
Chronic Ouabain Prevents Na,K-ATPase Dysfunction and Targets AMPK and IL-6 in Disused Rat Soleus Muscle


 ,
,
Abstract
:1. Introduction
2. Results
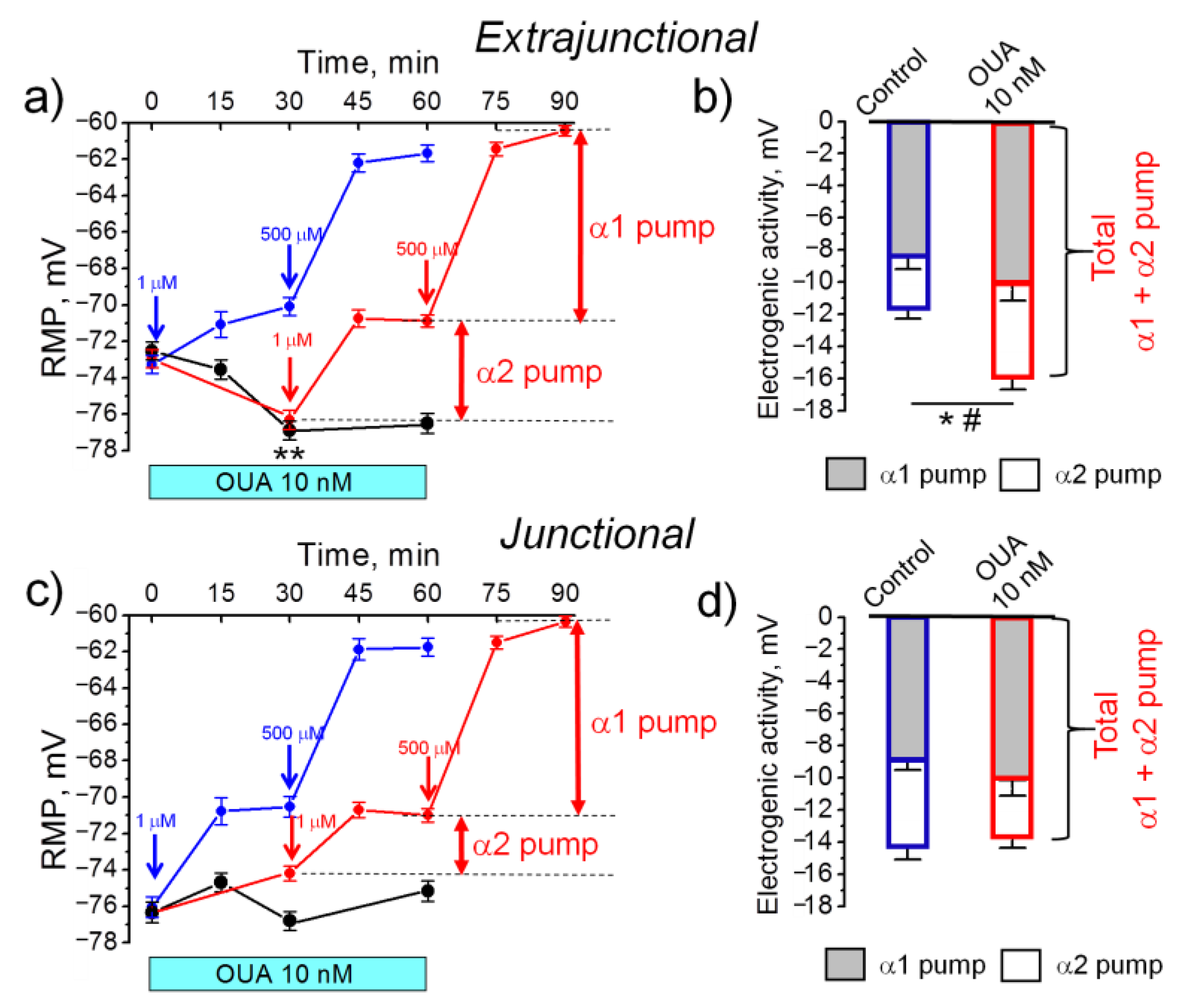
2.1. An Acute Exposure to Nanomolar Ouabain Hyperpolarizes Sarcolemma via Enhanced Electrogenic Activity of the α2 Na,K-ATPase
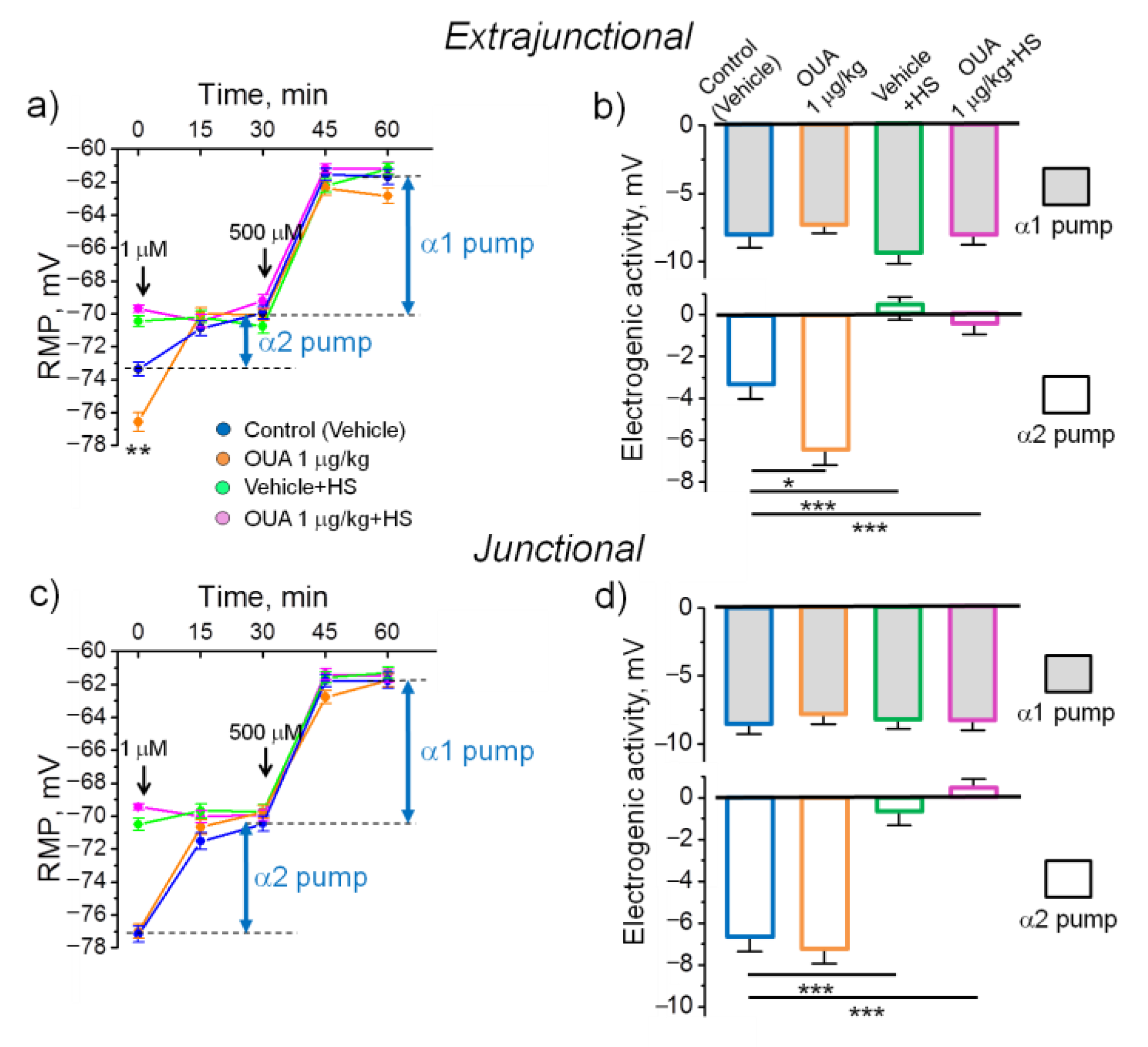
2.2. A Single Injection of Ouabain Does Not Prevent HS-Induced Disturbances in Muscle Electrogenesis
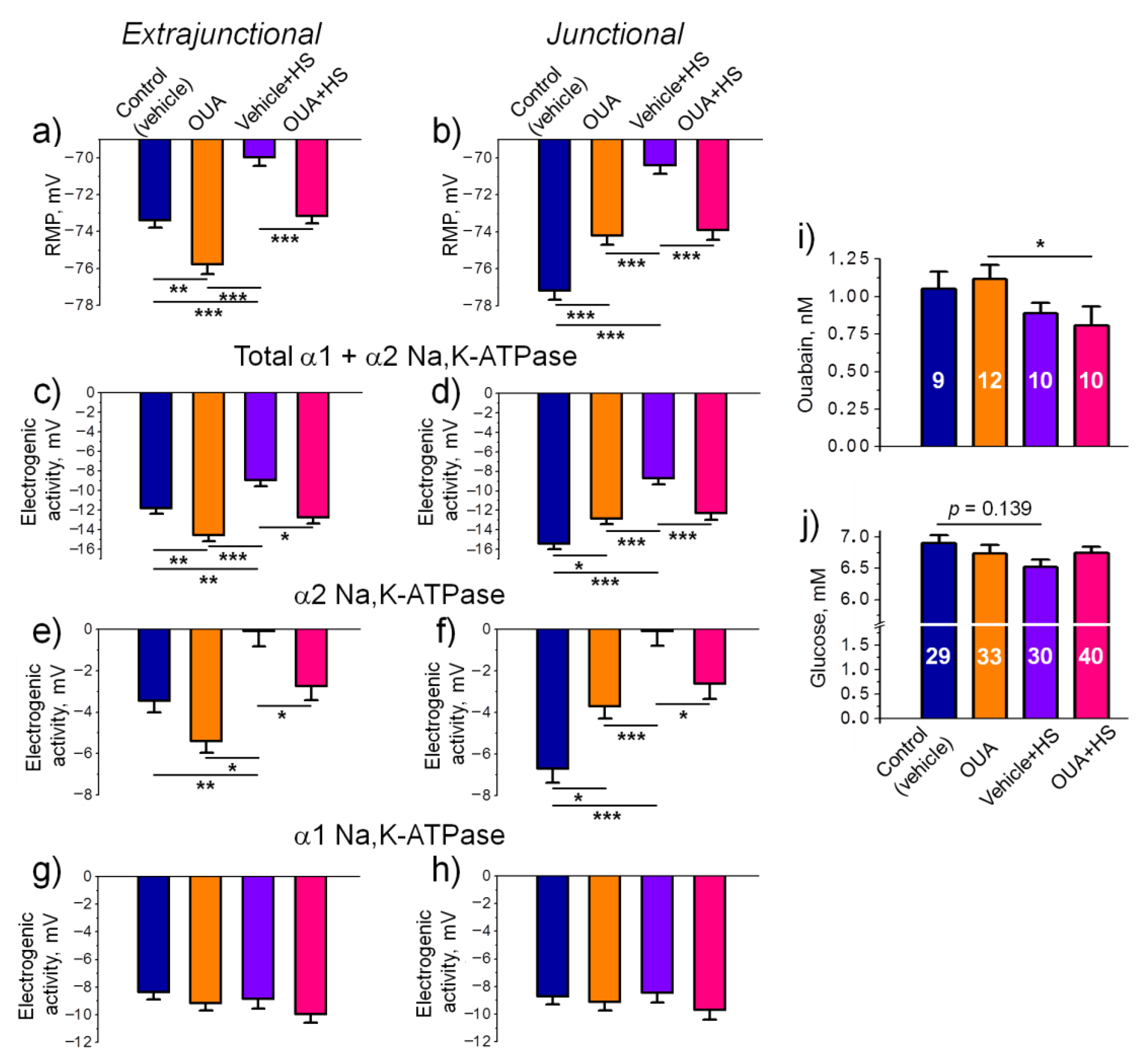
2.3. Chronic Ouabain Modulates the Na,K-ATPase Electrogenic Activity and Prevents the HS-Disturbed Electrogenesis
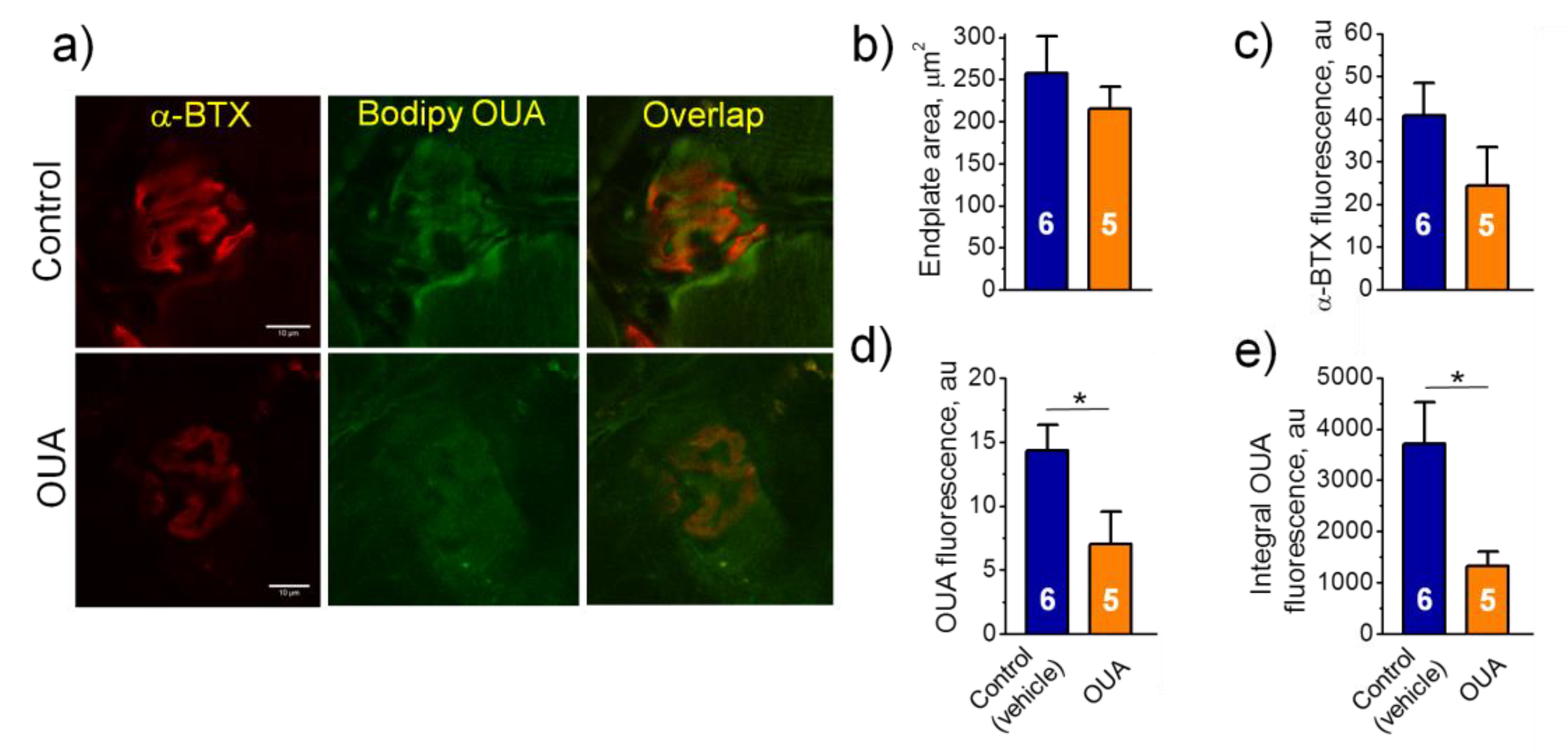
2.4. Chronic Ouabain Reduces the α2 Na,K-ATPase Membrane Localization
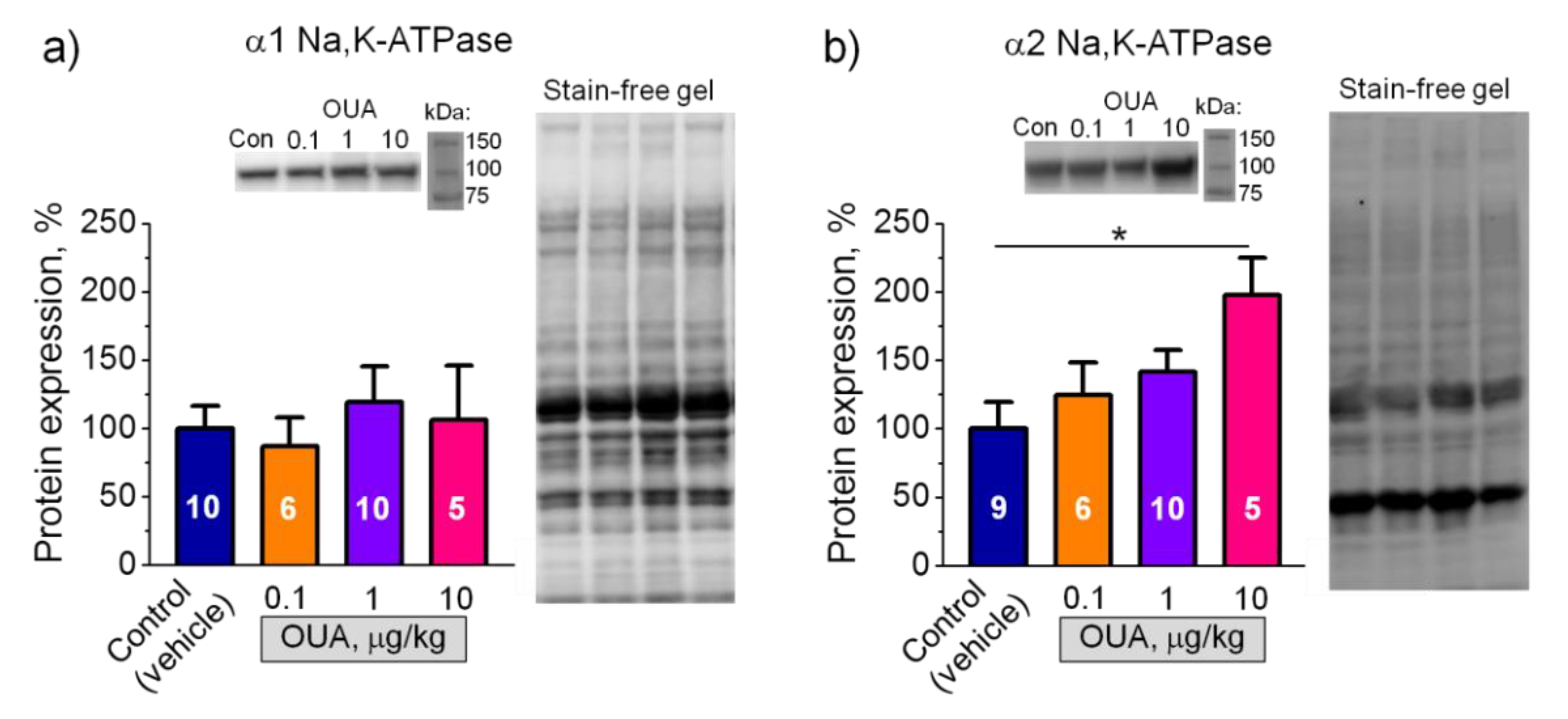
2.5. Chronic Ouabain Modulates Membrane Electrogenesis without Changes in the Na,K-ATPase Protein Content
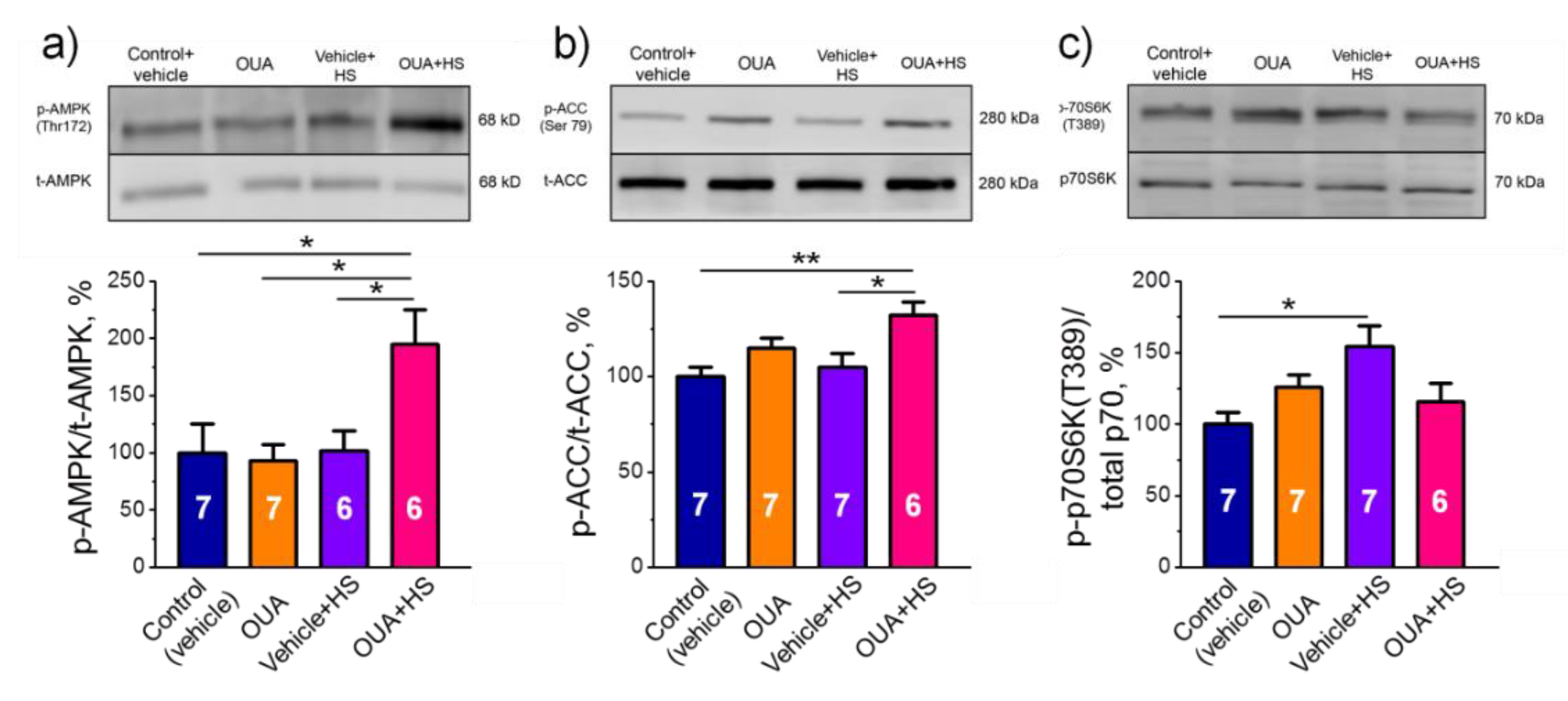
2.6. Chronic Ouabain Pretreatment Modulates the Phosphorylation Level of AMPK and mRNA Expression of IL-6 under Disuse
3. Discussion
- (1)
- Although acute administration of 10 nM ouabain hyperpolarized the plasma membrane ex-vivo, a single injection of ouabain (1 µg/kg) prior the HS is unable to prevent the HS-induced membrane depolarization.
- (2)
- Chronic administration of ouabain (1 µg/kg) alone for four days reduces the α2 Na,K-ATPase membrane localization without changes in the total level of α1 and α2 Na,K-ATPase proteins.
- (3)
- Chronic administration of ouabain diminishes the HS-induced loss of the α2 Na,K-ATPase electrogenic activity and sarcolemma depolarization without changes in the α2 Na,K-ATPase localization in the junctional membrane.
- (4)
- Chronic ouabain treatment increased the phosphorylation levels of AMPK and its substrate ACC accompanied with increased mRNA expression of IL-6 and IL-6R, suggesting the involvement IL-6/AMPK signaling pathways in preventive effects of ouabain.
4. Materials and Methods
4.1. Animals
4.2. Membrane Potential Recording
4.3. Measurement of the Na,K-ATPase Electrogenic Activity in Intact Muscle
4.4. Confocal Microscopy Imaging
4.5. Western Blot Assays
4.6. Cytoplasmic Extracts Preparation and Immunoblots
4.7. RT-qPCR Analysis
4.8. Materials
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sejersted, O.M.; Sjogaard, G. Dynamics and consequences of potassium shifts in skeletal muscle and heart during exercise. Physiol. Rev. 2000, 80, 1411–1481. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T. Na+-K+ pump regulation and skeletal muscle contractility. Physiol. Rev. 2003, 83, 1269–1324. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T. Quantification of Na+, K+ pumps and their transport rate in skeletal muscle: Functional significance. J. Gen. Physiol. 2013, 142, 327–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skou, J.C. The influence of some cations on an adenosine triphosphatase from peripheral nerves. Biochim. Biophys. Acta 1957, 23, 394–401. [Google Scholar] [CrossRef]
- Blanco, G.; Mercer, R.W. Isozymes of the Na-K-ATPase: Heterogeneity in structure, diversity in function. Am. J. Physiol. 1998, 275, F633–F655. [Google Scholar] [CrossRef]
- Pirkmajer, S.; Chibalin, A.V. Na,K-ATPase regulation in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E1–E31. [Google Scholar] [CrossRef] [Green Version]
- Clausen, M.V.; Hilbers, F.; Poulsen, H. The structure and function of the Na, K-ATPase isoforms in health and disease. Front. Physiol. 2017, 8, 371. [Google Scholar] [CrossRef]
- Matchkov, V.V.; Krivoi, I.I. Specialized functional diversity and interactions of the Na, K-ATPase. Front Physiol. 2016, 7, 179. [Google Scholar] [CrossRef] [Green Version]
- Blaustein, M.P.; Hamlyn, J.M. Ouabain, endogenous ouabain and ouabain-like factors: The Na+ pump/ouabain receptor, its linkage to NCX, and its myriad functions. Cell Calcium 2020, 102159. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, J.; Lingrel, J.B. Tissue-Specific and developmental regulation of rat Na, K-ATPase catalytic α isoform and β subunit mRNAs. J. Biol. Chem. 1988, 263, 10436–10442. [Google Scholar] [CrossRef]
- He, S.; Shelly, D.A.; Moseley, A.E.; James, P.F.; James, J.H.; Paul, R.J.; Lingrel, J.B. The α1- and α2-isoforms of Na-K-ATPase play different roles in skeletal muscle contractility. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R917–R925. [Google Scholar] [CrossRef]
- Radzyukevich, T.L.; Neumann, J.C.; Rindler, T.N.; Oshiro, N.; Goldhamer, D.J.; Lingrel, J.B.; Heiny, J.A. Tissue-Specific role of the Na,K-ATPase α2 isozyme in skeletal muscle. J. Biol. Chem. 2013, 288, 1226–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiFranco, M.; Hakimjavadi, H.; Lingrel, J.B.; Heiny, J.A. Na, K-ATPase α2 activity in mammalian skeletal muscle T-tubules is acutely stimulated by extracellular K+. J. Gen. Physiol. 2015, 146, 281–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kravtsova, V.V.; Petrov, A.M.; Matchkov, V.V.; Bouzinova, E.V.; Vasiliev, A.N.; Benziane, B.; Zefirov, A.L.; Chibalin, A.V.; Heiny, J.A.; Krivoi, I.I. Distinct α2 Na,K-ATPase membrane pools are differently involved in early skeletal muscle remodeling during disuse. J. Gen. Physiol. 2016, 147, 175–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutz, L.C.; Mukherji, S.T.; Wang, X.; Bryant, A.; Larre, I.; Heiny, J.A.; Lingrel, J.B.; Pierre, S.V.; Xie, Z. Isoform-Specific role of Na/K-ATPase α1 in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E620–E629. [Google Scholar] [CrossRef]
- Heiny, J.A.; Kravtsova, V.V.; Mandel, F.; Radzyukevich, T.L.; Benziane, B.; Prokofiev, A.V.; Pedersen, S.E.; Chibalin, A.V.; Krivoi, I.I. The nicotinic acetylcholine receptor and the Na,K-ATPase α2 isoform interact to regulate membrane electrogenesis in skeletal muscle. J. Biol. Chem. 2010, 285, 28614–28626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krivoi, I.I.; Petrov, A.M. Cholesterol and the safety factor for neuromuscular transmission. Int. J. Mol. Sci. 2019, 20, 1046. [Google Scholar] [CrossRef] [Green Version]
- Krivoi, I.; Vasiliev, A.; Kravtsova, V.; Dobretsov, M.; Mandel, F. Porcine kidney extract contains factor(s) that inhibit the ouabain-sensitive isoform of Na, K-ATPase (α2) in rat skeletal muscle: A convenient electrophysiological assay. Ann. N. Y. Acad. Sci. 2003, 986, 639–641. [Google Scholar] [CrossRef]
- Radzyukevich, T.L.; Moseley, A.E.; Shelly, D.A.; Redden, G.A.; Behbehani, M.M.; Lingrel, J.B.; Paul, R.J.; Heiny, J.A. The Na, K-ATPase α2 subunit isoform modulates contractility in the perinatal mouse diaphragm. Am. J. Physiol. Cell Physiol. 2004, 287, C1300–C1310. [Google Scholar] [CrossRef] [Green Version]
- Kravtsova, V.V.; Bouzinova, E.V.; Chibalin, A.V.; Matchkov, V.V.; Krivoi, I.I. Isoform-Specific Na, K-ATPase and membrane cholesterol remodeling in the motor endplates in distinct mouse models of myodystrophy. Am. J. Physiol. Cell Physiol. 2020, 318, C1030–C1041. [Google Scholar] [CrossRef]
- Filatov, G.N.; Pinter, M.J.; Rich, M.M. Resting potential-dependent regulation of the voltage sensitivity of sodium channel gating in rat skeletal muscle in vivo. J. Gen. Physiol. 2005, 126, 161–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, M.T.; Cottey, E.; Cottey, A.; Stefanski, C.; Carlson, C.G. Reduced resting potentials in dystrophic (mdx) muscle fibers are secondary to NF-κB-dependent negative modulation of ouabain sensitive Na+-K+ pump activity. J. Neurosci. 2011, 303, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kravtsova, V.V.; Timonina, N.A.; Zakir’yanova, G.F.; Sokolova, A.V.; Mikhailov, V.M.; Zefirov, A.L.; Krivoi, I.I. The structural and functional characteristics of the motor end plates of dysferlin-deficient mice. Neurochem. J. 2018, 12, 305–310. [Google Scholar] [CrossRef]
- Kravtsova, V.V.; Matchkov, V.V.; Bouzinova, E.V.; Vasiliev, A.N.; Razgovorova, I.A.; Heiny, J.A.; Krivoi, I.I. Isoform-Specific Na,K-ATPase alterations precede disuse-induced atrophy of rat soleus muscle. Biomed. Res. Int. 2015, 720172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaustein, M.P. Physiological effects of endogenous ouabain: Control of intracellular Ca2+ stores and cell responsiveness. Am. J. Physiol. Cell Physiol. 1993, 264, C1367–C1387. [Google Scholar] [CrossRef]
- Bagrov, A.Y.; Shapiro, J.I.; Fedorova, O.V. Endogenous cardiotonic steroids: Physiology, pharmacology, and novel therapeutic targets. Pharmacol. Rev. 2009, 61, 9–38. [Google Scholar] [CrossRef]
- Lingrel, J.B. The physiological significance of the cardiotonic steroid/ouabain-binding site of the Na,K-ATPase. Annu. Rev. Physiol. 2010, 72, 395–412. [Google Scholar] [CrossRef] [Green Version]
- Hamlyn, J.M.; Blaustein, M.P.; Bova, S.; DuCharme, D.W.; Harris, D.W.; Mandel, F.; Mathews, W.R.; Ludens, J.H. Identification and characterization of a ouabain-like compound from human plasma. Proc. Natl. Acad. Sci. USA 1991, 88, 6259–6263. [Google Scholar] [CrossRef] [Green Version]
- Bauer, N.; Müller-Ehmsen, J.; Krämer, U.; Hambarchian, N.; Zobel, C.; Schwinger, R.H.; Neu, H.; Kirch, U.; Grünbaum, E.G.; Schoner, W. Ouabain-Like compound changes rapidly on physical exercise in humans and dogs: Effects of β-blockade and angiotensin-converting enzyme inhibition. Hypertension 2005, 45, 1024–1028. [Google Scholar] [CrossRef] [Green Version]
- Radzyukevich, T.L.; Lingrel, J.B.; Heiny, J.A. The cardiac glycoside binding site on the Na,K-ATPase α2 isoform plays a role in the dynamic regulation of active transport in skeletal muscle. Proc. Natl. Acad. Sci. USA 2009, 106, 2565–2570. [Google Scholar] [CrossRef] [Green Version]
- Hamlyn, J.M.; Manunta, P. Endogenous cardiotonic steroids in kidney failure: A review and an hypothesis. Adv. Chronic Kidney Dis. 2015, 22, 232–244. [Google Scholar] [CrossRef] [Green Version]
- Lichtstein, D.; Ilani, A.; Rosen, H.; Horesh, N.; Singh, S.V.; Buzaglo, N.; Hodes, A. Na+, K+-ATPase signaling and bipolar disorder. Int. J. Mol. Sci. 2018, 19, 2314. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Wymore, R.S.; Wang, Y.; Gaudette, G.R.; Krukenkamp, I.B.; Cohen, I.S.; Mathias, R.T. Isoform-Specific stimulation of cardiac Na/K pumps by nanomolar concentrations of glycosides. J. Gen. Physiol. 2002, 119, 297–312. [Google Scholar] [CrossRef]
- Holthouser, K.A.; Mandal, A.; Merchant, M.L.; Schelling, J.R.; Delamere, N.A.; Valdes, R.R., Jr.; Tyagi, S.C.; Lederer, E.D.; Khundmiri, S.J. Ouabain stimulates Na-K-ATPase through a sodium/hydrogen exchanger-1 (NHE-1)-dependent mechanism in human kidney proximal tubule cells. Am. J. Physiol. Ren. Physiol. 2010, 299, F77–F90. [Google Scholar] [CrossRef] [Green Version]
- Ketchem, C.J.; Conner, C.D.; Murray, R.D.; DuPlessis, M.; Lederer, E.D.; Wilkey, D.; Merchant, M.; Khundmiri, S.J. Low dose ouabain stimulates Na-K ATPase α1 subunit association with angiotensin II type 1 receptor in renal proximal tubule cells. Biochim. Biophys. Acta 2016, 1863, 2624–2636. [Google Scholar] [CrossRef] [PubMed]
- Kravtsova, V.V.; Bouzinova, E.V.; Matchkov, V.V.; Krivoi, I.I. Skeletal muscle Na, K-ATPase as a target for circulating ouabain. Int. J. Mol. Sci. 2020, 21, 2875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirkmajer, S.; Bezjak, K.; Matkovic, U.; Dolinar, K.; Jiang, L.Q.; Miš, K.; Gros, K.; Milovanova, K.; Pirkmajer, K.P.; Marš, T.; et al. Ouabain suppresses IL-6/STAT3 signaling and promotes cytokine secretion in cultured skeletal muscle cells. Front. Physiol. 2020, 11, 566584. [Google Scholar] [CrossRef] [PubMed]
- Mijatovic, T.; Van Quaquebeke, E.; Delest, B.; Debeir, O.; Darro, F.; Kiss, R. Cardiotonic steroids on the road to anti-cancer therapy. Biochim. Biophys. Acta 2007, 1776, 32–57. [Google Scholar] [CrossRef] [PubMed]
- Garcia, I.J.P.; Kinoshita, P.F.; Silva, L.N.D.E.; De Souza Busch, M.; Atella, G.C.; Scavone, C.; Cortes, V.F.; Barbosa, L.A.; De Lima Santos, H. Ouabain attenuates oxidative stress and modulates lipid composition in hippocampus of rats in lipopolysaccharide-induced hypocampal neuroinflammation in rats. J. Cell. Biochem. 2019, 120, 4081–4091. [Google Scholar] [CrossRef]
- Dvela-Levitt, M.; Ami, H.C.; Rosen, H.; Shohami, E.; Lichtstein, D. Ouabain improves functional recovery following traumatic brain injury. J. Neurotrauma 2014, 31, 1942–1947. [Google Scholar] [CrossRef] [Green Version]
- Agalakova, N.I.; Kolodkin, N.I.; Adair, C.D.; Trashkov, A.P.; Bagrov, A.Y. Preeclampsia: Cardiotonic steroids, fibrosis, fli1 and hint to carcinogenesis. Int. J. Mol. Sci. 2021, 22, 1941. [Google Scholar] [CrossRef] [PubMed]
- Morey-Holton, E.; Globus, R.K.; Kaplansky, A.; Durnova, G. The hindlimb unloading rat model: Literature overview, technique update and comparison with space flight data. Adv. Space Biol. Med. 2005, 10, 7–40. [Google Scholar] [CrossRef]
- Pirkmajer, S.; Petric, M.; Chibalin, A.V. The role of AMPK in regulation of Na+, K+-ATPase in skeletal muscle: Does the gauge always plug the sink? J. Muscle Res. Cell. Motil. 2021, 42, 77–97. [Google Scholar] [CrossRef] [PubMed]
- Vilchinskaya, N.A.; Mochalova, E.P.; Nemirovskaya, T.L.; Mirzoev, T.M.; Turtikova, O.V.; Shenkman, B.S. Rapid decline in MyHC I(β) mRNA expression in rat soleus during hindlimb unloading is associated with AMPK dephosphorylation. J. Physiol. 2017, 595, 7123–7134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilchinskaya, N.A.; Krivoi, I.I.; Shenkman, B.S. AMP-Activated protein kinase as a key trigger for the disuse-induced skeletal muscle remodeling. Int. J. Mol. Sci. 2018, 19, 3558. [Google Scholar] [CrossRef] [Green Version]
- Chibalin, A.V.; Benziane, B.; Zakyrjanova, G.F.; Kravtsova, V.V.; Krivoi, I.I. Early endplate remodeling and skeletal muscle signaling events following rat hindlimb suspension. J. Cell. Physiol. 2018, 233, 6329–6336. [Google Scholar] [CrossRef]
- Blaustein, M.P.; Chen, L.; Hamlyn, J.M.; Leenen, F.H.; Lingrel, J.B.; Wier, W.G.; Zhang, J. Pivotal role of α2 Na+ pumps and their high affinity ouabain binding site in cardiovascular health and disease. J. Physiol. 2016, 594, 6079–6103. [Google Scholar] [CrossRef] [Green Version]
- Kotova, O.; Al-Khalili, L.; Talia, S.; Hooke, C.; Fedorova, O.V.; Bagrov, A.Y.; Chibalin, A.V. Cardiotonic steroids stimulate glycogen synthesis in human skeletal muscle cells via a Src-and ERK1/2-dependent mechanism. J. Biol. Chem. 2006, 281, 20085–20094. [Google Scholar] [CrossRef] [Green Version]
- Bodine, S.C.; Baehr, L.M. Skeletal muscle atrophy and the E3 ubiquitin ligases MuRF-1 and MAFbx/Atrogin-1. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E469–E484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenkman, B.S. How postural muscle senses disuse? Early signs and signals. Int. J. Mol. Sci. 2020, 21, 5037. [Google Scholar] [CrossRef]
- Gorza, L.; Sorge, M.; Seclì, L.; Brancaccio, M. Master regulators of muscle atrophy: Role of costamere components. Cells 2021, 10, 61. [Google Scholar] [CrossRef]
- Hardie, D.G.; Schaffer, B.E.; Brunet, A. AMPK: An energy-sensing pathway with multiple inputs and outputs. Trends Cell. Biol. 2016, 26, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Dial, A.G.; Ng, S.Y.; Manta, A.; Ljubicic, V. The role of AMPK in neuromuscular biology and disease. Trends Endocrinol. Metab. 2018, 29, 300–312. [Google Scholar] [CrossRef]
- Cerveró, C.; Montull, N.; Tarabal, O.; Piedrafita, L.; Esquerda, J.E.; Calderó, J. Chronic treatment with the AMPK agonist AICAR prevents skeletal muscle pathology but fails to improve clinical outcome in a mouse model of severe spinal muscular atrophy. Neurotherapeutics 2016, 13, 198–216. [Google Scholar] [CrossRef]
- Benziane, B.; Björnholm, M.; Pirkmajer, S.; Austin, R.L.; Kotova, O.; Viollet, B.; Zierath, J.R.; Chibalin, A.V. Activation of AMP-activated protein kinase stimulates Na+,K+-ATPase activity in skeletal muscle cells. J. Biol. Chem. 2012, 287, 23451–23463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kravtsova, V.V.; Vilchinskaya, N.A.; Rozlomii, V.L.; Shenkman, B.S.; Krivoi, I.I. Low ouabain doses and AMP-activated protein kinase as factors supporting electrogenesis in skeletal muscle. Biochemistry 2019, 84, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Ruderman, N.B.; Keller, C.; Richard, A.M.; Saha, A.K.; Luo, Z.; Xiang, X.; Giralt, M.; Ritov, V.B.; Menshikova, E.V.; Kelley, D.E.; et al. Interleukin-6 regulation of AMP-activated protein kinase. Potential role in the systemic response to exercise and prevention of the metabolic syndrome. Diabetes 2006, 55, S48–S54. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, B.K.; Febbraio, M.A. Muscle as an endocrine organ: Focus on muscle-derived interleukin-6. Physiol. Rev. 2008, 88, 1379–1406. [Google Scholar] [CrossRef] [Green Version]
- Mirzoev, T.; Tyganov, S.; Vilchinskaya, N.; Lomonosova, Y.; Shenkman, B. Key markers of mTORC1-dependent and mTORC1-independent signaling pathways regulating protein synthesis in rat soleus muscle during early stages of hindlimb unloading. Cell. Physiol. Biochem. 2016, 39, 1011–1020. [Google Scholar] [CrossRef]
- Belova, S.P.; Vilchinskaya, N.A.; Mochalova, E.P.; Mirzoev, T.M.; Nemirovskaya, T.L.; Shenkman, B.S. Elevated p70S6K phosphorylation in rat soleus during the early stage of unloading: Causes and consequences. Arch. Biochem. Biophys. 2019, 674, 108105. [Google Scholar] [CrossRef]
- Selden, R.; Smith, T.W. Ouabain pharmacokinetics in dog and man. Determination by radioimmunoassay. Circulation 1972, 45, 1176–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matyushkin, D.P.; Krivoi, I.I.; Drabkina, T.M. Synaptic feed-backs mediated by potassium ions. Gen. Physiol. Biophys. 1995, 14, 369–381. [Google Scholar]
- Kuntzweiler, T.A.; Arguello, J.M.; Lingrel, J.B. Asp804 and Asp808 in the transmembrane domain of the Na, K-ATPase α subunit are cation coordinating residues. J. Biol. Chem. 1996, 271, 29682–29687. [Google Scholar] [CrossRef] [Green Version]
- Cheek, D.; Holt, A.; Hill, D.; Talbert, J.L. Skeletal muscle cell mass and growth: The concept of the deoxyribonucleic acid unit. Pediatr. Res. 1971, 5, 312–328. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, R.B.; Phelps, R.O. Muscle fiber type composition of the rat hindlimb. Am. J. Anat. 1984, 171, 259–272. [Google Scholar] [CrossRef]
- Lawler, J.M.; Song, W.; Demaree, S.R. Hindlimb unloading increases oxidative stress and disrupts antioxidant capacity in skeletal muscle. Free Radic. Biol. Med. 2003, 35, 9–16. [Google Scholar] [CrossRef]
- Nuoc, T.N.; Kim, S.; Ahn, S.H.; Lee, J.S.; Park, B.J.; Lee, T.H. The analysis of antioxidant expression during muscle atrophy induced by hindlimb suspension in mice. J. Physiol. Sci. 2017, 67, 121–129. [Google Scholar] [CrossRef]
- Bogdanova, A.; Petrushanko, I.Y.; Hernansanz-Agustín, P.; Martínez-Ruiz, A. “Oxygen sensing” by Na,K-ATPase: These miraculous thiols. Front. Physiol. 2016, 7, 314. [Google Scholar] [CrossRef]
- Bolotta, A.; Visconti, P.; Fedrizzi, G.; Ghezzo, A.; Marini, M.; Manunta, P.; Messaggio, E.; Posar, A.; Vignini, A.; Abruzzo, P.M. Na+, K+-ATPase activity in children with autism spectrum disorder: Searching for the reason(s) of its decrease in blood cells. Autism Res. 2018, 11, 1388–1403. [Google Scholar] [CrossRef]
- Juel, C.; Hostrup, M.; Bangsbo, J. The effect of exercise and beta2-adrenergic stimulation on glutathionylation and function of the Na,K-ATPase in human skeletal muscle. Physiol. Rep. 2015, 3, e12515. [Google Scholar] [CrossRef]
- Cavalcante-Silva, L.H.A.; Lima, É.A.; Carvalho, D.C.M.; Sales-Neto, J.M.; Alves, A.K.A.; Galvão, J.G.F.M.; Silva, J.S.F.; Rodrigues-Mascarenhas, S. Much more than a cardiotonic steroid: Modulation of inflammation by ouabain. Front. Physiol. 2017, 8, 895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, P.F.; Yshii, L.M.; Orellana, A.M.M.; Paixão, A.G.; Vasconcelos, A.R.; Lima, L.S.; Kawamoto, E.M.; Scavone, C. α2 Na+, K+-ATPase silencing induces loss of inflammatory response and ouabain protection in glial cells. Sci. Rep. 2017, 7, 4894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011; pp. 1–246.
- Krivoi, I.I.; Drabkina, T.M.; Kravtsova, V.V.; Vasiliev, A.N.; Eaton, M.J.; Skatchkov, S.N.; Mandel, F. On the functional interaction between nicotinic acetylcholine receptor and Na+,K+-ATPase. Pflug. Arch. 2006, 452, 756–765. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Description | Forward Primer Reverse Primer | GeneBank |
|---|---|---|
| Rpl 19 | 5′-GTACCCTTCCTCTTCCCTATGC-3′ 5′-CAATGCCAACTCTCGTCAACAG-3′ | NM_031103.1 |
| Interleukin-6 | 5′-CCGGAGAGGAGACTTCACAG-3′ 5′-ACAGTGCATCATCGCTGTTC-3′ | NM_012589.2 |
| Interleukin-6 receptor | 5′-TCACAGAGCAGAGAATGGACT-3′ 5′-GTATGGCTGATACCACAAGGT-3′ | NM_017020.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kravtsova, V.V.; Paramonova, I.I.; Vilchinskaya, N.A.; Tishkova, M.V.; Matchkov, V.V.; Shenkman, B.S.; Krivoi, I.I. Chronic Ouabain Prevents Na,K-ATPase Dysfunction and Targets AMPK and IL-6 in Disused Rat Soleus Muscle. Int. J. Mol. Sci. 2021, 22, 3920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083920
Kravtsova VV, Paramonova II, Vilchinskaya NA, Tishkova MV, Matchkov VV, Shenkman BS, Krivoi II. Chronic Ouabain Prevents Na,K-ATPase Dysfunction and Targets AMPK and IL-6 in Disused Rat Soleus Muscle. International Journal of Molecular Sciences. 2021; 22(8):3920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083920
Chicago/Turabian StyleKravtsova, Violetta V., Inna I. Paramonova, Natalia A. Vilchinskaya, Maria V. Tishkova, Vladimir V. Matchkov, Boris S. Shenkman, and Igor I. Krivoi. 2021. "Chronic Ouabain Prevents Na,K-ATPase Dysfunction and Targets AMPK and IL-6 in Disused Rat Soleus Muscle" International Journal of Molecular Sciences 22, no. 8: 3920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083920