
Metabolic, Organoleptic and Transcriptomic Impact of Saccharomyces cerevisiae Genes Involved in the Biosynthesis of Linear and Substituted Esters

 , and
, and
Abstract
:1. Introduction
2. Results
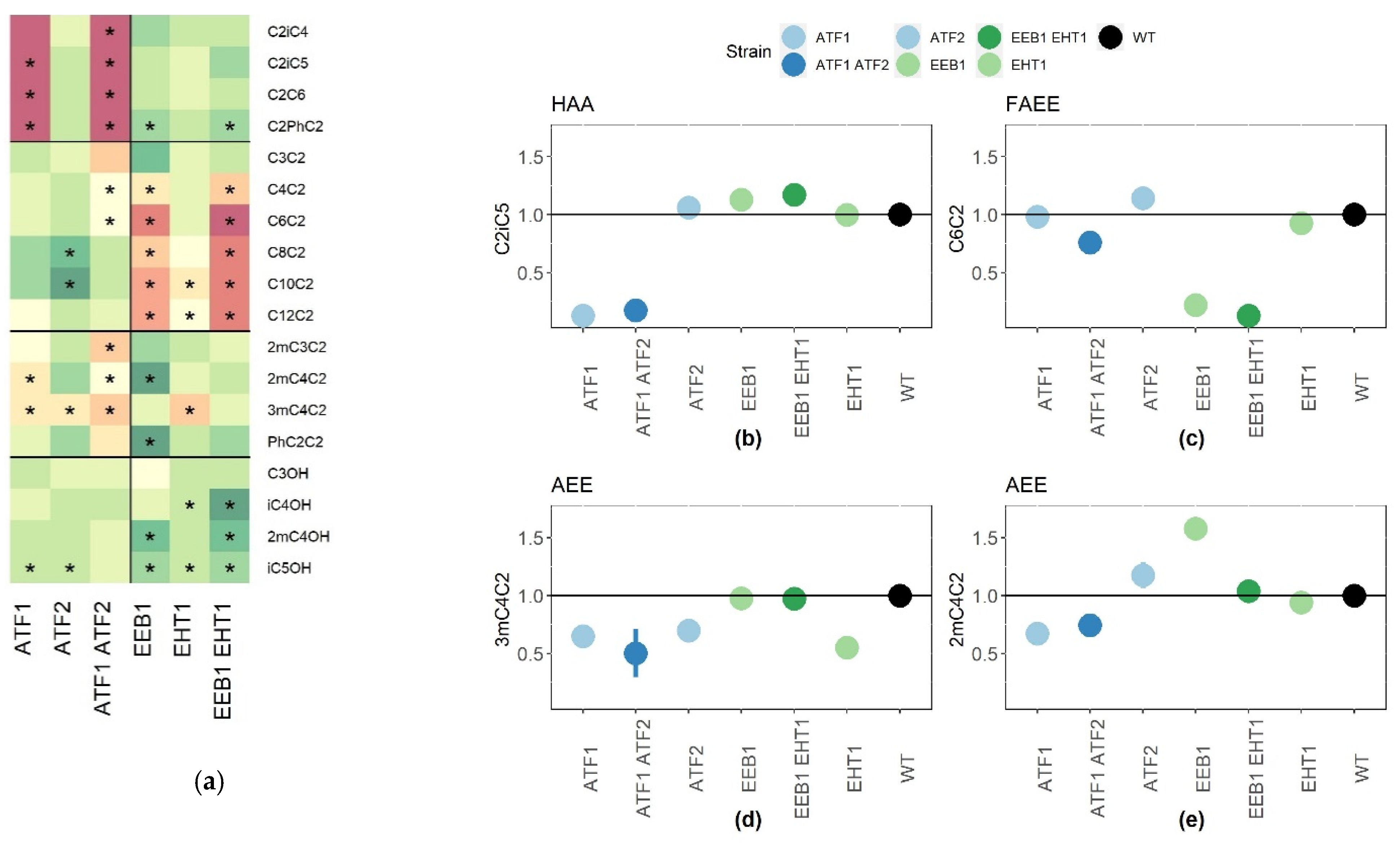
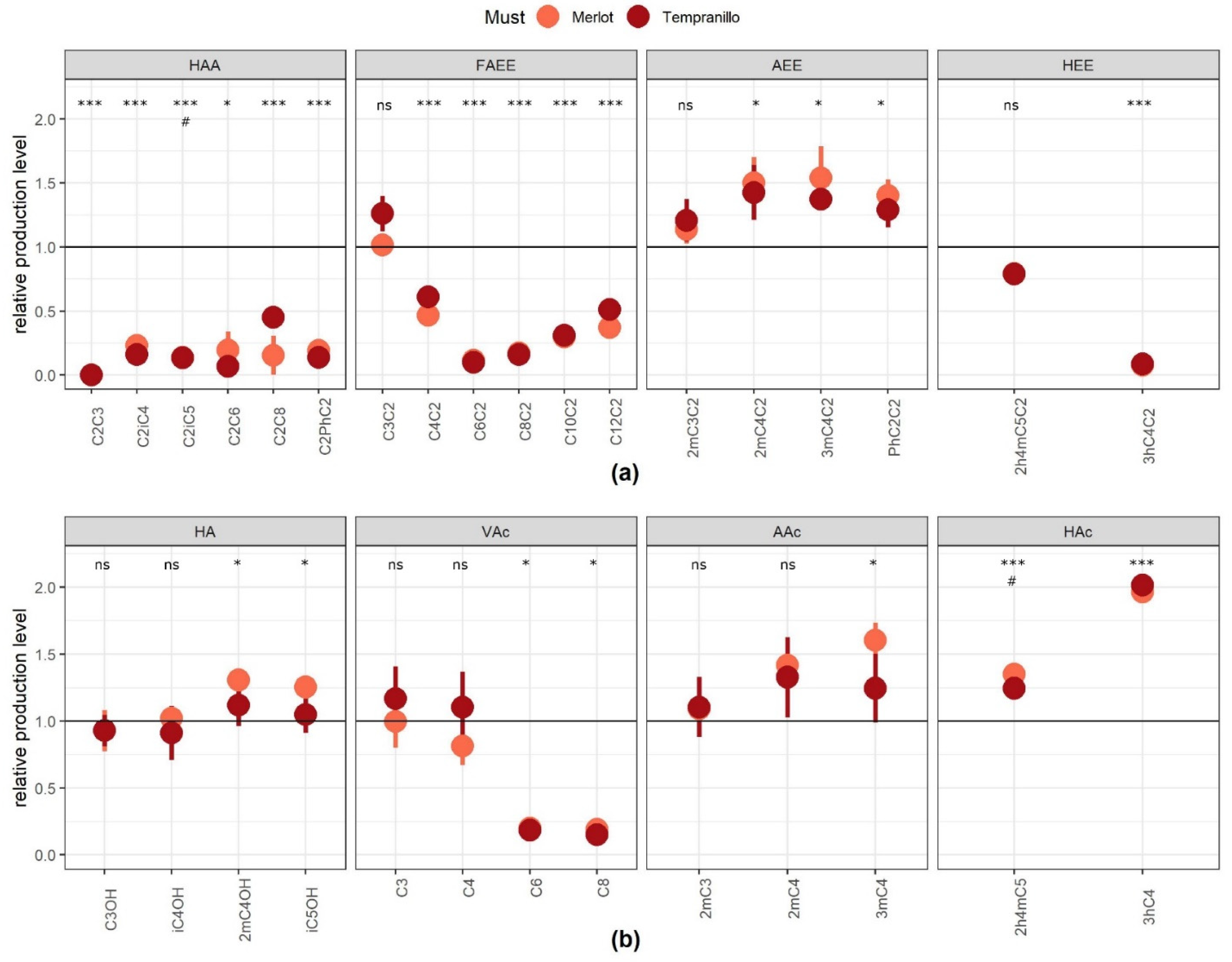
2.1. Validation of the Role of AATase and AEATase in a Red Wine Fermentation
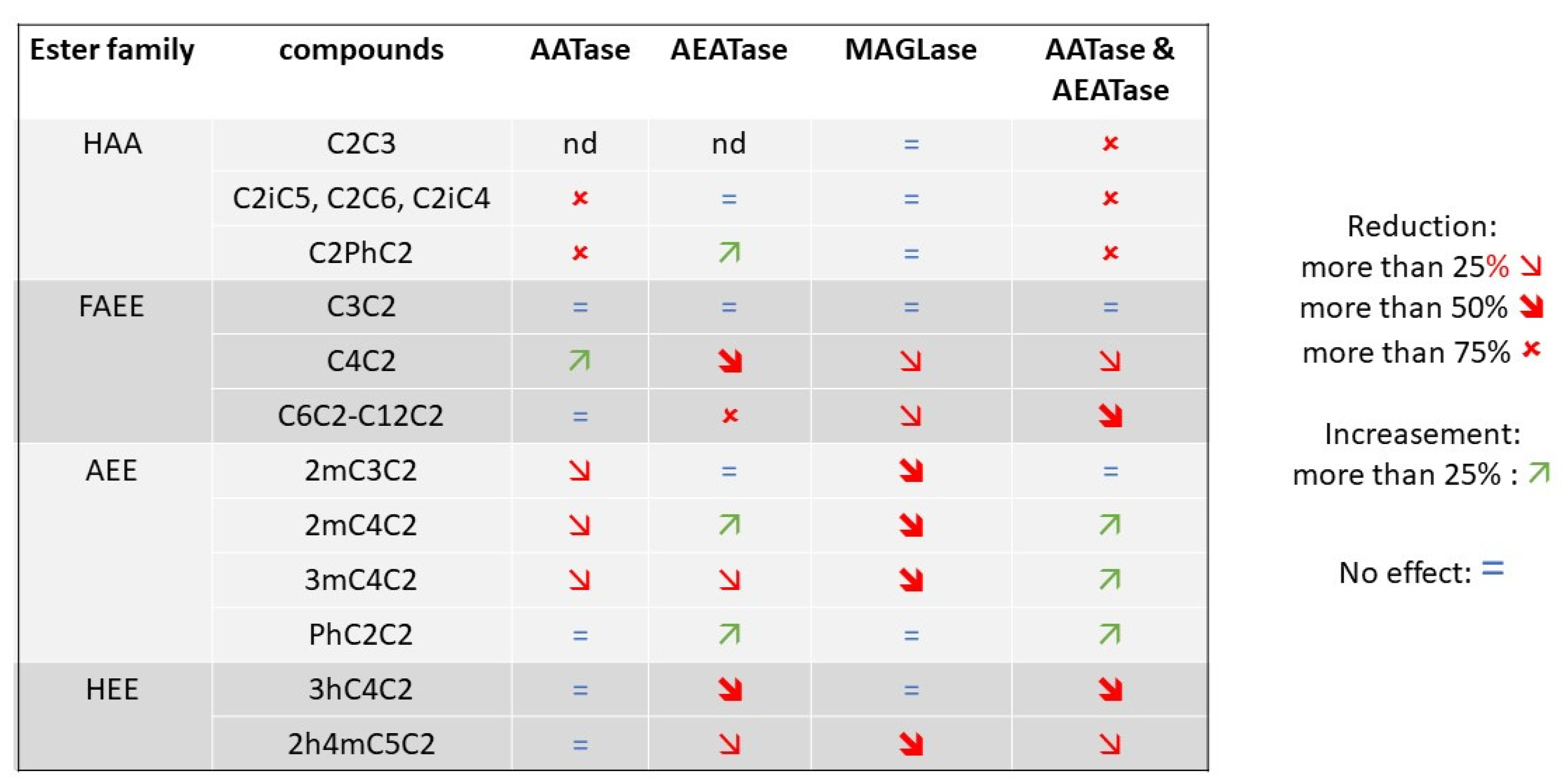
2.2. Functional Characterization of a Nearly-Esterase-Free Strain
2.3. Sensory Profiling of a Wine with Reduced Ester Content
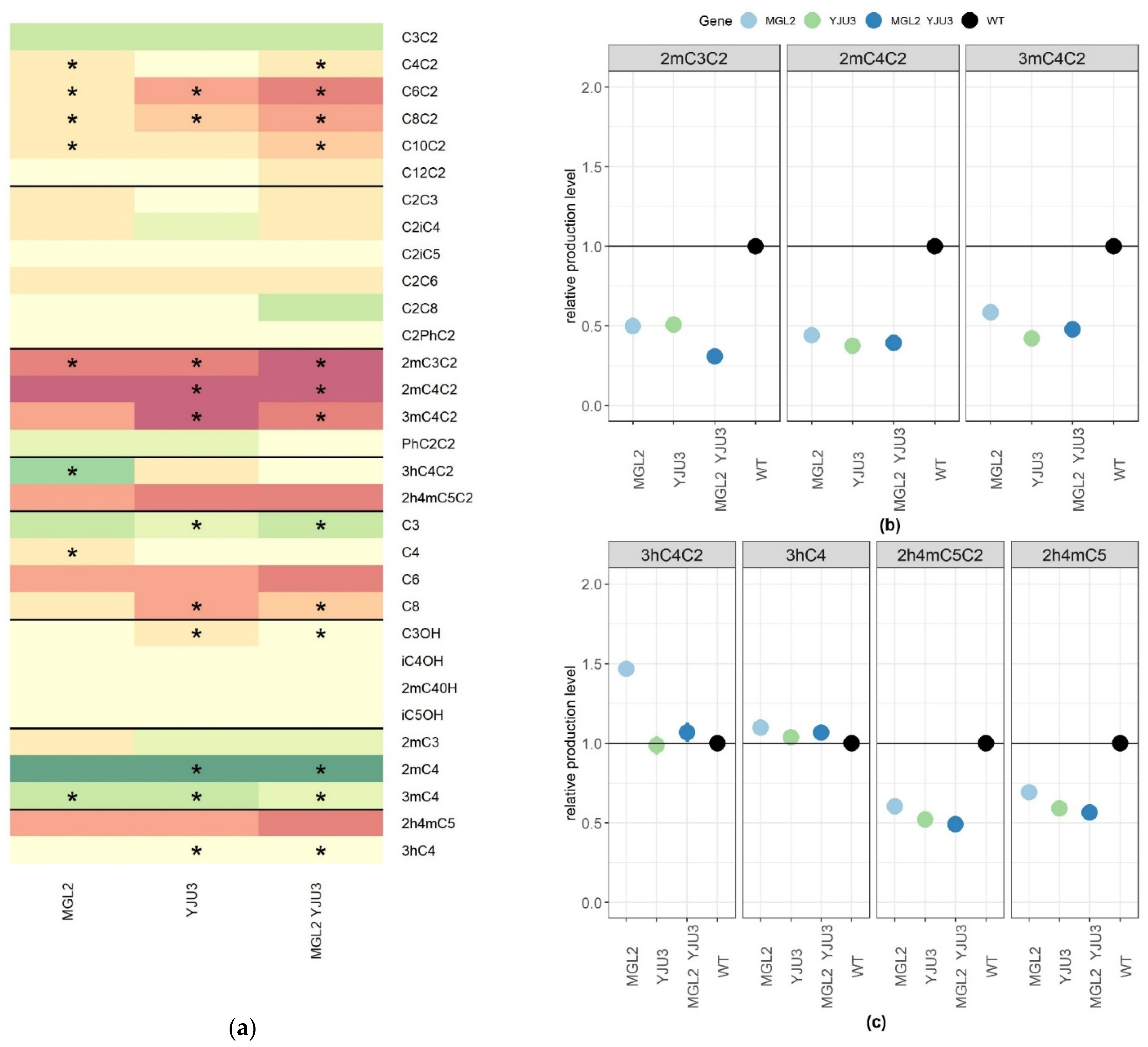
2.4. Functional Characterization of Mgl2p and Yju3p, Two Mono-Acyl Glycerol Lipases Involved in the Synthesis of Substituted Esters
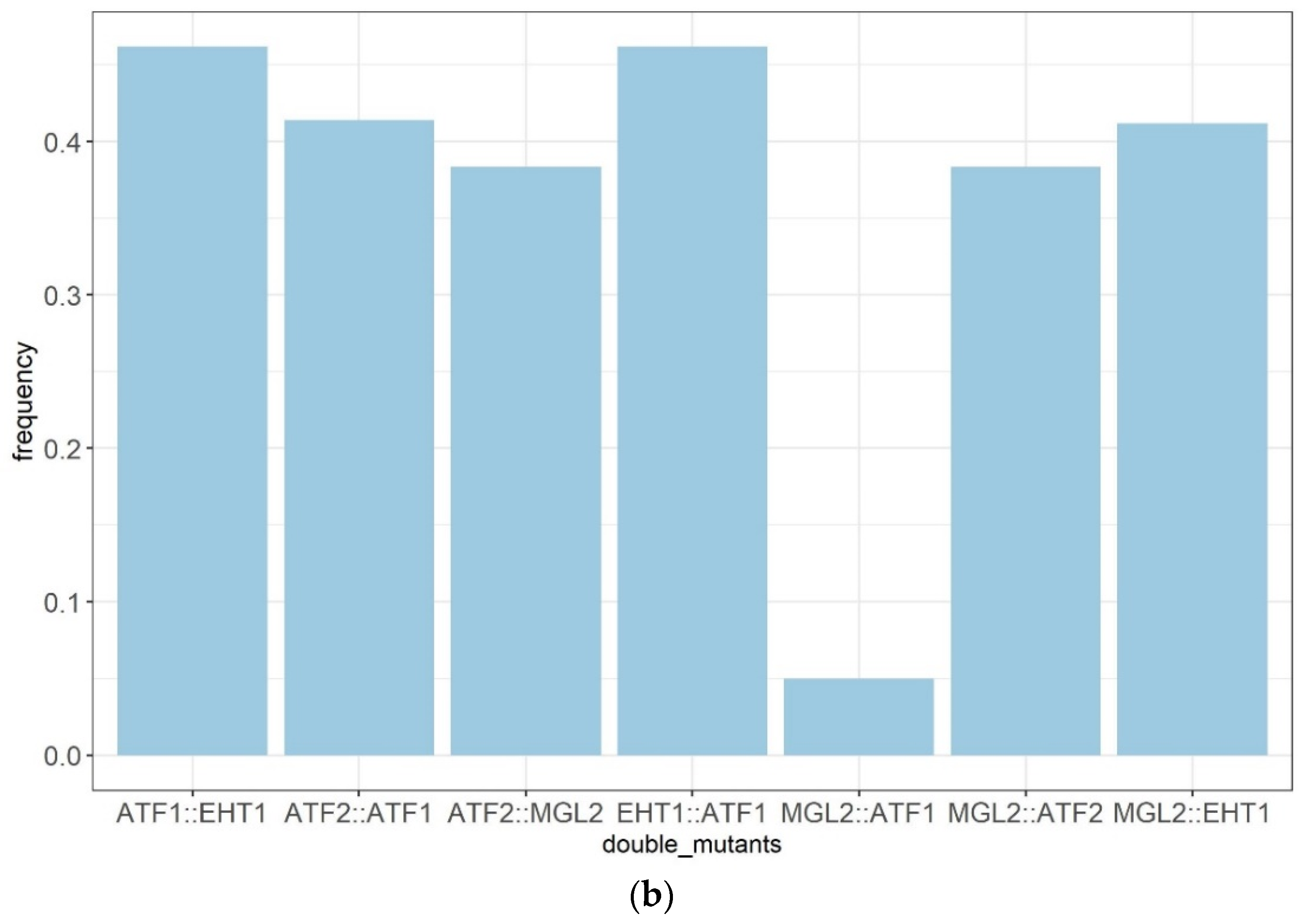
2.5. Attempts to Construct a Fully Esterase-Free Yeast Strain
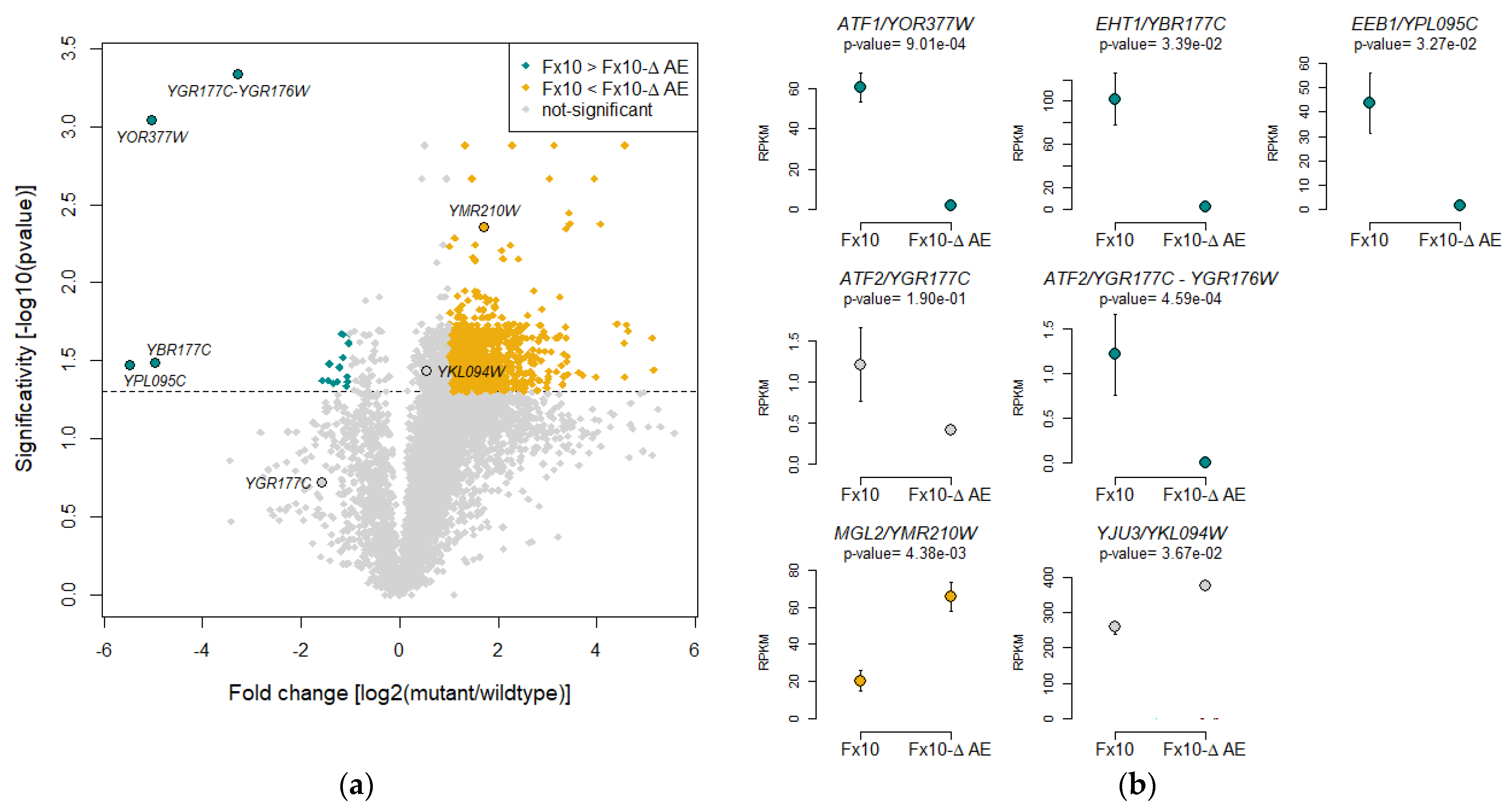
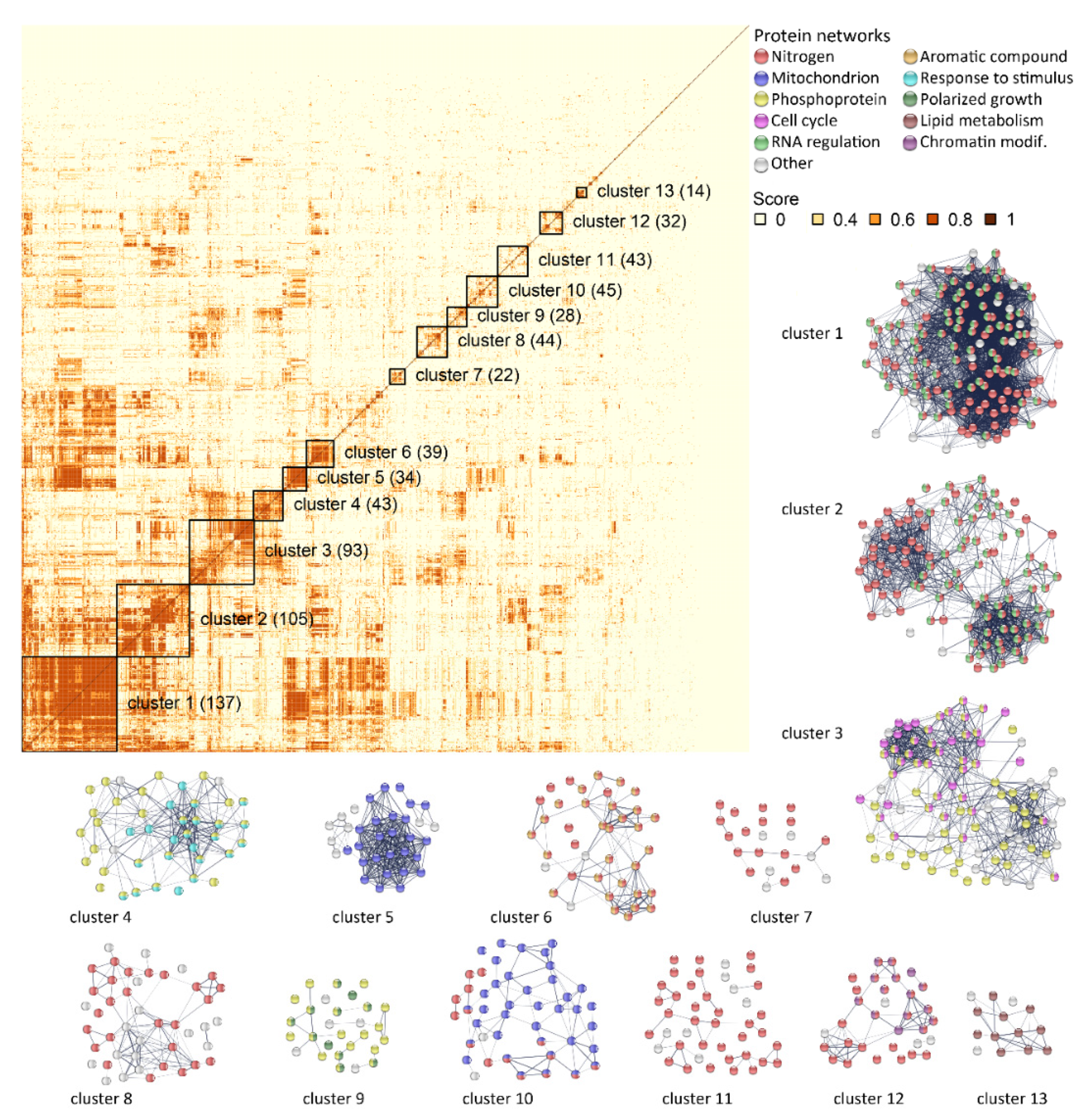
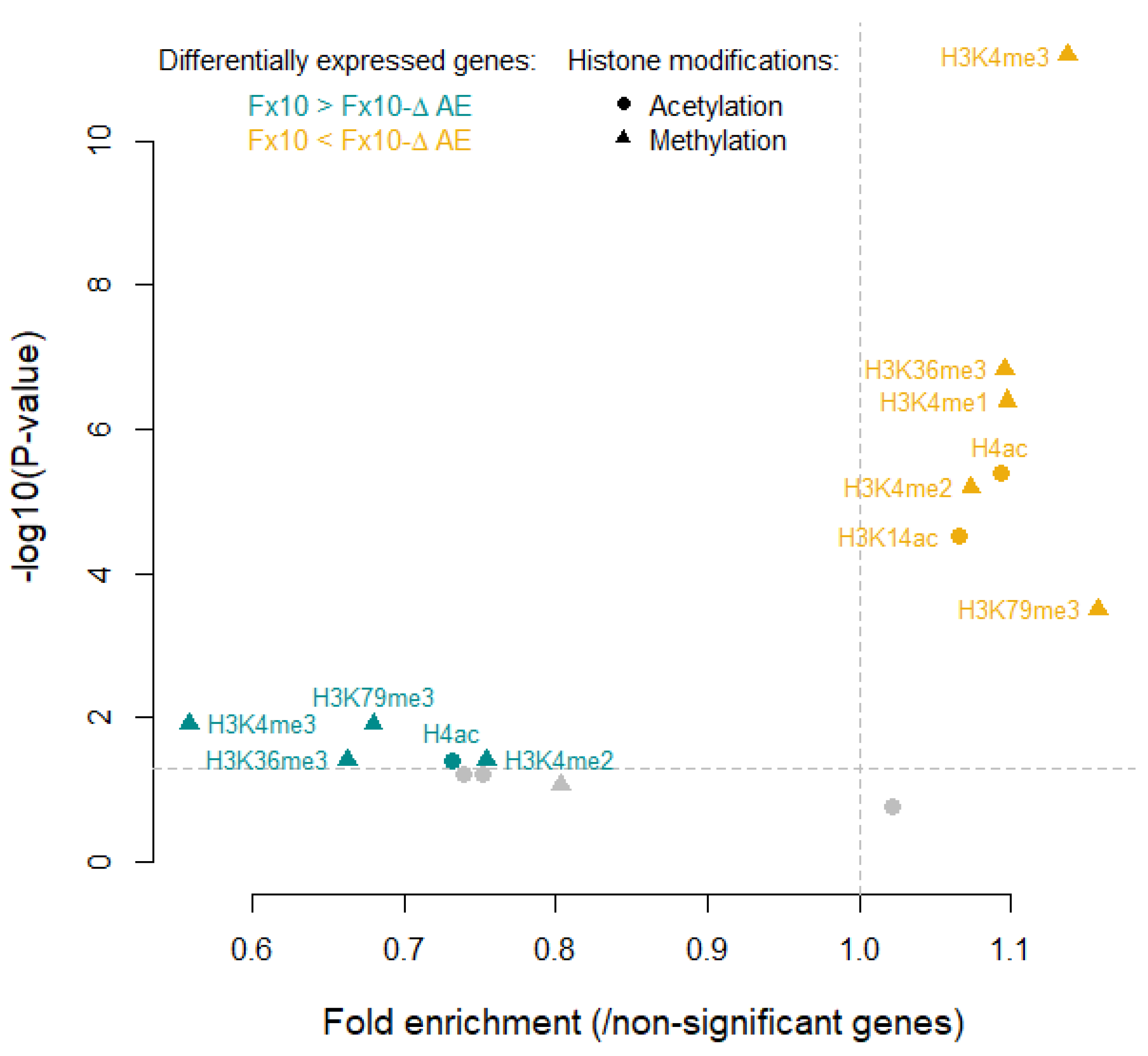
2.6. Transcriptomic Analysis of the Fx10-ΔAE Strain Reveals Unsuspected Consequences of the Depletion of Esterase Activities
3. Discussion
3.1. General Reassessment of the Esterase Contribution in Red Winemaking
3.2. Genetic and Transcriptomic Experiments Revealed Unsuspected Physiological Consequences of Esterase Activity
4. Materials and Methods
4.1. Culture Conditions and Classical Genetics Manipulations
4.2. Construction of Esterase-Free Yeast Strains
4.3. Fermentation Conditions
4.4. Total RNA Isolation and mRNA Sequencing
4.5. RNA-Seq Alignment and Quantification
4.6. RNA-Seq Statistical Analyses
4.7. Chemical Analyses
4.7.1. Apolar Ester Analysis
4.7.2. Quantification of Volatile Acids and Hydroxylated Esters by Liquid–Liquid Extraction and GC/MS Analysis
4.7.3. Quantification of Hydroxylated Acids
4.7.4. Higher Alcohol Analyses
4.8. Sensory Analyses
4.8.1. General Conditions
4.8.2. Sensory Panels
4.8.3. Discriminative Tests
4.8.4. Descriptive Testing Methods
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sicard, D.; Legras, J.L. Bread, beer and wine: Yeast domestication in the Saccharomyces sensu stricto complex. Comptes Rendus–Biol. 2011, 334, 229–236. [Google Scholar] [CrossRef]
- Pisarnitskii, A.F. Formation of wine aroma: Tones and imperfections caused by minor components. Appl. Biochem. Microbiol. 2001, 37, 552–560. [Google Scholar] [CrossRef]
- Cordente, A.G.; Curtin, C.D.; Varela, C.; Pretorius, I.S. Flavour-active wine yeasts. Appl. Microbiol. Biotechnol. 2012, 96, 601–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, S.; Derdelinckx, G.; Dufour, J.-P. Relationships between the chemical composition and sensory evaluation of lager beers. Food Qual. Prefer. 1994, 5, 145–149. [Google Scholar] [CrossRef]
- Gammacurta, M.; Marchand, S.; Albertin, W.; Moine, V.; De Revel, G. Impact of yeast strain on ester levels and fruity aroma persistence during aging of bordeaux red wines. J. Agric. Food Chem. 2014, 62, 5378–5389. [Google Scholar] [CrossRef] [PubMed]
- Tempère, S.; Marchal, A.; Barbe, J.-C.; Bely, M.; Masneuf-Pomarede, I.; Marullo, P.; Albertin, W. The complexity of wine: Clarifying the role of microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 3995–4007. [Google Scholar] [CrossRef] [PubMed]
- Hirst, M.B.; Richter, C.L. Review of aroma formation through metabolic pathways of Saccharomyces cerevisia in beverage fermentations. Am. J. Enol. Vitic. 2016, 67, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Trindade de Carvalho, B.; Holt, S.; Souffriau, B.; Lopes Brandão, R.; Foulquié-Moreno, M.R.; Thevelein, J.M. Identification of novel alleles conferring superior production of rose flavor phenylethyl acetate using polygenic analysis in yeast. MBio 2017, 8, e01173-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steyer, D.; Ambroset, C.; Brion, C.; Claudel, P.; Delobel, P.; Sanchez, I.; Erny, C.; Blondin, B.; Karst, F.; Legras, J.-L. QTL mapping of the production of wine aroma compounds by yeast. BMC Genom. 2012, 13, 573. [Google Scholar] [CrossRef] [Green Version]
- Eder, M.; Sanchez, I.; Brice, C.; Camarasa, C.; Legras, J.-L.; Dequin, S. QTL mapping of volatile compound production in Saccharomyces cerevisiae during alcoholic fermentation. BMC Genom. 2018, 19, 166. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazelwood, L.A.; Daran, J.M.; van Maris, A.J.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 3920. [Google Scholar] [CrossRef] [Green Version]
- Procopio, S.; Qian, F.; Becker, T. Function and regulation of yeast genes involved in higher alcohol and ester metabolism during beverage fermentation. Eur. Food Res. Technol. 2011, 233, 721–729. [Google Scholar] [CrossRef]
- Dickinson, J.R.; Salgado, L.E.J.; Hewlins, M.J.E. The catabolism of amino acids to long chain and complex alcohols in Saccharomyces cerevisiae. J. Biol. Chem. 2003, 278, 8028–8034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshioka, K.; Hashimoto, N. Ester formation by alcohol acetyltransferase from brewers’ yeast. Agric. Biol. Chem. 1981, 45, 2183–2190. [Google Scholar] [CrossRef]
- Mason, A.B.; Dufour, J. Alcohol acetyltransferases and the significance of ester synthesis in yeast. Yeast 2000, 16, 1287–1298. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Van Laere, S.D.M.; Vanderhaegen, B.M.P.; Derdelinckx, G.; Dufour, J.; Pretorius, I.S.; Winderickx, J.; Thevelein, J.M.; Delvaux, F.R. Expression levels of the yeast alcohol acetyltransferase genes. Society 2003, 69, 5228–5237. [Google Scholar] [CrossRef]
- Lilly, M.; Lambrechts, M.G.; Pretorius, I.S. Effect of increased yeast alcohol acetyltransferase activity on flavor profiles of wine and distillates. Appl. Environ. Microbiol. 2000, 66, 744–753. [Google Scholar] [CrossRef] [Green Version]
- Kruis, A.J.; Gallone, B.; Jonker, T.; Mars, A.E.; van Rijswijck, I.M.H.; Wolkers-Rooijackers, J.C.M.; Smid, E.J.; Steensels, J.; Verstrepen, K.J.; Kengen, S.W.; et al. Contribution of Eat1 and other alcohol acyltransferases to ester production in Saccharomyces cerevisiae. Front. Microbiol. 2018, 9, 3202. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Verstrepen, K.J.; Van Laere, S.D.M.; Voet, A.R.D.; Van Dijck, P.; Delvaux, F.R.; Thevelein, J.M. The Saccharomyces cerevisiae EHT1 and EEB1 genes encode novel enzymes with medium-chain fatty acid ethyl ester synthesis and hydrolysis capacity. J. Biol. Chem. 2006, 281, 4446–4456. [Google Scholar] [CrossRef] [Green Version]
- Knight, M.J.; Bull, I.; Curnow, P. The yeast enzyme Eht1 is an octanoyl-CoA:ethanol acyltransferase that also functions as a thioesterase. Yeast 2014, 2–5. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Maroto, M.C.; Schneider, R.; Baumes, R. Formation pathways of ethyl esters of branched short-chain fatty acids during wine aging. J. Agric. Food Chem. 2005, 53, 3503–3509. [Google Scholar] [CrossRef] [PubMed]
- Matheis, K.; Granvogl, M.; Schieberle, P. Quantitation and enantiomeric ratios of aroma compounds formed by an Ehrlich degradation of l-isoleucine in fermented foods. J. Agric. Food Chem. 2016, 64, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Lytra, G.; Franc, C.; Cameleyre, M.; Barbe, J.-C. Study of substituted ester formation in red wine by the development of a new method for quantitative determination and enantiomeric separation of their corresponding acids. J. Agric. Food Chem. 2017, 65, 5018–5025. [Google Scholar] [CrossRef] [PubMed]
- Selvaraju, K.; Gowsalya, R.; Vijayakumar, R.; Nachiappan, V. MGL2/YMR210w encodes a monoacylglycerol lipase in Saccharomyces cerevisiae. FEBS Lett. 2016, 590, 1174–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heier, C.; Taschler, U.; Rengachari, S.; Oberer, M.; Wolinski, H.; Natter, K.; Kohlwein, S.D.; Leber, R.; Zimmermann, R. Identification of Yju3p as functional orthologue of mammalian monoglyceride lipase in the yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta 2010, 1801, 1063–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokholok, D.K.; Harbison, C.T.; Levine, S.; Cole, M.; Hannett, N.M.; Tong, I.L.; Bell, G.W.; Walker, K.; Rolfe, P.A.; Herbolsheimer, E.; et al. Genome-wide map of nucleosome acetylation and methylation in yeast. Cell 2005, 122, 517–527. [Google Scholar] [CrossRef] [Green Version]
- Rollero, S.; Bloem, A.; Camarasa, C.; Sanchez, I.; Ortiz-Julien, A.; Sablayrolles, J.-M.; Dequin, S.; Mouret, J.-R. Combined effects of nutrients and temperature on the production of fermentative aromas by Saccharomyces cerevisiae during wine fermentation. Appl. Microbiol. Biotechnol. 2014. [Google Scholar] [CrossRef]
- Seguinot, P.; Bloem, A.; Brial, P.; Meudec, E.; Ortiz-Julien, A.; Camarasa, C. Analysing the impact of the nature of the nitrogen source on the formation of volatile compounds to unravel the aroma metabolism of two non-Saccharomyces strains. Int. J. Food Microbiol. 2020, 316, 108441. [Google Scholar] [CrossRef]
- Lytra, G.; Tempere, S.; de Revel, G.; Barbe, J.-C. Impact of Perceptive Interactions on Red Wine Fruity Aroma. J. Agric. Food Chem. 2012, 60, 12260–12269. [Google Scholar] [CrossRef]
- Smit, B.A.; Engels, W.J.M.; Smit, G. Branched chain aldehydes: Production and breakdown pathways and relevance for flavour in foods. Appl. Microbiol. Biotechnol. 2008, 81, 987–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lytra, G.; Cameleyre, M.; Tempere, S.; Barbe, J.-C. Distribution and organoleptic impact of ethyl 3-hydroxybutanoate enantiomers in wine. J. Agric. Food Chem. 2015, 63, 10484–10491. [Google Scholar] [CrossRef]
- Galdieri, L.; Zhang, T.; Rogerson, D.; Lleshi, R.; Vancura, A. Protein acetylation and acetyl coenzyme a metabolism in budding yeast. Eukaryot. Cell 2014, 13, 1472–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchal, A.; Marullo, P.; Moine, V.; Dubourdieu, D. Influence of yeast macromolecules on sweetness in dry wines: Role of the Saccharomyces cerevisiae protein Hsp12. J. Agric. Food Chem. 2011, 59, 2004–2010. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Marullo, P.; Bely, M.; Masneuf-Pomarède, I.; Pons, M.; Aigle, M.; Dubourdieu, D.; Masneuf-Pomarede, I.; Pons, M.; Aigle, M.; Dubourdieu, D. Breeding strategies for combining fermentative qualities and reducing off-flavor production in a wine yeast model. FEMS Yeast Res. 2006, 6, 268–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marullo, P.; Mansour, C.; Dufour, M.; Albertin, W.; Sicard, D.; Bely, M.; Dubourdieu, D. Genetic improvement of thermo-tolerance in wine Saccharomyces cerevisiae strains by a backcross approach. FEMS Yeast Res. 2009, 9, 1148–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, A.; Durand, C.; Loira, N.; Durrens, P.; Sherman, D.J.; Marullo, P. QTL dissection of lag phase in wine fermentation reveals a new translocation responsible for Saccharomyces cerevisiae adaptation to sulfite. PLoS ONE 2014, 9, e86298. [Google Scholar] [CrossRef] [Green Version]
- da Silva, T.; Albertin, W.; Dillmann, C.; Bely, M.; Guerche, S.L.; Giraud, C.; Huet, S.; Sicard, D.; Masneuf-Pomarede, I.; Vienne, D.D.; et al. Hybridization within Saccharomyces genus results in homoeostasis and phenotypic novelty in winemaking conditions. PLoS ONE 2015, 10, e0123834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novo, M.; Bigey, F.; Beyne, E.; Galeote, V.; Gavory, F.; Mallet, S.; Cambon, B.; Legras, J.-L.; Wincker, P.; Casaregola, S.; et al. Eukaryote-to-eukaryote gene transfer events revealed by the genome sequence of the wine yeast Saccharomyces cerevisiae EC1118. Proc. Natl. Acad. Sci. USA 2009, 106, 16333–16338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goecks, J.; Nekrutenko, A.; Taylor, J.; Team, T.G. Galaxy: A comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 2010, 11, R86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq-a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; Volume 1, p. 409. [Google Scholar]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; Von Mering, C.; et al. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, D808–D815. [Google Scholar] [CrossRef] [Green Version]
- Antalick, G.; Perello, M.-C.; de Revel, G. Development, validation and application of a specific method for the quantitative determination of wine esters by headspace-solid-phase microextraction-gas chromatography–mass spectrometry. Food Chem. 2010, 121, 1236–1245. [Google Scholar] [CrossRef]
- Martin, N.; de Revel, G. Sensory evaluation: Scientific bases and oenological applications. J. Int. Sci. Vigne Vin 1999, 33, 81–93. [Google Scholar]
- Pineau, B.; Barbe, J.-C.; Van Leeuwen, C.; Dubourdieu, D. Examples of Perceptive Interactions Involved in Specific “Red-” and “Black-berry” Aromas in Red Wines. J. Agric. Food Chem. 2009, 57, 3702–3708. [Google Scholar] [CrossRef] [PubMed]
- Malcorps, P.; Cheval, J.M.; Jamil, S.; Dufour, J.-P. A new model for the regulation of ester synthesis by alcohol acetyltransferases in Saccharomyces cerevisiae. J. Am. Soc. Brew. Chem. 1991, 49, 47–53. [Google Scholar]
- Christiaens, J.F.; Franco, L.M.; Yaksi, E.; Verstrepen, K.J.; Cools, T.L.; De Meester, L.; Michiels, J.; Wenseleers, T.; Hassan, B.A. The fungal aroma gene ATF1 promotes dispersal of yeast cells through insect vectors. Cell Rep. 2014, 9, 425–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Esters | Metabolic Precursors | ||||
|---|---|---|---|---|---|
| Compounds | Family | Abbreviation | Compounds | Family | Abbreviation |
| ethyl propanoate | Fatty acid ethyl esters (FAEE) | C3C2 | propanoic acid | Volatile acids (VAc) | C3 |
| ethyl butanoate | C4C2 | butanoid acid | C4 | ||
| ethyl hexanoate | C6C2 | hexanoic acid | C6 | ||
| ethyl octanoate | C8C2 | octanoic acid | C8 | ||
| ethyl decanoate | C10C2 | ||||
| ethyl dodecanoate | C12C2 | ||||
| propyl acetate | Higher alcohol acetates (HAA) | C2C3 | propan-1-ol | Higher alcohols (HA) | C3OH |
| 2-methylpropyl acetate | C2iC4 | 2-methylpropan-1-ol | iC4OH | ||
| 3-methylbutyl acetate (isoamyl acetate) | C2iC5 | 3-methylbutanol (isoamyl alcohol) | iC5OH | ||
| hexyl acetate | C2C6 | ||||
| octyl acetate | C2C8 | ||||
| 2-phenylethyl acetate | C2PhC2 | ||||
| ethyl 2-methylpropanoate | Alkylated ethyl esters (AEE) | 2mC3C2 | 2-methylpropanoic acid | Alkylated acids (AAc) | 2mC3 |
| ethyl 2-methylbutanoate | 2mC4C2 | 2-methylbutanoic acid | 2mC4 | ||
| ethyl 3-methylbutanoate | 3mC4C2 | 3-methylbutanoic acid | 3mC4 | ||
| ethyl phenylacetate | PhC2C2 | ||||
| ethyl 2-hydroxy-4-methyl-pentanoate | Hydroxylated ethyl esters (HEE) | 2h4mC5C2 | ethyl 2-hydroxy-4-methyl-pentanoic acid | Hydroxylated acids (HAc) | 2h4mC5 |
| ethyl 3-hydroxy-butanoate | 3hC4C2 | ethyl 3-hydroxy-butanoic acid | 3hC4 | ||
| Strain | Background | Genotype | Description | Origin |
|---|---|---|---|---|
| Y31674 | BY4743 | BY4743;Mata/α;his3Δ1/his3Δ1;leu2Δ0/leu2Δ0;lys2Δ0/LYS2;MET15/met15Δ0,ura3Δ0/ura3Δ0;YOR377::kanMx4/YOR377::kanMx4 | ATF1 deletion | Euroscarf |
| Y34807 | BY4743 | BY4743;Mata/α;his3Δ1/his3Δ1;leu2Δ0/leu2Δ0;lys2Δ0/LYS2;MET15/met15Δ0,ura3Δ0/ura3Δ0;YGR177::kanMx4/YGR177::kanMx4 | ATF2 deletion | Euroscarf |
| Y33317 | BY4743 | BY4743;Mata/α;his3Δ1/his3Δ1;leu2Δ0/leu2Δ0;lys2Δ0/LYS2;MET15/met15Δ0,ura3Δ0/ura3Δ0;YRR177::kanMx4/YRR177::kanMx4 | EEB1 deletion | Euroscarf |
| Y32157 | BY4743 | BY4743;Mata/α;his3Δ1/his3Δ1;leu2Δ0/leu2Δ0;lys2Δ0/LYS2;MET15/met15Δ0,ura3Δ0/ura3Δ0;YPL095::kanMx4/YPL095::kanMx4 | EHT1 deletion | Euroscarf |
| Y30796 | BY4743 | BY4743;Mata/α;his3Δ1/his3Δ1;leu2Δ0/leu2Δ0;lys2Δ0/LYS2;MET15/met15Δ0,ura3Δ0/ura3Δ0;YMR210w::kanMx4/YMR210w::kanMx4 | MGL2 deletion | Euroscarf |
| Y34943 | BY4743 | BY4743;Mata/α;his3Δ1/his3Δ1;leu2Δ0/leu2Δ0;lys2Δ0/LYS2;MET15/met15Δ0,ura3Δ0/ura3Δ0;YKL094w::kanMx4/YKL094w::kanMx4 | YJU3 deletion | Euroscarf |
| Fx10 HO/ho::HYG | Fx10 | Fx10; Mata/α, HO/ho::HYG | Diploid homozygous | ISVV collection |
| Fx10-ΔA1 | Fx10 | Fx10; Mata/α; HO/ho::HYG; YOR377::kanMx4/YOR377::kanMx4 | ATF1 deletion | this study |
| Fx10-ΔA2 | Fx10 | Fx10; Mata/α, HO/ho::HYG; YGR177::kanMx4/YGR177::kanMx4 | ATF2 deletion | this study |
| Fx10-ΔE1 | Fx10 | Fx10; Mata/α, HO/ho::HYG; YRR177::kanMx4/YRR177::kanMx4 | EEB1 deletion | this study |
| Fx10-ΔE2 | Fx10 | Fx10; Mata/α, HO/ho::HYG; YPL095::kanMx4/YPL095::kanMx4 | EHT1 deletion | this study |
| Fx10-ΔA12 | Fx10 | Fx10; Mata/α, HO/ho::HYG; YOR377::kanMx4/YOR377::kanMx4; YGR177::kanMx4/YGR177::kanMx4 | ATF1, ATF2 deletion | this study |
| Fx10-ΔE12 | Fx10 | Fx10; Mata/α, HO/ho::HYG; YRR177::kanMx4/YRR177::kanMx4; YPL095::kanMx4/YPL095::kanMx4 | EEB1, EHT1 deletion | this study |
| Fx10-ΔAE | Fx10 | Fx10; Mata/α, HO/ho::HYG; YOR377::kanMx4/YOR377::kanMx4; YGR177::kanMx4/YGR177::kanMx4; YRR177::kanMx4/YRR177::kanMx4; YPL095::kanMx4/YPL095::kanMx4 | ATF1, ATF2, EEB1, EHT1 deletion | this study |
| Fx10-ΔM | Fx10 | Fx10; Mata/α, HO/ho::HYG, YMR210w::kanMx4/YMR210w::kanMx4 | MGL2 deletion | this study |
| Fx10-ΔY | Fx10 | Fx10; Mata/α, HO/ho::HYG; ;YKL094w::kanMx4/YKL094w::kanMx4 | YJU3 deletion | this study |
| Fx10-ΔME | Fx10 | Fx10; Mata/α, HO/ho::HYG, YMR210w::kanMx4/YMR210w::kanMx4, YRR177::kanMx4/YRR177::kanMx4; YPL095::kanMx4/YPL095::kanMx4 | MGL2, EEB1, EHT1 deletion | this study |
| Fx10-ΔMY | Fx10 | Fx10; Mata/α, HO/ho::HYG, YMR210w::kanMx4/YMR210w::kanMx4, ;YKL094w::kanMx4/YKL094w::kanMx4 | MGL2, YJU3 deletion | this study |
| Fx10-ΔAEM | Fx10 | Fx10; Mata/a, HO/ho::HYG; YOR377 /YOR377::kanMx4; YGR177 /YGR177::kanMx4; YRR177 /YRR177::kanMx4; YPL095 /YPL095::kanMx4; YMR210w /YMR210w::kanMx4 | Heterozygous hybrid for ATF1, ATF2, EEB1, EHT1, MGL2 deletion | this study |
| H1xH5 | Fx10 | Fx10; Mata/a, HO/ho::HYG; YOR377 /YOR377::kanMx4; YGR177 ::kanMx4/YGR177::kanMx4; YRR177::kanMx4/YRR177::kanMx4; YPL095 /YPL095::kanMx4; YMR210w /YMR210w::kanMx4 | this study | |
| H1xH6 | Fx10 | Fx10; Mata/a, HO/ho::HYG; YOR377 /YOR377::kanMx4; YGR177/YGR177::kanMx4; YRR177::kanMx4/YRR177::kanMx4; YPL095::kanMx4/YPL095::kanMx4; YMR210w /YMR210w::kanMx4 | this study | |
| Y31674 | BY4743 | BY4743;Mata/α;his3Δ1/his3Δ1;leu2Δ0/leu2Δ0;lys2Δ0/LYS2;MET15/met15Δ0,ura3Δ0/ura3Δ0;YOR377::kanMx4/YOR377::kanMx4 | ATF1 deletion | Euroscarf |
| Y34807 | BY4743 | BY4743;Mata/α;his3Δ1/his3Δ1;leu2Δ0/leu2Δ0;lys2Δ0/LYS2;MET15/met15Δ0,ura3Δ0/ura3Δ0;YGR177::kanMx4/YGR177::kanMx4 | ATF2 deletion | Euroscarf |
| Pedigree | Ploidy | Haplotype 1 | Viable | Unviable | Number of Functional Copies | ||||
|---|---|---|---|---|---|---|---|---|---|
| ATF1 | ATF2 | EEB1 | EHT1 | MGL2 | |||||
| Fx10-Delta5-progeny | n | H1 | 5 | 0 | 0 | 0 | 0 | 0 | 2 |
| n | H2 | 1 | 0 | 0 | 2 | 2 | 2 | 0 | |
| n | H3 | 1 | 0 | 0 | 2 | 2 | 0 | 0 | |
| n | H4 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | |
| n | H5 | 2 | 0 | 2 | 2 | 0 | 0 | 0 | |
| n | H6 | 2 | 0 | 2 | 0 | 0 | 2 | 0 | |
| F2-hybrids | 2n | H1/H2 | 0 | 20 | 0 | 1 | 1 | 1 | 1 |
| 2n | H1/H3 | 0 | 20 | 0 | 1 | 1 | 0 | 1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marullo, P.; Trujillo, M.; Viannais, R.; Hercman, L.; Guillaumie, S.; Colonna-Ceccaldi, B.; Albertin, W.; Barbe, J.-C. Metabolic, Organoleptic and Transcriptomic Impact of Saccharomyces cerevisiae Genes Involved in the Biosynthesis of Linear and Substituted Esters. Int. J. Mol. Sci. 2021, 22, 4026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084026
Marullo P, Trujillo M, Viannais R, Hercman L, Guillaumie S, Colonna-Ceccaldi B, Albertin W, Barbe J-C. Metabolic, Organoleptic and Transcriptomic Impact of Saccharomyces cerevisiae Genes Involved in the Biosynthesis of Linear and Substituted Esters. International Journal of Molecular Sciences. 2021; 22(8):4026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084026
Chicago/Turabian StyleMarullo, Philippe, Marine Trujillo, Rémy Viannais, Lucas Hercman, Sabine Guillaumie, Benoit Colonna-Ceccaldi, Warren Albertin, and Jean-Christophe Barbe. 2021. "Metabolic, Organoleptic and Transcriptomic Impact of Saccharomyces cerevisiae Genes Involved in the Biosynthesis of Linear and Substituted Esters" International Journal of Molecular Sciences 22, no. 8: 4026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084026