathway. International Journal of Molecular Sciences. 2021; 22(7):3782. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073782
Chicago/Turabian StyleChen, Chih-Wei, Cheng-Han Yang, Yuan-Ho Lin, Ya-Chin Hou, Tain-Junn Cheng, Sheng-Tsung Chang, Yu-Hua Huang, Shang-Ting Chung, Chung-Ching Chio, Yan-Shen Shan, and et al. 2021. "The Fibronectin Expression Determines the Distinct Progressions of Malignant Gliomas via Transforming Growth Factor-Beta Pathway" International Journal of Molecular Sciences 22, no. 7: 3782. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073782