
Antarctic Rahnella inusitata: A Producer of Cold-Stable β-Galactosidase Enzymes

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
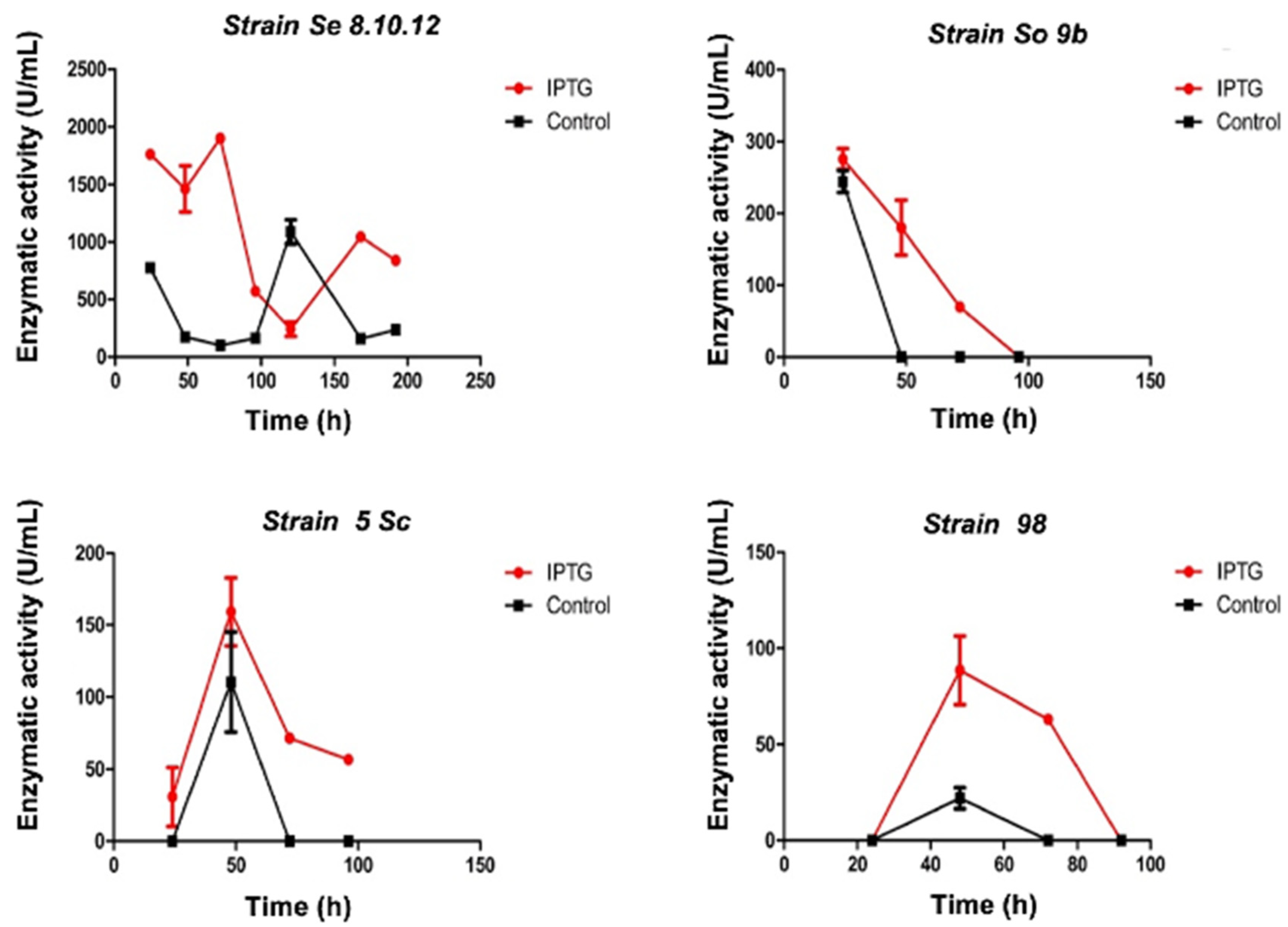
2.1. A Rahnella Strain Exhibited the Highest Enzymatic Activity among Multiple Antarctic Strains with β-Galactosidase Activity
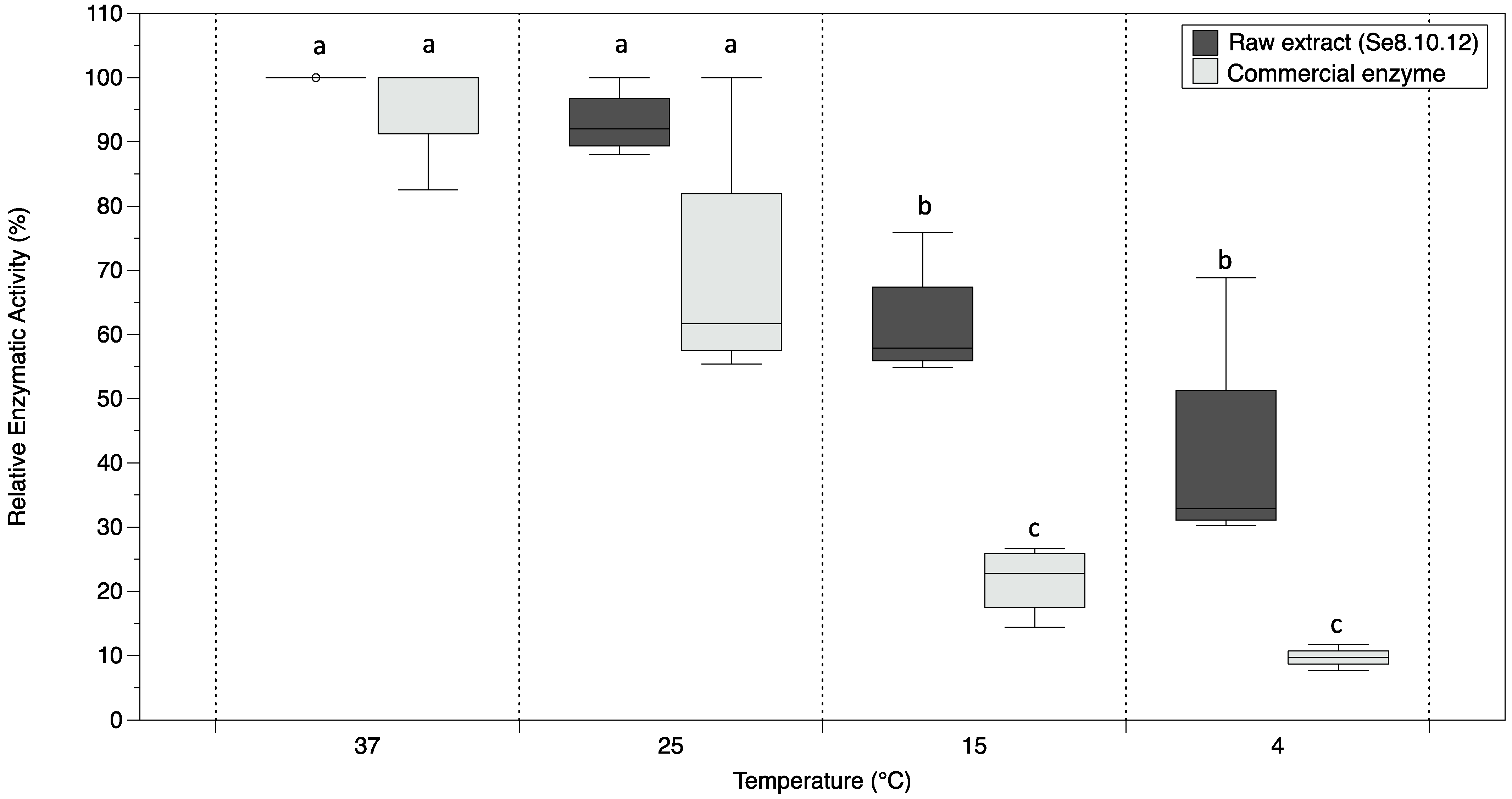
2.2. Antarctic Strain Se8.10.12 Showed Cold-Stable β-galactosidase Activity
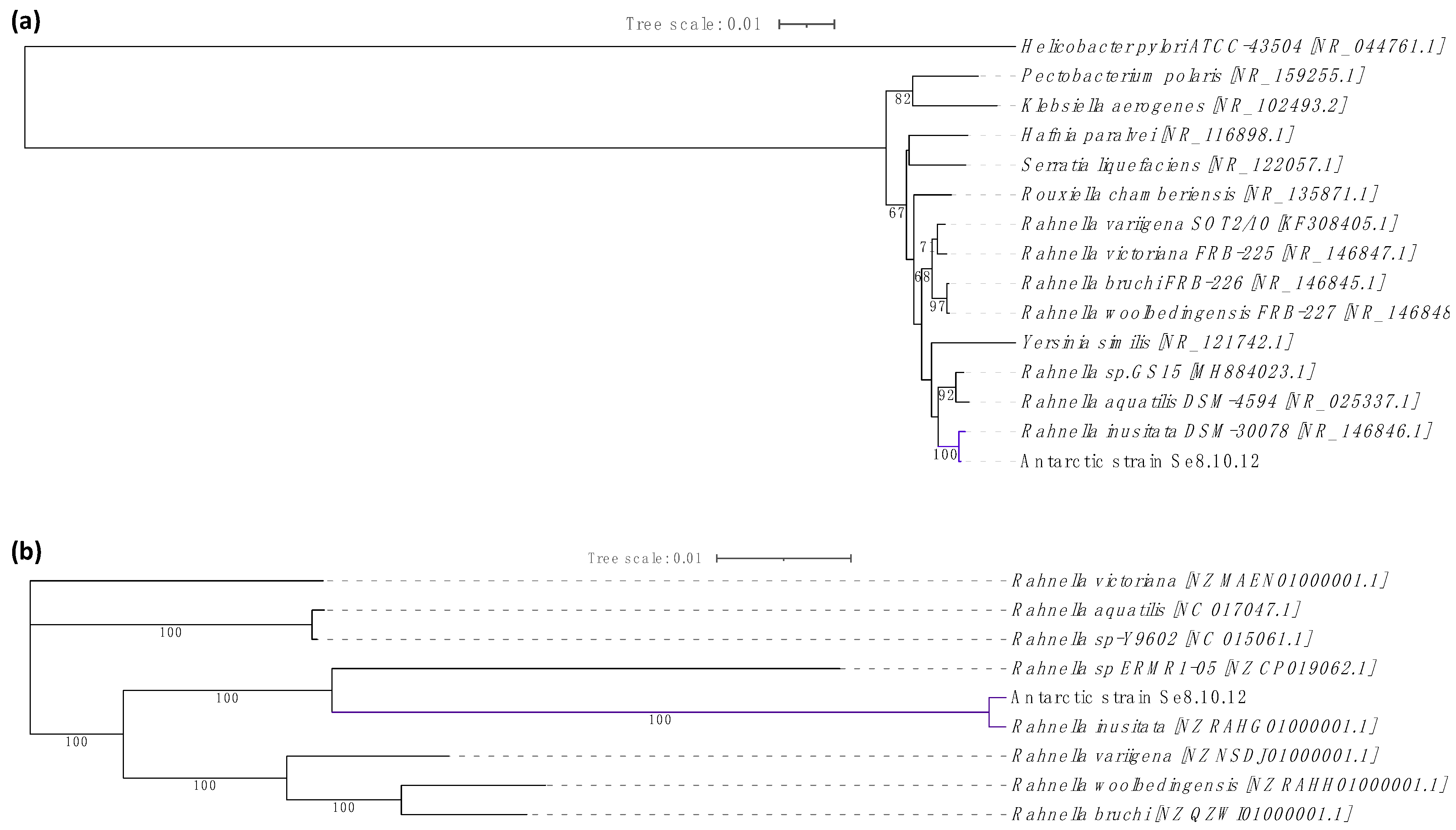
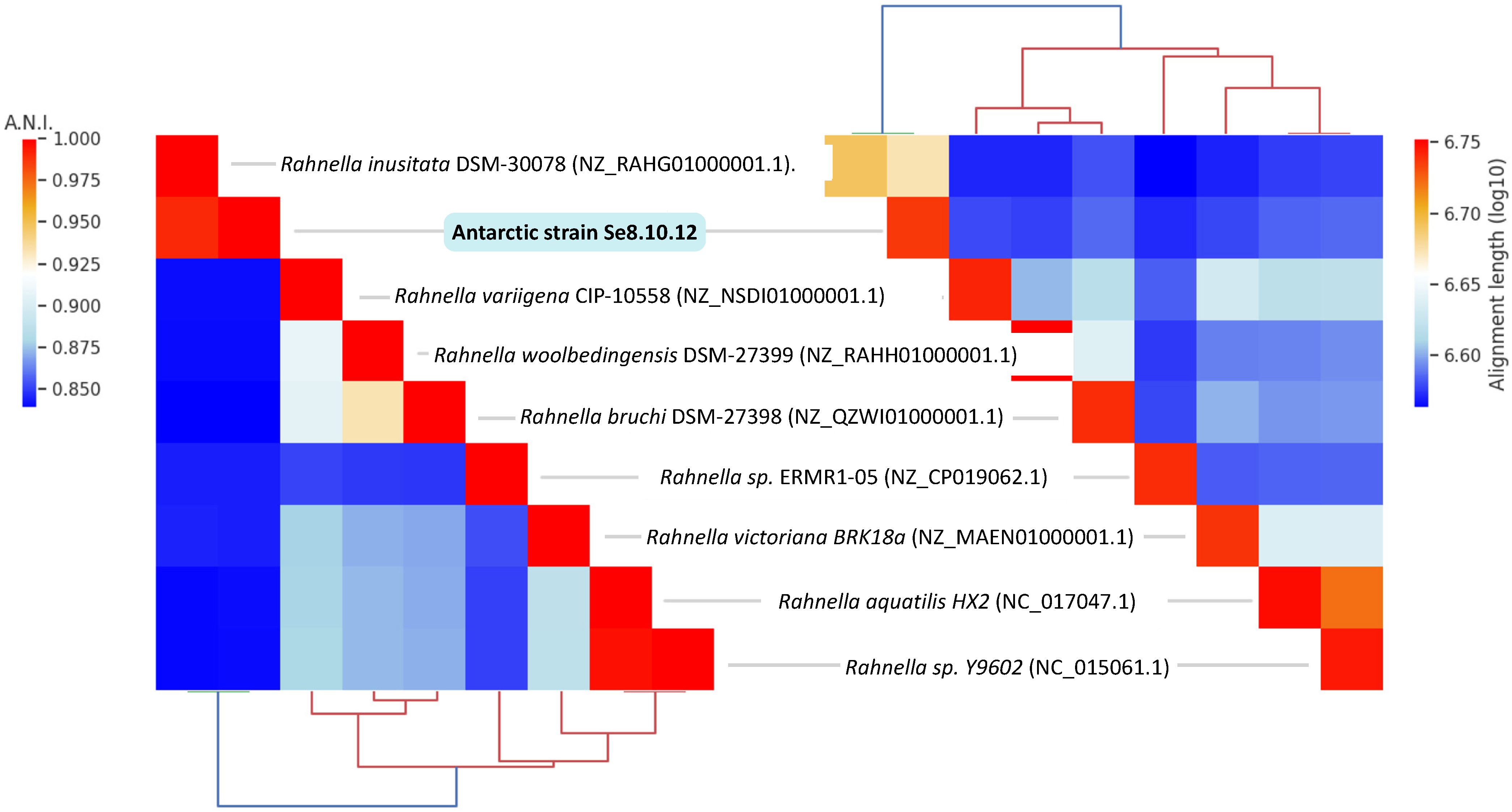
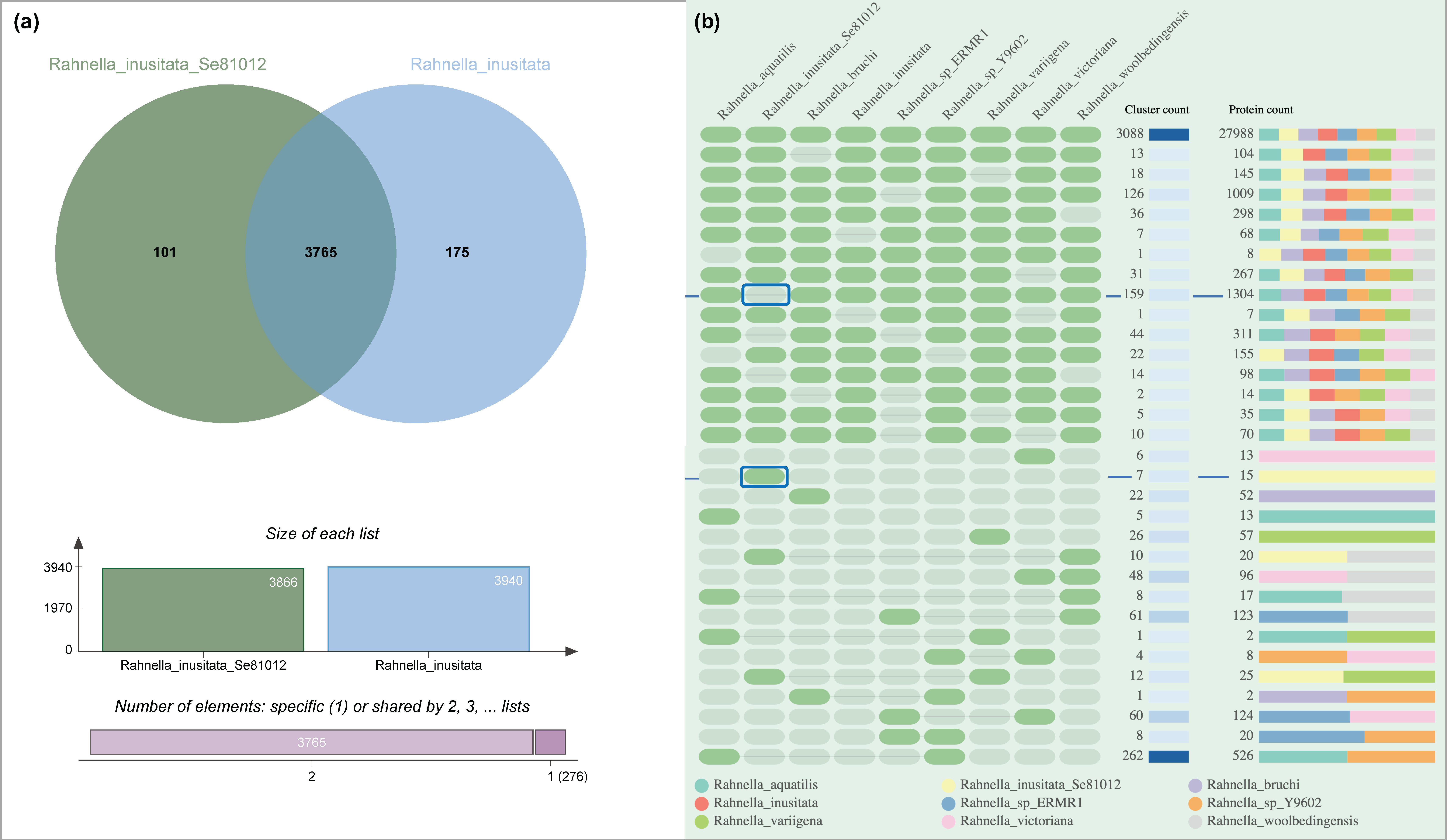
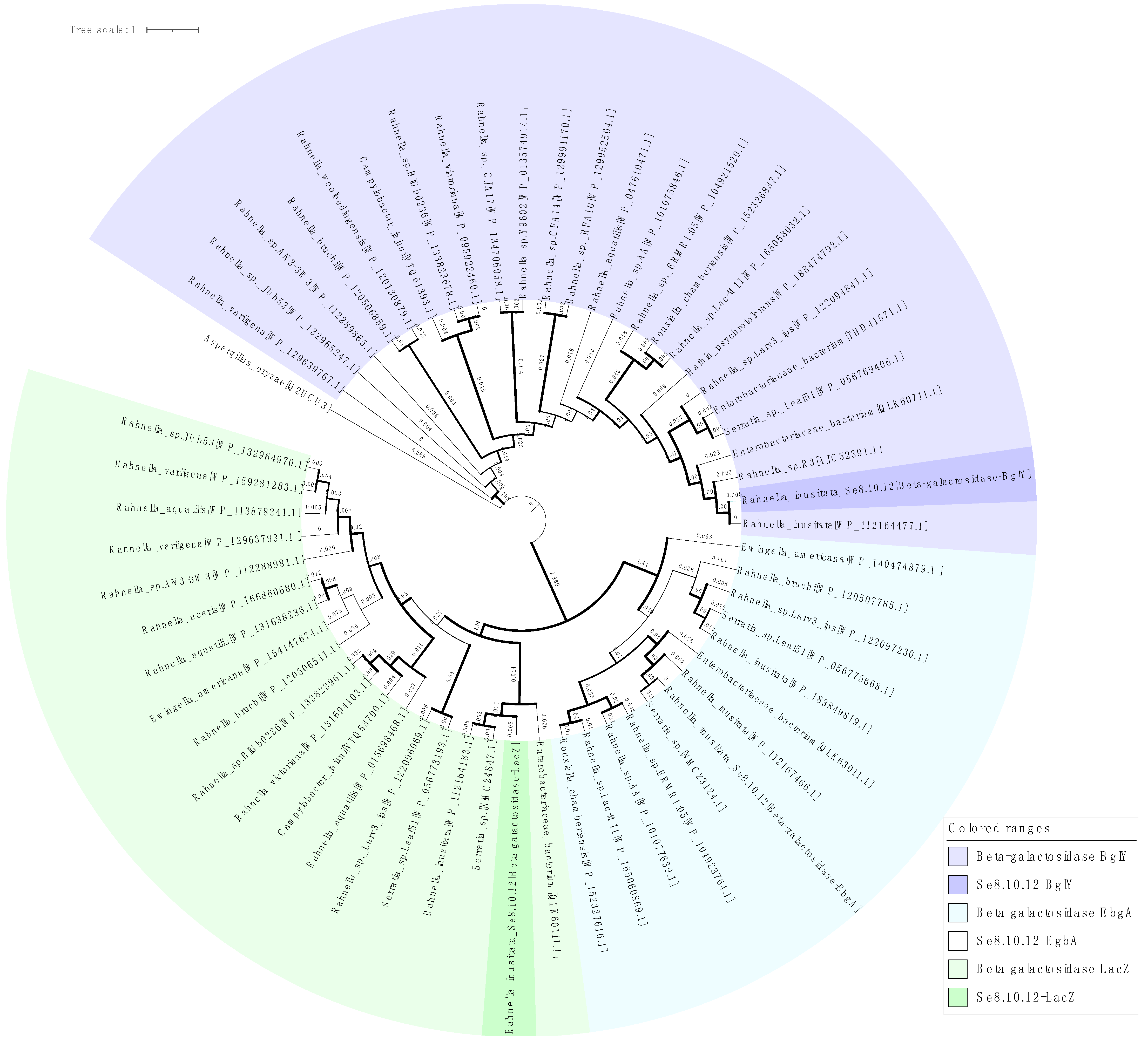
2.3. Characterization of Strain Se8.10.12 Revealed an Antarctic Rahnella inusitata with Unique Metal-Resistance Related Genes among the Rahnella Species
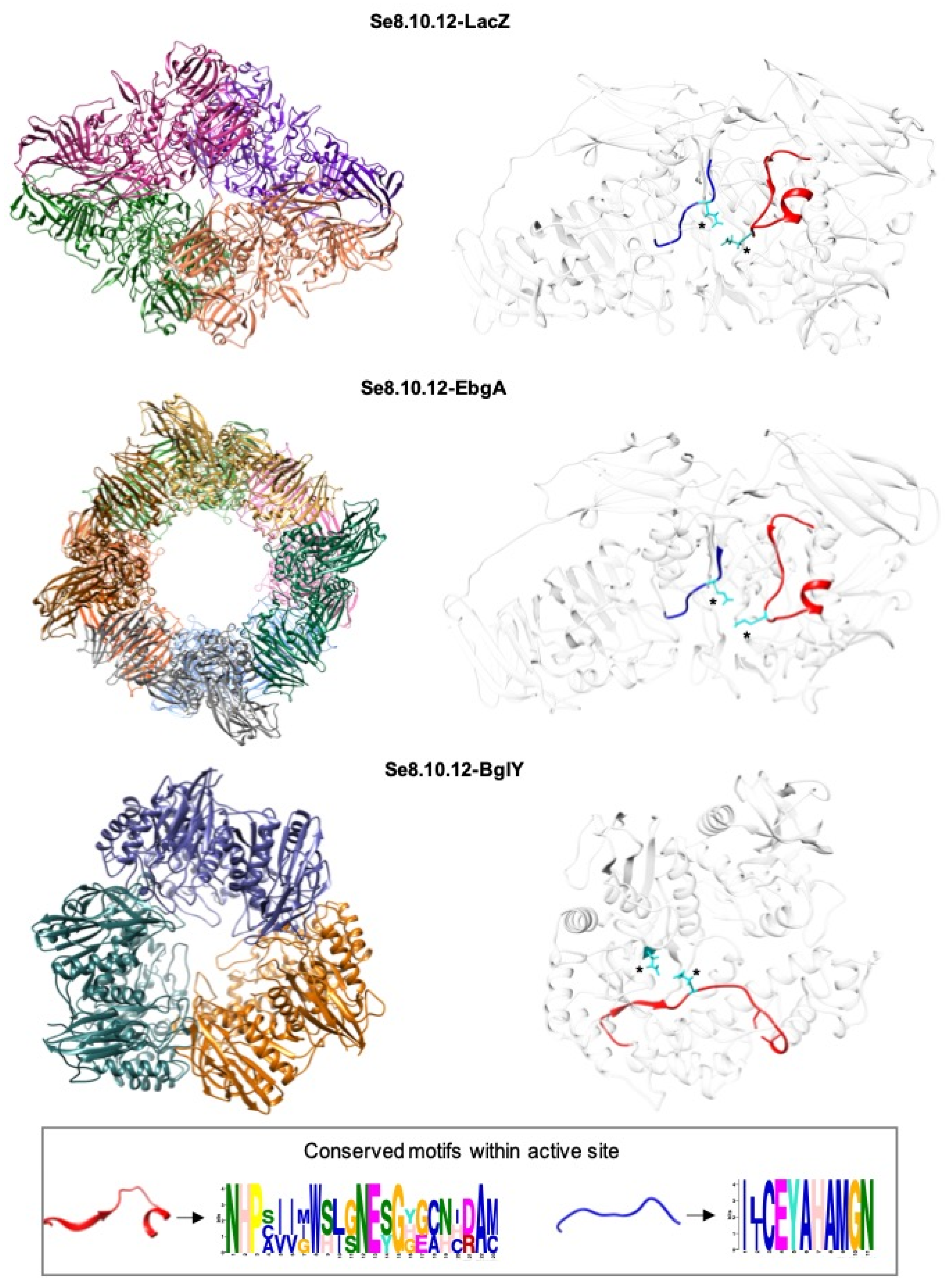
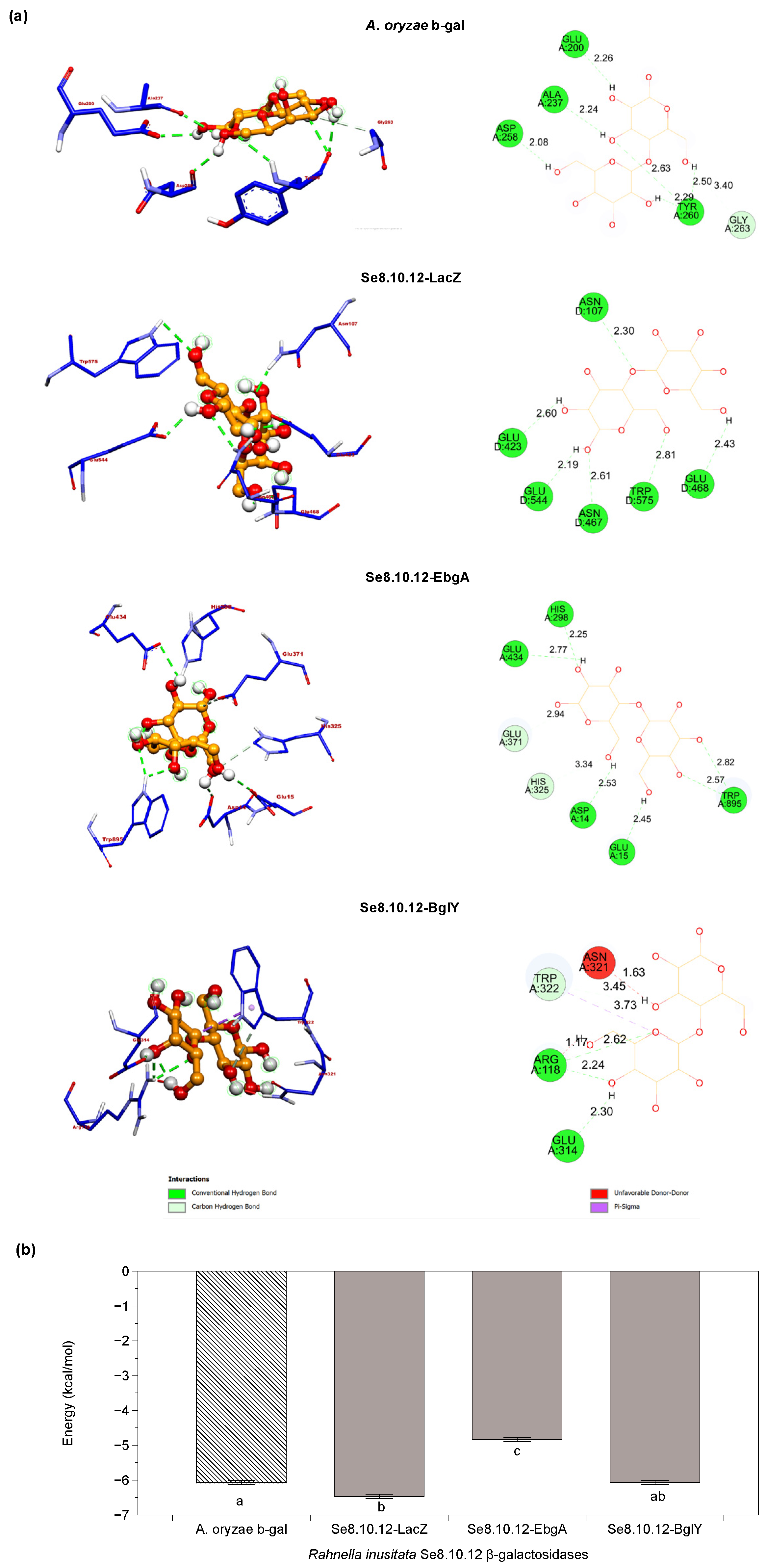
2.4. Rahnella inusitata Se8.10.12 Genome Harbors Three β-Galactosidase Genes, In Silico Molecular Docking Showed That Two of Them Have a Higher Predicted Affinity for Lactose Than the Commercial Enzyme
3. Materials and Methods
3.1. Biological Material
3.2. Screening of β-Galactosidase Activity in Antarctic Bacteria
3.3. Molecular Identification of β-Galactosidase Producing Antarctic Strains
3.4. β-Galactosidase Activity Quantification
3.5. Morphological and Biochemical Characterization of Strain Se8.10.12
3.6. Determination of the Effect of Temperature on the β-Galactosidase Relative Activity of Strain Se8.10.12
3.7. Whole-Genome Sequencing and Assembly
3.8. Genome-Wide Analysis for Taxonomic Identification and Characterization
3.9. Characterization of β-Galactosidase Enzymes of the Strain Se8.10.12 Based on Genomic Data
3.10. Statistic Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vásquez-Ponce, F.; Higuera-Llantén, S.; Pavlov, M.S.; Marshall, S.H.; Olivares-Pacheco, J. Phylogenetic MLSA and phenotypic analysis identification of three probable novel Pseudomonas species isolated on King George Island, South Shetland, Antarctica. Braz. J. Microbiol. 2018, 49, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Cid, F.P.; Maruyama, F.; Murase, K.; Graether, S.P.; Larama, G.; Bravo, L.A.; Jorquera, M.A. Draft genome sequences of bacteria isolated from the Deschampsia antarctica phyllosphere. Extremophiles 2018, 22, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Montero, K.; Barrientos, L. Advances in Antarctic Research for Antimicrobial Discovery: A Comprehensive Narrative Review of Bacteria from Antarctic Environments as Potential Sources of Novel Antibiotic Compounds Against Human Pathogens and Microorganisms of Industrial Importance. Antibiotics 2018, 7, 90. [Google Scholar] [CrossRef]
- Ferrer, M.; Golyshina, O.; Beloqui, A.; Golyshin, P.N. Mining enzymes from extreme environments. Curr. Opin. Microbiol. 2007, 10, 207–214. [Google Scholar] [CrossRef]
- Nichols, D.; Bowman, J.; Sanderson, K.; Nichols, C.M.; Lewis, T.; McMeekin, T.; Nichols, P.D. Developments with Antarctic microorganisms: Culture collections, bioactivity screening, taxonomy, PUFA production and cold-adapted enzymes. Curr. Opin. Biotechnol. 1999, 10, 240–246. [Google Scholar] [CrossRef]
- Antony, R.; Sanyal, A.; Kapse, N.; Dhakephalkar, P.K.; Thamban, M.; Nair, S. Microbial communities associated with Antarctic snow pack and their biogeochemical implications. Microbiol. Res. 2016, 192, 192–202. [Google Scholar] [CrossRef]
- Karasová-Lipovová, P.; Strnad, H.; Spiwok, V.; Malá, Š.; Králová, B.; Russell, N.J. The cloning, purification and characterisation of a cold-active β-galactosidase from the psychrotolerant Antarctic bacterium Arthrobacter sp. C2-2. Enzyme Microb. Technol. 2003, 6, 836–844. [Google Scholar] [CrossRef]
- Itan, Y.; Jones, B.L.; Ingram, C.J.; Swallow, D.M.; Thomas, M.G. A worldwide correlation of lactase persistence phenotype and genotypes. BMC Evol. Biol. 2010, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Vesa, T.H.; Korpela, R.; Marteau, P. Lactose Intolerance. J. Am. Coll. Nutr. 2000, 9, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, P.; Wanarska, M.; Kur, J. A new cold-adapted β-D-galactosidase from the Antarctic Arthrobacter sp. 32c—Gene cloning, overexpression, purification and properties. BMC Microbiol. 2009, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Yi, J.; Hua, X.; Zhang, Y.; Yang, R. Preparation and characterization of gellan gum microspheres containing a cold-adapted β-galactosidase from Rahnella sp. R3. Carbohydr. Polym. 2017, 162, 10–15. [Google Scholar] [CrossRef]
- Margesin, R.; Schinner, F. (Eds.) Cold-Adapted Organisms: Ecology, Physiology, Enzymology; Springer: Berlin/Heidelberg, Germany, 1999; ISBN 9788578110796. [Google Scholar]
- Fan, Y.; Hua, X.; Zhang, Y.; Feng, Y.; Shen, Q.; Dong, J.; Zhao, W.; Zhang, W.; Jin, Z.; Yang, R. Cloning, expression and structural stability of a cold-adapted β-galactosidase from Rahnella sp. R3. Protein Expr. Purif. 2015, 115, 158–164. [Google Scholar] [CrossRef]
- Nakagawa, T.; Fujimoto, Y.; Uchino, M.; Miyaji, T.; Takano, K.; Tomizuka, N. Isolation and characterization of psychrophiles producing cold-active -galactosidase. Lett. Appl. Microbiol. 2003, 37, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.; Geueke, B.; Delgado, O.; Coleman, J.; Hatti-Kaul, R. β-Galactosidase from a cold-adapted bacterium: Purification, characterization and application for lactose hydrolysis. Appl. Microbiol. Biotechnol. 2002, 58, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Turkiewicz, M.; Kur, J.; Białkowska, A.; Cieśliński, H.; Kalinowska, H.; Bielecki, S. Antarctic marine bacterium Pseudoalteromonas sp. 22b as a source of cold-adapted β-galactosidase. Biomol. Eng. 2003, 20, 317–324. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, K.; Mo, Z.; Liu, Y.; Hu, X. Crystallization and preliminary X-ray analysis of a cold-active β-galactosidase from the psychrotrophic and halotolerant Planococcus sp. L4. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 911–913. [Google Scholar] [CrossRef]
- Wierzbicka-Woś, A.; Cieśliński, H.; Wanarska, M.; Kozłowska-Tylingo, K.; Hildebrandt, P.; Kur, J. A novel cold-active β-D-galactosidase from the Paracoccus sp. 32d—gene cloning, purification and characterization. Microb. Cell Fact. 2011, 10, 1–12. [Google Scholar] [CrossRef]
- Alikunju, A.P.; Joy, S.; Salam, J.A.; Silvester, R.; Antony, A.C.; Rahiman, K.M.M.; Krishnan, K.P.; Hatha, A.A.M. Functional Characterization of a New Cold-Adapted β-Galactosidase from an Arctic Fjord Sediment Bacteria Enterobacter ludwigii MCC 3423. Catal. Lett. 2018, 148, 3223–3235. [Google Scholar] [CrossRef]
- Wang, G.X.; Gao, Y.; Hu, B.; Lu, X.L.; Liu, X.Y.; Jiao, B. hua A novel cold-adapted β-galactosidase isolated from Halomonas sp. S62: Gene cloning, purification and enzymatic characterization. World J. Microbiol. Biotechnol. 2013, 29, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Stougaard, P. Identification, cloning and expression of a cold-active β-galactosidase from a novel Arctic bacterium, Alkalilactibacillus ikkense. Environ. Technol. 2010, 31, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Mangiagalli, M.; Lapi, M.; Maione, S.; Orlando, M.; Brocca, S.; Pesce, A.; Barbiroli, A.; Camilloni, C.; Pucciarelli, S.; Lotti, M.; et al. The co-existence of cold activity and thermal stability in an Antarctic GH42 β-galactosidase relies on its hexameric quaternary arrangement. FEBS J. 2020, 288, 546–565. [Google Scholar] [CrossRef] [PubMed]
- Saqib, S.; Akram, A.; Halim, S.A.; Tassaduq, R. Sources of β-galactosidase and its applications in food industry. 3 Biotech 2017, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Groudieva, T.; Kambourova, M.; Yusef, H.; Royter, M.; Grote, R.; Trinks, H.; Antranikian, G. Diversity and cold-active hydrolytic enzymes of culturable bacteria associated with Arctic sea ice, Spitzbergen. Extremophiles 2004, 8, 475–488. [Google Scholar] [CrossRef]
- Vincent, V.; Aghajari, N.; Pollet, N.; Boisson, A.; Boudebbouze, S.; Haser, R.; Maguin, E.; Rhimi, M. The acid tolerant and cold-active β-galactosidase from Lactococcus lactis strain is an attractive biocatalyst for lactose hydrolysis. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2013, 103, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.-H.; Splechtna, B.; Krasteva, S.; Kneifel, W.; Kulbe, K.D.; Divne, C.; Haltrich, D. Characterization and molecular cloning of a heterodimeric Î2-galactosidase from the probiotic strain Lactobacillus acidophilus R22. FEMS Microbiol. Lett. 2007, 269, 136–144. [Google Scholar] [CrossRef]
- Rhimi, M.; Boisson, A.; Dejob, M.; Boudebouze, S.; Maguin, E.; Haser, R.; Aghajari, N. Efficient bioconversion of lactose in milk and whey: Immobilization and biochemical characterization of a β-galactosidase from the dairy Streptococcus thermophilus LMD9 strain. Res. Microbiol. 2010, 161, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Rhimi, M.; Aghajari, N.; Jaouadi, B.; Juy, M.; Boudebbouze, S.; Maguin, E.; Haser, R.; Bejar, S. Exploring the acidotolerance of β-galactosidase from Lactobacillus delbrueckii subsp. bulgaricus: An attractive enzyme for lactose bioconversion. Res. Microbiol. 2009, 160, 775–784. [Google Scholar] [CrossRef]
- Peeters, K.; Verleyen, E.; Hodgson, D.A.; Convey, P.; Ertz, D.; Vyverman, W.; Willems, A. Heterotrophic bacterial diversity in aquatic microbial mat communities from Antarctica. Polar Biol. 2012, 35, 543–554. [Google Scholar] [CrossRef]
- Tashyrev, O.; Romanovskaya, V.; Rokitko, P.; Tashyreva, H.; Prytula, I.; Suslova, O.; Govorukha, V.; Prekrasna, I.; Gladka, G. Autecology and Taxonomy of Bacteria Isolated from Extreme Environments. Mikrobiol. Zh. 2017, 79, 100–113. [Google Scholar] [CrossRef]
- Park, J.W.; Oh, Y.S.; Lim, J.Y.; Roh, D.H. Isolation and characterization of cold-adapted strains producing β-galactosidase. J. Microbiol. 2006, 44, 396–402. [Google Scholar]
- Liu, W.Y.; Shi, Y.W.; Wang, X.Q.; Wang, Y.; Wei, C.Q.; Lou, K. Isolation and identification of a strain producing cold-adapted β-galactosidase, and purification and characterisation of the enzyme. Czech J. Food Sci. 2008, 26, 284–290. [Google Scholar] [CrossRef]
- Mahdian, S.M.A.; Karimi, E.; Tanipour, M.H.; Parizadeh, S.M.R.; Ghayour-Mobarhan, M.; Bazaz, M.M.; Mashkani, B. Expression of a functional cold active β-galactosidase from Planococcus sp-L4 in Pichia pastoris. Protein Expr. Purif. 2016, 125, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Coker, J.A.; Sheridan, P.P.; Loveland-Curtze, J.; Gutshall, K.R.; Auman, A.J.; Brenchley, J.E. Biochemical characterization of a β-galactosidase with a low temperature optimum obtained from an Antarctic Arthrobacter isolate. J. Bacteriol. 2003, 185, 5473–5482. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, P.P.; Brenchley, J.E. Characterization of a salt-tolerant family 42 β-galactosidase from a psychrophilic antarctic Planococcus isolate. Appl. Environ. Microbiol. 2000, 66, 2438–2444. [Google Scholar] [CrossRef]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [CrossRef]
- Brady, C.; Hunter, G.; Kirk, S.; Arnold, D.; Denman, S. Rahnella victoriana sp. nov., Rahnella bruchi sp. nov., Rahnella woolbedingensis sp. nov., classification of Rahnella genomospecies 2 and 3 as Rahnella variigena sp. nov. and Rahnella inusitata sp. nov., respectively and emended description of the genus Rahnella. Syst. Appl. Microbiol. 2014, 37, 545–552. [Google Scholar] [CrossRef]
- Izard, D.; Gavini, F.; Trinel, P.A.; Leclerc, H. “Rahnella aquatilis”, a new member of the “Enterobacteriaceae”. Ann. Microbiol. (Paris) 1979, 130, 163–177. [Google Scholar] [PubMed]
- Hugoson, E.; Lam, W.T.; Guy, L. miComplete: Weighted quality evaluation of assembled microbial genomes. Bioinformatics 2019. [Google Scholar] [CrossRef]
- Santos, I.R.; Silva-Filho, E.V.; Schaefer, C.E.G.R.; Albuquerque-Filho, M.R.; Campos, L.S. Heavy metal contamination in coastal sediments and soils near the Brazilian Antarctic Station, King George Island. Mar. Pollut. Bull. 2005, 50, 185–194. [Google Scholar] [CrossRef]
- Chu, W.L.; Dang, N.L.; Kok, Y.Y.; Ivan Yap, K.S.; Phang, S.M.; Convey, P. Heavy metal pollution in Antarctica and its potential impacts on algae. Polar Sci. 2019, 20, 75–83. [Google Scholar] [CrossRef]
- Marina-Montes, C.; Pérez-Arribas, L.V.; Escudero, M.; Anzano, J.; Cáceres, J.O. Heavy metal transport and evolution of atmospheric aerosols in the Antarctic region. Sci. Total Environ. 2020, 721, 137702. [Google Scholar] [CrossRef] [PubMed]
- Romaniuk, K.; Ciok, A.; Decewicz, P.; Uhrynowski, W.; Budzik, K.; Nieckarz, M.; Pawlowska, J.; Zdanowski, M.K.; Bartosik, D.; Dziewit, L. Insight into heavy metal resistome of soil psychrotolerant bacteria originating from King George Island (Antarctica). Polar Biol. 2018, 41, 1319–1333. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Yang, P.; Wang, Y.; Meng, K.; Luo, H.; Zhang, W.; Wu, N.; Fan, Y.; Yao, B. Heterologous expression of a gene encoding a thermostable β-galactosidase from Alicyclobacillus acidocaldarius. Biotechnol. Lett. 2008, 30, 343–348. [Google Scholar] [CrossRef]
- Coker, J.A.; Brenchley, J.E. Protein engineering of a cold-active β-galactosidase from Arthrobacter sp. SB to increase lactose hydrolysis reveals new sites affecting low temperature activity. Extremophiles 2006, 10, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Molina, F.; Simancas, A.; Tabla, R.; Roa, I.; Rebollodo, E. Horizontal gene transfer and the evolving definition of coliforms. In Microbes in the Spotlight: Recent Progress in the Understanding of Beneficial and Harmful Microorganisms; BrownWalker Press: Boca Raton, FL, USA, 2016; p. 496. ISBN 978-1-62734-612-2. [Google Scholar]
- Stokes, H.W.; Betts, P.W.; Hall, B.G. Sequence of the ebgA gene of Escherichia coli: Comparison with the lacZ gene. Mol. Biol. Evol. 1985, 2, 469–477. [Google Scholar] [CrossRef]
- Hall, B.G. Evolutionary potential of the ebgA gene. Mol. Biol. Evol. 1995, 12, 514–517. [Google Scholar] [CrossRef]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef]
- Karan, R.; Mathew, S.; Muhammad, R.; Bautista, D.B.; Vogler, M.; Eppinger, J.; Oliva, R.; Cavallo, L.; Arold, S.T.; Rueping, M. Understanding high-salt and cold adaptation of a polyextremophilic enzyme. Microorganisms 2020, 8, 1594. [Google Scholar] [CrossRef]
- Juers, D.H.; Heightman, T.D.; Vasella, A.; McCarter, J.D.; Mackenzie, L.; Withers, S.G.; Matthews, B.W. A structural view of the action of Escherichia coli (lacZ) β-galactosidase. Biochemistry 2001, 40, 14781–14794. [Google Scholar] [CrossRef]
- Juers, D.H.; Matthews, B.W.; Huber, R.E. LacZ β-galactosidase: Structure and function of an enzyme of historical and molecular biological importance. Protein Sci. 2012, 21, 1792–1807. [Google Scholar] [CrossRef] [PubMed]
- Hoyoux, A.; Jennes, I.; Dubois, P.; Genicot, S.; Dubail, F.; François, J.M.; Baise, E.; Feller, G.; Gerday, C. Cold-Adapted β-Galactosidase from the Antarctic Psychrophile Pseudoalteromonas haloplanktis. Appl. Environ. Microbiol. 2001, 67, 1529–1535. [Google Scholar] [CrossRef]
- Cieśliński, H.; Kur, J.; Białkowska, A.; Baran, I.; Makowski, K.; Turkiewicz, M. Cloning, expression, and purification of a recombinant cold-adapted β-galactosidase from antarctic bacterium Pseudoalteromonas sp. 22b. Protein Expr. Purif. 2005, 39, 27–34. [Google Scholar] [CrossRef]
- Yao, C.; Sun, J.; Wang, W.; Zhuang, Z.; Liu, J.; Hao, J. A novel cold-adapted β-galactosidase from Alteromonas sp. ML117 cleaves milk lactose effectively at low temperature. Process Biochem. 2019, 82, 94–101. [Google Scholar] [CrossRef]
- Douillard, F.P.; Kant, R.; Ritari, J.; Paulin, L.; Palva, A.; De Vos, W.M. Comparative genome analysis of Lactobacillus casei strains isolated from Actimel and Yakult products reveals marked similarities and points to a common origin. Microb. Biotechnol. 2013, 6, 576–587. [Google Scholar] [CrossRef] [PubMed]
- Míguez Amil, S.; Jiménez-Ortega, E.; Ramírez-Escudero, M.; Talens-Perales, D.; Marín-Navarro, J.; Polaina, J.; Sanz-Aparicio, J.; Fernandez-Leiro, R. The cryo-EM Structure of Thermotoga maritima β-Galactosidase: Quaternary Structure Guides Protein Engineering. ACS Chem. Biol. 2020, 15, 179–188. [Google Scholar] [CrossRef]
- Vukić, V.; Hrnjez, D.; Milanović, S.; Iličić, M.; Kanurić, K.; Petri, E. Comparative Molecular Modeling and Docking Analysis of β-galactosidase Enzymes from Commercially Important Starter Cultures Used in the Dairy Industry. Food Biotechnol. 2015, 28, 248–262. [Google Scholar] [CrossRef]
- Naumoff, D.G. Hierarchical classification of glycoside hydrolases. Biochem. 2011, 76, 622–635. [Google Scholar] [CrossRef]
- Davies, G.; Henrissat, B. Structures and mechanisms of glycosyl hydrolases. Structure 1995, 3, 853–859. [Google Scholar] [CrossRef]
- Li, S.; Zhu, X.; Xing, M. A new β-galactosidase from the antarctic bacterium Alteromonas sp. ANT48 and its potential in formation of prebiotic galacto-oligosaccharides. Mar. Drugs 2019, 17, 599. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Sun, J.; Tang, Y.J.; Xie, J.; Wei, D. Expression, characterization and structural profile of a heterodimeric β-galactosidase from the novel strain Lactobacillus curieae M2011381. Process Biochem. 2020, 97, 87–95. [Google Scholar] [CrossRef]
- Lam, K.K.; Labutti, K.; Khalak, A.; Tse, D. FinisherSC: A repeat-aware tool for upgrading de novo assembly using long reads. Bioinformatics 2015, 31, 3207–3209. [Google Scholar] [CrossRef]
- Chakraborty, M.; Baldwin-Brown, J.G.; Long, A.D.; Emerson, J.J. Contiguous and accurate de novo assembly of metazoan genomes with modest long read coverage. Nucleic Acids Res. 2016, 44, e147. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Avram, O.; Rapoport, D.; Portugez, S.; Pupko, T. M1CR0B1AL1Z3R—A user-friendly web server for the analysis of large-scale microbial genomics data. Nucleic Acids Res. 2019, 47, W88–W92. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017. [Google Scholar] [CrossRef] [PubMed]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Pritchard, L.; Glover, R.H.; Humphris, S.; Elphinstone, J.G.; Toth, I.K. Genomics and taxonomy in diagnostics for food security: Soft-rotting enterobacterial plant pathogens. Anal. Methods 2016, 8, 12–24. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A web server for whole-genome comparison and annotation of orthologous clusters across multiple species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef]
- Li, L.; Stoeckert, C.J.; Roos, D.S. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2014, 12, 59–60. [Google Scholar] [CrossRef]
- Dongen, S. Van A cluster algorithm for graphs. Inf. Syst. [INS] 2000, R0010, 40. [Google Scholar]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2004, 32, 2699. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Bayliss, S.; Thorpe, H.; Coyle, N.; Sheppard, S.; Feil, E. PIRATE: A fast and scalable pangenomics toolbox for clustering diverged orthologues in bacteria. bioRxiv 2019. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Krieger, E.; Joo, K.; Lee, J.; Lee, J.; Raman, S.; Thompson, J.; Tyka, M.; Baker, D.; Karplus, K. Improving physical realism, stereochemistry, and side-chain accuracy in homology modeling: Four approaches that performed well in CASP8. Proteins Struct. Funct. Bioinform. 2009, 77 (Suppl. 9), 114–122. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Rullmann, J.A.C.; MacArthur, M.W.; Kaptein, R.; Thornton, J.M. AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 1996, 8, 477–486. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 15, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Schüttelkopf, A.W.; Van Aalten, D.M.F. PRODRG: A tool for high-throughput crystallography of protein-ligand complexes. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, D60, 1355–1363. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 16, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Dekker, P.J.T.; Koenders, D.; Bruins, M.J. Lactose-free dairy products: Market developments, production, nutrition and health benefits. Nutrients 2019, 11, 551. [Google Scholar] [CrossRef] [PubMed]
- Biovia, D.S.; Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E.; Darden, T.; et al. Dassault Systèmes BIOVIA, Discovery Studio Visualizer, v.17.2, San Diego: Dassault Systèmes, 2016. J. Chem. Phys. 2000, 10, 21–9991. [Google Scholar] [CrossRef]
- JASP Team. JASP, version 0.13.1; Computer software; University of Amsterdam: Amsterdam, The Netherlands, 2020. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster-ID | SwissProt ID: Protein, Function | GO Annotation | Genome|Protein |
|---|---|---|---|
| 5124 | P0CE53: transposase InsH, involved in the transposition of the insertion sequence IS5 | GO:0006313; P:transposition, DNA-mediated | Rahnella_inusitata_Se81012|AJMHLFKD_00752 Rahnella_inusitata_Se81012|AJMHLFKD_00297 Rahnella_inusitata_Se81012|AJMHLFKD_03605 |
| 6148 | Q47459: protein PcoE, Required for the copper-inducible expression of copper resistance | GO:0046872; F: metal ion binding | Rahnella_inusitata_Se81012|AJMHLFKD_00787 Rahnella_inusitata_Se81012|AJMHLFKD_00806 |
| 6151 | Q9ZHD4: sensor kinase SilS, component of the soil cation-efflux system that confers resistance to silver | GO:0000155; F:phospho-relay sensor kinase activity | Rahnella_inusitata_Se81012|AJMHLFKD_00805 Rahnella_inusitata_Se81012|AJMHLFKD_00788 |
| 6149 | N/A | N/A | Rahnella_inusitata_Se81012|AJMHLFKD_02409 Rahnella_inusitata_Se81012|AJMHLFKD_03172 |
| 6147 | N/A | N/A | Rahnella_inusitata_Se81012|AJMHLFKD_02421 Rahnella_inusitata_Se81012|AJMHLFKD_03161 |
| 6146 | P76514: uncharacterized protein YfdR, probable cobalt ion binding | GO:0050897; F: cobalt ion binding | Rahnella_inusitata_Se81012|AJMHLFKD_03184 Rahnella_inusitata_Se81012|AJMHLFKD_02399 |
| 6150 | Q9XJJ6: Spanin inner membrane subunit, participates in cell lysis during virus exit | GO:0019076; P: viral release from the host cell | Rahnella_inusitata_Se81012|AJMHLFKD_03641 Rahnella_inusitata_Se81012|AJMHLFKD_03102 |
| 5653 | P41059: uncharacterized protein in gpA, probable zinc ion binding | GO:0008270; F: zinc ion binding | Rahnella_sp_ERMR1|LLFBIOOH_04925 Rahnella_inusitata_Se81012|AJMHLFKD_02768 |
| 4804 | P74068: uncharacterized transporter sll1263, cadmium, iron, and zinc transmembrane transport | GO:0015341; F: zinc efflux active transmembrane transporter activity | Rahnella_aquatilis|AEJLJCIE_02970 Rahnella_inusitata_Se81012AJMHLFKD_00295 Rahnella_sp_Y9602|CHBNGHAC_02899 |
| Organism | Protein | Sequence | Molecular Mass (Da) | pI | Glycoside Hydrolase Family | PDB Number | % Identity | ||
|---|---|---|---|---|---|---|---|---|---|
| Lac-Z | BglY | EbgA | |||||||
| Rahnella inusitata Se.8.10.12 | Lac-Z | 1032 | 116,913.82 | 5.30 | GH 2 | - | 100 | 29.79 | 34.10 |
| Rahnella inusitata Se.8.10.12 | BglY | 687 | 77,230.21 | 5.84 | GH 42 | - | 29.79 | 100 | 40.91 |
| Rahnella inusitata Se.8.10.12 | EbgA | 1023 | 116,917.20 | 5.20 | GH 2 | - | 34.10 | 40.91 | 100 |
| Rahnella sp. R3 | β-gal | 712 | 80,188.46 | 5.98 | GH 42 | 5E9A_1 | 31.82 | 99.13 | 36.84 |
| Escherichia coli | Lac-Z | 1023 | 116,385.64 | 5.28 | GH 2 | 1jz2.1.D | 68.9 | 36.54 | 34.51 |
| Thermotoga maritima | β-gal | 1083 | 127,476.78 | 5.52 | GH 2 | 6S6Z_A | 36.39 | 29.27 | 37.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Núñez-Montero, K.; Salazar, R.; Santos, A.; Gómez-Espinoza, O.; Farah, S.; Troncoso, C.; Hoffmann, C.; Melivilu, D.; Scott, F.; Barrientos Díaz, L. Antarctic Rahnella inusitata: A Producer of Cold-Stable β-Galactosidase Enzymes. Int. J. Mol. Sci. 2021, 22, 4144. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084144
Núñez-Montero K, Salazar R, Santos A, Gómez-Espinoza O, Farah S, Troncoso C, Hoffmann C, Melivilu D, Scott F, Barrientos Díaz L. Antarctic Rahnella inusitata: A Producer of Cold-Stable β-Galactosidase Enzymes. International Journal of Molecular Sciences. 2021; 22(8):4144. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084144
Chicago/Turabian StyleNúñez-Montero, Kattia, Rodrigo Salazar, Andrés Santos, Olman Gómez-Espinoza, Scandar Farah, Claudia Troncoso, Catalina Hoffmann, Damaris Melivilu, Felipe Scott, and Leticia Barrientos Díaz. 2021. "Antarctic Rahnella inusitata: A Producer of Cold-Stable β-Galactosidase Enzymes" International Journal of Molecular Sciences 22, no. 8: 4144. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084144