
Analysis of DnaK Expression from a Strain of Mycoplasma fermentans in Infected HCT116 Human Colon Carcinoma Cells

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Infection of Human HCT116 Cancer Cells Results in Reduced M. fermentans MF-I1 Replication, Increased Expression of dnaK DNA Copy Number and Detection of Intracellular DnaK
2.2. Analysis of MF-I1 dnaK Transcripts Exposes the Emergence of Bacteria with Several Mutations in Important Regulatory Regions
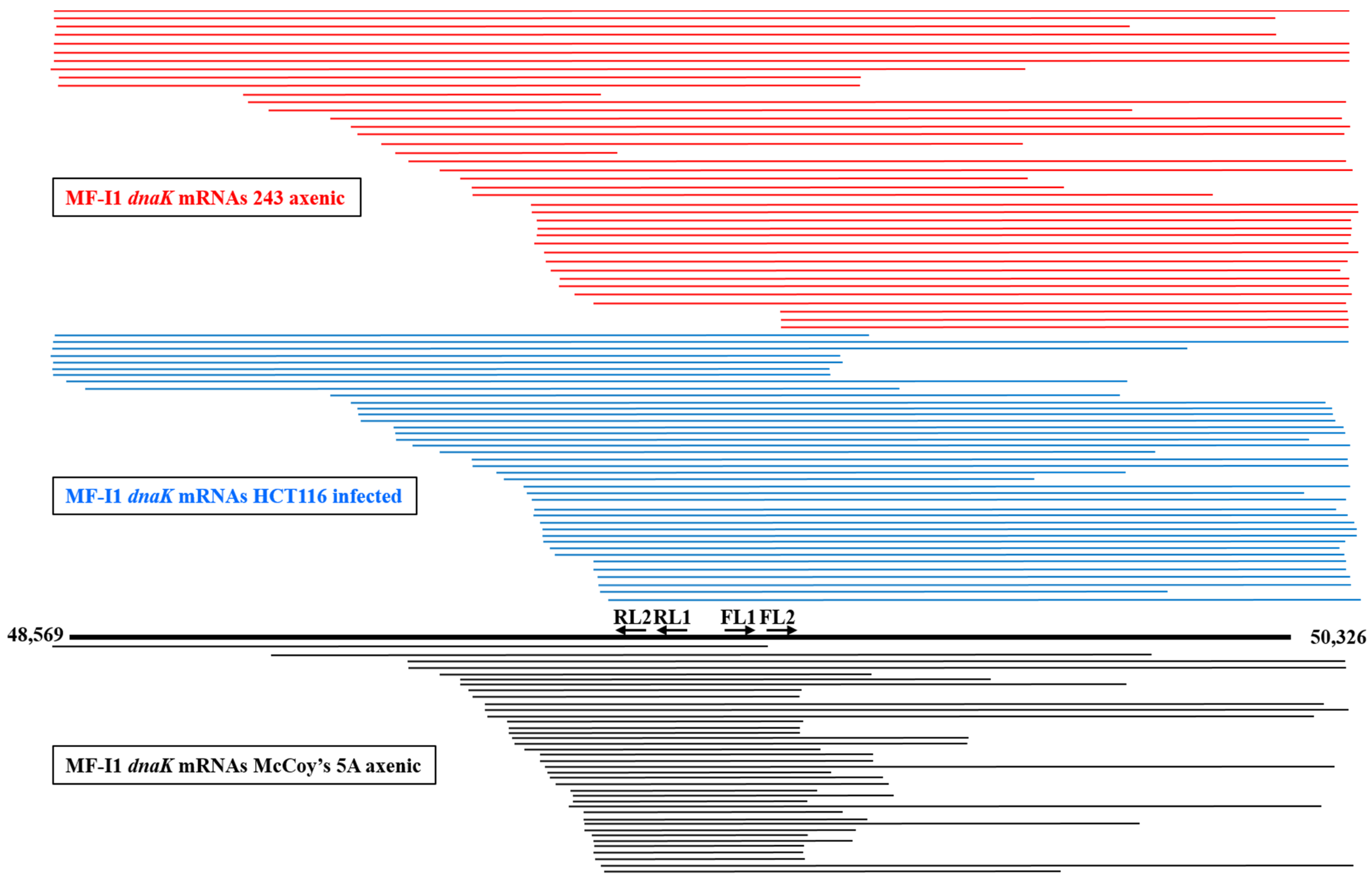
2.3. M. fermentans MF-I1 Strain dnaK mRNA Length Analysis Reveals Presence of Several Types of Transcripts
2.4. Bacteria Grown in Different Conditions Show Differences in dnaK mRNA and Its Internal Promoter
3. Materials and Methods
3.1. Culture of Bacteria
3.2. Colony Forming Unit Assay
3.3. Cells and MF-I1 Infection
3.4. Accession Numbers
3.5. DNA and RNA Extraction
3.6. Analysis of Subcellular Fractions by SDS-PAGE and Immunoblot
3.7. Cloning of MF-I1 dnaK
3.8. Quantitative PCR (qPCR) for MF-I1 and Host Cell Genome Copy Numbers
3.9. Circularized RT-PCR (cRT-PCR) for Analysis of 5′ and 3′ Ends of MF-I1 Isolate mRNA
3.10. Sequence Analyses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| MF-I1 | Mycoplasma fermentans-IHV-substrain 1 |
| PCR | Polymerase Chain Reaction |
| PARP1 | Poly (ADP-Ribose) Polymerase-1 |
| USP10 | Ubiquitin carboxyl-terminal hydrolase 10 |
| CFU | Colony Forming Units |
References
- Warren, J.R.; Marshall, B. Unidentified curved bacilli on gastric epithelium in active chronic gastritis. Lancet 1983, 1, 1273–1275. [Google Scholar]
- Buti, L.; Spooner, E.; Van der Veen, A.G.; Rappuoli, R.; Covacci, A.; Ploegh, H.L. Helicobacter pylori cytotoxin-associated gene A (CagA) subverts the apoptosis-stimulating protein of p53 (ASPP2) tumor suppressor pathway of the host. Proc. Natl. Acad. Sci. USA 2011, 108, 9238–9243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razin, S.; Yogev, D.; Naot, Y. Molecular biology and pathogenicity of mycoplasmas. Microbiol. Mol. Biol. Rev. 1998, 62, 1094–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yavlovich, A.; Katzenell, A.; Tarshis, M.; Higazi, A.A.; Rottem, S. Mycoplasma fermentans binds to and invades HeLa cells: Involvement of plasminogen and urokinase. Infect Immun. 2004, 72, 5004–5011. [Google Scholar] [CrossRef] [Green Version]
- Lo, S.C.; Hayes, M.M.; Kotani, H.; Pierce, P.F.; Wear, D.J.; Newton, P.B., 3rd; Tully, J.G.; Shih, J.W. Adhesion onto and invasion into mammalian cells by mycoplasma penetrans: A newly isolated mycoplasma from patients with AIDS. Mod. Pathol. 1993, 6, 276–280. [Google Scholar]
- Baseman, J.B.; Lange, M.; Criscimagna, N.L.; Giron, J.A.; Thomas, C.A. Interplay between mycoplasmas and host target cells. Microb. Pathog. 1995, 19, 105–116. [Google Scholar] [CrossRef]
- Hegde, S.; Hegde, S.; Spergser, J.; Brunthaler, R.; Rosengarten, R.; Chopra-Dewasthaly, R. In vitro and in vivo cell invasion and systemic spreading of Mycoplasma agalactiae in the sheep infection model. Int. J. Med Microbiol. IJMM 2014, 304, 1024–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunoya, T.; Omori, T.; Tomioka, H.; Umeda, F.; Suzuki, T.; Uetsuka, K. Intracellular Localization of Mycoplasma bovis in the Bronchiolar Epithelium of Experimentally Infected Calves. J. Comp. Pathol. 2020, 176, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Robinson, D.; Davies, H.A.; Sarathchandra, P.; Furr, P.M. Intracellular location of mycoplasmas in cultured cells demonstrated by immunocytochemistry and electron microscopy. Int. J. Exp. Pathol. 1991, 72, 705–714. [Google Scholar]
- Helaine, S.; Thompson, J.A.; Watson, K.G.; Liu, M.; Boyle, C.; Holden, D.W. Dynamics of intracellular bacterial replication at the single cell level. Proc. Natl. Acad. Sci. USA 2010, 107, 3746–3751. [Google Scholar] [CrossRef] [Green Version]
- Steele-Mortimer, O. The Salmonella-containing vacuole: Moving with the times. Curr. Opin. Microbiol. 2008, 11, 38–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik-Kale, P.; Winfree, S.; Steele-Mortimer, O. The bimodal lifestyle of intracellular Salmonella in epithelial cells: Replication in the cytosol obscures defects in vacuolar replication. PLoS ONE 2012, 7, e38732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribet, D.; Cossart, P. How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes Infect. 2015, 17, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, F.; Curreli, S.; Gallo, R.C.; Zella, D. Tampering of Viruses and Bacteria with Host DNA Repair: Implications for Cellular Transformation. Cancers 2021, 13, 241. [Google Scholar] [CrossRef]
- Ainsworth, J.G.; Easterbrook, P.J.; Clarke, J.; Gilroy, C.B.; Taylor-Robinson, D. An association of disseminated Mycoplasma fermentans in HIV-1 positive patients with non-Hodgkin’s lymphoma. Int. J. Std Aids 2001, 12, 499–504. [Google Scholar] [CrossRef]
- Barykova, Y.A.; Logunov, D.Y.; Shmarov, M.M.; Vinarov, A.Z.; Fiev, D.N.; Vinarova, N.A.; Rakovskaya, I.V.; Baker, P.S.; Shyshynova, I.; Stephenson, A.J.; et al. Association of Mycoplasma hominis infection with prostate cancer. Oncotarget 2011, 2, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Henrich, B.; Rumming, M.; Sczyrba, A.; Velleuer, E.; Dietrich, R.; Gerlach, W.; Gombert, M.; Rahn, S.; Stoye, J.; Borkhardt, A.; et al. Mycoplasma salivarium as a Dominant Coloniser of Fanconi Anaemia Associated Oral Carcinoma. PLoS ONE 2014, 9, e92297. [Google Scholar] [CrossRef]
- Jiang, S.; Zhang, S.; Langenfeld, J.; Lo, S.-C.; Rogers, M.B. Mycoplasma infection transforms normal lung cells and induces bone morphogenetic protein 2 expression by post-transcriptional mechanisms. J. Cell. Biochem. 2008, 104, 580–594. [Google Scholar] [CrossRef]
- Namiki, K.; Goodison, S.; Porvasnik, S.; Allan, R.W.; Iczkowski, K.A.; Urbanek, C.; Reyes, L.; Sakamoto, N.; Rosser, C.J. Persistent Exposure to Mycoplasma Induces Malignant Transformation of Human Prostate Cells. PLoS ONE 2009, 4, e6872. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Tsai, S.; Lo, S.-C. Alteration of gene expression profiles during mycoplasma-induced malignant cell transformation. BMC Cancer 2006, 6, 116. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Scheblyakov, D.V.; Zubkova, O.V.; Shmarov, M.M.; Rakovskaya, I.V.; Gurova, K.V.; Tararova, N.D.; Burdelya, L.G.; Naroditsky, B.S.; Ginzburg, A.L.; et al. Mycoplasma infection suppresses p53, activates NF-κB and cooperates with oncogenic Ras in rodent fibroblast transformation. Oncogene 2008, 27, 4521–4531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calloni, G.; Chen, T.; Schermann, S.M.; Chang, H.C.; Genevaux, P.; Agostini, F.; Tartaglia, G.G.; Hayer-Hartl, M.; Hartl, F.U. DnaK functions as a central hub in the E. coli chaperone network. Cell Rep. 2012, 1, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Saibil, H. Chaperone machines for protein folding, unfolding and disaggregation. Nat. Rev. Mol. Cell Biol. 2013, 14, 630–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zella, D.; Curreli, S.; Benedetti, F.; Krishnan, S.; Cocchi, F.; Latinovic, O.S.; Denaro, F.; Romerio, F.; Djavani, M.; Charurat, M.E.; et al. Mycoplasma promotes malignant transformation in vivo, and its DnaK, a bacterial chaperone protein, has broad oncogenic properties. Proc. Natl. Acad. Sci. USA 2018, 115, E12005–E12014. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Luo, K.; Zhang, L.; Cheville, J.C.; Lou, Z. USP10 Regulates p53 Localization and Stability by Deubiquitinating p53. Cell 2010, 140, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, F.; Cocchi, F.; Latinovic, O.S.; Curreli, S.; Krishnan, S.; Munawwar, A.; Gallo, R.C.; Zella, D. Role of Mycoplasma Chaperone DnaK in Cellular Transformation. Int. J. Mol. Sci. 2020, 21, 1311. [Google Scholar] [CrossRef] [Green Version]
- Costa, T.R.D.; Felisberto-Rodrigues, C.; Meir, A.; Prevost, M.S.; Redzej, A.; Trokter, M.; Waksman, G. Secretion systems in Gram-negative bacteria: Structural and mechanistic insights. Nat. Rev. Microbiol. 2015, 13, 343. [Google Scholar] [CrossRef]
- Holland, I.B. Translocation of bacterial proteins—An overview. Biochim. Biophys. Acta BBA Mol. Cell Res. 2004, 1694, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, F.; Krishnan, S.; Cocchi, F.; Tettelin, H.; Gallo, R.C.; Zella, D.; Curreli, S. Proteome analysis of Mycoplasma fermentans cultured under aerobic and anaerobic conditions. Transl. Med. Commun. 2019, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Musatovova, O.; Dhandayuthapani, S.; Baseman, J.B. Transcriptional heat shock response in the smallest known self-replicating cell, Mycoplasma genitalium. J. Bacteriol. 2006, 188, 2845–2855. [Google Scholar] [CrossRef] [Green Version]
- Perez-Rodriguez, R.; Fisher, A.C.; Perlmutter, J.D.; Hicks, M.G.; Chanal, A.; Santini, C.L.; Wu, L.F.; Palmer, T.; DeLisa, M.P. An essential role for the DnaK molecular chaperone in stabilizing over-expressed substrate proteins of the bacterial twin-arginine translocation pathway. J. Mol. Biol. 2007, 367, 715–730. [Google Scholar] [CrossRef] [PubMed]
- Guisbert, E.; Herman, C.; Lu, C.Z.; Gross, C.A. A chaperone network controls the heat shock response in E. coli. Genes Dev. 2004, 18, 2812–2821. [Google Scholar] [CrossRef] [Green Version]
- Andreev, J.; Borovsky, Z.; Rosenshine, I.; Rottem, S. Invasion of HeLa cells by Mycoplasma penetrans and the induction of tyrosine phosphorylation of a 145-kDa host cell protein. FEMS Microbiol. Lett. 1995, 132, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Blötz, C.; Stülke, J. Glycerol metabolism and its implication in virulence in Mycoplasma. FEMS Microbiol. Rev. 2017, 41, 640–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Dordet-Frisoni, E.; Gillard, L.; Ba, A.; Hygonenq, M.-C.; Sagné, E.; Nouvel, L.X.; Maillard, R.; Assié, S.; Guo, A.; et al. Extracellular DNA: A Nutritional Trigger of Mycoplasma bovis Cytotoxicity. Front. Microbiol. 2019, 10, 2753. [Google Scholar] [CrossRef]
- Chang, L.J.; Chen, W.H.; Minion, F.C.; Shiuan, D. Mycoplasmas regulate the expression of heat-shock protein genes through CIRCE-HrcA interactions. Biochem. Biophys. Res. Commun. 2008, 367, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Narberhaus, F. Negative regulation of bacterial heat shock genes. Mol. Microbiol. 1999, 31, 1–8. [Google Scholar] [CrossRef]
- De Hoon, M.J.; Makita, Y.; Nakai, K.; Miyano, S. Prediction of transcriptional terminators in Bacillus subtilis and related species. PLoS Comput. Biol. 2005, 1, e25. [Google Scholar] [CrossRef]
- Portnoy, V.; Schuster, G. Mycoplasma gallisepticum as the first analyzed bacterium in which RNA is not polyadenylated. FEMS Microbiol. Lett. 2008, 283, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Guell, M.; van Noort, V.; Yus, E.; Chen, W.H.; Leigh-Bell, J.; Michalodimitrakis, K.; Yamada, T.; Arumugam, M.; Doerks, T.; Kuhner, S.; et al. Transcriptome complexity in a genome-reduced bacterium. Science 2009, 326, 1268–1271. [Google Scholar] [CrossRef]
- Carpousis, A.J. The Escherichia coli RNA degradosome: Structure, function and relationship in other ribonucleolytic multienzyme complexes. Biochem. Soc. Trans. 2002, 30, 150–155. [Google Scholar] [CrossRef]
- Commichau, F.M.; Rothe, F.M.; Herzberg, C.; Wagner, E.; Hellwig, D.; Lehnik-Habrink, M.; Hammer, E.; Volker, U.; Stulke, J. Novel activities of glycolytic enzymes in Bacillus subtilis: Interactions with essential proteins involved in mRNA processing. Mol. Cell. Proteom. MCP 2009, 8, 1350–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Slomovic, S.; Schuster, G. Circularized RT-PCR (cRT-PCR): Analysis of the 5′ ends, 3′ ends, and poly(A) tails of RNA. Methods Enzymol. 2013, 530, 227–251. [Google Scholar] [CrossRef]
- Song, M.G.; Bail, S.; Kiledjian, M. Multiple Nudix family proteins possess mRNA decapping activity. RNA 2013, 19, 390–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, P.P.; Barquist, L.; Bateman, A.; Nawrocki, E.P.; Weinberg, Z. RNIE: Genome-wide prediction of bacterial intrinsic terminators. Nucleic Acids Res. 2011, 39, 5845–5852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2--a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitrov, D.S.; Franzoso, G.; Salman, M.; Blumenthal, R.; Tarshis, M.; Barile, M.F.; Rottem, S. Mycoplasma fermentans (Incognitus Strain) Cells Are Able to Fuse with T Lymphocytes. Clin. Infect. Dis. 1993, 17, S305–S308. [Google Scholar] [CrossRef]
- Hartl, F.U.; Hayer-Hartl, M. Converging concepts of protein folding in vitro and in vivo. Nat. Struct. Mol. Biol. 2009, 16, 574–581. [Google Scholar] [CrossRef]
- Roncarati, D.; Scarlato, V. Regulation of heat-shock genes in bacteria: From signal sensing to gene expression output. FEMS Microbiol. Rev. 2017, 41, 549–574. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, J.A.; Khodursky, A.B.; Lin, P.H.; Lin-Chao, S.; Cohen, S.N. Global analysis of mRNA decay and abundance in Escherichia coli at single-gene resolution using two-color fluorescent DNA microarrays. Proc. Natl. Acad. Sci. USA 2002, 99, 9697–9702. [Google Scholar] [CrossRef] [Green Version]
- Jain, C.; Deana, A.; Belasco, J.G. Consequences of RNase E scarcity in Escherichia coli. Mol. Microbiol. 2002, 43, 1053–1064. [Google Scholar] [CrossRef]
- Hui, M.P.; Foley, P.L.; Belasco, J.G. Messenger RNA degradation in bacterial cells. Annu. Rev. Genet. 2014, 48, 537–559. [Google Scholar] [CrossRef] [Green Version]
- Matin, A. The molecular basis of carbon-starvation-induced general resistance in Escherichia coli. Mol. Microbiol. 1991, 5, 3–10. [Google Scholar] [CrossRef]
- Jin, D.J.; Cagliero, C.; Zhou, Y.N. Growth rate regulation in Escherichia coli. FEMS Microbiol. Rev. 2012, 36, 269–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klumpp, S.; Zhang, Z.; Hwa, T. Growth rate-dependent global effects on gene expression in bacteria. Cell 2009, 139, 1366–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bremer, H.; Dennis, P.P. Modulation of Chemical Composition and Other Parameters of the Cell at Different Exponential Growth Rates. Ecosal Plus 2008, 3. [Google Scholar] [CrossRef] [PubMed]
- Cooper, S.; Helmstetter, C.E. Chromosome replication and the division cycle of Escherichia coli Br. J. Mol. Biol. 1968, 31, 519–540. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Point | Internalized MF-I1 (CFU/106 HCT116 Cells) | MF-I1 in Supernatant from HCT116 Culture (CFU/mL) | MF-I1 in McCoy’s 5A Medium Axenic Culture (CFU/mL) |
|---|---|---|---|
| Day 1 | 8242 ± 2224 | 8193 ± 3461 | 905 ± 75 |
| Day 3 | 62,647 ± 3512 | 33,303 ± 3461 | 3568 ± 1275 |
| Day 6 | 26,371 ± 596 | 336,665 ± 42,757 | 7874 ± 903 |
| dnaK Promoters | −35 Box | −10 Box |
|---|---|---|
| Pr. 1 | (−173) TTGATT | (−153) ATTTAATTT |
| Pr. 2 | (+197) TTGCAT | (+219) GGGTACAAA |
| Pr. 3 | (+617) ATGAAA | (+638) TGCTAAAAT |
| Pr.4 | (+1205) TTGAAA | (+1224) TGTTACAAT |
| CLONE | POINT MUTATIONS | INSERTIONS | DELETIONS | |
|---|---|---|---|---|
| Nucleotide substitution | Amino acid substitution | |||
| H5 | A1190G | Glu397Gly | ||
| H6 | C1578G | Asn526Lys | ||
| H8 | A756G | Leu255Leu | ||
| H9 | A1190G | Glu397Gly | ||
| H10 | A1190G | Glu397Gly | ||
| H11 | A1190G | Glu397Gly | ||
| H12 | G373T, T600C, A1236G | Ala125Ser, Gly200Gly, Lys412Lys | ||
| H16 | A1488G | Glu483Gly | ||
| H19 | A1512G | Ala504Ala | ||
| H21 | A1348 (Lys450) | |||
| H24 | A1299G | Thr433Thr | ||
| H28 | T1304C, G1357A | Ile435Thr, Ala453Thr | ||
| H32 | G1321A | Gly441Arg | ||
| H33 | A1714G | Ser572Gly | ||
| H35 | G1471A, A1484G | Glu491Lys, Asn495Ser | ||
| H39 | A1211G | Asn404Thr | ||
| H41 | A1200G | Thr400Thr | ||
| C2 | A1368G, T1564C | Ile456Met, Val522Ala | ||
| C9 | 790G (His264) | |||
| C10 | T648C | Ile216Ile | ||
| C11 | G4A, C1309T | Pro2Ser, Ala437Thr | ||
| C13 | C528G | Phe176Leu | ||
| C15 | T1263C | Ala421Ala | ||
| C16 | G1715A | Ser572Asn | ||
| C20 | C732T | Asp244Asp | ||
| C24 | A1075G, T1076C | Ile359Ala | ||
| C26 | A11G, T57C | Glu4Gly, Ala19Ala | ||
| C34 | A1503C | Lys501Asn | ||
| C35 | C761T, A1396G | Ser254Leu, Thr466Ala | ||
| C37 | A37G | Thr13Gly | ||
| C40 | C710T | Ala237Val | ||
| M25 | T1062G | Val354Val | ||
| M26 | T1070C | Arg390Arg | ||
| M30 | G632T | Trp211Leu | ||
| M34 | T764G, G765T | Leu259Leu, Val260Phe | ||
| M35 | C1298T | Thr433Ile | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curreli, S.; Tettelin, H.; Benedetti, F.; Krishnan, S.; Cocchi, F.; Reitz, M.; Gallo, R.C.; Zella, D. Analysis of DnaK Expression from a Strain of Mycoplasma fermentans in Infected HCT116 Human Colon Carcinoma Cells. Int. J. Mol. Sci. 2021, 22, 3885. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083885
Curreli S, Tettelin H, Benedetti F, Krishnan S, Cocchi F, Reitz M, Gallo RC, Zella D. Analysis of DnaK Expression from a Strain of Mycoplasma fermentans in Infected HCT116 Human Colon Carcinoma Cells. International Journal of Molecular Sciences. 2021; 22(8):3885. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083885
Chicago/Turabian StyleCurreli, Sabrina, Hervé Tettelin, Francesca Benedetti, Selvi Krishnan, Fiorenza Cocchi, Marvin Reitz, Robert C. Gallo, and Davide Zella. 2021. "Analysis of DnaK Expression from a Strain of Mycoplasma fermentans in Infected HCT116 Human Colon Carcinoma Cells" International Journal of Molecular Sciences 22, no. 8: 3885. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083885