
BCOR Internal Tandem Duplication Expression in Neural Stem Cells Promotes Growth, Invasion, and Expression of PRC2 Targets

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
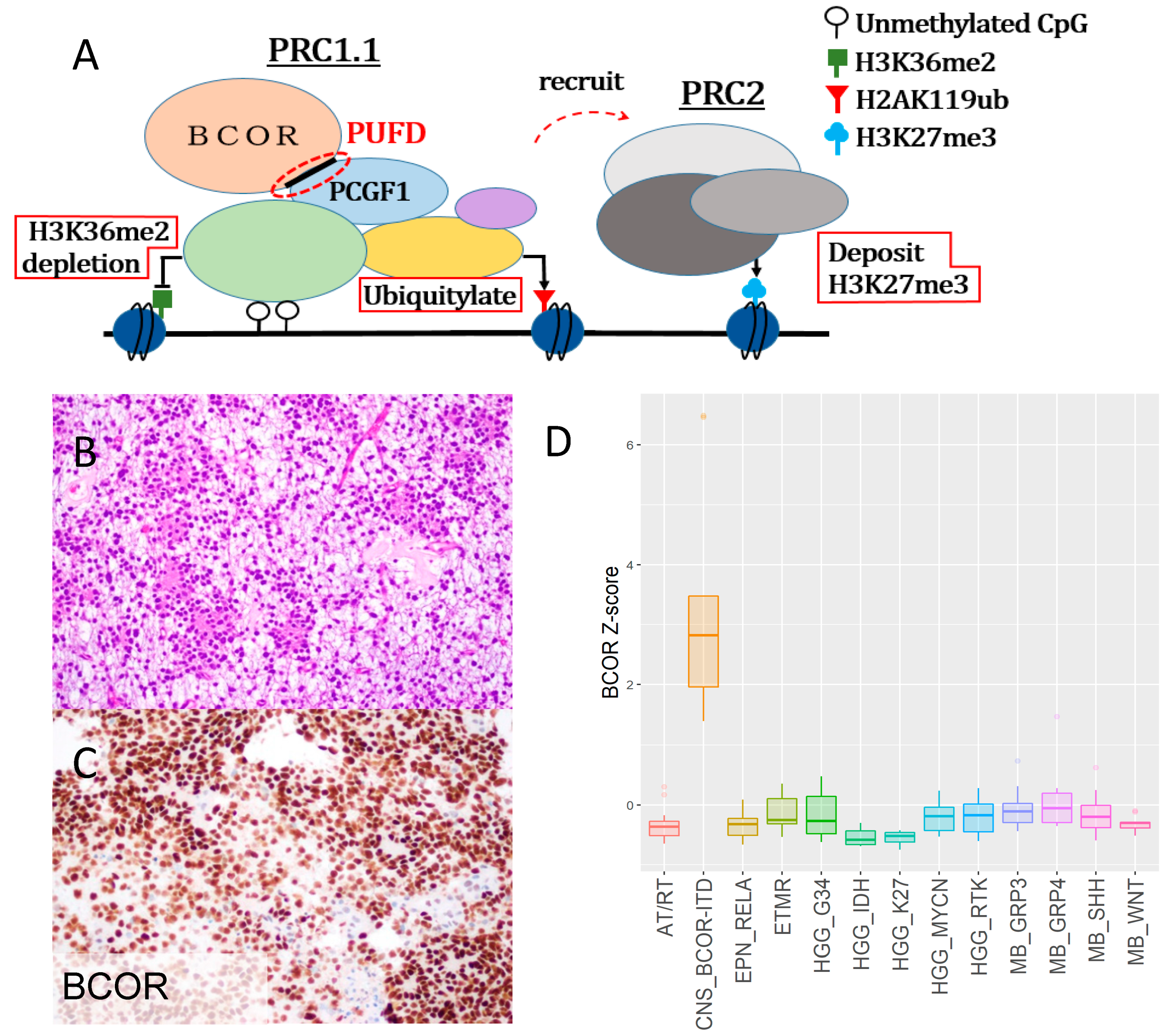
2.1. CNS-BCOR ITD is an Aggressive Tumor with Overexpression of Mutant-BCOR
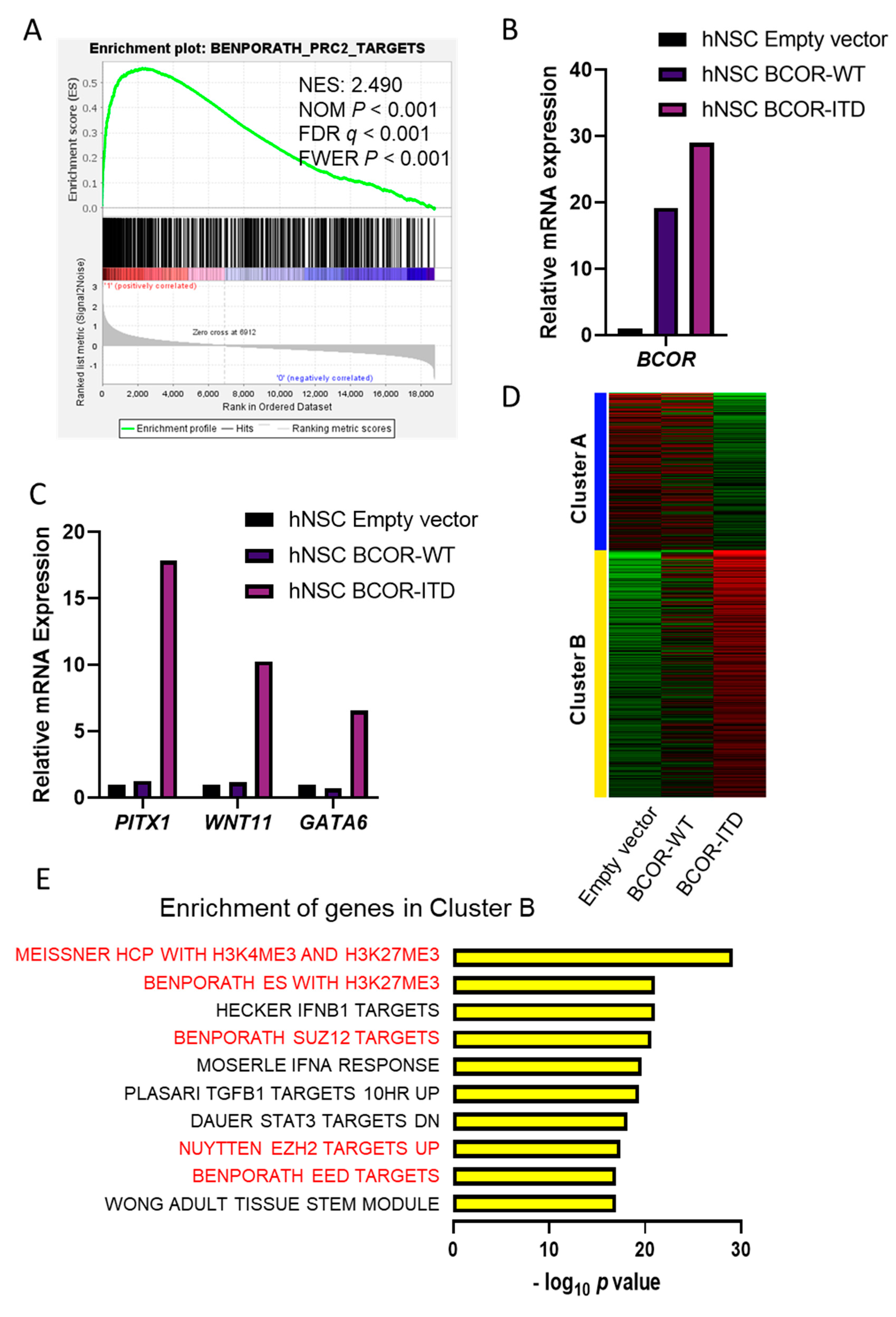
2.2. CNS BCOR-ITD Shows Upregulation of PRC2 Targets, and hNSC with BCOR-ITD Recapitulated This Induction
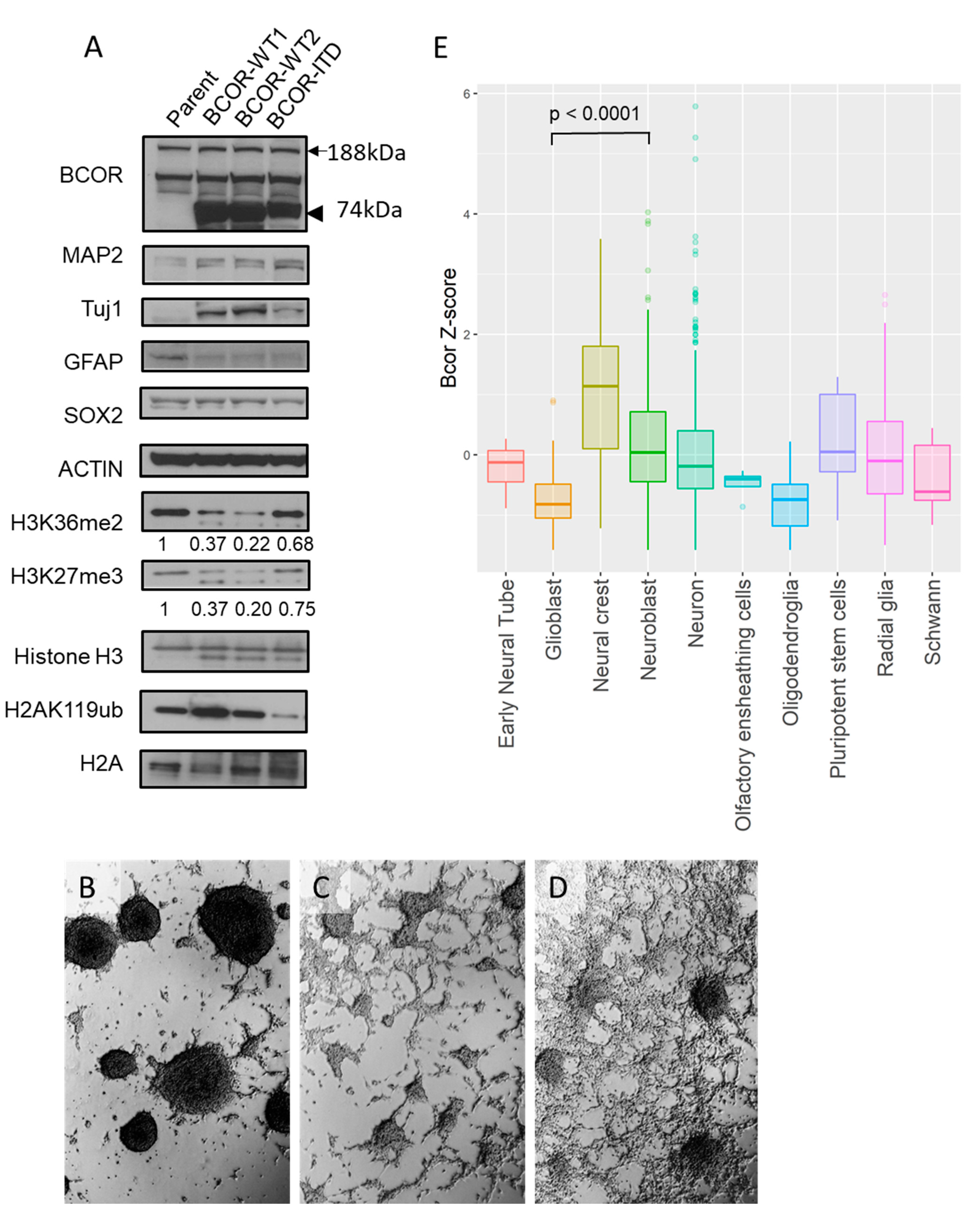
2.3. Both BCORE7–15-WT and BCORE7–15-ITD Alter Differentiation of mNSC
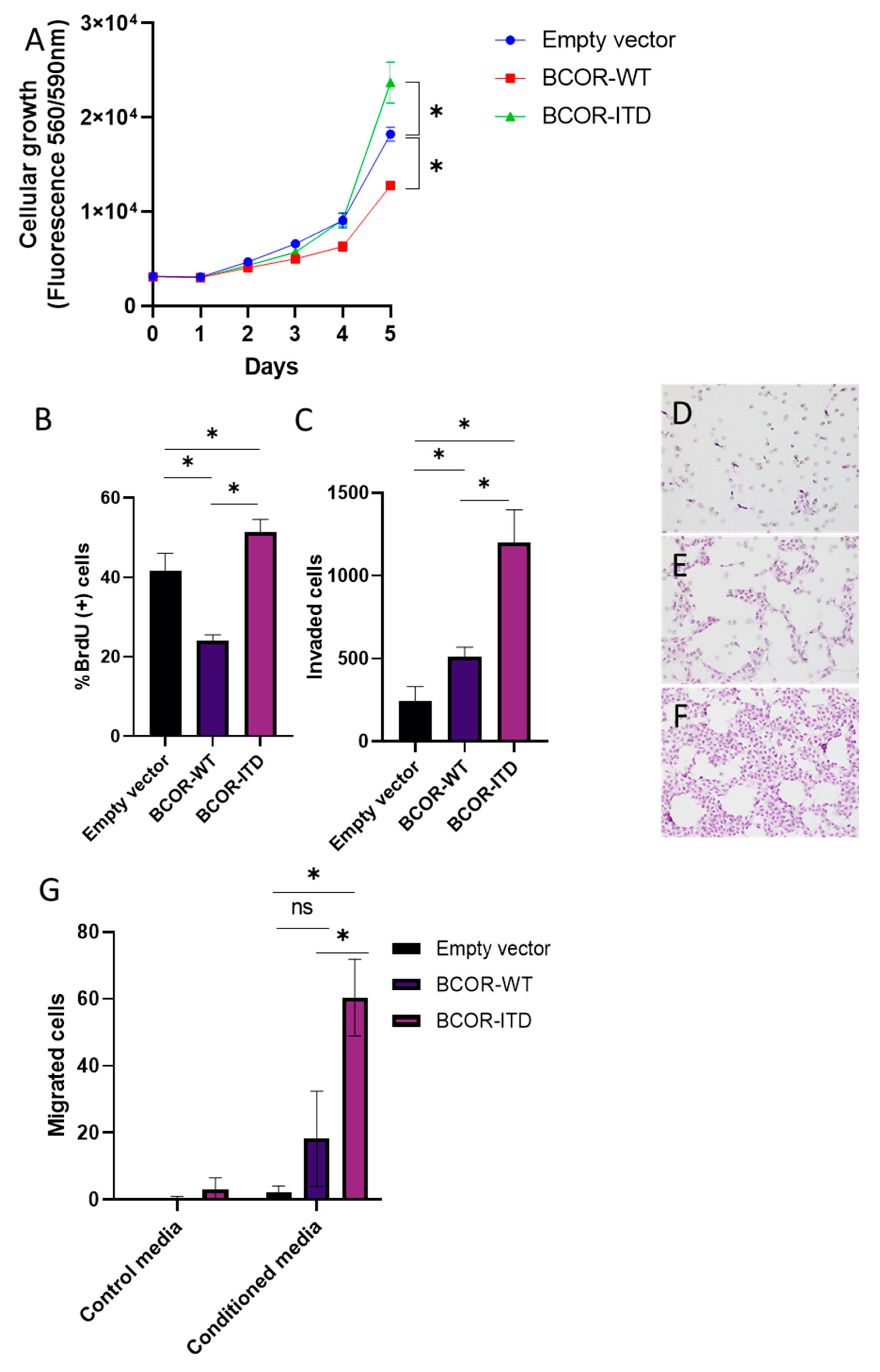
2.4. BCORE7–15-ITD Overexpression Increased Cellular Growth, Invasion, and Migration of mNSC, while BCORE7–15-WT Decreased Cellular Growth
3. Discussion and Conclusions
4. Materials and Methods
4.1. CNS-BCOR ITD Tumor Samples
4.2. Survival Analysis
4.3. BCOR Expression in Primary CNS Tumors
4.4. Gene-Set Enrichment Analysis (GSEA)
4.5. Generation of hNSC Models
4.6. Quantitative RT-PCR (RTqPCR)
4.7. RNA Sequencing of hNSC Models
4.8. Examination Bcor Expression in Developing Brains Using Public Single-Cell RNA-seq Data
4.9. Generation of mNSC Models
4.10. Western Blotting and Antibodies
4.11. Growth and Proliferation Assays
4.12. Transwell Invasion and Migration Assays
4.13. Colony Formation Assay
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BCL6 | B-cell lymphoma 6 protein |
| BCOR-WT | Wildtype BCOR |
| BCOR-ITD | BCOR internal tandem duplication |
| BCORE7–15 | BCOR exons 7 to 15 |
| CCSK | Clear cell sarcoma of the kidney |
| CNS | Central Nervous System |
| CNS-BCOR ITD | CNS tumor with BCL6-corepressor internal tandem duplication |
| cIMPACT | Consortium to Inform Molecular and Practical Approaches to CNS Tumor Taxonomy |
| FWER | Family-wise error rate |
| GSEA | Gene-set enrichment analysis |
| hNSC | Human neural stem cells |
| IGF1R | Insulin Growth Factor 1 Receptor |
| IGF2 | Insulin Growth Factor 2 |
| mNSC | Murine neural stem cells |
| NSC | Neural stem cells |
| OFCD | Oculo-facio-cardio-dental syndrome |
| PRC | Polycomb repressive complex |
| PUFD | PCGF Ub-like fold discriminator |
| RTqPCR | Reverse transcription quantitative polymerase chain reaction |
| RNA-seq | RNA sequencing |
References
- Sturm, D.; Orr, B.A.; Toprak, U.H.; Hovestadt, V.; Jones, D.T.; Capper, D.; Sill, M.; Buchhalter, I.; Northcott, P.A.; Leis, I.; et al. New Brain Tumor Entities Emerge from Molecular Classification of CNS-PNETs. Cell 2016, 164, 1060–1072. [Google Scholar] [CrossRef] [Green Version]
- Louis, D.N.; Wesseling, P.; Aldape, K.; Brat, D.J.; Capper, D.; Cree, L.A.; Eberhart, G.G.; Figarella-Brangeret, D.; Fouladi, M.; Fuller, G.N.; et al. cIMPACT-NOW update 6: New entity and diagnostic principle recommendations of the cIMPACT-Utrecht meeting on future CNS tumor classification and grading. Brain Pathol. 2020, 30, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez Castro, L.N.; Wesseling, P. The cIMPACT-NOW updates and their significance to current neuro-oncology practice. Neuro-Oncol. Pract. 2021, 8, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Appay, R.; Macagno, N.; Padovani, L.; Korshunov, A.; Kool, M.; André, N.; Scavarda, D.; Pietsch, T.; Figarella-Branger, D. HGNET-BCOR Tumors of the Cerebellum: Clinicopathologic and Molecular Characterization of 3 Cases. Am. J. Surg. Pathol. 2017, 41, 1254–1260. [Google Scholar] [CrossRef] [PubMed]
- Paret, C.; Theruvath, J.; Russo, A.; Kron, B.; El Malki, K.; Lehmann, N.; Wingerter, A.; Neu, M.A.; Gerhold-Ay, A.; Wagner, W.; et al. Activation of the basal cell carcinoma pathway in a patient with CNS HGNET-BCOR diagnosis: Consequences for personalized targeted therapy. Oncotarget 2016, 7, 83378–83391. [Google Scholar] [CrossRef] [Green Version]
- Paret, C.; Russo, A.; Otto, H.; Mayer, A.; Zahnreich, S.; Wagner, W.; Samuel, D.; Scharnhorst, D.; Solomon, D.A.; Dhall, G.; et al. Personalized therapy: CNS HGNET-BCOR responsiveness to arsenic trioxide combined with radiotherapy. Oncotarget 2017, 8, 114210–114225. [Google Scholar] [CrossRef] [Green Version]
- Vewinger, N.; Huprich, S.; Seidmann, L.; Russo, A.; Alt, F.; Bender, H.; Sommer, C.; Samuel, D.; Lehmann, N.; Backes, N.; et al. Igf1r is a potential new therapeutic target for hgnet-bcor brain tumor patients. Int. J. Mol. Sci. 2019, 20, 3027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, Y.; Nobusawa, S.; Nakata, S.; Nakada, M.; Arakawa, Y.; Mineharu, Y.; Sugita, Y.; Yoshioka, T.; Araki, A.; Satoshi, N.; et al. CNS high-grade neuroepithelial tumor with BCOR internal tandem duplication: A comparison with its counterparts in the kidney and soft tissue. Brain Pathol. 2018, 28, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Ferris, S.P.; Vega, J.V.; Aboian, M.; Lee, J.C.; Van Ziffle, J.; Onodera, C.; Grenert, J.P.; Saunders, T.; Chen, Y.; Banerjee, A.; et al. High-grade neuroepithelial tumor with BCOR exon 15 internal tandem duplication—a comprehensive clinical, radiographic, pathologic, and genomic analysis. Brain Pathol. 2020, 30, 46–62. [Google Scholar] [CrossRef] [Green Version]
- Kirkman, M.A.; Pickles, J.C.; Fairchild, A.R.; Avery, A.; Pietsch, T.; Jacques, T.S. Early Wound Site Seeding in a Patient with Central Nervous System High-Grade Neuroepithelial Tumor with BCOR Alteration. World Neurosurg. 2018, 116, 279–284. [Google Scholar] [CrossRef]
- Al-Battashi, A.; Al Hajri, Z.; Perry, A.; Al-Kindi, H.; Al-Ghaithi, I.A. Cerebellar High-Grade Neuroepithelial Tumour with BCOR Alteration in a five-year-old Child: A case report. Sultan. Qaboos. Univ. Med. J. 2019, 19, e153–e156. [Google Scholar] [CrossRef] [PubMed]
- Haberler, C.; Reiniger, L.; Rajnai, H.; Kalev, O.; Gelpi, E.; Tamesberger, M.; Pietsch, T. Case of the month 1-2019: CNS high-grade neuroepithelial tumor with BCOR alteration. Clin. Neuropathol. 2019, 38, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Bremer, J.; Kottke, R.; Johann, P.D.; Von Hoff, K.; Brazzola, P.; Grotzer, M.A.; Kool, M.; Rushing, E.; Gerber, N.U. A single supratentorial high-grade neuroepithelial tumor with two distinct BCOR mutations, exceptionally long complete remission and survival. Pediatr. Blood Cancer 2020, 67, e28384. [Google Scholar] [CrossRef] [PubMed]
- Łastowska, M.; Trubicka, J.; Sobocińska, A.; Wojtas, B.; Niemira, M.; Szałkowska, A.; Krętowski, A.; Karkucińska-Więckowska, A.; Kaleta, M.; Ejmont, M.; et al. Molecular identification of CNS NB-FOXR2, CNS EFT-CIC, CNS HGNET-MN1 and CNS HGNET-BCOR pediatric brain tumors using tumor-specific signature genes. Acta. Neuropathol. Commun. 2020, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Mitra, S.; Sugur, H.; Vazhayil, V.; Rao, B.R.M.; Annayappa, S.K.; Nandeesh, B.N.; Yasha, T.C.; Santosh, V. Central nervous system high grade neuroepithelial tumor with BCOR immunopositivity: Is there a molecular heterogeneity? Brain Tumor. Pathol. 2021, 38, 41–49. [Google Scholar] [CrossRef]
- Fukuoka, K.; Kanemura, Y.; Shofuda, T.; Fukushima, S.; Yamashita, S.; Narushima, D.; Kato, M.; Honda-Kitahara, M.; Ichikawa, H.; Kohno, T.; et al. Significance of molecular classification of ependymomas: C11orf95-RELA fusion-negative supratentorial ependymomas are a heterogeneous group of tumors. Acta Neuropathol. Commun. 2018, 6, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno-Yokohata, H.; Okita, H.; Nakasato, K.; Akimoto, S.; Hata, J.-I.; Koshinaga, T.; Fukuzawa, M.; Kiyokawa, N. Consistent in-frame internal tandem duplications of BCOR characterize clear cell sarcoma of the kidney. Nat. Genet. 2015, 47, 861–863. [Google Scholar] [CrossRef]
- Roy, A.; Kumar, V.; Zorman, B.; Fang, E.; Haines, K.M.; Doddapaneni, H.; Hampton, O.A.; White, S.; Bavle, A.A.; Patel, N.R.; et al. Recurrent internal tandem duplications of BCOR in clear cell sarcoma of the kidney. Nat. Commun. 2015, 6, 8891. [Google Scholar] [CrossRef]
- Wang, Z.; Gearhart, M.D.; Lee, Y.W.; Kumar, I.; Ramazanov, B.; Zhang, Y.; Hernandez, C.; Lu, A.Y.; Neuenkirchen, N.; Deng, J.J.; et al. A Non-canonical BCOR-PRC1.1 Complex Represses Differentiation Programs in Human ESCs. Cell Stem. 2018, 22, 235–251.e9. [Google Scholar] [CrossRef] [Green Version]
- Blackledge, N.P.; Farcas, A.M.; Kondo, T.; King, H.W.; McGouran, J.F.; Hanssen, L.L.; Ito, S.; Cooper, S.; Kondo, K.; Koseki, Y.; et al. Variant PRC1 complex-dependent H2A ubiquitylation drives PRC2 recruitment and polycomb domain formation. Cell 2014, 157, 1445–1459. [Google Scholar] [CrossRef] [Green Version]
- Wamstad, J.A.; Corcoran, C.M.; Keating, A.M.; Bardwell, V.J. Role of the transcriptional corepressor Bcor in embryonic stem cell differentiation and early embryonic development. PLoS ONE 2008, 3, e2814. [Google Scholar] [CrossRef] [Green Version]
- Ng, D.; Thakker, N.; Corcoran, C.M.; Donnai, D.; Perveen, R.; Schneider, A.; Hadley, D.W.; Tifft, C.J.; Zhang, L.; Wilkie, A.O.M.; et al. Oculofaciocardiodental and Lenz microphthalmia syndromes result from distinct classes of mutations in BCOR. Nat. Genet. 2004, 36, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Tiberi, L.; Bonnefont, J.; van den Ameele, J.; Le Bon, S.D.; Herpoel, A.; Biheu, A.; Baron, B.W.; Vanderhaeghen, P. A BCL6/BCOR/SIRT1 complex triggers neurogenesis and suppresses medulloblastoma by repressing Sonic Hedgehog signaling. Cancer Cell 2014, 26, 797–812. [Google Scholar] [CrossRef] [Green Version]
- Kutscher, L.M.; Okonechnikov, K.; Batora, N.V.; Clark, J.; Silva, P.B.; Vouri, M.; Van Rijn, S.; Sieber, L.; Statz, B.; Gearhart, M.D.; et al. Functional loss of a noncanonical BCOR-PRC1.1 complex accelerates SHH-driven medulloblastoma formation. Genes. Dev. 2020, 34, 1161–1176. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.J.; So, J.; Rogers, A.J.; Gregory, G.; Li, J.; Zethoven, M.; Gearhart, M.D.; Bardwell, V.J.; Johnstone, R.W.; Vervoort, S.J.; et al. Bcor loss perturbs myeloid differentiation and promotes leukaemogenesis. Nat. Commun. 2019, 10, 1347. [Google Scholar] [CrossRef]
- Kelly, M.J.; So, J.; Rogers, A.J.; Gregory, G.; Li, J.; Zethoven, M.; Gearhart, M.D.; Bardwell, V.J.; Johnstone, R.W.; Vervoort, S.J.; et al. A single institution experience of the incidence of extracranial metastasis in glioma. J. Clin. Neurosci. 2012, 19, 1511–1515. [Google Scholar] [CrossRef]
- Fan, Z.; Yamaza, T.; Lee, J.S.; Yu, J.; Wang, S.; Fan, G.; Shi, S.; Wang, C.-Y. BCOR regulates mesenchymal stem cell function by epigenetic mechanisms. Nat. Cell Biol. 2009, 11, 1002–1009. [Google Scholar] [CrossRef] [PubMed]
- Buchberger, E.; El Harchi, M.; Payrhuber, D.; Zommer, A.; Schauer, D.; Simonitsch-Klupp, I.; Bilban, M.; Brostjan, C. Overexpression of the transcriptional repressor complex BCL-6/BCoR leads to nuclear aggregates distinct from classical aggresomes. PLoS ONE 2013, 8, e76845. [Google Scholar] [CrossRef] [Green Version]
- Wamstad, J.A.; Bardwell, V.J. Characterization of Bcor expression in mouse development. Gene. Expr. Patterns. 2007, 7, 550–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalli, F.M.; Remke, M.; Rampasek, L.; Peacock, J.; Shih, D.J.; Luu, B.; Garzia, L.; Torchia, J.; Nor, C.; Morrissy, A.S.; et al. Intertumoral Heterogeneity within Medulloblastoma Subgroups. Cancer. Cell 2017, 31, 737–754.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casey, S.; Greene, D.; Hu, R.; Jones, W.W.; Stephanie, L.; David, S.; Mejia, R.; Patro, S.; Piccolo, R.; Romero, A.R.; et al. refine.bio: A resource of uniformly processed publicly available gene expression datasets. Available online: https://www.refine.bio. (accessed on 8 April 2021).
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.-F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1α-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Lopez, W.O.C.; Nikkhah, G.; Kahlert, U.D.; Maciaczyk, D.; Bogiel, T.; Moellers, S.; Schültke, E.; Döbrössy, M.; Maciaczyk, J. Clinical neurotransplantation protocol for Huntington’s and Parkinson’s disease. Restor. Neurol. Neurosci. 2013, 31, 579–595. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinformatics 2018, 19, 534. [Google Scholar] [CrossRef] [Green Version]
- La Manno, G.; Siletti, K.; Furlan, A.; Gyllborg, D.; Vinsland, E.; Langseth, C.M.; Khven, I.; Johnsson, A.; Nilsson, M.; Lönnerberg, P.; et al. Molecular architecture of the developing mouse brain. BioRxiv 2020. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Yuan, M.; Da Silva, A.C.A.L.; Arnold, A.; Okeke, L.; Ames, H.; Correa-Cerro, L.S.; Vizcaino, M.A.; Ho, C.-Y.; Eberhart, C.G.; Rodriguez, F.J. MicroRNA (miR) 125b regulates cell growth and invasion in pediatric low grade glioma. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Nakata, S.; Price, A.; Eberhart, C.; Morris, M. Increased Tau Expression Correlates With IDH Mutation in Infiltrating Gliomas and Impairs Cell Migration. J. Neuropathol. Exp. Neurol. 2020, 79, 493–499. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CNS-BCOR ITD | |
|---|---|
| Gender | Male: Female = 1:1.05 |
| Median Age at Diagnosis | 4 years old (range: 0 to 22 years old) |
| Locations of Tumor (Frequency) | Cerebellum (46%), Cerebral hemisphere (44%), Other posterior fossa (7.5%), Basal ganglia (2.5%) |
| Median Overall Survival | 3 years |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakata, S.; Yuan, M.; Rubens, J.A.; Kahlert, U.D.; Maciaczyk, J.; Raabe, E.H.; Eberhart, C.G. BCOR Internal Tandem Duplication Expression in Neural Stem Cells Promotes Growth, Invasion, and Expression of PRC2 Targets. Int. J. Mol. Sci. 2021, 22, 3913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083913
Nakata S, Yuan M, Rubens JA, Kahlert UD, Maciaczyk J, Raabe EH, Eberhart CG. BCOR Internal Tandem Duplication Expression in Neural Stem Cells Promotes Growth, Invasion, and Expression of PRC2 Targets. International Journal of Molecular Sciences. 2021; 22(8):3913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083913
Chicago/Turabian StyleNakata, Satoshi, Ming Yuan, Jeffrey A. Rubens, Ulf D. Kahlert, Jarek Maciaczyk, Eric H. Raabe, and Charles G. Eberhart. 2021. "BCOR Internal Tandem Duplication Expression in Neural Stem Cells Promotes Growth, Invasion, and Expression of PRC2 Targets" International Journal of Molecular Sciences 22, no. 8: 3913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083913