
Impaired Ca2+ Sensitivity of a Novel GCAP1 Variant Causes Cone Dystrophy and Leads to Abnormal Synaptic Transmission Between Photoreceptors and Bipolar Cells

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
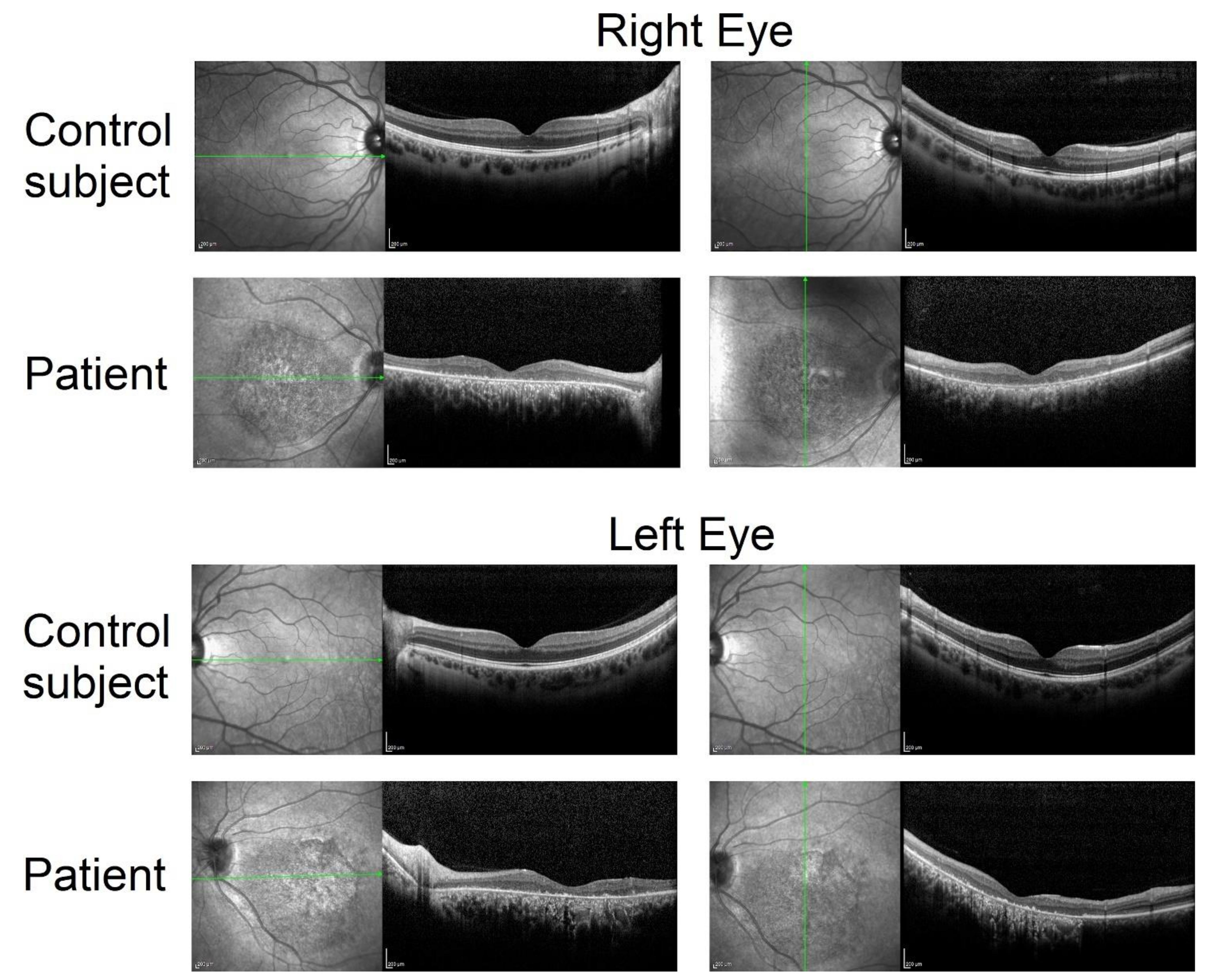
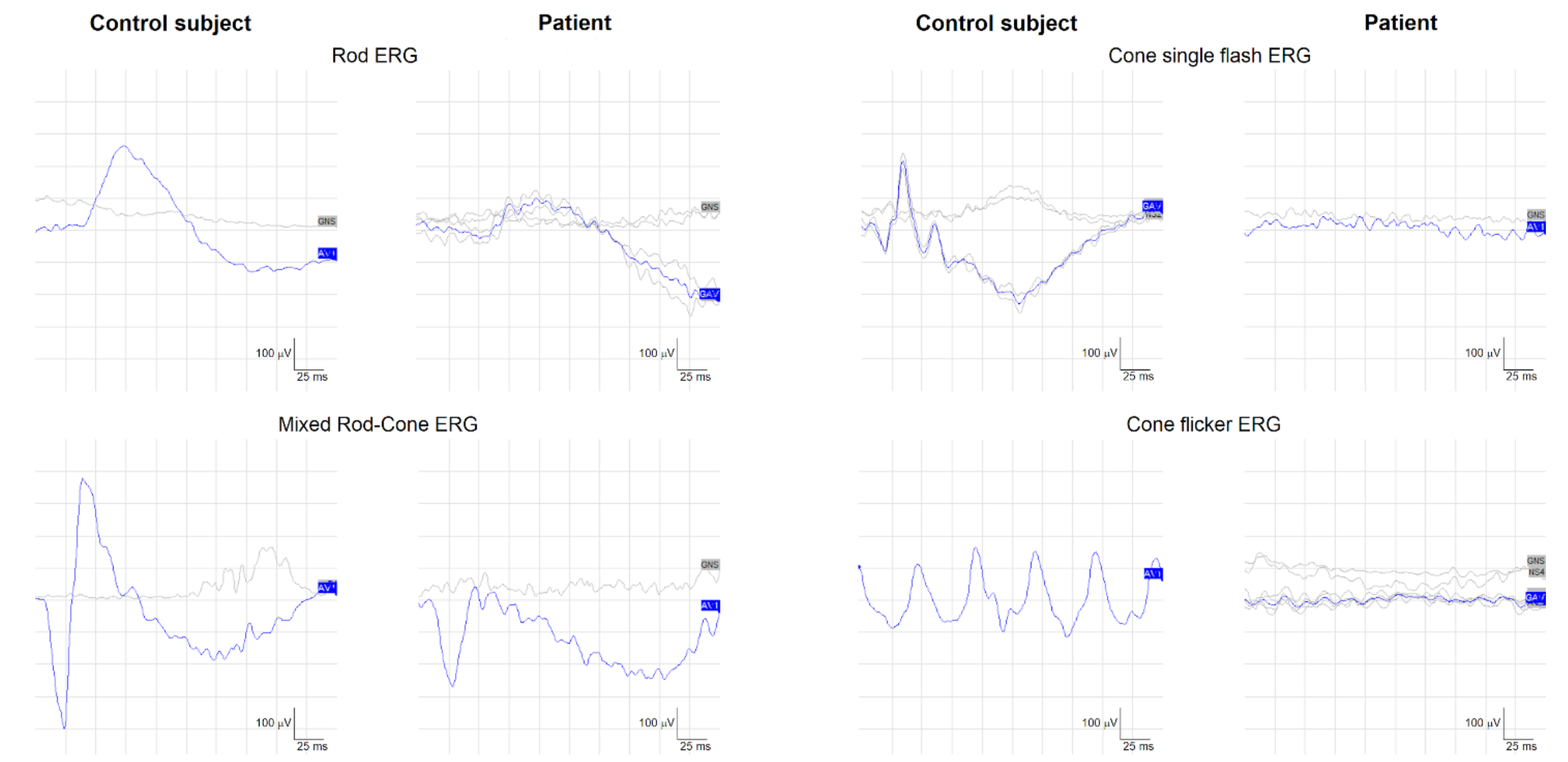
2.1. Clinical Phenotype and Disease Progression
2.2. Identification of a Novel Variant in GUCA1A in Heterozygosis
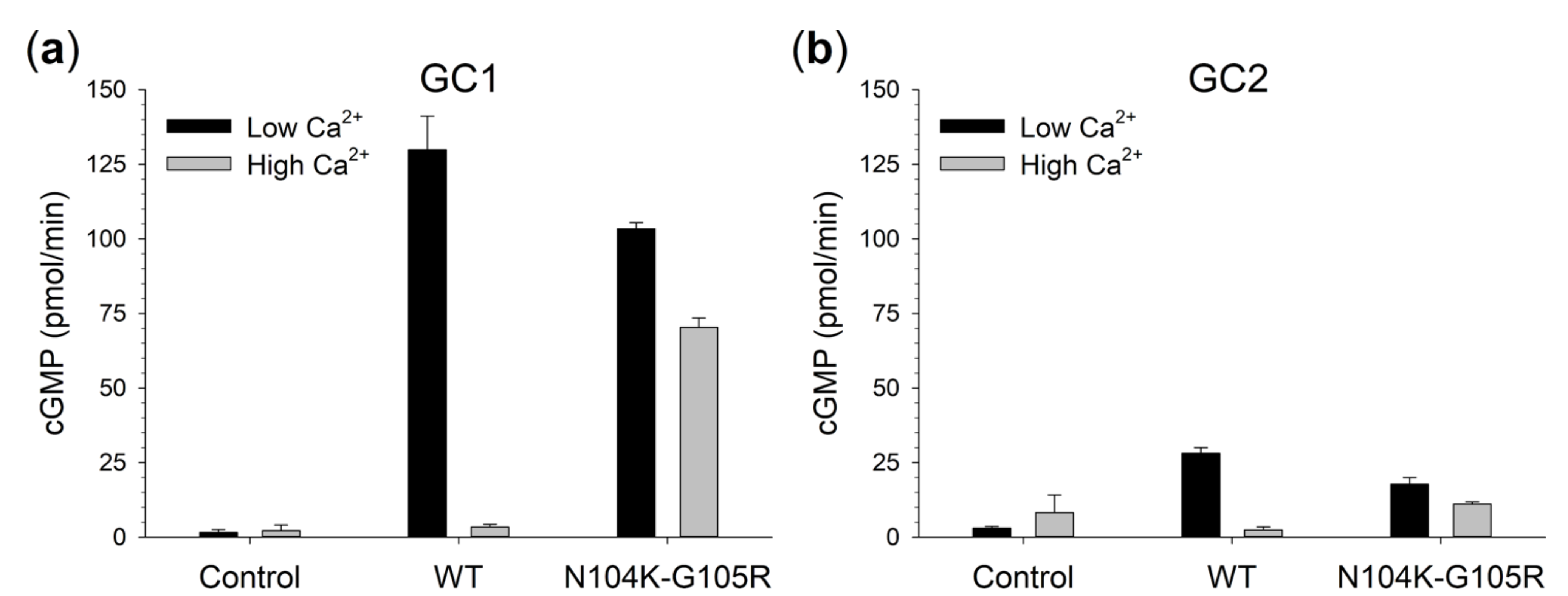
2.3. Guanylate Cyclase Regulation by N104K-G105R GCAP1
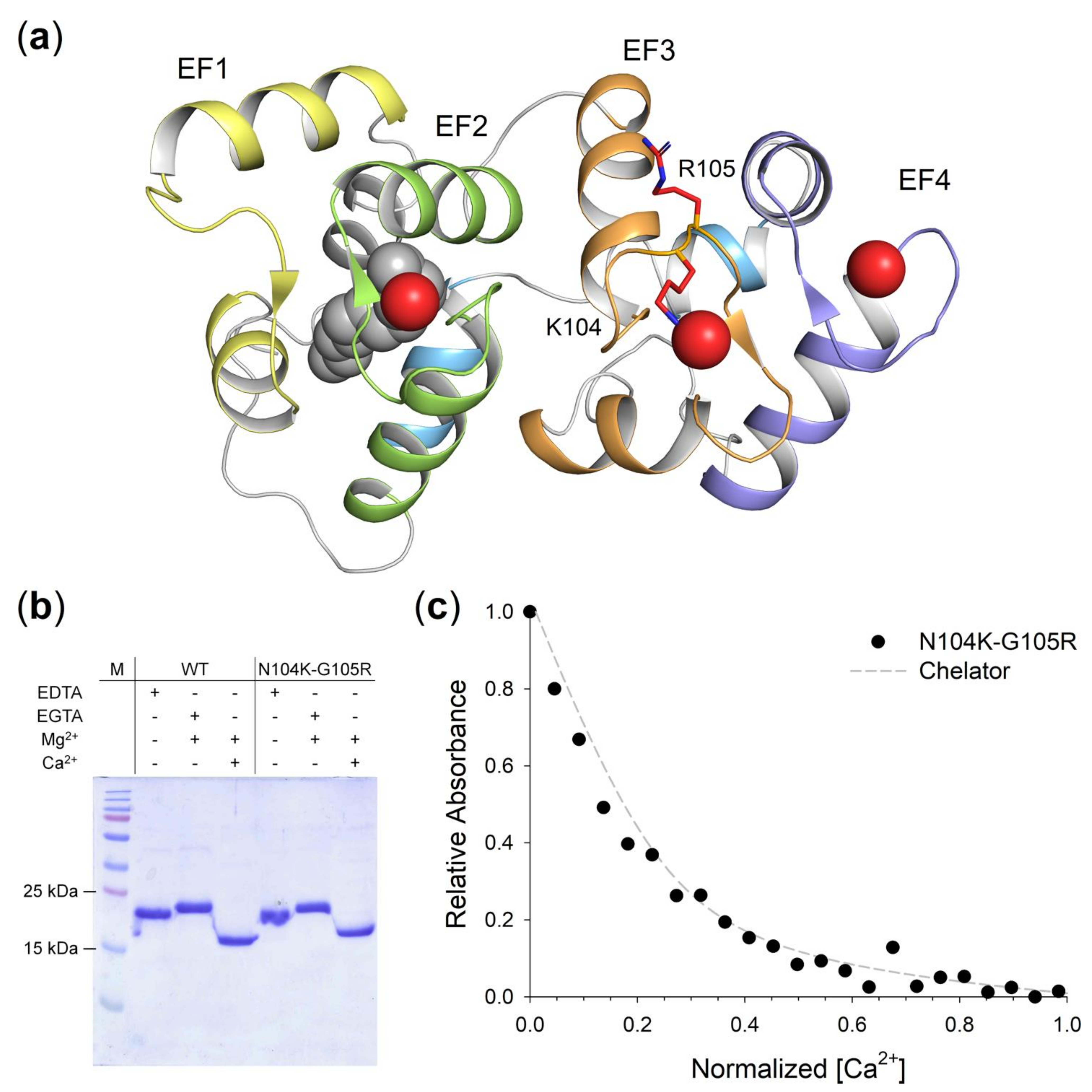
2.4. GCAP1 N104K-G105R Variant Presents Reduced Ca2+-Affinity
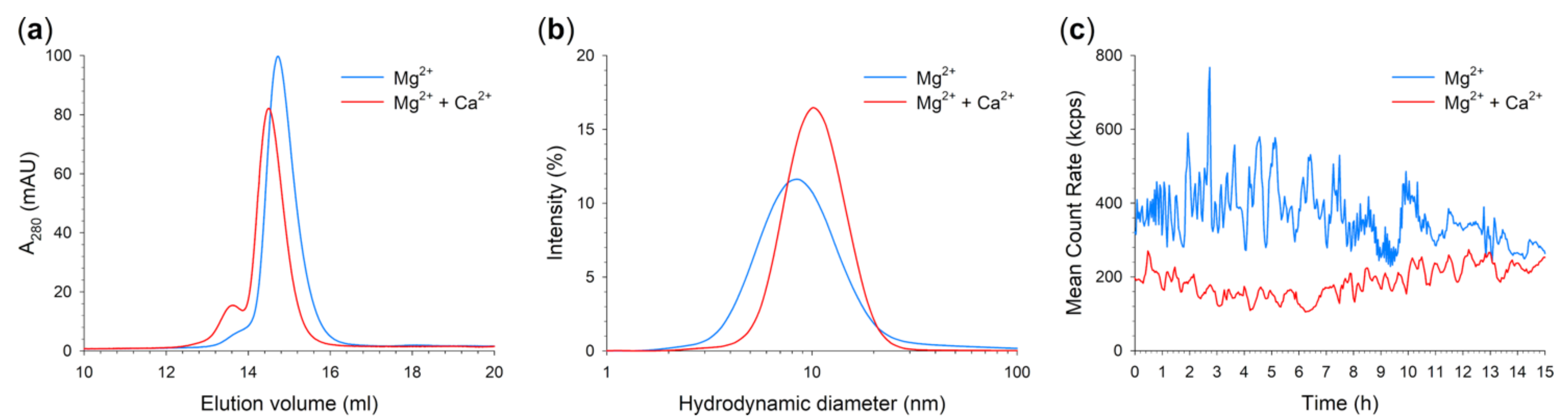
2.5. Quaternary Structure and Aggregation Propensity of N104K-G105R GCAP1
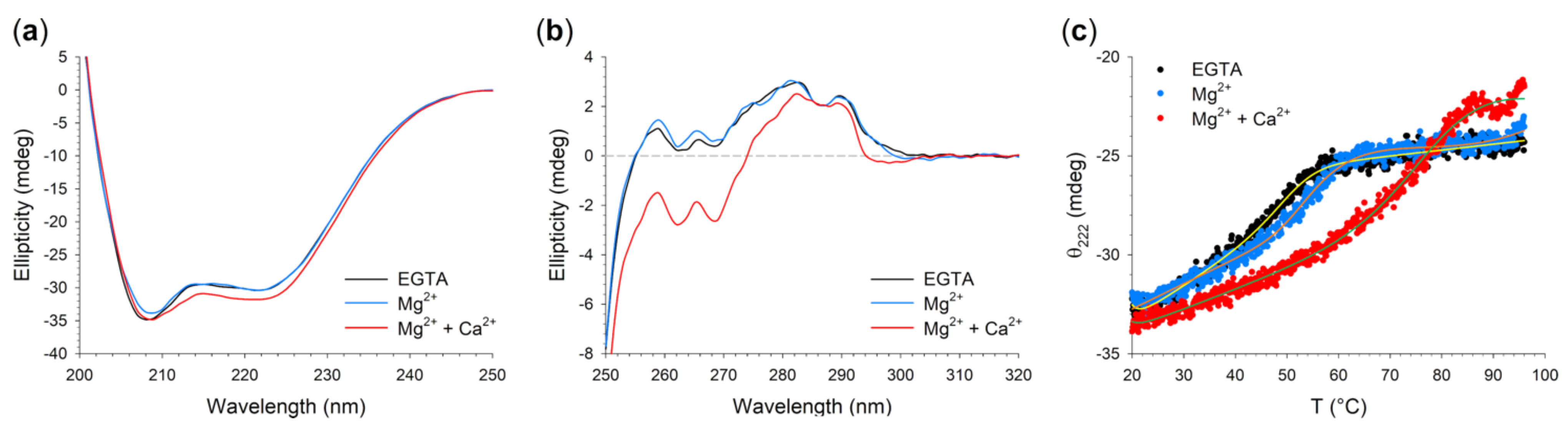
2.6. Structural and Stability Changes of N104K-G105R GCAP1 upon Cation Binding
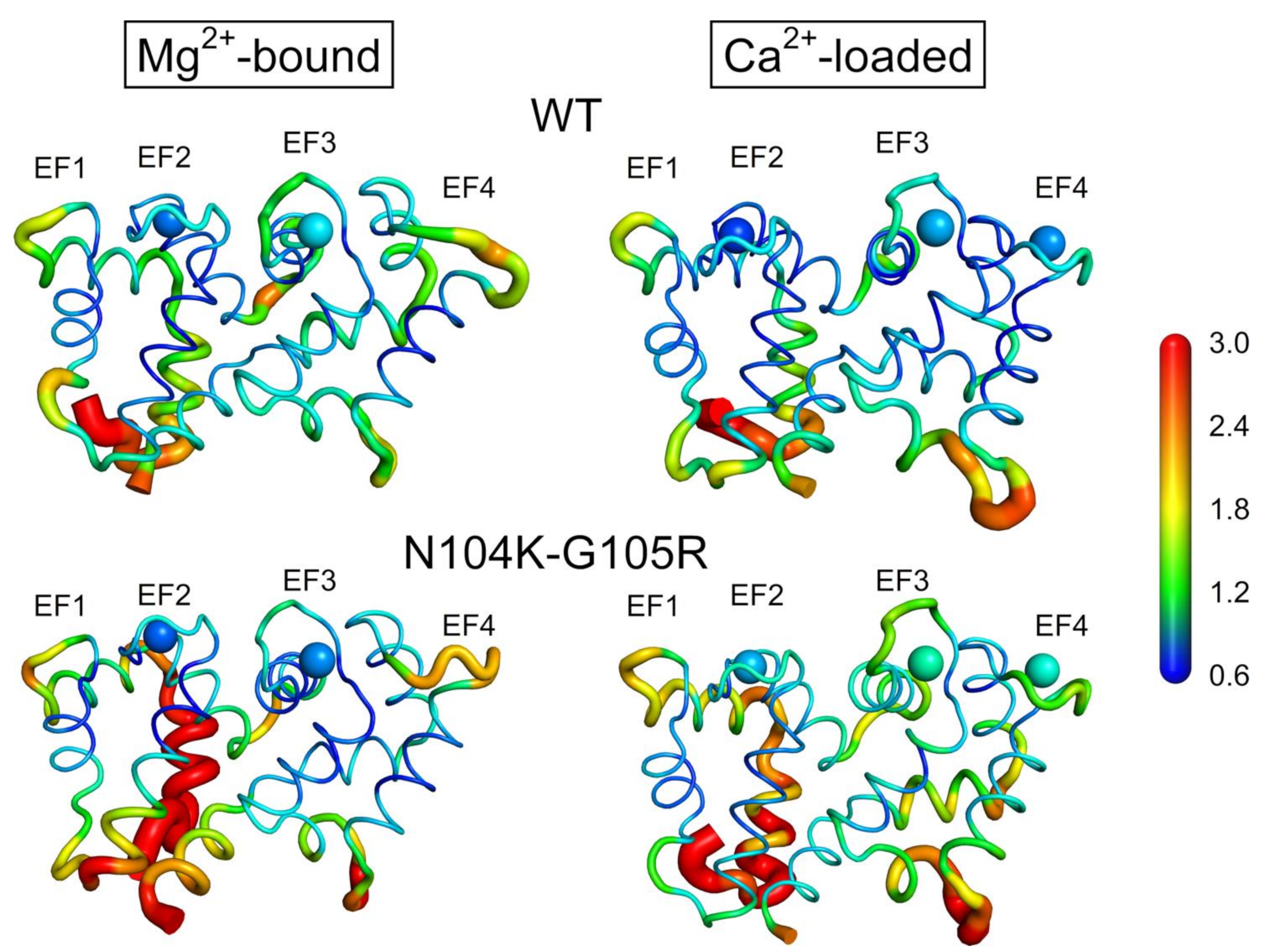
2.7. Flexibility of GCAP1 Variants at Atomistic Resolution Monitored by Molecular Dynamics Simulations
3. Discussion
4. Materials and Methods
4.1. Patient Studies, Clinical and Ophthalmological Examinations
4.2. Genetic Testing
4.3. Cloning, Protein Expression and Purification
4.4. Guanylate Cyclase Assay
4.5. Gel Shift Assay
4.6. Ca2+-Affinity Assay
4.7. Analytical Size Exclusion Chromatography
4.8. Circular Dichroism Spectroscopy and Thermal Denaturation Profiles
4.9. Dynamic Light Scattering
4.10. 8-Anilinonaphthalene-1-Sulfonic Acid Fluorescence Hydrophobicity Assay
4.11. Molecular Dynamics Simulations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koch, K.W.; Dell’Orco, D. A calcium-relay mechanism in vertebrate phototransduction. ACS Chem. Neurosci. 2013, 4, 909–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, K.W.; Dell’Orco, D. Protein and signaling networks in vertebrate photoreceptor cells. Front. Mol. Neurosci. 2015, 8, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korenbrot, J.I. Speed, sensitivity, and stability of the light response in rod and cone photoreceptors: Facts and models. Prog. Retin. Eye Res. 2012, 31, 442–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peshenko, I.V.; Dizhoor, A.M. Guanylyl cyclase-activating proteins (GCAPs) are Ca2+/Mg2+ sensors: Implications for photoreceptor guanylyl cyclase (RetGC) regulation in mammalian photoreceptors. J. Biol. Chem. 2004, 279, 16903–16906. [Google Scholar] [CrossRef] [Green Version]
- Peshenko, I.V.; Dizhoor, A.M. Ca2+ and Mg2+ binding properties of GCAP-1. Evidence that Mg2+-bound form is the physiological activator of photoreceptor guanylyl cyclase. J. Biol. Chem. 2006, 281, 23830–23841. [Google Scholar] [CrossRef] [Green Version]
- Koch, K.W.; Duda, T.; Sharma, R.K. Ca(2+)-modulated vision-linked ROS-GC guanylate cyclase transduction machinery. Mol. Cell Biochem. 2010, 334, 105–115. [Google Scholar] [CrossRef]
- Dizhoor, A.M.; Olshevskaya, E.V.; Peshenko, I.V. Mg2+/Ca2+ cation binding cycle of guanylyl cyclase activating proteins (GCAPs): Role in regulation of photoreceptor guanylyl cyclase. Mol. Cell Biochem. 2010, 334, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Peshenko, I.V.; Olshevskaya, E.V.; Savchenko, A.B.; Karan, S.; Palczewski, K.; Baehr, W.; Dizhoor, A.M. Enzymatic properties and regulation of the native isozymes of retinal membrane guanylyl cyclase (RetGC) from mouse photoreceptors. Biochemistry 2011, 50, 5590–5600. [Google Scholar] [CrossRef] [Green Version]
- Helten, A.; Saftel, W.; Koch, K.W. Expression level and activity profile of membrane bound guanylate cyclase type 2 in rod outer segments. J. Neurochem. 2007, 103, 1439–1446. [Google Scholar] [CrossRef]
- Manes, G.; Mamouni, S.; Herald, E.; Richard, A.C.; Senechal, A.; Aouad, K.; Bocquet, B.; Meunier, I.; Hamel, C.P. Cone dystrophy or macular dystrophy associated with novel autosomal dominant GUCA1A mutations. Mol. Vis. 2017, 23, 198–209. [Google Scholar]
- Abbas, S.; Marino, V.; Weisschuh, N.; Kieninger, S.; Solaki, M.; Dell’Orco, D.; Koch, K.W. Neuronal calcium sensor GCAP1 Encoded by GUCA1A exhibits heterogeneous functional properties in two cases of retinitis pigmentosa. ACS Chem. Neurosci. 2020, 11, 1458–1470. [Google Scholar] [CrossRef]
- Marino, V.; Dal Cortivo, G.; Oppici, E.; Maltese, P.E.; D’Esposito, F.; Manara, E.; Ziccardi, L.; Falsini, B.; Magli, A.; Bertelli, M.; et al. A novel p.(Glu111Val) missense mutation in GUCA1A associated with cone-rod dystrophy leads to impaired calcium sensing and perturbed second messenger homeostasis in photoreceptors. Hum. Mol. Genet. 2018, 27, 4204–4217. [Google Scholar] [CrossRef]
- Marino, V.; Scholten, A.; Koch, K.W.; Dell’Orco, D. Two retinal dystrophy-associated missense mutations in GUCA1A with distinct molecular properties result in a similar aberrant regulation of the retinal guanylate cyclase. Hum. Mol. Genet. 2015, 24, 6653–6666. [Google Scholar] [CrossRef] [Green Version]
- Dizhoor, A.M.; Boikov, S.G.; Olshevskaya, E.V. Constitutive activation of photoreceptor guanylate cyclase by Y99C mutant of GCAP-1. Possible role in causing human autosomal dominant cone degeneration. J. Biol. Chem. 1998, 273, 17311–17314. [Google Scholar] [CrossRef] [Green Version]
- Sokal, I.; Li, N.; Surgucheva, I.; Warren, M.J.; Payne, A.M.; Bhattacharya, S.S.; Baehr, W.; Palczewski, K. GCAP1 (Y99C) mutant is constitutively active in autosomal dominant cone dystrophy. Mol. Cell 1998, 2, 129–133. [Google Scholar] [CrossRef]
- Wilkie, S.E.; Li, Y.; Deery, E.C.; Newbold, R.J.; Garibaldi, D.; Bateman, J.B.; Zhang, H.; Lin, W.; Zack, D.J.; Bhattacharya, S.S.; et al. Identification and functional consequences of a new mutation (E155G) in the gene for GCAP1 that causes autosomal dominant cone dystrophy. Am. J. Hum. Genet. 2001, 69, 471–480. [Google Scholar] [CrossRef] [Green Version]
- Nishiguchi, K.M.; Sokal, I.; Yang, L.; Roychowdhury, N.; Palczewski, K.; Berson, E.L.; Dryja, T.P.; Baehr, W. A novel mutation (I143NT) in guanylate cyclase-activating protein 1 (GCAP1) associated with autosomal dominant cone degeneration. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3863–3870. [Google Scholar] [CrossRef]
- Sokal, I.; Dupps, W.J.; Grassi, M.A.; Brown, J., Jr.; Affatigato, L.M.; Roychowdhury, N.; Yang, L.; Filipek, S.; Palczewski, K.; Stone, E.M.; et al. A novel GCAP1 missense mutation (L151F) in a large family with autosomal dominant cone-rod dystrophy (adCORD). Investig. Ophthalmol. Vis. Sci. 2005, 46, 1124–1132. [Google Scholar] [CrossRef]
- Jiang, L.; Wheaton, D.; Bereta, G.; Zhang, K.; Palczewski, K.; Birch, D.G.; Baehr, W. A novel GCAP1(N104K) mutation in EF-hand 3 (EF3) linked to autosomal dominant cone dystrophy. Vis. Res. 2008, 48, 2425–2432. [Google Scholar] [CrossRef] [Green Version]
- Dell’Orco, D.; Behnen, P.; Linse, S.; Koch, K.W. Calcium binding, structural stability and guanylate cyclase activation in GCAP1 variants associated with human cone dystrophy. Cell Mol. Life Sci. 2010, 67, 973–984. [Google Scholar] [CrossRef]
- Kamenarova, K.; Corton, M.; Garcia-Sandoval, B.; Fernandez-San Jose, P.; Panchev, V.; Avila-Fernandez, A.; Lopez-Molina, M.I.; Chakarova, C.; Ayuso, C.; Bhattacharya, S.S. Novel GUCA1A mutations suggesting possible mechanisms of pathogenesis in cone, cone-rod, and macular dystrophy patients. Biomed. Res. Int. 2013, 2013, 517570. [Google Scholar] [CrossRef] [Green Version]
- Kitiratschky, V.B.; Behnen, P.; Kellner, U.; Heckenlively, J.R.; Zrenner, E.; Jagle, H.; Kohl, S.; Wissinger, B.; Koch, K.W. Mutations in the GUCA1A gene involved in hereditary cone dystrophies impair calcium-mediated regulation of guanylate cyclase. Hum. Mutat. 2009, 30, E782–E796. [Google Scholar] [CrossRef]
- Vocke, F.; Weisschuh, N.; Marino, V.; Malfatti, S.; Jacobson, S.G.; Reiff, C.M.; Dell’Orco, D.; Koch, K.W. Dysfunction of cGMP signalling in photoreceptors by a macular dystrophy-related mutation in the calcium sensor GCAP1. Hum. Mol. Genet. 2017, 26, 133–144. [Google Scholar] [CrossRef]
- Peshenko, I.V.; Cideciyan, A.V.; Sumaroka, A.; Olshevskaya, E.V.; Scholten, A.; Abbas, S.; Koch, K.W.; Jacobson, S.G.; Dizhoor, A.M. A G86R mutation in the calcium-sensor protein GCAP1 alters regulation of retinal guanylyl cyclase and causes dominant cone-rod degeneration. J. Biol. Chem. 2019, 294, 3476–3488. [Google Scholar] [CrossRef] [Green Version]
- Gill, J.S.; Georgiou, M.; Kalitzeos, A.; Moore, A.T.; Michaelides, M. Progressive cone and cone-rod dystrophies: Clinical features, molecular genetics and prospects for therapy. Br. J. Ophthalmol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Dell’Orco, D.; Dal Cortivo, G. Normal GCAPs partly compensate for altered cGMP signaling in retinal dystrophies associated with mutations in GUCA1A. Sci. Rep. 2019, 9, 20105. [Google Scholar] [CrossRef]
- Abed, E.; Placidi, G.; Calandriello, L.; Piccardi, M.; Campagna, F.; Bertelli, M.; Minnella, A.M.; Savastano, M.C.; Falsini, B. Correlation of macular focal electroretinogram with ellipsoid zone extension in stargardt disease. J. Ophthalmol. 2017, 2017, 3643495. [Google Scholar] [CrossRef] [Green Version]
- Galli-Resta, L.; Falsini, B.; Rossi, G.; Piccardi, M.; Ziccardi, L.; Fadda, A.; Minnella, A.; Marangoni, D.; Placidi, G.; Campagna, F.; et al. Bilateral symmetry of visual function loss in cone-rod dystrophies. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3759–3768. [Google Scholar] [CrossRef] [Green Version]
- Abed, E.; Placidi, G.; Campagna, F.; Federici, M.; Minnella, A.; Guerri, G.; Bertelli, M.; Piccardi, M.; Galli-Resta, L.; Falsini, B. Early impairment of the full-field photopic negative response in patients with Stargardt disease and pathogenic variants of the ABCA4 gene. Clin. Exp. Ophthalmol. 2018, 46, 519–530. [Google Scholar] [CrossRef]
- Sieving, P.A. Photopic ON- and OFF-pathway abnormalities in retinal dystrophies. Trans. Am. Ophthalmol. Soc. 1993, 91, 701–773. [Google Scholar]
- Fujii, N.; Shiono, T.; Wada, Y.; Nakazawa, M.; Tamai, M.; Yamada, N. Autosomal dominant cone-rod dystrophy with negative electroretinogram. Br. J. Ophthalmol. 1995, 79, 916–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American college of medical genetics and genomics and the association for molecular pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Viviano, J.; Krishnan, A.; Wu, H.; Venkataraman, V. Electrophoretic mobility shift in native gels indicates calcium-dependent structural changes of neuronal calcium sensor proteins. Anal. Biochem. 2016, 494, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Dell’Orco, D.; Sulmann, S.; Linse, S.; Koch, K.W. Dynamics of conformational Ca2+-switches in signaling networks detected by a planar plasmonic device. Anal. Chem. 2012, 84, 2982–2989. [Google Scholar] [CrossRef] [PubMed]
- Linse, S. Calcium binding to proteins studied via competition with chromophoric chelators. Methods Mol. Biol. 2002, 173, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; Roseman, G.; Peshenko, I.; Manchala, G.; Cudia, D.; Dizhoor, A.M.; Millhauser, G.; Ames, J.B. Retinal guanylyl cyclase activating protein 1 forms a functional dimer. PLoS ONE 2018, 13, e0193947. [Google Scholar] [CrossRef] [Green Version]
- Boni, F.; Marino, V.; Bidoia, C.; Mastrangelo, E.; Barbiroli, A.; Dell’Orco, D.; Milani, M. Modulation of guanylate cyclase activating protein 1 (GCAP1) dimeric assembly by Ca(2+) or Mg(2+): Hints to understand protein activity. Biomolecules 2020, 10, 1408. [Google Scholar] [CrossRef]
- Dal Cortivo, G.; Marino, V.; Boni, F.; Milani, M.; Dell’Orco, D. Missense mutations affecting Ca(2+)-coordination in GCAP1 lead to cone-rod dystrophies by altering protein structural and functional properties. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118794. [Google Scholar] [CrossRef]
- Dal Cortivo, G.; Marino, V.; Iacobucci, C.; Vallone, R.; Arlt, C.; Rehkamp, A.; Sinz, A.; Dell’Orco, D. Oligomeric state, hydrodynamic properties and target recognition of human Calcium and Integrin Binding protein 2 (CIB2). Sci. Rep. 2019, 9, 15058. [Google Scholar] [CrossRef] [Green Version]
- Abbas, S.; Marino, V.; Bielefeld, L.; Koch, K.W.; Dell’Orco, D. Constitutive activation of guanylate cyclase by the G86R GCAP1 variant is due to “Locking” Cation-pi interactions that impair the activator-to-inhibitor structural transition. Int. J. Mol. Sci. 2020, 21, 752. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, F. Presynaptic [Ca(2+)] and GCAPs: Aspects on the structure and function of photoreceptor ribbon synapses. Front. Mol. Neurosci. 2014, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Spiwoks-Becker, I.; Glas, M.; Lasarzik, I.; Vollrath, L. Mouse photoreceptor synaptic ribbons lose and regain material in response to illumination changes. Eur. J. Neurosci. 2004, 19, 1559–1571. [Google Scholar] [CrossRef] [PubMed]
- Regus-Leidig, H.; Specht, D.; Tom Dieck, S.; Brandstatter, J.H. Stability of active zone components at the photoreceptor ribbon complex. Mol. Vis. 2010, 16, 2690–2700. [Google Scholar]
- Venkatesan, J.K.; Natarajan, S.; Schwarz, K.; Mayer, S.I.; Alpadi, K.; Magupalli, V.G.; Sung, C.H.; Schmitz, F. Nicotinamide adenine dinucleotide-dependent binding of the neuronal Ca2+ sensor protein GCAP2 to photoreceptor synaptic ribbons. J. Neurosci. 2010, 30, 6559–6576. [Google Scholar] [CrossRef]
- Lopez-del Hoyo, N.; Fazioli, L.; Lopez-Begines, S.; Fernandez-Sanchez, L.; Cuenca, N.; Llorens, J.; de la Villa, P.; Mendez, A. Overexpression of guanylate cyclase activating protein 2 in rod photoreceptors in vivo leads to morphological changes at the synaptic ribbon. PLoS ONE 2012, 7, e42994. [Google Scholar] [CrossRef] [Green Version]
- Avesani, A.; Marino, V.; Zanzoni, S.; Koch, K.W.; Dell’Orco, D. Molecular properties of human guanylate cyclase-activating protein 2 (GCAP2) and its retinal dystrophy-associated variant G157R. J. Biol. Chem. 2021. [Google Scholar] [CrossRef]
- Dizhoor, A.M.; Olshevskaya, E.V.; Henzel, W.J.; Wong, S.C.; Stults, J.T.; Ankoudinova, I.; Hurley, J.B. Cloning, sequencing, and expression of a 24-kDa Ca(2+)-binding protein activating photoreceptor guanylyl cyclase. J. Biol. Chem. 1995, 270, 25200–25206. [Google Scholar] [CrossRef] [Green Version]
- Frins, S.; Bonigk, W.; Muller, F.; Kellner, R.; Koch, K.W. Functional characterization of a guanylyl cyclase-activating protein from vertebrate rods. Cloning, heterologous expression, and localization. J. Biol. Chem. 1996, 271, 8022–8027. [Google Scholar] [CrossRef] [Green Version]
- Kachi, S.; Nishizawa, Y.; Olshevskaya, E.; Yamazaki, A.; Miyake, Y.; Wakabayashi, T.; Dizhoor, A.; Usukura, J. Detailed localization of photoreceptor guanylate cyclase activating protein-1 and -2 in mammalian retinas using light and electron microscopy. Exp. Eye Res. 1999, 68, 465–473. [Google Scholar] [CrossRef]
- Goraczniak, R.; Duda, T.; Sharma, R.K. Structural and functional characterization of a second subfamily member of the calcium-modulated bovine rod outer segment membrane guanylate cyclase, ROS-GC2. Biochem. Biophys. Res. Commun. 1997, 234, 666–670. [Google Scholar] [CrossRef]
- Helten, A.; Koch, K.W. Calcium-dependent conformational changes in guanylate cyclase-activating protein 2 monitored by cysteine accessibility. Biochem. Biophys. Res. Commun. 2007, 356, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Cooper, N.; Liu, L.; Yoshida, A.; Pozdnyakov, N.; Margulis, A.; Sitaramayya, A. The bovine rod outer segment guanylate cyclase, ROS-GC, is present in both outer segment and synaptic layers of the retina. J. Mol. Neurosci. 1995, 6, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, V.; Duda, T.; Vardi, N.; Koch, K.W.; Sharma, R.K. Calcium-modulated guanylate cyclase transduction machinery in the photoreceptor--bipolar synaptic region. Biochemistry 2003, 42, 5640–5648. [Google Scholar] [CrossRef] [Green Version]
- Duda, T.; Koch, K.W.; Venkataraman, V.; Lange, C.; Beyermann, M.; Sharma, R.K. Ca(2+) sensor S100beta-modulated sites of membrane guanylate cyclase in the photoreceptor-bipolar synapse. EMBO J. 2002, 21, 2547–2556. [Google Scholar] [CrossRef] [Green Version]
- Renner, A.B.; Kellner, U.; Cropp, E.; Foerster, M.H. Dysfunction of transmission in the inner retina: Incidence and clinical causes of negative electroretinogram. Graefes. Arch. Clin. Exp. Ophthalmol. 2006, 244, 1467–1473. [Google Scholar] [CrossRef]
- Kellner, U.; Foerster, M.H. Cone dystrophies with negative photopic electroretinogram. Br. J. Ophthalmol. 1993, 77, 404–409. [Google Scholar] [CrossRef]
- Miyake, Y.; Shiroyama, N.; Horiguchi, M.; Saito, A.; Yagasaki, K. Bull’s-eye maculopathy and negative electroretinogram. Retina 1989, 9, 210–215. [Google Scholar] [CrossRef]
- Cibis, G.W.; Fitzgerald, K.M. The negative ERG is not synonymous with nightblindness. Trans. Am. Ophthalmol. Soc. 2001, 99, 171–175; discussion 175–176. [Google Scholar]
- Hayashi, T.; Hosono, K.; Kurata, K.; Katagiri, S.; Mizobuchi, K.; Ueno, S.; Kondo, M.; Nakano, T.; Hotta, Y. Coexistence of GNAT1 and ABCA4 variants associated with Nougaret-type congenital stationary night blindness and childhood-onset cone-rod dystrophy. Doc. Ophthalmol. 2020, 140, 147–157. [Google Scholar] [CrossRef]
- Khan, N.W.; Kondo, M.; Hiriyanna, K.T.; Jamison, J.A.; Bush, R.A.; Sieving, P.A. Primate retinal signaling pathways: Suppressing ON-pathway activity in monkey with glutamate analogues mimics human CSNB1-NYX genetic night blindness. J. Neurophysiol. 2005, 93, 481–492. [Google Scholar] [CrossRef] [Green Version]
- Cideciyan, A.V.; Jacobson, S.G. Negative electroretinograms in retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 1993, 34, 3253–3263. [Google Scholar]
- Strettoi, E.; Pignatelli, V.; Rossi, C.; Porciatti, V.; Falsini, B. Remodeling of second-order neurons in the retina of rd/rd mutant mice. Vis. Res. 2003, 43, 867–877. [Google Scholar] [CrossRef] [Green Version]
- Power, M.; Das, S.; Schutze, K.; Marigo, V.; Ekstrom, P.; Paquet-Durand, F. Cellular mechanisms of hereditary photoreceptor degeneration—Focus on cGMP. Prog. Retin. Eye Res. 2020, 74, 100772. [Google Scholar] [CrossRef]
- Maltese, P.E.; Orlova, N.; Krasikova, E.; Emelyanchik, E.; Cheremisina, A.; Kuscaeva, A.; Salmina, A.; Miotto, R.; Bonizzato, A.; Guerri, G.; et al. Gene-targeted analysis of clinically diagnosed long QT russian families. Int. Heart J. 2017, 58, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Marceddu, G.; Dallavilla, T.; Guerri, G.; Manara, E.; Chiurazzi, P.; Bertelli, M. PipeMAGI: An integrated and validated workflow for analysis of NGS data for clinical diagnostics. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6753–6765. [Google Scholar] [CrossRef]
- Kopanos, C.; Tsiolkas, V.; Kouris, A.; Chapple, C.E.; Albarca Aguilera, M.; Meyer, R.; Massouras, A. VarSome: The human genomic variant search engine. Bioinformatics 2019, 35, 1978–1980. [Google Scholar] [CrossRef]
- Hwang, J.Y.; Koch, K.W. Calcium- and myristoyl-dependent properties of guanylate cyclase-activating protein-1 and protein-2. Biochemistry 2002, 41, 13021–13028. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zagel, P.; Dell’Orco, D.; Koch, K.W. The dimerization domain in outer segment guanylate cyclase is a Ca(2)(+)-sensitive control switch module. Biochemistry 2013, 52, 5065–5074. [Google Scholar] [CrossRef]
- Andre, I.; Linse, S. Measurement of Ca2+-binding constants of proteins and presentation of the CaLigator software. Anal. Biochem. 2002, 305, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Vallone, R.; Dal Cortivo, G.; D’Onofrio, M.; Dell’Orco, D. Preferential Binding of Mg(2+) Over Ca(2+) to CIB2 Triggers an Allosteric Switch Impaired in Usher Syndrome Type 1J. Front. Mol. Neurosci. 2018, 11, 274. [Google Scholar] [CrossRef] [PubMed]
- Marino, V.; Borsatto, A.; Vocke, F.; Koch, K.W.; Dell’Orco, D. CaF2 nanoparticles as surface carriers of GCAP1, a calcium sensor protein involved in retinal dystrophies. Nanoscale 2017, 9, 11773–11784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephen, R.; Bereta, G.; Golczak, M.; Palczewski, K.; Sousa, M.C. Stabilizing function for myristoyl group revealed by the crystal structure of a neuronal calcium sensor, guanylate cyclase-activating protein 1. Structure 2007, 15, 1392–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, V.; Sulmann, S.; Koch, K.W.; Dell’Orco, D. Structural effects of Mg(2)(+) on the regulatory states of three neuronal calcium sensors operating in vertebrate phototransduction. Biochim. Biophys. Acta 2015, 1853, 2055–2065. [Google Scholar] [CrossRef] [Green Version]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Rauscher, S.; Nawrocki, G.; Ran, T.; Feig, M.; de Groot, B.L.; Grubmuller, H.; MacKerell, A.D., Jr. CHARMM36m: An improved force field for folded and intrinsically disordered proteins. Nat. Methods 2017, 14, 71–73. [Google Scholar] [CrossRef] [Green Version]
- Marino, V.; Dell’Orco, D. Allosteric communication pathways routed by Ca(2+)/Mg(2+) exchange in GCAP1 selectively switch target regulation modes. Sci. Rep. 2016, 6, 34277. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Optical Coherence Tomography | Fundus Autofluorescence | |||||
|---|---|---|---|---|---|---|
| Central Macular Thickness | Ellipsoid Zone Length | Area of Atrophy | ||||
| Year | RE a | LE b | RE a | LE b | RE a | LE b |
| 2018 | 144 | 61 | 0 | 0 | 5.77 mm2 | 8.35 mm2 |
| 2019 | 143 | n.a. | 0 | 0 | 4.72 mm2 | 7.71 mm2 |
| 2020 | 97 | 105 | 0 | 0 | 5.97 mm2 | 8.45 mm2 |
| Variant | Condition | MM a (kDa) | d b (nm) | nc | PdI d |
|---|---|---|---|---|---|
| WT e | Mg2+ | 45.9 | 6.35 ± 0.07 | 27 | 0.342 ± 0.02 |
| Mg2+ + Ca2+ | 47.8 | 6.85 ± 0.17 | 20 | 0.306 ± 0.02 | |
| N104K-G105R | Mg2+ | 38.2 | 9.43 ± 0.07 | 400 | 0.278 ± 0.002 |
| Mg2+ + Ca2+ | 40.2 | 10.80 ± 0.05 | 400 | 0.214 ± 0.002 |
| Variant | Condition | Imax/IANS a | θ222/θ208 b | Δθ/θ (%) c | Tm (°C) d | Unfolding (%) e |
|---|---|---|---|---|---|---|
| WT | EGTA | 3.4 f | 0.90 f | - | 54.1 g | 24.6 g |
| Mg2+ | 2.5 f | 0.91 f | 2.8 f | 58.0 g | 30.8 g | |
| Mg2+ + Ca2+ | 2.4 f | 0.95 f | 7.7 f | >96 g | 30.4 g | |
| N104K-G105R | EGTA | 8.4 | 0.87 | - | 51.1 | 24.6 |
| Mg2+ | 4.8 | 0.90 | 0.1 | 60.8 | 26.3 | |
| Mg2+ + Ca2+ | 5.0 | 0.92 | 4.6 | 79.4 | 36.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marino, V.; Dal Cortivo, G.; Maltese, P.E.; Placidi, G.; De Siena, E.; Falsini, B.; Bertelli, M.; Dell’Orco, D. Impaired Ca2+ Sensitivity of a Novel GCAP1 Variant Causes Cone Dystrophy and Leads to Abnormal Synaptic Transmission Between Photoreceptors and Bipolar Cells. Int. J. Mol. Sci. 2021, 22, 4030. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084030
Marino V, Dal Cortivo G, Maltese PE, Placidi G, De Siena E, Falsini B, Bertelli M, Dell’Orco D. Impaired Ca2+ Sensitivity of a Novel GCAP1 Variant Causes Cone Dystrophy and Leads to Abnormal Synaptic Transmission Between Photoreceptors and Bipolar Cells. International Journal of Molecular Sciences. 2021; 22(8):4030. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084030
Chicago/Turabian StyleMarino, Valerio, Giuditta Dal Cortivo, Paolo Enrico Maltese, Giorgio Placidi, Elisa De Siena, Benedetto Falsini, Matteo Bertelli, and Daniele Dell’Orco. 2021. "Impaired Ca2+ Sensitivity of a Novel GCAP1 Variant Causes Cone Dystrophy and Leads to Abnormal Synaptic Transmission Between Photoreceptors and Bipolar Cells" International Journal of Molecular Sciences 22, no. 8: 4030. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084030