
Altered microRNA Transcriptome in Cultured Human Liver Cells upon Infection with Ebola Virus

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
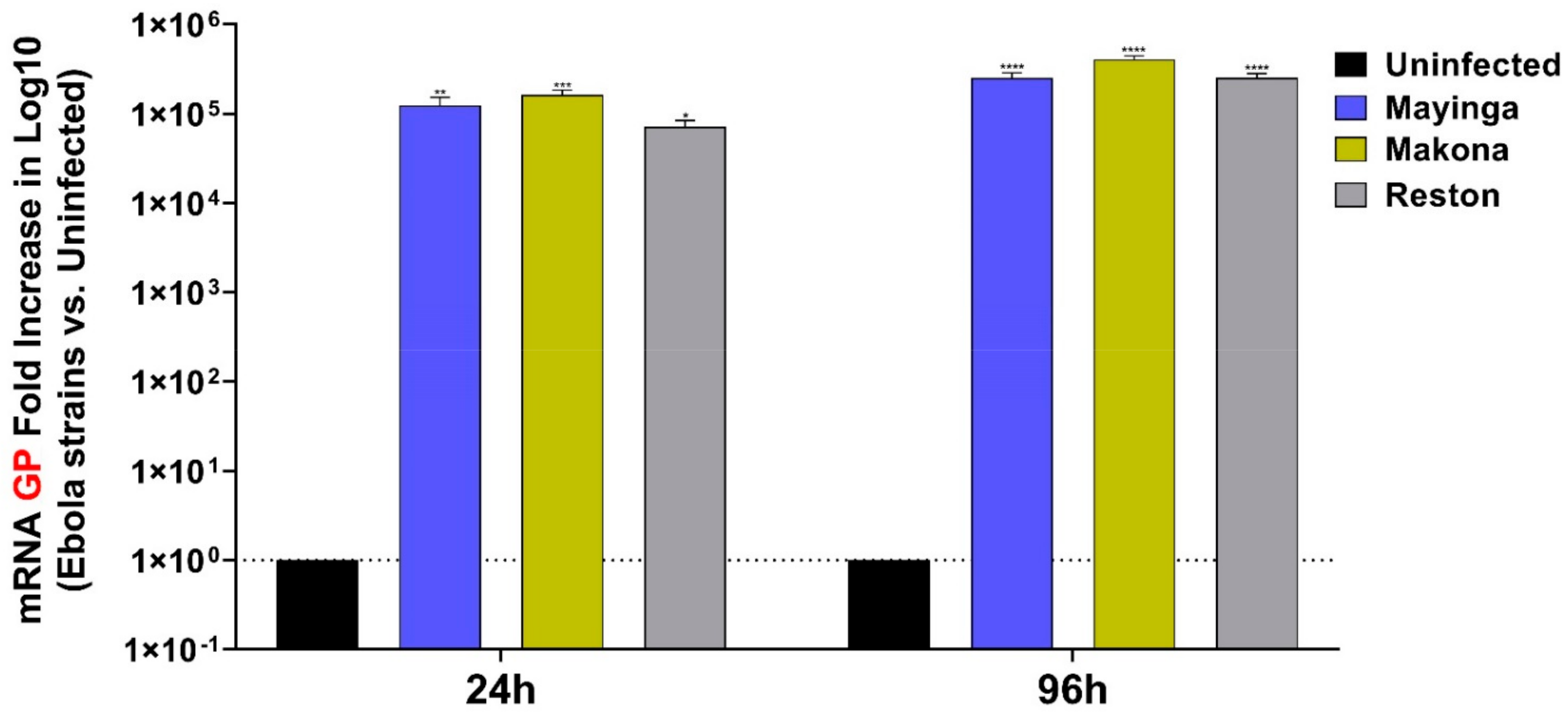
2.1. EBOV Relative Viral Replication
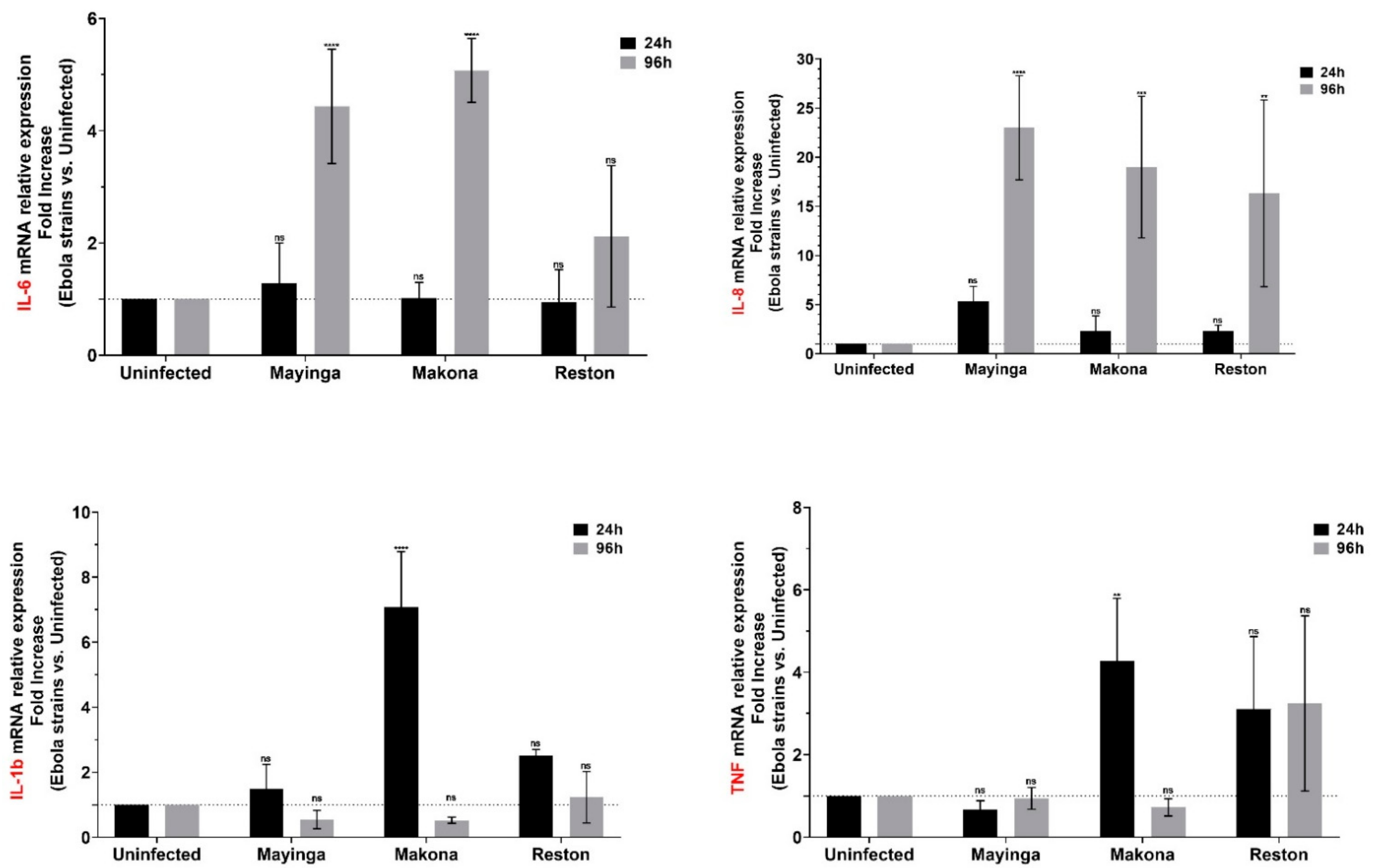
2.2. Makona May Elicit Proinflammatory Reaction Earlier Than Mayinga
2.3. Mayinga and Makona Haemorrhagic Phenotype May Result from an Imbalance of Coagulation Factors
2.4. Mayinga and Makona Do Not Seem to Promote Apoptosis in Cultured Hepatocytes
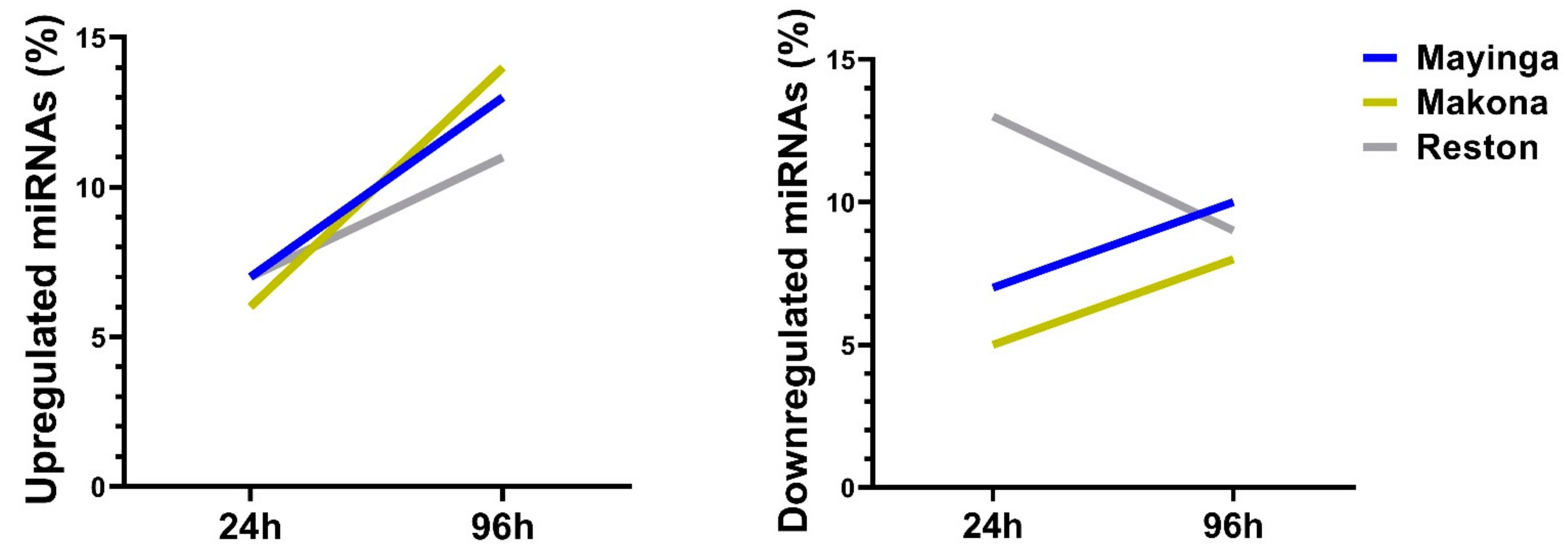
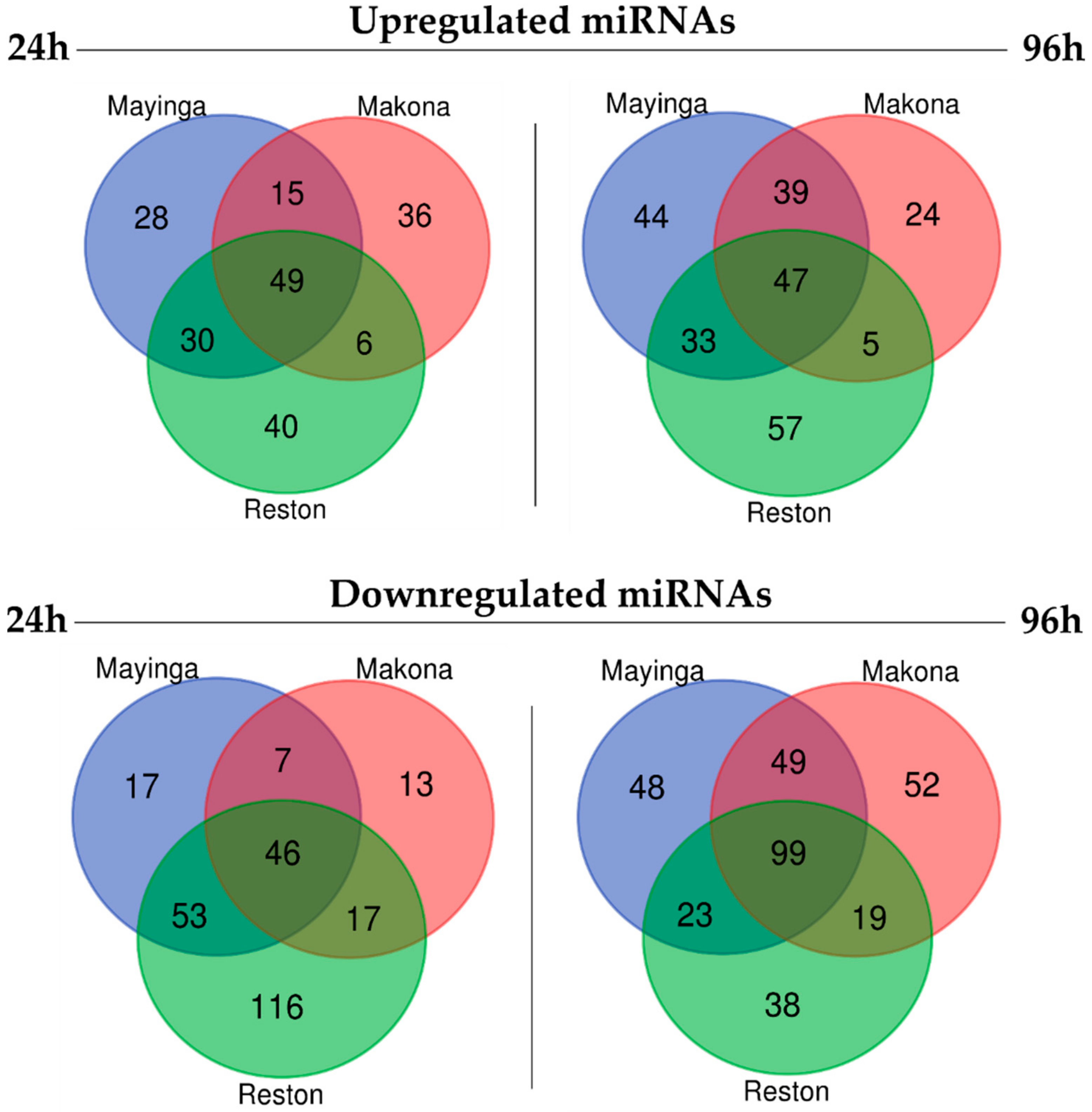
2.5. One Fifth of the miRNome Is Differentially Expressed over Time upon EBOV Infection
2.6. The Transient Downregulation of Certain Host miRNAs May Be an Achilles Heel for the RESTV
2.7. GO and KEGG Analyses of Differentially Expressed MiRNAs
2.7.1. Upregulated miRNAs
Early Stage
Later Stage
2.7.2. Downregulated miRNA
Early Stage
Later Stage
2.8. miR-122-5p, miR-148a-3p, miR-21-5p Are Selectively Modulated by EBOV
2.9. Filovirus-Targeting MiRNA
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cell Culture
5.2. Viruses
5.3. RNA Isolation
5.4. RT-qPCR
5.5. Statistical Analysis of qPCR Data
5.6. Illumina Nextseq Sequencing
5.6.1. Trimmed Reads
5.6.2. miRNA Expression Profiles
5.6.3. Differentially Expressed miRNA
5.6.4. Novel miRNAs
5.6.5. GO and KEGG Annotation/Enrichment
5.7. RNA22
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EGFR | Epidermal Growth Factor Receptor |
| GO | Gene Ontology |
| HCV | Hepatitis C Virus |
| ICAM-1 | Intercellular Adhesion Molecule-1 |
| IL | Interleukin |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| mTOR | mechanistic Target of Rapamycin |
| MOI | Multiplicity of infection |
| NF-kB | Nuclear factor kappa-light-chain-enhancer of activated β cells |
| RT-qPCR | Reverse Transcription-quantitative Polymerase Chain Reaction |
| TNF | Tumor Necrosis Factor |
| VCAM-1 | Vascular Cell Adhesion Molecule-1 |
References
- CDC. 2014–2016 Ebola Outbreak in West Africa|History|Ebola (Ebola Virus Disease)|CDC. Available online: https://www.cdc.gov/vhf/ebola/history/2014-2016-outbreak/index.html (accessed on 6 December 2020).
- Coltart, C.E.M.; Lindsey, B.; Ghinai, I.; Johnson, A.M.; Heymann, D.L. The Ebola Outbreak, 2013–2016: Old Lessons for New Epidemics. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Bào, Y.; Bavari, S.; Becker, S.; Bradfute, S.; Brauburger, K.; Brister, J.R.; Bukreyev, A.A.; Caì, Y.; Chandran, K.; et al. Virus Nomenclature below the Species Level: A Standardized Nomenclature for Filovirus Strains and Variants Rescued from CDNA. Arch. Virol. 2014, 159, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Cantoni, D.; Hamlet, A.; Michaelis, M.; Wass, M.N.; Rossman, J.S. Risks Posed by Reston, the Forgotten Ebolavirus. mSphere 2016, 1. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, T.; Anthony, S.J.; Gbakima, A.; Bird, B.H.; Bangura, J.; Tremeau-Bravard, A.; Belaganahalli, M.N.; Wells, H.; Dhanota, J.K.; Liang, E.; et al. Discovery of a New Ebolavirus (Bombali Virus) in Molossid Bats in Sierra Leone. Nat. Microbiol. 2018, 3, 1084–1089. [Google Scholar] [CrossRef]
- Rojas, M.; Monsalve, D.M.; Pacheco, Y.; Acosta-Ampudia, Y.; Ramírez-Santana, C.; Ansari, A.A.; Gershwin, M.E.; Anaya, J.-M. Ebola Virus Disease: An Emerging and Re-Emerging Viral Threat. J. Autoimmun. 2020, 106, 102375. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.; Messaoudi, I. Molecular Mechanisms of Ebola Pathogenesis. J. Leukoc. Biol. 2016, 100, 889–904. [Google Scholar] [CrossRef]
- Aleksandrowicz, P.; Marzi, A.; Biedenkopf, N.; Beimforde, N.; Becker, S.; Hoenen, T.; Feldmann, H.; Schnittler, H.-J. Ebola Virus Enters Host Cells by Macropinocytosis and Clathrin-Mediated Endocytosis. J. Infect. Dis. 2011, 204, S957–S967. [Google Scholar] [CrossRef] [Green Version]
- Davey, R.A.; Shtanko, O.; Anantpadma, M.; Sakurai, Y.; Chandran, K.; Maury, W. Mechanisms of Filovirus Entry. Curr. Top. Microbiol. Immunol. 2017, 411, 323–352. [Google Scholar] [CrossRef] [PubMed]
- Geisbert, T.W.; Hensley, L.E.; Larsen, T.; Young, H.A.; Reed, D.S.; Geisbert, J.B.; Scott, D.P.; Kagan, E.; Jahrling, P.B.; Davis, K.J. Pathogenesis of Ebola Hemorrhagic Fever in Cynomolgus Macaques: Evidence That Dendritic Cells Are Early and Sustained Targets of Infection. Am. J. Pathol. 2003, 163, 2347–2370. [Google Scholar] [CrossRef]
- Marcinkiewicz, J.; Bryniarski, K.; Nazimek, K. Ebola Haemorrhagic Fever Virus: Pathogenesis, Immune Responses, Potential Prevention. Folia Med. Cracov. 2014, 54, 39–48. [Google Scholar]
- Wong, G.; Kobinger, G.P.; Qiu, X. Characterization of Host Immune Responses in Ebola Virus Infections. Expert Rev. Clin. Immunol. 2014, 10, 781–790. [Google Scholar] [CrossRef]
- Basler, C.F. Innate Immune Evasion by Filoviruses. Virology 2015, 479–480, 122–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baseler, L.; Chertow, D.S.; Johnson, K.M.; Feldmann, H.; Morens, D.M. The Pathogenesis of Ebola Virus Disease. Annu. Rev. Pathol. Mech. Dis. 2017, 12, 387–418. [Google Scholar] [CrossRef] [PubMed]
- Reid, S.P.; Valmas, C.; Martinez, O.; Sanchez, F.M.; Basler, C.F. Ebola Virus VP24 Proteins Inhibit the Interaction of NPI-1 Subfamily Karyopherin Alpha Proteins with Activated STAT1. J. Virol. 2007, 81, 13469–13477. [Google Scholar] [CrossRef] [Green Version]
- Leung, D.W.; Ginder, N.D.; Fulton, D.B.; Nix, J.; Basler, C.F.; Honzatko, R.B.; Amarasinghe, G.K. Structure of the Ebola VP35 Interferon Inhibitory Domain. Proc. Natl. Acad. Sci. USA 2009, 106, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Olejnik, J.; Hume, A.J.; Leung, D.W.; Amarasinghe, G.K.; Basler, C.F.; Mühlberger, E. Filovirus Strategies to Escape Antiviral Responses. Curr. Top. Microbiol. Immunol. 2017, 411, 293–322. [Google Scholar] [CrossRef]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, J.; Rao, D.S. MicroRNAs in Inflammation and Immune Responses. Leukemia 2012, 26, 404–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, A.; Baltimore, D. MicroRNAs as Regulatory Elements in Immune System Logic. Nat. Rev. Immunol. 2016, 16, 279–294. [Google Scholar] [CrossRef]
- Wang, Z.; Li, J.; Fu, Y.; Zhao, Z.; Zhang, C.; Li, N.; Li, J.; Cheng, H.; Jin, X.; Lu, B.; et al. A Rapid Screen for Host-Encoded MiRNAs with Inhibitory Effects against Ebola Virus Using a Transcription- and Replication-Competent Virus-Like Particle System. Int. J. Mol. Sci. 2018, 19, 1488. [Google Scholar] [CrossRef] [Green Version]
- Sheng, M.; Zhong, Y.; Chen, Y.; Du, J.; Ju, X.; Zhao, C.; Zhang, G.; Zhang, L.; Liu, K.; Yang, N.; et al. Hsa-MiR-1246, Hsa-MiR-320a and Hsa-MiR-196b-5p Inhibitors Can Reduce the Cytotoxicity of Ebola Virus Glycoprotein in vitro. Sci. China Life Sci. 2014, 57, 959–972. [Google Scholar] [CrossRef]
- Duy, J.; Koehler, J.W.; Honko, A.N.; Schoepp, R.J.; Wauquier, N.; Gonzalez, J.-P.; Pitt, M.L.; Mucker, E.M.; Johnson, J.C.; O’Hearn, A.; et al. Circulating MicroRNA Profiles of Ebola Virus Infection. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Oliver, G.F.; Orang, A.V.; Appukuttan, B.; Marri, S.; Michael, M.Z.; Marsh, G.A.; Smith, J.R. Expression of MicroRNA in Human Retinal Pigment Epithelial Cells Following Infection with Zaire Ebolavirus. BMC Res. Notes 2019, 12, 639. [Google Scholar] [CrossRef] [PubMed]
- Golkar, Z.; Battaria, R.; Pace, D.G.; Bagasra, O. Inhibition of Ebola Virus by Anti-Ebola MiRNAs in Silico. J. Infect. Dev. Ctries. 2016, 10, 626–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grundhoff, A.; Sullivan, C.S. Virus-Encoded MicroRNAs. Virology 2011, 411, 325–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duy, J.; Honko, A.N.; Altamura, L.A.; Bixler, S.L.; Wollen-Roberts, S.; Wauquier, N.; O’Hearn, A.; Mucker, E.M.; Johnson, J.C.; Shamblin, J.D.; et al. Virus-Encoded MiRNAs in Ebola Virus Disease. Sci. Rep. 2018, 8, 6480. [Google Scholar] [CrossRef]
- Teng, Y.; Wang, Y.; Zhang, X.; Liu, W.; Fan, H.; Yao, H.; Lin, B.; Zhu, P.; Yuan, W.; Tong, Y.; et al. Systematic Genome-Wide Screening and Prediction of MicroRNAs in EBOV During the 2014 Ebolavirus Outbreak. Sci. Rep. 2015, 5, 9912. [Google Scholar] [CrossRef] [Green Version]
- Bosworth, A.; Dowall, S.D.; Armstrong, S.; Liu, X.; Dong, X.; Bruce, C.B.; Ng, L.F.P.; Carroll, M.W.; Hewson, R.; Hiscox, J.A. Investigating the Cellular Transcriptomic Response Induced by the Makona Variant of Ebola Virus in Differentiated THP-1 Cells. Viruses 2019, 11, 1023. [Google Scholar] [CrossRef] [Green Version]
- Marzi, A.; Feldmann, F.; Hanley, P.W.; Scott, D.; Günther, S.; Feldmann, H. Delayed Disease Progression in Cynomolgus Macaques Infected with Ebola Virus Makona Strain. Emerg. Infect. Dis. J. CDC 2015, 21. [Google Scholar] [CrossRef] [Green Version]
- Bosworth, A.; Dowall, S.D.; Garcia-Dorival, I.; Rickett, N.Y.; Bruce, C.B.; Matthews, D.A.; Fang, Y.; Aljabr, W.; Kenny, J.; Nelson, C.; et al. A Comparison of Host Gene Expression Signatures Associated with Infection in Vitro by the Makona and Ecran (Mayinga) Variants of Ebola Virus. Sci. Rep. 2017, 7, 43144. [Google Scholar] [CrossRef] [Green Version]
- Kash, J.C.; Mühlberger, E.; Carter, V.; Grosch, M.; Perwitasari, O.; Proll, S.C.; Thomas, M.J.; Weber, F.; Klenk, H.-D.; Katze, M.G. Global Suppression of the Host Antiviral Response by Ebola- and Marburgviruses: Increased Antagonism of the Type I Interferon Response is Associated with Enhanced Virulence. J. Virol. 2006, 80, 3009–3020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, S.; Rao, B.J.; Asgeirsson, B.; Dandekar, A.M. Correlating the Ability of VP24 Protein from Ebola and Marburg Viruses to Bind Human Karyopherin to Their Immune Suppression Mechanism and Pathogenicity Using Computational Methods. F1000Research 2014, 3, 265. [Google Scholar] [CrossRef] [Green Version]
- Jun, S.-R.; Leuze, M.R.; Nookaew, I.; Uberbacher, E.C.; Land, M.; Zhang, Q.; Wanchai, V.; Chai, J.; Nielsen, M.; Trolle, T.; et al. Ebolavirus Comparative Genomics. Fems. Microbiol. Rev. 2015, 39, 764–778. [Google Scholar] [CrossRef] [Green Version]
- Baize, S.; Pannetier, D.; Oestereich, L.; Rieger, T.; Koivogui, L.; Magassouba, N.; Soropogui, B.; Sow, M.S.; Keïta, S.; de Clerck, H.; et al. Emergence of Zaire Ebola Virus Disease in Guinea. N. Engl. J. Med. 2014, 371, 1418–1425. [Google Scholar] [CrossRef] [Green Version]
- Olejnik, J.; Forero, A.; Deflubé, L.R.; Hume, A.J.; Manhart, W.A.; Nishida, A.; Marzi, A.; Katze, M.G.; Ebihara, H.; Rasmussen, A.L.; et al. Ebolaviruses Associated with Differential Pathogenicity Induce Distinct Host Responses in Human Macrophages. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Ellis, D.S.; Simpson, I.H.; Francis, D.P.; Knobloch, J.; Bowen, E.T.; Lolik, P.; Deng, I.M. Ultrastructure of Ebola Virus Particles in Human Liver. J. Clin. Pathol. 1978, 31, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Speranza, E.; Muñoz-Fontela, C.; Haldenby, S.; Rickett, N.Y.; Garcia-Dorival, I.; Fang, Y.; Hall, Y.; Zekeng, E.-G.; Lüdtke, A.; et al. Transcriptomic Signatures Differentiate Survival from Fatal Outcomes in Humans Infected with Ebola Virus. Genome Biol. 2017, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martines, R.B.; Ng, D.L.; Greer, P.W.; Rollin, P.E.; Zaki, S.R. Tissue and Cellular Tropism, Pathology and Pathogenesis of Ebola and Marburg Viruses. J. Pathol. 2015, 235, 153–174. [Google Scholar] [CrossRef]
- Logue, J.; Vargas Licona, W.; Cooper, T.K.; Reeder, B.; Byrum, R.; Qin, J.; Deiuliis Murphy, N.; Cong, Y.; Bonilla, A.; Sword, J.; et al. Ebola Virus Isolation Using Huh-7 Cells Has Methodological Advantages and Similar Sensitivity to Isolation Using Other Cell Types and Suckling BALB/c Laboratory Mice. Viruses 2019, 11, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankeel, A.; Menicucci, A.R.; Woolsey, C.; Fenton, K.A.; Mendoza, N.; Versteeg, K.; Cross, R.W.; Geisbert, T.W.; Messaoudi, I. Early Transcriptional Changes within Liver, Adrenal Gland, and Lymphoid Tissues Significantly Contribute to Ebola Virus Pathogenesis in Cynomolgus Macaques. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Hartman, A.L.; Ling, L.; Nichol, S.T.; Hibberd, M.L. Whole-Genome Expression Profiling Reveals That Inhibition of Host Innate Immune Response Pathways by Ebola Virus Can Be Reversed by a Single Amino Acid Change in the VP35 Protein. J. Virol. 2008, 82, 5348–5358. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, H.; Geisbert, T.W. Ebola Haemorrhagic Fever. Lancet 2011, 377, 849–862. [Google Scholar] [CrossRef] [Green Version]
- Geisbert, T.W.; Jahrling, P.B. Use of Immunoelectron Microscopy to Show Ebola Virus during the 1989 United States Epizootic. J. Clin. Pathol. 1990, 43, 813–816. [Google Scholar] [CrossRef] [Green Version]
- Cilloniz, C.; Ebihara, H.; Ni, C.; Neumann, G.; Korth, M.J.; Kelly, S.M.; Kawaoka, Y.; Feldmann, H.; Katze, M.G. Functional Genomics Reveals the Induction of Inflammatory Response and Metalloproteinase Gene Expression during Lethal Ebola Virus Infection. J. Virol. 2011, 85, 9060–9068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisbert, T.W.; Young, H.A.; Jahrling, P.B.; Davis, K.J.; Kagan, E.; Hensley, L.E. Mechanisms Underlying Coagulation Abnormalities in Ebola Hemorrhagic Fever: Overexpression of Tissue Factor in Primate Monocytes/Macrophages Is a Key Event. J. Infect. Dis. 2003, 188, 1618–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, S.; Braspenning, J. Measurement of Blood Coagulation Factor Synthesis in Cultures of Human Hepatocytes. Methods Mol. Biol. 2015, 1250, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Gene Ontology Consortium. The Gene Ontology Resource: 20 Years and Still Going Strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Zhou, Q.; Lui, V.W.Y.; Yeo, W. Targeting the PI3K/Akt/MTOR Pathway in Hepatocellular Carcinoma. Future Oncol. 2011, 7, 1149–1167. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Chen, Z.; Shi, W.; Lian, Q. Tropomodulin 3 Promotes Liver Cancer Progression by Activating the MAPK/ERK Signaling Pathway. Oncol. Rep. 2019, 41, 3060–3068. [Google Scholar] [CrossRef] [PubMed]
- Sequera, C.; Manzano, S.; Guerrero, C.; Porras, A. How Rap and Its GEFs Control Liver Physiology and Cancer Development. C3G Alterations in Human Hepatocarcinoma. Hepat. Oncol. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López, P.; Girardi, E.; Pfeffer, S. Importance of cellular microRNAs in the regulation of viral infections. Med. Sci. 2019, 35, 667–673. [Google Scholar] [CrossRef]
- Girardi, E.; López, P.; Pfeffer, S. On the Importance of Host MicroRNAs During Viral Infection. Front. Genet. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Bruscella, P.; Bottini, S.; Baudesson, C.; Pawlotsky, J.-M.; Feray, C.; Trabucchi, M. Viruses and MiRNAs: More Friends than Foes. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Schult, P.; Roth, H.; Adams, R.L.; Mas, C.; Imbert, L.; Orlik, C.; Ruggieri, A.; Pyle, A.M.; Lohmann, V. MicroRNA-122 Amplifies Hepatitis C Virus Translation by Shaping the Structure of the Internal Ribosomal Entry Site. Nat. Commun. 2018, 9, 2613. [Google Scholar] [CrossRef]
- Pereira, A.; Moreira, F.; Vinasco-Sandoval, T.; Cunha, A.; Vidal, A.; Ribeiro-dos-Santos, A.M.; Pinto, P.; Magalhães, L.; Assumpção, M.; Demachki, S.; et al. MiRNome Reveals New Insights Into the Molecular Biology of Field Cancerization in Gastric Cancer. Front. Genet. 2019, 10. [Google Scholar] [CrossRef]
- Kendziorski, C.M.; Zhang, Y.; Lan, H.; Attie, A.D. The Efficiency of Pooling MRNA in Microarray Experiments. Biostatistics 2003, 4, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Kendziorski, C.; Irizarry, R.A.; Chen, K.-S.; Haag, J.D.; Gould, M.N. On the Utility of Pooling Biological Samples in Microarray Experiments. Proc. Natl. Acad. Sci. USA 2005, 102, 4252–4257. [Google Scholar] [CrossRef] [Green Version]
- Glass, Ä.; Henning, J.; Karopka, T.; Scheel, T.; Bansemer, S.; Koczan, D.; Gierl, L.; Rolfs, A.; Gimsa, U. Representation of Individual Gene Expression in Completely Pooled MRNA Samples. Biosci. Biotechnol. Biochem. 2005, 69, 1098–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, R.D.; O’Connor, M.D.; Griffith, M.; Kuchenbauer, F.; Delaney, A.; Prabhu, A.-L.; Zhao, Y.; McDonald, H.; Zeng, T.; Hirst, M.; et al. Application of Massively Parallel Sequencing to MicroRNA Profiling and Discovery in Human Embryonic Stem Cells. Genome Res. 2008, 18, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Ziemann, M.; Kaspi, A.; El-Osta, A. Evaluation of MicroRNA Alignment Techniques. RNA 2016, 22, 1120–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, M.; Benmoussa, A.; Provost, P. Small Non-Coding RNAs Derived From Eukaryotic Ribosomal RNA. Noncoding RNA 2019, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diallo, I.; Provost, P. RNA-Sequencing Analyses of Small Bacterial RNAs and Their Emergence as Virulence Factors in Host-Pathogen Interactions. Int. J. Mol. Sci. 2020, 21, 1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thankaswamy-Kosalai, S.; Sen, P.; Nookaew, I. Evaluation and Assessment of Read-Mapping by Multiple next-Generation Sequencing Aligners Based on Genome-Wide Characteristics. Genomics 2017, 109, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Lindner, R.; Friedel, C.C. A Comprehensive Evaluation of Alignment Algorithms in the Context of RNA-Seq. PLoS ONE 2012, 7, e52403. [Google Scholar] [CrossRef] [Green Version]
- Leshkowitz, D.; Horn-Saban, S.; Parmet, Y.; Feldmesser, E. Differences in MicroRNA Detection Levels Are Technology and Sequence Dependent. RNA 2013, 19, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Wauquier, N.; Becquart, P.; Padilla, C.; Baize, S.; Leroy, E.M. Human Fatal Zaire Ebola Virus Infection Is Associated with an Aberrant Innate Immunity and with Massive Lymphocyte Apoptosis. PLoS Negl. Trop. Dis. 2010, 4. [Google Scholar] [CrossRef]
- Andus, T.; Bauer, J.; Gerok, W. Effects of Cytokines on the Liver. Hepatology 1991, 13, 364–375. [Google Scholar] [CrossRef]
- Schmidt-Arras, D.; Rose-John, S. IL-6 Pathway in the Liver: From Physiopathology to Therapy. J. Hepatol. 2016, 64, 1403–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.-M.; An, J. Cytokines, Inflammation and Pain. Int. Anesth. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Schoenmakers, S.H.H.F.; Reitsma, P.H.; Spek, C.A. Blood Coagulation Factors as Inflammatory Mediators. Blood Cells Mol. Dis. 2005, 34, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Neumann, F.J.; Ott, I.; Marx, N.; Luther, T.; Kenngott, S.; Gawaz, M.; Kotzsch, M.; Schömig, A. Effect of Human Recombinant Interleukin-6 and Interleukin-8 on Monocyte Procoagulant Activity. Arter. Thromb. Vasc. Biol. 1997, 17, 3399–3405. [Google Scholar] [CrossRef] [PubMed]
- Grignani, G.; Maiolo, A. Cytokines and Hemostasis. Haematologica 2000, 85, 967–972. [Google Scholar] [PubMed]
- Smith, S.A.; Travers, R.J.; Morrissey, J.H. How It All Starts: Initiation of the Clotting Cascade. Crit. Rev. Biochem. Mol. Biol. 2015, 50, 326–336. [Google Scholar] [CrossRef] [Green Version]
- Smither, S.J.; O’Brien, L.M.; Eastaugh, L.; Woolley, T.; Lever, M.; Fletcher, T.; Parmar, K.; Hunt, B.J.; Watts, S.; Kirkman, E. Haemostatic Changes in Five Patients Infected with Ebola Virus. Viruses 2019, 11, 647. [Google Scholar] [CrossRef] [Green Version]
- Bhella, D. The Role of Cellular Adhesion Molecules in Virus Attachment and Entry. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140035. [Google Scholar] [CrossRef] [Green Version]
- Kondratowicz, A.S.; Lennemann, N.J.; Sinn, P.L.; Davey, R.A.; Hunt, C.L.; Moller-Tank, S.; Meyerholz, D.K.; Rennert, P.; Mullins, R.F.; Brindley, M.; et al. T-Cell Immunoglobulin and Mucin Domain 1 (TIM-1) Is a Receptor for Zaire Ebolavirus and Lake Victoria Marburgvirus. Proc. Natl. Acad. Sci. USA 2011, 108, 8426–8431. [Google Scholar] [CrossRef] [Green Version]
- Bradfute, S.B.; Swanson, P.E.; Smith, M.A.; Watanabe, E.; McDunn, J.E.; Hotchkiss, R.S.; Bavari, S. Mechanisms and Consequences of Ebolavirus-Induced Lymphocyte Apoptosis. J. Immunol. 2010, 184, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Zampieri, C.A.; Sullivan, N.J.; Nabel, G.J. Immunopathology of Highly Virulent Pathogens: Insights from Ebola Virus. Nat. Immunol. 2007, 8, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Boehmann, Y.; Enterlein, S.; Randolf, A.; Mühlberger, E. A Reconstituted Replication and Transcription System for Ebola Virus Reston and Comparison with Ebola Virus Zaire. Virology 2005, 332, 406–417. [Google Scholar] [CrossRef] [Green Version]
- Jopling, C.L.; Yi, M.; Lancaster, A.M.; Lemon, S.M.; Sarnow, P. Modulation of Hepatitis C Virus RNA Abundance by a Liver-Specific MicroRNA. Science 2005, 309, 1577–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedano, C.D.; Sarnow, P. Hepatitis C Virus Subverts Liver-Specific MiR-122 to Protect the Viral Genome from Exoribonuclease Xrn2. Cell Host Microbe 2014, 16, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Masaki, T.; Arend, K.C.; Li, Y.; Yamane, D.; McGivern, D.R.; Kato, T.; Wakita, T.; Moorman, N.J.; Lemon, S.M. MiR-122 Stimulates Hepatitis C Virus RNA Synthesis by Altering the Balance of Viral RNAs Engaged in Replication versus Translation. Cell Host Microbe 2015, 17, 217–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Wang, J.; Huang, M.; Xu, G.; Wei, W.; Qin, H. Inhibition of MiR-148a-3p Resists Hepatocellular Carcinoma Progress of Hepatitis C Virus Infection through Suppressing C-Jun and MAPK Pathway. J. Cell Mol. Med. 2019, 23, 1415–1426. [Google Scholar] [CrossRef] [Green Version]
- Clément, S.; Sobolewski, C.; Gomes, D.; Rojas, A.; Goossens, N.; Conzelmann, S.; Calo, N.; Negro, F.; Foti, M. Activation of the Oncogenic MiR-21-5p Promotes HCV Replication and Steatosis Induced by the Viral Core 3a Protein. Liver Int. 2019, 39, 1226–1236. [Google Scholar] [CrossRef]
- Hoofnagle, J.H. Course and Outcome of Hepatitis C. Hepatology 2002, 36, S21–S29. [Google Scholar] [CrossRef]
- Shi, G.; Suzuki, T. Molecular Basis of Encapsidation of Hepatitis C Virus Genome. Front. Microbiol. 2018, 9, 396. [Google Scholar] [CrossRef] [Green Version]
- Shabman, R.S.; Hoenen, T.; Groseth, A.; Jabado, O.; Binning, J.M.; Amarasinghe, G.K.; Feldmann, H.; Basler, C.F. An Upstream Open Reading Frame Modulates Ebola Virus Polymerase Translation and Virus Replication. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef]
- Beniac, D.R.; Booth, T.F. Structure of the Ebola Virus Glycoprotein Spike within the Virion Envelope at 11 Å Resolution. Sci. Rep. 2017, 7, 46374. [Google Scholar] [CrossRef] [Green Version]
- Niepmann, M.; Gerresheim, G.K. Hepatitis C Virus Translation Regulation. Int. J. Mol. Sci. 2020, 21, 2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choo, Q.L.; Kuo, G.; Weiner, A.J.; Overby, L.R.; Bradley, D.W.; Houghton, M. Isolation of a CDNA Clone Derived from a Blood-Borne Non-A, Non-B Viral Hepatitis Genome. Science 1989, 244, 359–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burlone, M.E.; Budkowska, A. Hepatitis C Virus Cell Entry: Role of Lipoproteins and Cellular Receptors. J. Gen. Virol. 2009, 90, 1055–1070. [Google Scholar] [CrossRef] [PubMed]
- Lanford, R.E.; Hildebrandt-Eriksen, E.S.; Petri, A.; Persson, R.; Lindow, M.; Munk, M.E.; Kauppinen, S.; Ørum, H. Therapeutic Silencing of MicroRNA-122 in Primates with Chronic Hepatitis C Virus Infection. Science 2010, 327, 198–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Sun, D.; Li, W.; Shen, H.; Zhu, Y.; Li, C.; Chen, Y.; Lu, L.; Li, W.; Zhang, J.; et al. A C-Myc-MicroRNA Functional Feedback Loop Affects Hepatocarcinogenesis. Hepatology 2013, 57, 2378–2389. [Google Scholar] [CrossRef] [PubMed]
- Drobna, M.; Szarzyńska, B.; Jaksik, R.; Sędek, Ł.; Kuchmiy, A.; Taghon, T.; van Vlierberghe, P.; Szczepański, T.; Witt, M.; Dawidowska, M. Hsa-MiR-20b-5p and Hsa-MiR-363-3p Affect Expression of PTEN and BIM Tumor Suppressor Genes and Modulate Survival of T-ALL Cells In Vitro. Cells 2020, 9, 1137. [Google Scholar] [CrossRef]
- Ayoubian, H.; Ludwig, N.; Fehlmann, T.; Menegatti, J.; Gröger, L.; Anastasiadou, E.; Trivedi, P.; Keller, A.; Meese, E.; Grässer, F.A. Epstein-Barr Virus Infection of Cell Lines Derived from Diffuse Large B-Cell Lymphomas Alters MicroRNA Loading of the Ago2 Complex. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Lin, X.; Yang, X.; Lu, G.; Zhang, Q.; Zhang, C. MicroRNA-132-3p Suppresses Type I IFN Response through Targeting IRF1 to Facilitate H1N1 Influenza A Virus Infection. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Xu, L.; Liu, Y.; Li, C.; Zhang, L.; Wang, T.; Zhao, D.; Xu, X.; Zhang, Y. MicroRNA-221-5p Inhibits Porcine Epidemic Diarrhea Virus Replication by Targeting Genomic Viral RNA and Activating the NF-ΚB Pathway. Int. J. Mol. Sci. 2018, 19, 3381. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Yao, Y.; Song, Q.; Li, S.; Hu, Z.; Yu, Y.; Hu, C.; Da, X.; Li, H.; Chen, Q.; et al. Up-Regulation of MiR-95-3p in Hepatocellular Carcinoma Promotes Tumorigenesis by Targeting P21 Expression. Sci. Rep. 2016, 6, 34034. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Gao, J.; Liang, Y.; Li, R.; Xiao, Q.; Zhang, Z.; Fan, X. Integration Analysis of a MiRNA-MRNA Expression in A549 Cells Infected with a Novel H3N2 Swine Influenza Virus and the 2009 H1N1 Pandemic Influenza Virus. Infect. Genet. Evol. 2019, 74, 103922. [Google Scholar] [CrossRef]
- Song, L.; Liu, H.; Gao, S.; Jiang, W.; Huang, W. Cellular MicroRNAs Inhibit Replication of the H1N1 Influenza A Virus in Infected Cells. J. Virol. 2010, 84, 8849–8860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodge, R.; Ferreira Barbosa, J.A.; Lombard-Vadnais, F.; Gilmore, J.C.; Deshiere, A.; Gosselin, A.; Wiche Salinas, T.R.; Bego, M.G.; Power, C.; Routy, J.-P.; et al. Host MicroRNAs-221 and -222 Inhibit HIV-1 Entry in Macrophages by Targeting the CD4 Viral Receptor. Cell Rep. 2017, 21, 141–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Zhang, Q.; Gao, L.; Li, N.; Chen, X.; Feng, W. Increasing Expression of MicroRNA 181 Inhibits Porcine Reproductive and Respiratory Syndrome Virus Replication and Has Implications for Controlling Virus Infection. J. Virol. 2013, 87, 1159–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santhakumar, D.; Forster, T.; Laqtom, N.N.; Fragkoudis, R.; Dickinson, P.; Abreu-Goodger, C.; Manakov, S.A.; Choudhury, N.R.; Griffiths, S.J.; Vermeulen, A.; et al. Combined Agonist–Antagonist Genome-Wide Functional Screening Identifies Broadly Active Antiviral MicroRNAs. Proc. Natl. Acad. Sci. USA 2010, 107, 13830–13835. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Wang, F.; Argyris, E.; Chen, K.; Liang, Z.; Tian, H.; Huang, W.; Squires, K.; Verlinghieri, G.; Zhang, H. Cellular MicroRNAs Contribute to HIV-1 Latency in Resting Primary CD4 + T Lymphocytes. Nat. Med. 2007, 13, 1241–1247. [Google Scholar] [CrossRef]
- Li, Y.; Jiao, Y.; Fu, Z.; Luo, Z.; Su, J.; Li, Y. High MiR-454-3p Expression Predicts Poor Prognosis in Hepatocellular Carcinoma. Cancer Manag. Res. 2019, 11, 2795–2802. [Google Scholar] [CrossRef] [Green Version]
- Versteeg, K.; Menicucci, A.R.; Woolsey, C.; Mire, C.E.; Geisbert, J.B.; Cross, R.W.; Agans, K.N.; Jeske, D.; Messaoudi, I.; Geisbert, T.W. Infection with the Makona Variant Results in a Delayed and Distinct Host Immune Response Compared to Previous Ebola Virus Variants. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-C.; Yu, H.-R.; Huang, L.-T.; Huang, H.-C.; Chen, R.-F.; Lin, I.-C.; Ou, C.-Y.; Hsu, T.-Y.; Yang, K.D. MiRNA-125b Regulates TNF-α Production in CD14+ Neonatal Monocytes via Post-Transcriptional Regulation. J. Leukoc. Biol. 2012, 92, 171–182. [Google Scholar] [CrossRef]
- Miranda, K.C.; Huynh, T.; Tay, Y.; Ang, Y.-S.; Tam, W.-L.; Thomson, A.M.; Lim, B.; Rigoutsos, I. A Pattern-Based Method for the Identification of MicroRNA Binding Sites and Their Corresponding Heteroduplexes. Cell 2006, 126, 1203–1217. [Google Scholar] [CrossRef] [Green Version]
- Menendez, D.; Shatz, M.; Resnick, M.A. Interactions between the Tumor Suppressor P53 and Immune Responses. Curr. Opin. Oncol. 2013, 25, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Hua, H.; Kong, Q.; Zhang, H.; Wang, J.; Luo, T.; Jiang, Y. Targeting MTOR for Cancer Therapy. J. Hematol. Oncol. 2019, 12, 71. [Google Scholar] [CrossRef] [PubMed]
- Cuenda, A.; Rousseau, S. P38 MAP-Kinases Pathway Regulation, Function and Role in Human Diseases. Biochim. Biophys. Acta 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, L.; Zhou, G.; Soufan, O.; Xia, J. MiRNet 2.0: Network-Based Visual Analytics for MiRNA Functional Analysis and Systems Biology. Nucleic Acids Res. 2020, 48, W244–W251. [Google Scholar] [CrossRef]
- Fischer, T.; Spohn, M.; Olearo, F.; Zinser, M.E.; Kasonta, R.; Stubbe, H.C.; Rechtien, A.; Ly, M.L.; Schmiedel, S.; Lohse, A.W.; et al. Dynamic Changes of Circulating MiRNAs Induced by the Ebola Virus Vaccine VSV-EBOV. Vaccine 2018, 36, 7083–7094. [Google Scholar] [CrossRef]
- Li, C.; Hu, X.; Li, L.; Li, J.-H. Differential MicroRNA Expression in the Peripheral Blood from Human Patients with COVID-19. J. Clin. Lab. Anal. 2020, 34, e23590. [Google Scholar] [CrossRef]
- Cucinotta, D.; Vanelli, M. WHO Declares COVID-19 a Pandemic. Acta Biomed. 2020, 91, 157–160. [Google Scholar] [CrossRef]
- Muller, J.E.; Nathan, D.G. COVID-19, Nuclear War, and Global Warming: Lessons for Our Vulnerable World. Lancet 2020, 395, 1967–1968. [Google Scholar] [CrossRef]
- Longo, P.A.; Kavran, J.M.; Kim, M.-S.; Leahy, D.J. Transient Mammalian Cell Transfection with Polyethylenimine (PEI). Methods Enzym. 2013, 529, 227–240. [Google Scholar] [CrossRef] [Green Version]
- Benmoussa, A.; Diallo, I.; Salem, M.; Michel, S.; Gilbert, C.; Sévigny, J.; Provost, P. Concentrates of Two Subsets of Extracellular Vesicles from Cow’s Milk Modulate Symptoms and Inflammation in Experimental Colitis. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A Tool to Design Target-Specific Primers for Polymerase Chain Reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, A.; Jung, M.; Miller, K.; Lein, M.; Kristiansen, G.; Erbersdobler, A.; Jung, K. Suitable Reference Genes for Relative Quantification of MiRNA Expression in Prostate Cancer. Exp. Mol. Med. 2010, 42, 749–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bullard, J.H.; Purdom, E.; Hansen, K.D.; Dudoit, S. Evaluation of Statistical Methods for Normalization and Differential Expression in MRNA-Seq Experiments. BMC Bioinform. 2010, 11, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedländer, M.R.; Chen, W.; Adamidi, C.; Maaskola, J.; Einspanier, R.; Knespel, S.; Rajewsky, N. Discovering MicroRNAs from Deep Sequencing Data Using MiRDeep. Nat. Biotechnol. 2008, 26, 407–415. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mayinga | Makona | Reston | |||
|---|---|---|---|---|---|
| Mature miRNA | FC vs. C | Mature miRNA | FC vs. C | Mature miRNA | FC vs. C |
| miR-363-3p | 5.18 | miR-454-3p | 3.36 | miR-374a-3p | 6.15 |
| miR-374a-5p | 4.57 | miR-374a-3p | 2.80 | miR-26a-2-3p | 4.14 |
| miR-374b-5p | 3.90 | miR-374b-5p | 2.37 | miR-374b-5p | 3.92 |
| miR-374a-3p | 3.80 | miR-132-3p | 2.36 | miR-454-3p | 3.90 |
| miR-454-3p | 3.78 | miR-4483 | 2.31 | miR-374a-5p | 3.79 |
| miR-32-5p | 3.64 | miR-novel-chr1_24913 | 2.23 | miR-3613-5p | 3.71 |
| miR-429 | 3.57 | miR-221-5p | 2.20 | miR-664a-3p | 3.50 |
| miR-20a-5p | 2.90 | miR-19a-3p | 2.16 | miR-3128 | 3.23 |
| miR-26a-2-3p | 2.86 | miR-664a-3p | 2.15 | miR-95-3p | 3.17 |
| miR-200a-3p | 2.81 | miR-429 | 2.14 | miR-126-5p | 3.00 |
| miR-126-5p | 2.50 | miR-652-5p | 2.14 | miR-652-5p | 3.00 |
| miR-216a-5p | 2.50 | miR-4521 | 2.14 | miR-582-5p | 2.87 |
| miR-3613-5p | 2.47 | miR-374a-5p | 2.13 | miR-429 | 2.79 |
| miR-novelchr16_13983 | 2.47 | miR-6894-3p | 2.00 | miR-32-5p | 2.77 |
| miR-16-5p | 2.41 | miR-novel-chr20_29712 | 2.00 | miR-582-3p | 2.69 |
| miR-16-5p | 2.39 | miR-novel-chr2_37626 | 2.00 | miR-450b-5p | 2.67 |
| miR-548-3p | 2.36 | miR-32-5p | 1.95 | miR-21-5p | 2.65 |
| miR-18a-5p | 2.32 | miR-20a-5p | 1.92 | miR-30e-5p | 2.62 |
| miR-novelchr4_42996 | 2.31 | miR-novel-chr10_1855 | 1.90 | miR-148a-3p | 2.55 |
| miR-19a-3p | 2.30 | miR-4455 | 1.90 | miR-548x-3p | 2.45 |
| Mayinga | Makona | Reston | |||
|---|---|---|---|---|---|
| Mature miRNA | FC vs. C | Mature miRNA | FC vs. C | Mature miRNA | FC vs. C |
| miR-27a-5p | 22.35 | miR-novel-chr1_24417 | 11.50 | miR-novel-chr1_23305 | 16.80 |
| miR-4443 | 21.58 | miR-novel-chr7_52141 | 6.83 | miR-193b-5p | 14.33 |
| miR-195-3p | 11.18 | miR-novel-chr4_43591 | 6.77 | miR-novel-chr19_22180 | 9.70 |
| miR-132-5p | 9.82 | miR-novel-chr1_26062 | 5.03 | miR-novel-chr3_39440 | 9.30 |
| miR-145-3p | 9.73 | miR-novel-chr14_11806 | 4.69 | miR-novel-chr19_21743 | 8.00 |
| miR-4485-3p | 9.55 | miR-novel-chr12_7186 | 4.67 | miR-novel-chr5_47105 | 7.56 |
| miR-novel-chr1_24417 | 9.19 | miR-novel-chr19_21089 | 4.64 | miR-1246 | 7.53 |
| miR-novel-chr4_43591 | 9.12 | miR-4443 | 4.50 | miR-novel-chr17_17480 | 6.45 |
| miR-novel-chr7_52141 | 8.87 | miR-novel-chr6_48321 | 4.46 | miR-novel-chr4_43591 | 6.23 |
| miR-181b-3p | 8.62 | miR-novel-chr22_33060 | 4.26 | miR-novel-chr7_52141 | 6.20 |
| miR-novel-chr19_21743 | 7.89 | miR-27a-5p | 4.23 | miR-27a-5p | 5.98 |
| miR-92a-1-5p | 6.95 | miR-92a-1-5p | 4.05 | miR-novel-chr12_8312 | 5.80 |
| miR-181a-3p | 6.91 | miR-novel-chr20_29712 | 4.01 | miR-novel-chr17_17167 | 5.77 |
| miR-novel-chr19_22180 | 6.65 | miR-195-3p | 4.00 | miR-4792 | 5.63 |
| miR-novel-chr17_17480 | 6.60 | miR-215-3p | 3.95 | miR-novel-chr12_6127 | 5.41 |
| miR-193b-5p | 6.39 | miR-novel-chr10_1111 | 3.94 | miR-26a-2-3p | 5.40 |
| miR-200a-5p | 5.88 | miR-novel-chr3_38884 | 3.81 | miR-4485-3p | 5.18 |
| miR-novel-chr12_7186 | 5.72 | miR-181b-3p | 3.67 | miR-1290 | 5.13 |
| miR-novel-chr1_26062 | 5.68 | miR-4497 | 3.64 | miR-146a-3p | 5.11 |
| miR-novel-chr19_21089 | 5.30 | miR-novel-chr1_24751 | 3.56 | miR-novel-chr20_30085 | 5.07 |
| Mayinga | Makona | Reston | |||
|---|---|---|---|---|---|
| Mature miRNA | FC vs. C | Mature miRNA | FC vs. C | Mature miRNA | FC vs. C |
| hsa-miR-novel-chr16_14823 | 0.19 | hsa-miR-novel-chr16_14823 | 0.03 | hsa-miR-novel-chr16_14823 | 0.04 |
| hsa-miR-451a | 0.27 | hsa-miR-novel-chr5_47105 | 0.24 | hsa-miR-novel-chr5_47105 | 0.15 |
| hsa-miR-novel-chr2_35636 | 0.30 | hsa-miR-451a | 0.25 | hsa-miR-novel-chr12_6591 | 0.17 |
| hsa-miR-210-5p | 0.31 | hsa-miR-novel-chr15_12117 | 0.32 | hsa-miR-3182 | 0.17 |
| hsa-miR-486-5p | 0.35 | hsa-miR-143-3p | 0.32 | hsa-miR-novel-chr19_20368 | 0.18 |
| hsa-miR-486-5p | 0.35 | hsa-miR-novel-chr2_36744 | 0.33 | hsa-miR-novel-chr2_36744 | 0.18 |
| hsa-miR-novel-chr5_47105 | 0.37 | hsa-miR-novel-chr9_56521 | 0.34 | hsa-miR-novel-chr2_34829 | 0.21 |
| hsa-miR-novel-chr2_36744 | 0.39 | hsa-miR-novel-chr8_54829 | 0.34 | hsa-miR-novel-chr4_41778 | 0.21 |
| hsa-miR-210-3p | 0.40 | hsa-miR-novel-chr2_34829 | 0.37 | hsa-miR-novel-chr16_14481 | 0.24 |
| hsa-miR-7974 | 0.40 | hsa-miR-3182 | 0.38 | hsa-miR-940 | 0.24 |
| hsa-miR-491-5p | 0.43 | hsa-miR-210-5p | 0.39 | hsa-miR-novel-chr3_38187 | 0.25 |
| hsa-miR-novel-chr17_18292 | 0.44 | hsa-miR-210-3p | 0.39 | hsa-miR-210-3p | 0.26 |
| hsa-miR-novel-chr15_12117 | 0.44 | hsa-miR-novel-chr2_35636 | 0.40 | hsa-miR-novel-chr2_35636 | 0.26 |
| hsa-miR-novel-chr4_41778 | 0.45 | hsa-miR-199a-5p | 0.40 | hsa-miR-novel-chr1_26376 | 0.26 |
| hsa-miR-novel-chr19_20494 | 0.45 | hsa-miR-199a-5p | 0.40 | hsa-miR-novel-chr17_16473 | 0.26 |
| hsa-miR-novel-chr9_56521 | 0.46 | hsa-miR-199b-5p | 0.40 | hsa-miR-1291 | 0.27 |
| hsa-miR-3182 | 0.47 | hsa-miR-486-5p | 0.42 | hsa-miR-novel-chr3_39344 | 0.27 |
| hsa-let-7i-5p | 0.47 | hsa-miR-novel-chr16_14481 | 0.42 | hsa-miR-novel-chr3_38154 | 0.27 |
| hsa-miR-224-5p | 0.48 | hsa-miR-novel-chr3_38187 | 0.42 | hsa-miR-671-5p | 0.28 |
| hsa-miR-1908-3p | 0.48 | hsa-miR-486-5p | 0.43 | hsa-miR-novel-chr17_16481 | 0.29 |
| Mayinga | Makona | Reston | |||
|---|---|---|---|---|---|
| Mature miRNA | FC vs. C | Mature miRNA | FC vs. C | Mature miRNA | FC vs. C |
| hsa-miR-671-5p | 0.08 | hsa-miR-7974 | 0.08 | hsa-miR-4454 | 0.18 |
| hsa-miR-7974 | 0.09 | hsa-miR-671-5p | 0.12 | hsa-miR-582-5p | 0.19 |
| hsa-miR-novel-chr15_13530 | 0.12 | hsa-miR-582-3p | 0.13 | hsa-miR-7974 | 0.20 |
| hsa-miR-582-5p | 0.15 | hsa-miR-582-5p | 0.15 | hsa-miR-novel-chr15_13530 | 0.21 |
| hsa-miR-326 | 0.16 | hsa-miR-novel-chr15_13530 | 0.17 | hsa-miR-novel-chr21_31339 | 0.27 |
| hsa-miR-3182 | 0.21 | hsa-miR-574-5p | 0.17 | hsa-miR-1306-3p | 0.28 |
| hsa-miR-novel-chr12_5743 | 0.21 | hsa-miR-326 | 0.18 | hsa-miR-1303 | 0.29 |
| hsa-miR-novel-chr18_19540 | 0.22 | hsa-miR-novel-chr4_41778 | 0.20 | hsa-miR-326 | 0.29 |
| hsa-miR-4454 | 0.22 | hsa-miR-novel-chr12_8466 | 0.20 | hsa-miR-582-3p | 0.30 |
| hsa-miR-1257 | 0.22 | hsa-miR-1291 | 0.23 | hsa-miR-5591-3p | 0.30 |
| hsa-miR-novel-chr16_14823 | 0.03 | hsa-miR-novel-chr17_16082 | 0.23 | hsa-miR-589-3p | 0.32 |
| hsa-miR-novel-chr5_47105 | 0.24 | hsa-miR-novel-chr21_31339 | 0.23 | hsa-miR-573 | 0.32 |
| hsa-miR-451a | 0.25 | hsa-miR-novel-chr18_19540 | 0.24 | hsa-miR-671-5p | 0.33 |
| hsa-miR-novel-chr15_12117 | 0.32 | hsa-miR-1257 | 0.26 | hsa-miR-342-3p | 0.36 |
| hsa-miR-143-3p | 0.32 | hsa-miR-3679-5p | 0.27 | hsa-miR-500a-5p | 0.36 |
| hsa-miR-novel-chr2_36744 | 0.33 | hsa-miR-3182 | 0.27 | hsa-miR-1304-3p | 0.37 |
| hsa-miR-novel-chr9_56521 | 0.34 | hsa-miR-3187-3p | 0.28 | hsa-miR-653-5p | 0.39 |
| hsa-miR-novel-chr8_54829 | 0.34 | hsa-miR-652-3p | 0.28 | hsa-miR-574-5p | 0.39 |
| hsa-miR-novel-chr2_34829 | 0.37 | hsa-miR-5591-3p | 0.29 | hsa-miR-3201 | 0.39 |
| hsa-miR-3182 | 0.38 | hsa-miR-novel-chr12_5743 | 0.29 | hsa-miR-novel-chr12_5743 | 0.40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diallo, I.; Ho, J.; Laffont, B.; Laugier, J.; Benmoussa, A.; Lambert, M.; Husseini, Z.; Soule, G.; Kozak, R.; Kobinger, G.P.; et al. Altered microRNA Transcriptome in Cultured Human Liver Cells upon Infection with Ebola Virus. Int. J. Mol. Sci. 2021, 22, 3792. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073792
Diallo I, Ho J, Laffont B, Laugier J, Benmoussa A, Lambert M, Husseini Z, Soule G, Kozak R, Kobinger GP, et al. Altered microRNA Transcriptome in Cultured Human Liver Cells upon Infection with Ebola Virus. International Journal of Molecular Sciences. 2021; 22(7):3792. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073792
Chicago/Turabian StyleDiallo, Idrissa, Jeffrey Ho, Benoit Laffont, Jonathan Laugier, Abderrahim Benmoussa, Marine Lambert, Zeinab Husseini, Geoff Soule, Robert Kozak, Gary P. Kobinger, and et al. 2021. "Altered microRNA Transcriptome in Cultured Human Liver Cells upon Infection with Ebola Virus" International Journal of Molecular Sciences 22, no. 7: 3792. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073792