
Atypical p38 Signaling, Activation, and Implications for Disease

 , and
, and
Abstract
:1. Introduction
2. Classical Activation of Mitogen-Activated Protein Kinases (MAPK)
2.1. Activation of p38 by MAPKK3 and MAPKK6
2.2. Distribution, Activation, and Function of the p38 Isoforms
2.3. P38 Substrate Activation
2.4. Signal Termination
2.5. Molecular Inhibition
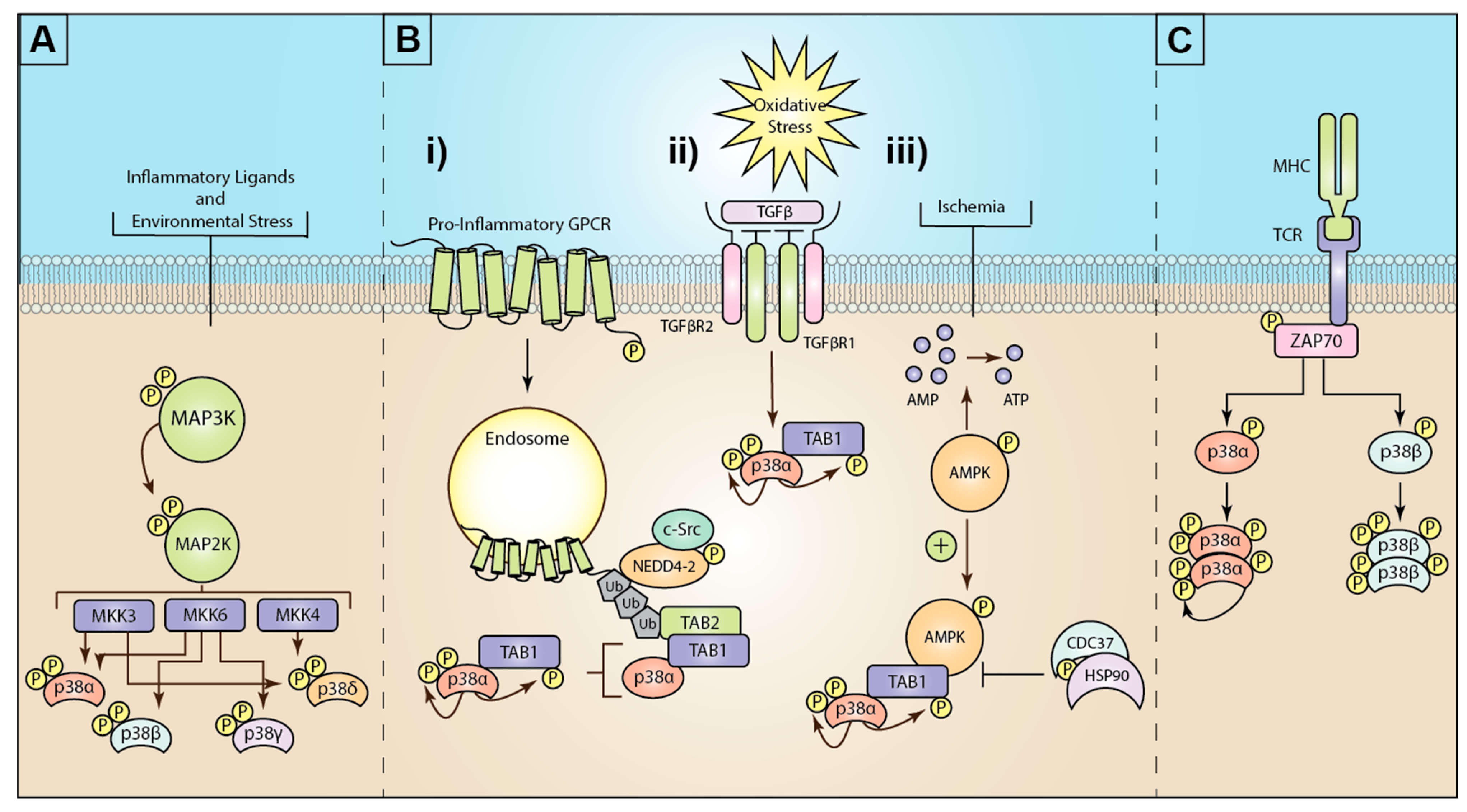
3. Mechanisms of Atypical p38 Activation
4. Activation of Atypical p38 by GPCRs
5. Pathophysiological Implications of MKK3/6-Dependent p38 MAPKs
6. Pathophysiological Implications of Atypical p38 Signaling
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, J.; Lee, J.D.; Bibbs, L.; Ulevitch, R.J. A MAP kinase targeted by endotoxin and hyperosmolarity in mammalian cells. Science 1994, 265, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, T.E., Jr.; Bradford, W.Z.; Castro-Bernardini, S.; Fagan, E.A.; Glaspole, I.; Glassberg, M.K.; Gorina, E.; Hopkins, P.M.; Kardatzke, D.; Lancaster, L.; et al. A phase 3 trial of pirfenidone in patients with idiopathic pulmonary fibrosis. N. Engl. J. Med. 2014, 370, 2083–2092. [Google Scholar] [CrossRef] [Green Version]
- Valeyre, D.; Albera, C.; Bradford, W.Z.; Costabel, U.; King, T.E., Jr.; Leff, J.A.; Noble, P.W.; Sahn, S.A.; du Bois, R.M. Comprehensive assessment of the long-term safety of pirfenidone in patients with idiopathic pulmonary fibrosis. Respirology 2014, 19, 740–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, X.; Akella, R.; He, H.; Humphreys, J.M.; Tsutakawa, S.E.; Lee, S.J.; Tainer, J.A.; Cobb, M.H.; Goldsmith, E.J. The structure of the MAP2K MEK6 reveals an autoinhibitory dimer. Structure 2009, 17, 96–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuadrado, A.; Nebreda, A.R. Mechanisms and functions of p38 MAPK signalling. Biochem. J. 2010, 429, 403–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raingeaud, J.; Whitmarsh, A.J.; Barrett, T.; Derijard, B.; Davis, R.J. MKK3- and MKK6-regulated gene expression is mediated by the p38 mitogen-activated protein kinase signal transduction pathway. Mol. Cell. Biol. 1996, 16, 1247–1255. [Google Scholar] [CrossRef] [Green Version]
- Derijard, B.; Raingeaud, J.; Barrett, T.; Wu, I.H.; Han, J.; Ulevitch, R.J.; Davis, R.J. Independent human MAP-kinase signal transduction pathways defined by MEK and MKK isoforms. Science 1995, 267, 682–685. [Google Scholar] [CrossRef]
- Moriguchi, T.; Kuroyanagi, N.; Yamaguchi, K.; Gotoh, Y.; Irie, K.; Kano, T.; Shirakabe, K.; Muro, Y.; Shibuya, H.; Matsumoto, K.; et al. A novel kinase cascade mediated by mitogen-activated protein kinase kinase 6 and MKK3. J. Biol. Chem. 1996, 271, 13675–13679. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, N.; Kamanaka, M.; Enslen, H.; Dong, C.; Wysk, M.; Davis, R.J.; Flavell, R.A. Differential involvement of p38 mitogen-activated protein kinase kinases MKK3 and MKK6 in T-cell apoptosis. EMBO Rep. 2002, 3, 785–791. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.T.; Yang, D.D.; Wysk, M.; Gatti, E.; Mellman, I.; Davis, R.J.; Flavell, R.A. Defective IL-12 production in mitogen-activated protein (MAP) kinase kinase 3 (Mkk3)-deficient mice. EMBO J. 1999, 18, 1845–1857. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.; Minden, A.; Martinetto, H.; Claret, F.X.; Lange-Carter, C.; Mercurio, F.; Johnson, G.L.; Karin, M. Identification of a dual specificity kinase that activates the Jun kinases and p38-Mpk2. Science 1995, 268, 286–290. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Mei, Z.Q.; Wu, J.W.; Wang, Z.X. Enzymatic activity and substrate specificity of mitogen-activated protein kinase p38alpha in different phosphorylation states. J. Biol. Chem. 2008, 283, 26591–26601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brancho, D.; Tanaka, N.; Jaeschke, A.; Ventura, J.J.; Kelkar, N.; Tanaka, Y.; Kyuuma, M.; Takeshita, T.; Flavell, R.A.; Davis, R.J. Mechanism of p38 MAP kinase activation in vivo. Genes Dev. 2003, 17, 1969–1978. [Google Scholar] [CrossRef] [Green Version]
- Remy, G.; Risco, A.M.; Inesta-Vaquera, F.A.; Gonzalez-Teran, B.; Sabio, G.; Davis, R.J.; Cuenda, A. Differential activation of p38MAPK isoforms by MKK6 and MKK3. Cell Signal. 2010, 22, 660–667. [Google Scholar] [CrossRef]
- Marshall, C.J. Specificity of receptor tyrosine kinase signaling: Transient versus sustained extracellular signal-regulated kinase activation. Cell 1995, 80, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Irie, T.; Muta, T.; Takeshige, K. TAK1 mediates an activation signal from toll-like receptor(s) to nuclear factor-kappaB in lipopolysaccharide-stimulated macrophages. FEBS Lett. 2000, 467, 160–164. [Google Scholar] [CrossRef] [Green Version]
- Takaesu, G.; Surabhi, R.M.; Park, K.J.; Ninomiya-Tsuji, J.; Matsumoto, K.; Gaynor, R.B. TAK1 is critical for IkappaB kinase-mediated activation of the NF-kappaB pathway. J. Mol. Biol. 2003, 326, 105–115. [Google Scholar] [CrossRef]
- Brown, K.; Vial, S.C.; Dedi, N.; Long, J.M.; Dunster, N.J.; Cheetham, G.M. Structural basis for the interaction of TAK1 kinase with its activating protein TAB1. J. Mol. Biol. 2005, 354, 1013–1020. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.H.; Xiao, C.; Paschal, A.E.; Bailey, S.T.; Rao, P.; Hayden, M.S.; Lee, K.Y.; Bussey, C.; Steckel, M.; Tanaka, N.; et al. TAK1, but not TAB1 or TAB2, plays an essential role in multiple signaling pathways in vivo. Genes Dev. 2005, 19, 2668–2681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Chen, C.; Li, Z.; Guo, W.; Gegner, J.A.; Lin, S.; Han, J. Characterization of the structure and function of a new mitogen-activated protein kinase (p38beta). J. Biol. Chem. 1996, 271, 17920–17926. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Gram, H.; Zhao, M.; New, L.; Gu, J.; Feng, L.; Di Padova, F.; Ulevitch, R.J.; Han, J. Characterization of the structure and function of the fourth member of p38 group mitogen-activated protein kinases, p38delta. J. Biol. Chem. 1997, 272, 30122–30128. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Jiang, Y.; Ulevitch, R.J.; Han, J. The primary structure of p38 gamma: A new member of p38 group of MAP kinases. Biochem. Biophys. Res. Commun. 1996, 228, 334–340. [Google Scholar] [CrossRef]
- O’Callaghan, C.; Fanning, L.J.; Barry, O.P. p38δ MAPK: Emerging Roles of a Neglected Isoform. Int. J. Cell. Biol. 2014, 2014, 272689. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.H.; Porras, A.; Alonso, G.; Jones, M.; Vintersten, K.; Panelli, S.; Valladares, A.; Perez, L.; Klein, R.; Nebreda, A.R. Essential role of p38alpha MAP kinase in placental but not embryonic cardiovascular development. Mol. Cell 2000, 6, 109–116. [Google Scholar] [CrossRef]
- Mudgett, J.S.; Ding, J.; Guh-Siesel, L.; Chartrain, N.A.; Yang, L.; Gopal, S.; Shen, M.M. Essential role for p38alpha mitogen-activated protein kinase in placental angiogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 10454–10459. [Google Scholar] [CrossRef] [Green Version]
- Beardmore, V.A.; Hinton, H.J.; Eftychi, C.; Apostolaki, M.; Armaka, M.; Darragh, J.; McIlrath, J.; Carr, J.M.; Armit, L.J.; Clacher, C.; et al. Generation and characterization of p38beta (MAPK11) gene-targeted mice. Mol. Cell. Biol. 2005, 25, 10454–10464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenblatt, M.B.; Shim, J.H.; Zou, W.; Sitara, D.; Schweitzer, M.; Hu, D.; Lotinun, S.; Sano, Y.; Baron, R.; Park, J.M.; et al. The p38 MAPK pathway is essential for skeletogenesis and bone homeostasis in mice. J. Clin. Investig. 2010, 120, 2457–2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Johnson, G.L. PB1 domains of MEKK2 and MEKK3 interact with the MEK5 PB1 domain for activation of the ERK5 pathway. J. Biol. Chem. 2003, 278, 36989–36992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meister, M.; Tomasovic, A.; Banning, A.; Tikkanen, R. Mitogen-Activated Protein (MAP) Kinase Scaffolding Proteins: A Recount. Int. J. Mol. Sci. 2013, 14, 4854–4884. [Google Scholar] [CrossRef] [Green Version]
- Grimsey, N.J.; Aguilar, B.; Smith, T.H.; Le, P.; Soohoo, A.L.; Puthenveedu, M.A.; Nizet, V.; Trejo, J. Ubiquitin plays an atypical role in GPCR-induced p38 MAP kinase activation on endosomes. J. Cell Biol. 2015, 210, 1117–1131. [Google Scholar] [CrossRef]
- Grimsey, N.J.; Narala, R.; Rada, C.C.; Mehta, S.; Stephens, B.S.; Kufareva, I.; Lapek, J.; Gonzalez, D.J.; Handel, T.M.; Zhang, J.; et al. A Tyrosine Switch on NEDD4-2 E3 Ligase Transmits GPCR Inflammatory Signaling. Cell Rep. 2018, 24, 3312–3323.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlik, M.T.; Abell, A.N.; Johnson, N.L.; Sun, W.; Cuevas, B.D.; Lobel-Rice, K.E.; Horne, E.A.; Dell’Acqua, M.L.; Johnson, G.L. Rac-MEKK3-MKK3 scaffolding for p38 MAPK activation during hyperosmotic shock. Nat. Cell Biol. 2003, 5, 1104–1110. [Google Scholar] [CrossRef]
- Galperin, E.; Sorkin, A. Endosomal targeting of MEK2 requires RAF, MEK kinase activity and clathrin-dependent endocytosis. Traffic 2008, 9, 1776–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canovas, B.; Nebreda, A.R. Diversity and versatility of p38 kinase signalling in health and disease. Nat. Rev. Mol. Cell Biol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wu, J.; Silke, J. An overview of mammalian p38 mitogen-activated protein kinases, central regulators of cell stress and receptor signaling. F1000Research 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Cuenda, A.; Sanz-Ezquerro, J.J. p38γ and p38δ: From Spectators to Key Physiological Players. Trends Biochem. Sci. 2017, 42, 431–442. [Google Scholar] [CrossRef]
- Zu, Y.L.; Wu, F.; Gilchrist, A.; Ai, Y.; Labadia, M.E.; Huang, C.K. The primary structure of a human MAP kinase activated protein kinase 2. Biochem. Biophys. Res. Commun. 1994, 200, 1118–1124. [Google Scholar] [CrossRef]
- Soni, S.; Anand, P.; Padwad, Y.S. MAPKAPK2: The master regulator of RNA-binding proteins modulates transcript stability and tumor progression. J. Exp. Clin. Cancer Res. 2019, 38, 121. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Rouse, J.; Zhang, A.; Cariati, S.; Cohen, P.; Comb, M.J. FGF and stress regulate CREB and ATF-1 via a pathway involving p38 MAP kinase and MAPKAP kinase-2. EMBO J. 1996, 15, 4629–4642. [Google Scholar] [CrossRef]
- Guay, J.; Lambert, H.; Gingras-Breton, G.; Lavoie, J.N.; Huot, J.; Landry, J. Regulation of actin filament dynamics by p38 map kinase-mediated phosphorylation of heat shock protein 27. J. Cell Sci. 1997, 110, 357–368. [Google Scholar]
- Soni, S.; Saroch, M.K.; Chander, B.; Tirpude, N.V.; Padwad, Y.S. MAPKAPK2 plays a crucial role in the progression of head and neck squamous cell carcinoma by regulating transcript stability. J. Exp. Clin. Cancer Res. 2019, 38, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyskens, K.M.S.E.; Arthur, J.S.C. Emerging Roles of the Mitogen and Stress Activated Kinases MSK1 and MSK2. Front. Cell Dev. Biol. 2016, 4, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trempolec, N.; Dave-Coll, N.; Nebreda, A.R. SnapShot: p38 MAPK substrates. Cell 2013, 152, 924–924.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes-Xavier, C.; Romá-Mateo, C.; Ríos, P.; Tárrega, C.; Cejudo-Marín, R.; Tabernero, L.; Pulido, R. Dual-specificity MAP kinase phosphatases as targets of cancer treatment. Anticancer Agents Med. Chem. 2011, 11, 109–132. [Google Scholar] [CrossRef]
- Sun, H.; Charles, C.H.; Lau, L.F.; Tonks, N.K. MKP-1 (3CH134), an immediate early gene product, is a dual specificity phosphatase that dephosphorylates MAP kinase in vivo. Cell 1993, 75, 487–493. [Google Scholar] [CrossRef]
- Chen, H.F.; Chuang, H.C.; Tan, T.H. Regulation of Dual-Specificity Phosphatase (DUSP) Ubiquitination and Protein Stability. Int. J. Mol. Sci. 2019, 20, 2668. [Google Scholar] [CrossRef] [Green Version]
- Tomida, T.; Takekawa, M.; Saito, H. Oscillation of p38 activity controls efficient pro-inflammatory gene expression. Nat. Commun. 2015, 6, 8350. [Google Scholar] [CrossRef] [Green Version]
- Takekawa, M.; Maeda, T.; Saito, H. Protein phosphatase 2Calpha inhibits the human stress-responsive p38 and JNK MAPK pathways. EMBO J. 1998, 17, 4744–4752. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Hu, X.; Sun, B.; Yang, T.; Shi, J.; Zhang, L.; Zhao, Y. Phosphatase Wip1 negatively regulates neutrophil development through p38 MAPK-STAT1. Blood 2013, 121, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Topolska-Woś, A.M.; Rosińska, S.; Filipek, A. MAP kinase p38 is a novel target of CacyBP/SIP phosphatase. Amino Acids 2017, 49, 1069–1076. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Boehm, J.; Lee, J.C. p38 MAP kinases: Key signalling molecules as therapeutic targets for inflammatory diseases. Nat. Rev. Drug Discov. 2003, 2, 717–726. [Google Scholar] [CrossRef]
- Adams, J.L.; Badger, A.M.; Kumar, S.; Lee, J.C. 1 p38 MAP Kinase: Molecular Target for the Inhibition of Pro-inflammatory Cytokines. In Progress in Medicinal Chemistry; King, F.D., Oxford, A.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; Volume 38, pp. 1–60. [Google Scholar]
- Xing, L.; Shieh, H.S.; Selness, S.R.; Devraj, R.V.; Walker, J.K.; Devadas, B.; Hope, H.R.; Compton, R.P.; Schindler, J.F.; Hirsch, J.L.; et al. Structural bioinformatics-based prediction of exceptional selectivity of p38 MAP kinase inhibitor PH-797804. Biochemistry 2009, 48, 6402–6411. [Google Scholar] [CrossRef]
- Wrobleski, S.T.; Lin, S.; Dhar, T.G.; Dyckman, A.J.; Li, T.; Pitt, S.; Zhang, R.; Fan, Y.; Doweyko, A.M.; Tokarski, J.S.; et al. The identification of novel p38α isoform selective kinase inhibitors having an unprecedented p38α binding mode. Bioorg. Med. Chem. Lett. 2013, 23, 4120–4126. [Google Scholar] [CrossRef]
- Das, J.; Moquin, R.V.; Pitt, S.; Zhang, R.; Shen, D.R.; McIntyre, K.W.; Gillooly, K.; Doweyko, A.M.; Sack, J.S.; Zhang, H.; et al. Pyrazolo-pyrimidines: A novel heterocyclic scaffold for potent and selective p38 alpha inhibitors. Bioorg. Med. Chem. Lett. 2008, 18, 2652–2657. [Google Scholar] [CrossRef]
- Xing, L. Clinical candidates of small molecule p38 MAPK inhibitors for inflammatory diseases. MAP Kinase 2015, 4, 7. [Google Scholar] [CrossRef]
- Devadas, B.; Selness, S.R.; Xing, L.; Madsen, H.M.; Marrufo, L.D.; Shieh, H.; Messing, D.M.; Yang, J.Z.; Morgan, H.M.; Anderson, G.D.; et al. Substituted N-aryl-6-pyrimidinones: A new class of potent, selective, and orally active p38 MAP kinase inhibitors. Bioorg. Med. Chem. Lett. 2011, 21, 3856–3860. [Google Scholar] [CrossRef]
- Selness, S.R.; Devraj, R.V.; Devadas, B.; Walker, J.K.; Boehm, T.L.; Durley, R.C.; Shieh, H.; Xing, L.; Rucker, P.V.; Jerome, K.D.; et al. Discovery of PH-797804, a highly selective and potent inhibitor of p38 MAP kinase. Bioorg. Med. Chem. Lett. 2011, 21, 4066–4071. [Google Scholar] [CrossRef] [PubMed]
- Grimes, J.M.; Grimes, K.V. p38 MAPK inhibition: A promising therapeutic approach for COVID-19. J. Mol. Cell. Cardiol. 2020, 144, 63–65. [Google Scholar] [CrossRef]
- Cheriyan, J.; Webb, A.J.; Sarov-Blat, L.; Elkhawad, M.; Wallace, S.M.; Mäki-Petäjä, K.M.; Collier, D.J.; Morgan, J.; Fang, Z.; Willette, R.N.; et al. Inhibition of p38 mitogen-activated protein kinase improves nitric oxide-mediated vasodilatation and reduces inflammation in hypercholesterolemia. Circulation 2011, 123, 515–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbour, A.M.; Sarov-Blat, L.; Cai, G.; Fossler, M.J.; Sprecher, D.L.; Graggaber, J.; McGeoch, A.T.; Maison, J.; Cheriyan, J. Safety, tolerability, pharmacokinetics and pharmacodynamics of losmapimod following a single intravenous or oral dose in healthy volunteers. Br. J. Clin. Pharmacol. 2013, 76, 99–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie, J.D.; Vaslef, S.; Chang, P.K.; May, A.K.; Gunn, S.R.; Yang, S.; Hardes, K.; Kahl, L.; Powley, W.M.; Lipson, D.A.; et al. A Randomized Dose-Escalation Study of the Safety and Anti-Inflammatory Activity of the p38 Mitogen-Activated Protein Kinase Inhibitor Dilmapimod in Severe Trauma Subjects at Risk for Acute Respiratory Distress Syndrome. Crit. Care Med. 2015, 43, 1859–1869. [Google Scholar] [CrossRef] [PubMed]
- Strâmbu, I.R.; Kobalava, Z.D.; Magnusson, B.P.; MacKinnon, A.; Parkin, J.M. Phase II Study of Single/Repeated Doses of Acumapimod (BCT197) to Treat Acute Exacerbations of COPD. J. Chronic Obstr. Pulm. Dis. 2019, 16, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Hölscher, C.; Gräb, J.; Hölscher, A.; Müller, A.L.; Schäfer, S.C.; Rybniker, J. Chemical p38 MAP kinase inhibition constrains tissue inflammation and improves antibiotic activity in Mycobacterium tuberculosis-infected mice. Sci. Rep. 2020, 10, 13629. [Google Scholar] [CrossRef] [PubMed]
- Nichols, C.; Ng, J.; Keshu, A.; Kelly, G.; Conte, M.R.; Marber, M.S.; Fraternali, F.; De Nicola, G.F. Mining the PDB for Tractable Cases Where X-ray Crystallography Combined with Fragment Screens Can Be Used to Systematically Design Protein-Protein Inhibitors: Two Test Cases Illustrated by IL1β-IL1R and p38α-TAB1 Complexes. J. Med. Chem. 2020, 63, 7559–7568. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.G.; Tulapurkar, M.E.; Ramarathnam, A.; Brophy, A.; Martinez, R., 3rd; Hom, K.; Hodges, T.; Samadani, R.; Singh, I.S.; MacKerell, A.D., Jr.; et al. Novel Noncatalytic Substrate-Selective p38α-Specific MAPK Inhibitors with Endothelial-Stabilizing and Anti-Inflammatory Activity. J. Immunol. 2017, 198, 3296–3306. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Sun, X.; Ye, Y.; Lu, Y.; Zuo, J.; Liu, W.; Elcock, A.; Zhu, S. p38α Mitogen-Activated Protein Kinase Is a Druggable Target in Pancreatic Adenocarcinoma. Front. Oncol. 2019, 9, 1294. [Google Scholar] [CrossRef]
- Wang, C.; Hockerman, S.; Jacobsen, E.J.; Alippe, Y.; Selness, S.R.; Hope, H.R.; Hirsch, J.L.; Mnich, S.J.; Saabye, M.J.; Hood, W.F.; et al. Selective inhibition of the p38α MAPK-MK2 axis inhibits inflammatory cues including inflammasome priming signals. J. Exp. Med. 2018, 215, 1315–1325. [Google Scholar] [CrossRef]
- De Nicola, G.F.; Bassi, R.; Nichols, C.; Fernandez-Caggiano, M.; Golforoush, P.A.; Thapa, D.; Anderson, R.; Martin, E.D.; Verma, S.; Kleinjung, J.; et al. The TAB1-p38alpha complex aggravates myocardial injury and can be targeted by small molecules. JCI Insight 2018, 3. [Google Scholar] [CrossRef]
- Astolfi, A.; Manfroni, G.; Cecchetti, V.; Barreca, M.L. A Comprehensive Structural Overview of p38α Mitogen-Activated Protein Kinase in Complex with ATP-Site and Non-ATP-Site Binders. ChemMedChem 2018, 13, 7–14. [Google Scholar] [CrossRef]
- De Nicola, G.F.; Martin, E.D.; Chaikuad, A.; Bassi, R.; Clark, J.; Martino, L.; Verma, S.; Sicard, P.; Tata, R.; Atkinson, R.A.; et al. Mechanism and consequence of the autoactivation of p38α mitogen-activated protein kinase promoted by TAB1. Nat. Struct. Mol. Biol. 2013, 20, 1182–1190. [Google Scholar] [CrossRef] [Green Version]
- Ge, B.; Gram, H.; Di Padova, F.; Huang, B.; New, L.; Ulevitch, R.J.; Luo, Y.; Han, J. MAPKK-independent activation of p38alpha mediated by TAB1-dependent autophosphorylation of p38alpha. Science 2002, 295, 1291–1294. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, M.; Omori, E.; Kim, J.Y.; Komatsu, Y.; Scott, G.; Ray, M.K.; Yamada, G.; Matsumoto, K.; Mishina, Y.; Ninomiya-Tsuji, J. TAK1-binding protein 1, TAB1, mediates osmotic stress-induced TAK1 activation but is dispensable for TAK1-mediated cytokine signaling. J. Biol. Chem. 2008, 283, 33080–33086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, R.; Sidler, C.L.; Thali, R.F.; Winssinger, N.; Cheung, P.C.; Neumann, D. Autoactivation of transforming growth factor beta-activated kinase 1 is a sequential bimolecular process. J. Biol. Chem. 2010, 285, 25753–25766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, K.; Matsumoto, K.; Ninomiya-Tsuji, J. TAK1 mitogen-activated protein kinase kinase kinase is activated by autophosphorylation within its activation loop. J. Biol. Chem. 2000, 275, 7359–7364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, P.C.; Campbell, D.G.; Nebreda, A.R.; Cohen, P. Feedback control of the protein kinase TAK1 by SAPK2a/p38alpha. EMBO J. 2003, 22, 5793–5805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, A.; Beuerlein, K.; Eckart, C.; Weiser, H.; Dickkopf, B.; Muller, H.; Sakurai, H.; Kracht, M. Identification and functional characterization of novel phosphorylation sites in TAK1-binding protein (TAB) 1. PLoS ONE 2011, 6, e29256. [Google Scholar] [CrossRef]
- Richardson, L.; Dixon, C.L.; Aguilera-Aguirre, L.; Menon, R. Oxidative stress-induced TGF-beta/TAB1-mediated p38MAPK activation in human amnion epithelial cells. Biol. Reprod. 2018, 99, 1100–1112. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Miller, E.J.; Ninomiya-Tsuji, J.; Russell, R.R., 3rd; Young, L.H. AMP-activated protein kinase activates p38 mitogen-activated protein kinase by increasing recruitment of p38 MAPK to TAB1 in the ischemic heart. Circ. Res. 2005, 97, 872–879. [Google Scholar] [CrossRef] [Green Version]
- Ota, A.; Zhang, J.; Ping, P.; Han, J.; Wang, Y. Specific regulation of noncanonical p38alpha activation by Hsp90-Cdc37 chaperone complex in cardiomyocyte. Circ. Res. 2010, 106, 1404–1412. [Google Scholar] [CrossRef] [Green Version]
- Theivanthiran, B.; Kathania, M.; Zeng, M.; Anguiano, E.; Basrur, V.; Vandergriff, T.; Pascual, V.; Wei, W.Z.; Massoumi, R.; Venuprasad, K. The E3 ubiquitin ligase Itch inhibits p38alpha signaling and skin inflammation through the ubiquitylation of Tab1. Sci. Signal. 2015, 8, ra22. [Google Scholar] [CrossRef]
- Wang, Q.; Feng, J.; Wang, J.; Zhang, X.; Zhang, D.; Zhu, T.; Wang, W.; Wang, X.; Jin, J.; Cao, J.; et al. Disruption of TAB1/p38alpha interaction using a cell-permeable peptide limits myocardial ischemia/reperfusion injury. Mol. Ther. 2013, 21, 1668–1677. [Google Scholar] [CrossRef] [Green Version]
- Pei, Y.J.; Wang, Q.Y.; Zhang, J.Y.; Guo, Y.H.; Feng, J.N. Characterization and Evaluation of Key Sites in the Peptide Inhibitor of TAB1/p38 alpha Interaction. Int. J. Pept. Res. Ther. 2018, 24, 225–233. [Google Scholar] [CrossRef]
- Zhou, H.; Zheng, M.; Chen, J.; Xie, C.; Kolatkar, A.R.; Zarubin, T.; Ye, Z.; Akella, R.; Lin, S.; Goldsmith, E.J.; et al. Determinants that control the specific interactions between TAB1 and p38alpha. Mol. Cell. Biol. 2006, 26, 3824–3834. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, Y.; Shibuya, H.; Takeda, N.; Ninomiya-Tsuji, J.; Yasui, T.; Miyado, K.; Sekimoto, T.; Ueno, N.; Matsumoto, K.; Yamada, G. Targeted disruption of the Tab1 gene causes embryonic lethality and defects in cardiovascular and lung morphogenesis. Mech. Dev. 2002, 119, 239–249. [Google Scholar] [CrossRef]
- Grimsey, N.J.; Lin, Y.; Narala, R.; Rada, C.C.; Mejia-Pena, H.; Trejo, J. G protein-coupled receptors activate p38 MAPK via a non-canonical TAB1-TAB2 and TAB1-TAB3 dependent pathway in endothelial cells. J. Biol. Chem. 2019, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvador, J.M.; Mittelstadt, P.R.; Guszczynski, T.; Copeland, T.D.; Yamaguchi, H.; Appella, E.; Fornace, A.J., Jr.; Ashwell, J.D. Alternative p38 activation pathway mediated by T cell receptor-proximal tyrosine kinases. Nat. Immunol. 2005, 6, 390–395. [Google Scholar] [CrossRef]
- Diskin, R.; Lebendiker, M.; Engelberg, D.; Livnah, O. Structures of p38alpha active mutants reveal conformational changes in L16 loop that induce autophosphorylation and activation. J. Mol. Biol. 2007, 365, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Flower, D.R. Modelling G-protein-coupled receptors for drug design. Biochim. Biophys. Acta 1999, 1422, 207–234. [Google Scholar] [CrossRef]
- Azzi, M.; Charest, P.G.; Angers, S.; Rousseau, G.; Kohout, T.; Bouvier, M.; Piñeyro, G. β-Arrestin-mediated activation of MAPK by inverse agonists reveals distinct active conformations for G protein-coupled receptors. Proc. Natl. Acad. Sci. USA 2003, 100, 11406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutkind, J.S. Regulation of Mitogen-Activated Protein Kinase Signaling Networks by G Protein-Coupled Receptors. Sci. STKE 2000, 2000, re1. [Google Scholar] [CrossRef]
- McDonald, P.H.; Chow, C.-W.; Miller, W.E.; Laporte, S.A.; Field, M.E.; Lin, F.-T.; Davis, R.J.; Lefkowitz, R.J. β-Arrestin 2: A Receptor-Regulated MAPK Scaffold for the Activation of JNK3. Science 2000, 290, 1574. [Google Scholar] [CrossRef]
- Shenoy, S.K.; Lefkowitz, R.J. Seven-transmembrane receptor signaling through beta-arrestin. Sci. STKE 2005, 2005, cm10. [Google Scholar] [CrossRef]
- Burton, J.C.; Grimsey, N.J. Ubiquitination as a Key Regulator of Endosomal Signaling by GPCRs. Front. Cell Dev. Biol. 2019, 7, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimsey, N.J.; Trejo, J. Integration of endothelial protease-activated receptor-1 inflammatory signaling by ubiquitin. Curr. Opin. Hematol. 2016, 23, 274–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dores, M.R.; Chen, B.; Lin, H.; Soh, U.J.; Paing, M.M.; Montagne, W.A.; Meerloo, T.; Trejo, J. ALIX binds a YPX(3)L motif of the GPCR PAR1 and mediates ubiquitin-independent ESCRT-III/MVB sorting. J. Cell. Biol. 2012, 197, 407–419. [Google Scholar] [CrossRef]
- Dores, M.R.; Grimsey, N.J.; Mendez, F.; Trejo, J. ALIX Regulates the Ubiquitin-Independent Lysosomal Sorting of the P2Y1 Purinergic Receptor via a YPX3L Motif. PLoS ONE 2016, 11, e0157587. [Google Scholar] [CrossRef]
- Dores, M.R.; Lin, H.; Grimsey, N.J.; Mendez, F.; Trejo, J. The alpha-arrestin ARRDC3 mediates ALIX ubiquitination and G protein-coupled receptor lysosomal sorting. Mol. Biol. Cell 2015, 26, 4660–4673. [Google Scholar] [CrossRef]
- Kulathu, Y.; Akutsu, M.; Bremm, A.; Hofmann, K.; Komander, D. Two-sided ubiquitin binding explains specificity of the TAB2 NZF domain. Nat. Struct. Mol. Biol. 2009, 16, 1328–1330. [Google Scholar] [CrossRef]
- Romero-Becerra, R.; Santamans, A.M.; Folgueira, C.; Sabio, G. p38 MAPK pathway in the heart: New insights in health and disease. Int. J. Mol. Sci. 2020, 21, 7412. [Google Scholar] [CrossRef]
- Arabacilar, P.; Marber, M. The case for inhibiting p38 mitogen-activated protein kinase in heart failure. Front. Pharmacol. 2015, 6, 102. [Google Scholar] [CrossRef] [Green Version]
- Song, N.; Ma, J.; Meng, X.W.; Liu, H.; Wang, H.; Song, S.Y.; Chen, Q.C.; Liu, H.Y.; Zhang, J.; Peng, K.; et al. Heat Shock Protein 70 Protects the Heart from Ischemia/Reperfusion Injury through Inhibition of p38 MAPK Signaling. Oxid. Med. Cell. Longev. 2020, 2020, 3908641. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, B.; Feil, R.; Hofmann, F.; Willenbockel, C.; Drexler, H.; Smolenski, A.; Lohmann, S.M.; Wollert, K.C. cGMP-dependent protein kinase type I inhibits TAB1-p38 mitogen-activated protein kinase apoptosis signaling in cardiac myocytes. J. Biol. Chem. 2006, 281, 32831–32840. [Google Scholar] [CrossRef] [Green Version]
- Tanno, M.; Bassi, R.; Gorog, D.A.; Saurin, A.T.; Jiang, J.; Heads, R.J.; Martin, J.L.; Davis, R.J.; Flavell, R.A.; Marber, M.S. Diverse mechanisms of myocardial p38 mitogen-activated protein kinase activation: Evidence for MKK-independent activation by a TAB1-associated mechanism contributing to injury during myocardial ischemia. Circ. Res. 2003, 93, 254–261. [Google Scholar] [CrossRef] [Green Version]
- Zheng, D.Y.; Zhou, M.; Jin, J.; He, M.; Wang, Y.; Du, J.; Xiao, X.Y.; Li, P.Y.; Ye, A.Z.; Liu, J.; et al. Inhibition of P38 MAPK Downregulates the Expression of IL-1beta to Protect Lung from Acute Injury in Intestinal Ischemia Reperfusion Rats. Mediators Inflamm. 2016, 2016, 9348037. [Google Scholar] [CrossRef] [Green Version]
- Du, C.S.; Yang, R.F.; Song, S.W.; Wang, Y.P.; Kang, J.H.; Zhang, R.; Su, D.F.; Xie, X. Magnesium Lithospermate B Protects Cardiomyocytes from Ischemic Injury Via Inhibition of TAB1-p38 Apoptosis Signaling. Front. Pharmacol. 2010, 1, 111. [Google Scholar] [CrossRef] [Green Version]
- Thapa, D.; Nichols, C.; Bassi, R.; Martin, E.D.; Verma, S.; Conte, M.R.; De Santis, V.; De Nicola, G.F.; Marber, M.S. TAB1-Induced Autoactivation of p38α Mitogen-Activated Protein Kinase Is Crucially Dependent on Threonine 185. Mol. Cell. Biol. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Ding, L.; Ji, H.; Xu, Z.; Liu, Q.; Zheng, Y. The Role of p38 MAPK in the Development of Diabetic Cardiomyopathy. Int. J. Mol. Sci. 2016, 17, 1037. [Google Scholar] [CrossRef] [Green Version]
- Al-Shabrawey, M.; Hussein, K.; Wang, F.; Wan, M.; Elmasry, K.; Elsherbiny, N.; Saleh, H.; Yu, P.B.; Tawfik, A.; Ibrahim, A.S. Bone Morphogenetic Protein-2 Induces Non-Canonical Inflammatory and Oxidative Pathways in Human Retinal Endothelial Cells. Front. Immunol. 2020, 11, 568795. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Zhou, D.; Zhang, L.S.; Deng, F.X.; Shu, S.; Wang, L.J.; Wu, Y.; Guo, N.; Zhou, J.; et al. Angiotensin II deteriorates advanced atherosclerosis by promoting MerTK cleavage and impairing efferocytosis through the AT(1)R/ROS/p38 MAPK/ADAM17 pathway. Am. J. Physiol. Cell Physiol. 2019, 317, C776–C787. [Google Scholar] [CrossRef]
- Corre, I.; Paris, F.; Huot, J. The p38 pathway, a major pleiotropic cascade that transduces stress and metastatic signals in endothelial cells. Oncotarget 2017, 8, 55684–55714. [Google Scholar] [CrossRef]
- Fisk, M.; Gajendragadkar, P.R.; Maki-Petaja, K.M.; Wilkinson, I.B.; Cheriyan, J. Therapeutic potential of p38 MAP kinase inhibition in the management of cardiovascular disease. Am. J. Cardiovasc. Drugs 2014, 14, 155–165. [Google Scholar] [CrossRef]
- Reustle, A.; Torzewski, M. Role of p38 MAPK in Atherosclerosis and Aortic Valve Sclerosis. Int. J. Mol. Sci. 2018, 19, 3761. [Google Scholar] [CrossRef] [Green Version]
- Seitz, I.; Hess, S.; Schulz, H.; Eckl, R.; Busch, G.; Montens, H.P.; Brandl, R.; Seidl, S.; Schomig, A.; Ott, I. Membrane-type serine protease-1/matriptase induces interleukin-6 and -8 in endothelial cells by activation of protease-activated receptor-2: Potential implications in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 769–775. [Google Scholar] [CrossRef]
- Renda, T.; Baraldo, S.; Pelaia, G.; Bazzan, E.; Turato, G.; Papi, A.; Maestrelli, P.; Maselli, R.; Vatrella, A.; Fabbri, L.M.; et al. Increased activation of p38 MAPK in COPD. Eur. Respir. J. 2008, 31, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Pelaia, C.; Vatrella, A.; Sciacqua, A.; Terracciano, R.; Pelaia, G. Role of p38-mitogen-activated protein kinase in COPD: Pathobiological implications and therapeutic perspectives. Expert Rev. Respir. Med. 2020, 14, 485–491. [Google Scholar] [CrossRef]
- Gaffey, K.; Reynolds, S.; Plumb, J.; Kaur, M.; Singh, D. Increased phosphorylated p38 mitogen-activated protein kinase in COPD lungs. Eur. Respir. J. 2013, 42, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, J.; Harbron, C.; Lea, S.; Booth, G.; Cadden, P.; Wreggett, K.A.; Singh, D. Synergistic effects of p38 mitogen-activated protein kinase inhibition with a corticosteroid in alveolar macrophages from patients with chronic obstructive pulmonary disease. J. Pharmacol. Exp. Ther. 2011, 338, 732–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Xie, M.; He, X.; Gao, H. Activity of sputum p38 MAPK is correlated with airway inflammation and reduced FEV1 in COPD patients. Med. Sci. Monit. 2013, 19, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Amano, H.; Murata, K.; Matsunaga, H.; Tanaka, K.; Yoshioka, K.; Kobayashi, T.; Ishida, J.; Fukamizu, A.; Sugiyama, F.; Sudo, T.; et al. p38 Mitogen-activated protein kinase accelerates emphysema in mouse model of chronic obstructive pulmonary disease. J. Recept. Signal. Transduct. 2014, 34, 299–306. [Google Scholar] [CrossRef]
- Feng, Y.; Fang, Z.; Liu, B.; Zheng, X. p38MAPK plays a pivotal role in the development of acute respiratory distress syndrome. Clinics 2019, 74, e509. [Google Scholar] [CrossRef]
- Fang, W.; Cai, S.X.; Wang, C.L.; Sun, X.X.; Li, K.; Yan, X.W.; Sun, Y.B.; Sun, X.Z.; Gu, C.K.; Dai, M.Y.; et al. Modulation of mitogenactivated protein kinase attenuates sepsisinduced acute lung injury in acute respiratory distress syndrome rats. Mol. Med. Rep. 2017, 16, 9652–9658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, X.; Fan, L.; He, T.; Jia, W.; Yang, L.; Zhang, J.; Liu, Y.; Shi, J.; Su, L.; Hu, D. SIRT1 protects rat lung tissue against severe burn-induced remote ALI by attenuating the apoptosis of PMVECs via p38 MAPK signaling. Sci. Rep. 2015, 5, 10277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, L.L.; Tan, Y.; Ma, H.Y.; Dai, P.; Qin, Y.X.; Yang, R.A.; Xu, Y.Y.; Deng, Z.; Zhao, W.; Xia, Q.J.; et al. Administration of SB239063, a potent p38 MAPK inhibitor, alleviates acute lung injury induced by intestinal ischemia reperfusion in rats associated with AQP4 downregulation. Int. Immunopharmacol. 2016, 38, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Ren, W.; Jiang, Z.; Zhu, L. Regulation of the NLRP3 inflammasome and macrophage pyroptosis by the p38 MAPK signaling pathway in a mouse model of acute lung injury. Mol. Med. Rep. 2018, 18, 4399–4409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Sun, F.; Wang, L.; Gao, M.; Xie, Y.; Sun, Y.; Liu, H.; Yuan, Y.; Yi, W.; Huang, Z.; et al. Virus-induced p38 MAPK activation facilitates viral infection. Theranostics 2020, 10, 12223–12240. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, S.S.; Jensen, S.B.; Chiliveru, S.; Melchjorsen, J.; Julkunen, I.; Gaestel, M.; Arthur, J.S.; Flavell, R.A.; Ghosh, S.; Paludan, S.R. RIG-I-mediated activation of p38 MAPK is essential for viral induction of interferon and activation of dendritic cells: Dependence on TRAF2 and TAK1. J. Biol. Chem. 2009, 284, 10774–10782. [Google Scholar] [CrossRef] [Green Version]
- Börgeling, Y.; Schmolke, M.; Viemann, D.; Nordhoff, C.; Roth, J.; Ludwig, S. Inhibition of p38 mitogen-activated protein kinase impairs influenza virus-induced primary and secondary host gene responses and protects mice from lethal H5N1 infection. J. Biol. Chem. 2014, 289, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Yang, Q.; Shao, J.; Zhang, L.; Ma, J.; Wang, Y.; Jiang, B.H.; Leng, J.; Bai, X. Cyclooxygenase-2 induced β1-integrin expression in NSCLC and promoted cell invasion via the EP1/MAPK/E2F-1/FoxC2 signal pathway. Sci. Rep. 2016, 6, 33823. [Google Scholar] [CrossRef]
- Patel, S.; Vetale, S.; Teli, P.; Mistry, R.; Chiplunkar, S. IL-10 production in non-small cell lung carcinoma patients is regulated by ERK, P38 and COX-2. J. Cell. Mol. Med. 2012, 16, 531–544. [Google Scholar] [CrossRef]
- Singh, R.K.; Najmi, A.K. Novel Therapeutic Potential of Mitogen-Activated Protein Kinase Activated Protein Kinase 2 (MK2) in Chronic Airway Inflammatory Disorders. Curr. Drug Targets 2019, 20, 367–379. [Google Scholar] [CrossRef]
- Wada, M.; Canals, D.; Adada, M.; Coant, N.; Salama, M.F.; Helke, K.L.; Arthur, J.S.; Shroyer, K.R.; Kitatani, K.; Obeid, L.M.; et al. P38 delta MAPK promotes breast cancer progression and lung metastasis by enhancing cell proliferation and cell detachment. Oncogene 2017, 36, 6649–6657. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Zhang, X.J.; Zou, H.; Zhang, Y.Y.; Xia, J.W.; Zhang, P.; Zhang, Y.Z.; Li, J.; Dong, L.; Wumaier, G.; et al. PTPL1 suppresses lung cancer cell migration via inhibiting TGF-β1-induced activation of p38 MAPK and Smad 2/3 pathways and EMT. Acta Pharmacol. Sin. 2021. [Google Scholar] [CrossRef]
- Koul, H.K.; Pal, M.; Koul, S. Role of p38 MAP Kinase Signal Transduction in Solid Tumors. Genes Cancer 2013, 4, 342–359. [Google Scholar] [CrossRef]
- Igea, A.; Nebreda, A.R. The Stress Kinase p38alpha as a Target for Cancer Therapy. Cancer Res. 2015, 75, 3997–4002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leelahavanichkul, K.; Amornphimoltham, P.; Molinolo, A.A.; Basile, J.R.; Koontongkaew, S.; Gutkind, J.S. A role for p38 MAPK in head and neck cancer cell growth and tumor-induced angiogenesis and lymphangiogenesis. Mol. Oncol. 2014, 8, 105–118. [Google Scholar] [CrossRef]
- Roy, S.; Roy, S.; Anuja, K.; Thakur, S.; Akhter, Y.; Padhi, S.; Banerjee, B. p38 Mitogen-activated protein kinase modulates cisplatin resistance in Head and Neck Squamous Cell Carcinoma cells. Arch. Oral Biol. 2021, 122, 104981. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sadat, S.H.; Ebisumoto, K.; Sakai, A.; Panuganti, B.A.; Ren, S.; Goto, Y.; Haft, S.; Fukusumi, T.; Ando, M.; et al. Cannabinoids Promote Progression of HPV-Positive Head and Neck Squamous Cell Carcinoma via p38 MAPK Activation. Clin. Cancer Res. 2020, 26, 2693–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avendano, M.S.; Garcia-Redondo, A.B.; Zalba, G.; Gonzalez-Amor, M.; Aguado, A.; Martinez-Revelles, S.; Beltran, L.M.; Camacho, M.; Cachofeiro, V.; Alonso, M.J.; et al. mPGES-1 (Microsomal Prostaglandin E Synthase-1) Mediates Vascular Dysfunction in Hypertension Through Oxidative Stress. Hypertension 2018, 72, 492–502. [Google Scholar] [CrossRef]
- Cánovas, B.; Igea, A.; Sartori, A.A.; Gomis, R.R.; Paull, T.T.; Isoda, M.; Pérez-Montoyo, H.; Serra, V.; González-Suárez, E.; Stracker, T.H.; et al. Targeting p38α Increases DNA Damage, Chromosome Instability, and the Anti-tumoral Response to Taxanes in Breast Cancer Cells. Cancer Cell 2018, 33, 1094–1110.e8. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Limón, A.; Joaquin, M.; Caballero, M.; Posas, F.; de Nadal, E. The p38 Pathway: From Biology to Cancer Therapy. Int. J. Mol. Sci. 2020, 21, 1913. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.; Koul, S.; Petersen, J.; Khandrika, L.; Hwa, J.S.; Meacham, R.B.; Wilson, S.; Koul, H.K. p38 Mitogen-Activated Protein Kinase–Driven MAPKAPK2 Regulates Invasion of Bladder Cancer by Modulation of MMP-2 and MMP-9 Activity. Cancer Res. 2010, 70, 832. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.K.; Kim, N.J. Recent Advances in the Inhibition of p38 MAPK as a Potential Strategy for the Treatment of Alzheimer’s Disease. Molecules 2017, 22, 1287. [Google Scholar] [CrossRef] [Green Version]
- Corrêa, S.A.; Eales, K.L. The Role of p38 MAPK and Its Substrates in Neuronal Plasticity and Neurodegenerative Disease. J. Signal. Transduct. 2012, 2012, 649079. [Google Scholar] [CrossRef] [Green Version]
- Germann, U.A.; Alam, J.J. P38α MAPK Signaling-A Robust Therapeutic Target for Rab5-Mediated Neurodegenerative Disease. Int. J. Mol. Sci. 2020, 21, 5485. [Google Scholar] [CrossRef]
- Ittner, A.; Asih, P.R.; Tan, A.R.P.; Prikas, E.; Bertz, J.; Stefanoska, K.; Lin, Y.; Volkerling, A.M.; Ke, Y.D.; Delerue, F.; et al. Reduction of advanced tau-mediated memory deficits by the MAP kinase p38γ. Acta Neuropathol. 2020, 140, 279–294. [Google Scholar] [CrossRef]
- He, J.; Zhong, W.; Zhang, M.; Zhang, R.; Hu, W. P38 Mitogen-activated Protein Kinase and Parkinson’s Disease. Transl. Neurosci. 2018, 9, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, X.; Niu, M.; Zhang, X.; Wang, J.; Zhou, C.; Xie, A. RAGE Silencing Ameliorates Neuroinflammation by Inhibition of p38-NF-κB Signaling Pathway in Mouse Model of Parkinson’s Disease. Front. Neurosci. 2020, 14, 353. [Google Scholar] [CrossRef]
- Wang, Q.; Zheng, H.; Zhang, Z.F.; Zhang, Y.X. Ginsenoside Rg1 modulates COX-2 expression in the substantia nigra of mice with MPTP-induced Parkinson disease through the P38 signaling pathway. Nan Fang Yi Ke Da Xue Xue Bao 2008, 28, 1594–1598. [Google Scholar] [PubMed]
- De Vos, K.J.; Hafezparast, M. Neurobiology of axonal transport defects in motor neuron diseases: Opportunities for translational research? Neurobiol. Dis. 2017, 105, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, K.L.; Greensmith, L.; Schiavo, G. Regulation of Axonal Transport by Protein Kinases. Trends Biochem. Sci. 2015, 40, 597–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, K.L.; Kalmar, B.; Rhymes, E.R.; Fellows, A.D.; Ahmed, M.; Whiting, P.; Davies, C.H.; Greensmith, L.; Schiavo, G. Inhibiting p38 MAPK alpha rescues axonal retrograde transport defects in a mouse model of ALS. Cell Death Dis. 2018, 9, 596. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.M.; Van Alstyne, M.; Lotti, F.; Bianchetti, E.; Tisdale, S.; Watterson, D.M.; Mentis, G.Z.; Pellizzoni, L. Stasimon Contributes to the Loss of Sensory Synapses and Motor Neuron Death in a Mouse Model of Spinal Muscular Atrophy. Cell Rep. 2019, 29, 3885–3901.e5. [Google Scholar] [CrossRef] [Green Version]
- Kyosseva, S.V. Targeting MAPK Signaling in Age-Related Macular Degeneration. Ophthalmol Eye Dis. 2016, 8, 23–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pons, M.; Cousins, S.W.; Alcazar, O.; Striker, G.E.; Marin-Castaño, M.E. Angiotensin II–Induced MMP-2 Activity and MMP-14 and Basigin Protein Expression Are Mediated via the Angiotensin II Receptor Type 1–Mitogen-Activated Protein Kinase 1 Pathway in Retinal Pigment Epithelium: Implications for Age-Related Macular Degeneration. Am. J. Pathol. 2011, 178, 2665–2681. [Google Scholar] [CrossRef] [Green Version]
- Zou, W.; Luo, S.; Zhang, Z.; Cheng, L.; Huang, X.; Ding, N.; Pan, Y.; Wu, Z. ASK1/p38-mediated NLRP3 inflammasome signaling pathway contributes to aberrant retinal angiogenesis in diabetic retinopathy. Int. J. Mol. Med. 2021, 47, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Zhang, C.; Yang, Q.; Xie, H.; Liu, D.; Tian, H.; Lu, L.; Xu, J.Y.; Li, W.; Xu, G.; et al. Melatonin maintains inner blood-retinal barrier via inhibition of p38/TXNIP/NF-κB pathway in diabetic retinopathy. J. Cell. Physiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.J.; Byeon, H.E.; Cho, C.S.; Kim, Y.H.; Kim, J.H.; Che, J.H.; Seok, S.H.; Kwon, J.W.; Kim, J.H.; Lee, K. Histamine causes an imbalance between pro-angiogenic and anti-angiogenic factors in the retinal pigment epithelium of diabetic retina via H4 receptor/p38 MAPK axis. BMJ Open Diabetes Res. Care 2020, 8. [Google Scholar] [CrossRef]
- Zou, W.; Zhang, Z.; Luo, S.; Cheng, L.; Huang, X.; Ding, N.; Yu, J.; Pan, Y.; Wu, Z. p38 promoted retinal micro-angiogenesis through up-regulated RUNX1 expression in diabetic retinopathy. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Tang, J.; Li, G.; Berti-Mattera, L.; Lee, C.A.; Bartkowski, D.; Gale, D.; Monahan, J.; Niesman, M.R.; Alton, G.; et al. Effects of p38 MAPK inhibition on early stages of diabetic retinopathy and sensory nerve function. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2158–2164. [Google Scholar] [CrossRef] [Green Version]
- Dapper, J.D.; Crish, S.D.; Pang, I.H.; Calkins, D.J. Proximal inhibition of p38 MAPK stress signaling prevents distal axonopathy. Neurobiol. Dis. 2013, 59, 26–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, M.; Lipton, S.A. Targeting excitotoxic/free radical signaling pathways for therapeutic intervention in glaucoma. Prog. Brain Res. 2008, 173, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Cuenda, A.; Rousseau, S. p38 MAP-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schieven, G.L. The p38alpha kinase plays a central role in inflammation. Curr. Top. Med. Chem. 2009, 9, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Choy, E.H.; Panayi, G.S. Cytokine pathways and joint inflammation in rheumatoid arthritis. N. Engl. J. Med. 2001, 344, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Stokes, D.G.; Kremer, J.M. Potential of tumor necrosis factor neutralization strategies in rheumatologic disorders other than rheumatoid arthritis. Semin. Arth. Rheum. 2003, 33, 1–18. [Google Scholar] [CrossRef]
- Han, J.; Jiang, Y.; Li, Z.; Kravchenko, V.V.; Ulevitch, R.J. Activation of the transcription factor MEF2C by the MAP kinase p38 in inflammation. Nature 1997, 386, 296–299. [Google Scholar] [CrossRef]
- Guan, Z.; Buckman, S.Y.; Pentland, A.P.; Templeton, D.J.; Morrison, A.R. Induction of cyclooxygenase-2 by the activated MEKK1 --> SEK1/MKK4 --> p38 mitogen-activated protein kinase pathway. J. Biol. Chem. 1998, 273, 12901–12908. [Google Scholar] [CrossRef] [Green Version]
- Badger, A.M.; Roshak, A.K.; Cook, M.N.; Newman-Tarr, T.M.; Swift, B.A.; Carlson, K.; Connor, J.R.; Lee, J.C.; Gowen, M.; Lark, M.W.; et al. Differential effects of SB 242235, a selective p38 mitogen-activated protein kinase inhibitor, on IL-1 treated bovine and human cartilage/chondrocyte cultures. Osteoarthr. Cartil. 2000, 8, 434–443. [Google Scholar] [CrossRef] [Green Version]
- Wiehler, S.; Cuvelier, S.L.; Chakrabarti, S.; Patel, K.D. p38 MAP kinase regulates rapid matrix metalloproteinase-9 release from eosinophils. Biochem. Biophys. Res. Commun. 2004, 315, 463–470. [Google Scholar] [CrossRef]
- Da Silva, J.; Pierrat, B.; Mary, J.L.; Lesslauer, W. Blockade of p38 mitogen-activated protein kinase pathway inhibits inducible nitric-oxide synthase expression in mouse astrocytes. J. Biol. Chem. 1997, 272, 28373–28380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koprak, S.; Staruch, M.J.; Dumont, F.J. A specific inhibitor of the p38 mitogen activated protein kinase affects differentially the production of various cytokines by activated human T cells: Dependence on CD28 signaling and preferential inhibition of IL-10 production. Cell Immunol. 1999, 192, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Ananieva, O.; Darragh, J.; Johansen, C.; Carr, J.M.; McIlrath, J.; Park, J.M.; Wingate, A.; Monk, C.E.; Toth, R.; Santos, S.G.; et al. The kinases MSK1 and MSK2 act as negative regulators of Toll-like receptor signaling. Nat. Immunol. 2008, 9, 1028–1036. [Google Scholar] [CrossRef] [Green Version]
- Lang, R.; Raffi, F.A.M. Dual-Specificity Phosphatases in Immunity and Infection: An Update. Int. J. Mol. Sci. 2019, 20, 2710. [Google Scholar] [CrossRef] [Green Version]
- de la Cruz-Morcillo, M.A.; García-Cano, J.; Arias-González, L.; García-Gil, E.; Artacho-Cordón, F.; Ríos-Arrabal, S.; Valero, M.L.; Cimas, F.J.; Serrano-Oviedo, L.; Villas, M.V.; et al. Abrogation of the p38 MAPK α signaling pathway does not promote radioresistance but its activity is required for 5-Fluorouracil-associated radiosensitivity. Cancer Lett. 2013, 335, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Lepore Signorile, M.; Grossi, V.; Di Franco, S.; Forte, G.; Disciglio, V.; Fasano, C.; Sanese, P.; De Marco, K.; Susca, F.C.; Mangiapane, L.R.; et al. Pharmacological targeting of the novel β-catenin chromatin-associated kinase p38α in colorectal cancer stem cell tumorspheres and organoids. Cell Death Dis. 2021, 12, 316. [Google Scholar] [CrossRef]
- Pereira, L.; Igea, A.; Canovas, B.; Dolado, I.; Nebreda, A.R. Inhibition of p38 MAPK sensitizes tumour cells to cisplatin-induced apoptosis mediated by reactive oxygen species and JNK. EMBO Mol. Med. 2013, 5, 1759–1774. [Google Scholar] [CrossRef] [Green Version]
- Roche, O.; Fernández-Aroca, D.M.; Arconada-Luque, E.; García-Flores, N.; Mellor, L.F.; Ruiz-Hidalgo, M.J.; Sánchez-Prieto, R. p38β and Cancer: The Beginning of the Road. Int. J. Mol. Sci. 2020, 21, 7524. [Google Scholar] [CrossRef]
- Sahu, V.; Nigam, L.; Agnihotri, V.; Gupta, A.; Shekhar, S.; Subbarao, N.; Bhaskar, S.; Dey, S. Diagnostic Significance of p38 Isoforms (p38α, p38β, p38γ, p38δ) in Head and Neck Squamous Cell Carcinoma: Comparative Serum Level Evaluation and Design of Novel Peptide Inhibitor Targeting the Same. Cancer Res. Treat. 2019, 51, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Krementsov, D.N.; Thornton, T.M.; Teuscher, C.; Rincon, M. The emerging role of p38 mitogen-activated protein kinase in multiple sclerosis and its models. Mol. Cell. Biol. 2013, 33, 3728–3734. [Google Scholar] [CrossRef] [Green Version]
- Ruano, D.; Revilla, E.; Gavilán, M.P.; Vizuete, M.L.; Pintado, C.; Vitorica, J.; Castaño, A. Role of p38 and inducible nitric oxide synthase in the in vivo dopaminergic cells’ degeneration induced by inflammatory processes after lipopolysaccharide injection. Neuroscience 2006, 140, 1157–1168. [Google Scholar] [CrossRef]
- Lu, G.; Kang, Y.J.; Han, J.; Herschman, H.R.; Stefani, E.; Wang, Y. TAB-1 modulates intracellular localization of p38 MAP kinase and downstream signaling. J. Biol. Chem. 2006, 281, 6087–6095. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Guan, J.; Jiang, B.; Brenner, D.A.; Del Monte, F.; Ward, J.E.; Connors, L.H.; Sawyer, D.B.; Semigran, M.J.; Macgillivray, T.E.; et al. Amyloidogenic light chains induce cardiomyocyte contractile dysfunction and apoptosis via a non-canonical p38alpha MAPK pathway. Proc. Natl. Acad. Sci. USA 2010, 107, 4188–4193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Guan, J.; Plovie, E.; Seldin, D.C.; Connors, L.H.; Merlini, G.; Falk, R.H.; MacRae, C.A.; Liao, R. Human amyloidogenic light chain proteins result in cardiac dysfunction, cell death, and early mortality in zebrafish. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H95–H103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkusu-Tsukada, K.; Toda, M.; Udono, H.; Kawakami, Y.; Takahashi, K. Targeted inhibition of IL-10-secreting CD25- Treg via p38 MAPK suppression in cancer immunotherapy. Eur. J. Immunol. 2010, 40, 1011–1021. [Google Scholar] [CrossRef]
- Singh, R. Model Predicts That MKP1 and TAB1 Regulate p38α Nuclear Pulse and Its Basal Activity through Positive and Negative Feedback Loops in Response to IL-1. PLoS ONE 2016, 11, e0157572. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Das, P.K.; Ukil, A. Antileishmanial effect of 18β-glycyrrhetinic acid is mediated by Toll-like receptor-dependent canonical and noncanonical p38 activation. Antimicrob. Agents Chemother. 2015, 59, 2531–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, L.; Del Rio, L.; Butcher, B.A.; Mogensen, T.H.; Paludan, S.R.; Flavell, R.A.; Denkers, E.Y. p38 MAPK autophosphorylation drives macrophage IL-12 production during intracellular infection. J. Immunol. 2005, 174, 4178–4184. [Google Scholar] [CrossRef] [Green Version]
- Hallé, M.; Gomez, M.A.; Stuible, M.; Shimizu, H.; McMaster, W.R.; Olivier, M.; Tremblay, M.L. The Leishmania surface protease GP63 cleaves multiple intracellular proteins and actively participates in p38 mitogen-activated protein kinase inactivation. J. Biol. Chem. 2009, 284, 6893–6908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angé, M.; Castanares-Zapatero, D.; De Poortere, J.; Dufeys, C.; Courtoy, G.E.; Bouzin, C.; Quarck, R.; Bertrand, L.; Beauloye, C.; Horman, S. α1AMP-Activated Protein Kinase Protects against Lipopolysaccharide-Induced Endothelial Barrier Disruption via Junctional Reinforcement and Activation of the p38 MAPK/HSP27 Pathway. Int. J. Mol. Sci. 2020, 21, 5581. [Google Scholar] [CrossRef]
- Wang, S.; Li, M.; Yin, B.; Li, H.; Xiao, B.; Lǚ, K.; Huang, Z.; Li, S.; He, J.; Li, C. Shrimp TAB1 interacts with TAK1 and p38 and activates the host innate immune response to bacterial infection. Mol. Immunol. 2017, 88, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Makeeva, N.; Roomans, G.M.; Welsh, N. Role of TAB1 in nitric oxide-induced p38 activation in insulin-producing cells. Int. J. Biol. Sci. 2006, 3, 71–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makeeva, N.; Roomans, G.M.; Myers, J.W.; Welsh, N. Transforming growth factor-beta-activated protein kinase 1-binding protein (TAB)-1alpha, but not TAB1beta, mediates cytokine-induced p38 mitogen-activated protein kinase phosphorylation and cell death in insulin-producing cells. Endocrinology 2008, 149, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Ohkusu-Tsukada, K.; Tominaga, N.; Udono, H.; Yui, K. Regulation of the maintenance of peripheral T-cell anergy by TAB1-mediated p38 alpha activation. Mol. Cell. Biol. 2004, 24, 6957–6966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ten Hove, W.; Houben, L.A.; Raaijmakers, J.A.M.; Bracke, M.; Koenderman, L. Differential regulation of TNFα and GM-CSF induced activation of P38 MAPK in neutrophils and eosinophils. Mol. Immunol. 2007, 44, 2492–2496. [Google Scholar] [CrossRef] [Green Version]
- Lanna, A.; Henson, S.M.; Escors, D.; Akbar, A.N. The kinase p38 activated by the metabolic regulator AMPK and scaffold TAB1 drives the senescence of human T cells. Nat. Immunol. 2014, 15, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Lanna, A.; Gomes, D.C.; Muller-Durovic, B.; McDonnell, T.; Escors, D.; Gilroy, D.W.; Lee, J.H.; Karin, M.; Akbar, A.N. A sestrin-dependent Erk-Jnk-p38 MAPK activation complex inhibits immunity during aging. Nat. Immunol. 2017, 18, 354–363. [Google Scholar] [CrossRef] [Green Version]
- Richardson, L.S.; Taylor, R.N.; Menon, R. Reversible EMT and MET mediate amnion remodeling during pregnancy and labor. Sci. Signal. 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Seit-Nebi, A.; Davis, R.J.; Han, J. Multiple activation mechanisms of p38alpha mitogen-activated protein kinase. J. Biol. Chem. 2006, 281, 26225–26234. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.I.; Kwak, J.H.; Zachariah, M.; He, Y.; Wang, L.; Choi, M.E. TGF-β-activated kinase 1 and TAK1-binding protein 1 cooperate to mediate TGF-β1-induced MKK3-p38 MAPK activation and stimulation of type I collagen. Am. J. Physiol. Ren. Physiol. 2007, 292, F1471–F1478. [Google Scholar] [CrossRef]
- Ge, B.; Xiong, X.; Jing, Q.; Mosley, J.L.; Filose, A.; Bian, D.; Huang, S.; Han, J. TAB1β (Transforming Growth Factor-β-activated Protein Kinase 1-binding Protein 1β), a Novel Splicing Variant of TAB1 That Interacts with p38α but Not TAK1. J. Biol. Chem. 2003, 278, 2286–2293. [Google Scholar] [CrossRef] [Green Version]
- Xin, F.; Wu, J. Crystal structure of the p38α MAP kinase in complex with a docking peptide from TAB1. Sci. China Life Sci. 2013, 56, 653–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvador, J.M.; Mittelstadt, P.R.; Belova, G.I.; Fornace, A.J.; Ashwell, J.D. The autoimmune suppressor Gadd45α inhibits the T cell alternative p38 activation pathway. Nat. Immunol. 2005, 6, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Dorn, T.; Kuhn, U.; Bungartz, G.; Stiller, S.; Bauer, M.; Ellwart, J.; Peters, T.; Scharffetter-Kochanek, K.; Semmrich, M.; Laschinger, M.; et al. RhoH is important for positive thymocyte selection and T-cell receptor signaling. Blood 2007, 109, 2346–2355. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Humphries, L.A.; Tomassian, T.; Mittelstadt, P.; Zhang, M.; Miceli, M.C. Scaffold protein Dlgh1 coordinates alternative p38 kinase activation, directing T cell receptor signals toward NFAT but not NF-kappaB transcription factors. Nat. Immunol. 2007, 8, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Yi, P.; Wang, X.; Zhang, B.; Jie, Z.; Soong, L.; Sun, J. Retinoic Acid Modulates Hyperactive T Cell Responses and Protects Vitamin A–Deficient Mice against Persistent Lymphocytic Choriomeningitis Virus Infection. J. Immunol. 2020, 204, 2984–2994. [Google Scholar] [CrossRef]
- Hirata, S.; Fukamachi, T.; Sakano, H.; Tarora, A.; Saito, H.; Kobayashi, H. Extracellular acidic environments induce phosphorylation of ZAP-70 in Jurkat T cells. Immunol. Lett. 2008, 115, 105–109. [Google Scholar] [CrossRef]
- Giardino Torchia, M.L.; Dutta, D.; Mittelstadt, P.R.; Guha, J.; Gaida, M.M.; Fish, K.; Barr, V.A.; Akpan, I.O.; Samelson, L.E.; Tagad, H.D.; et al. Intensity and duration of TCR signaling is limited by p38 phosphorylation of ZAP-70(T293) and destabilization of the signalosome. Proc. Natl. Acad. Sci. USA 2018, 115, 2174–2179. [Google Scholar] [CrossRef] [Green Version]
- Jun, J.E.; Kulhanek, K.R.; Chen, H.; Chakraborty, A.; Roose, J.P. Alternative ZAP70-p38 signals prime a classical p38 pathway through LAT and SOS to support regulatory T cell differentiation. Sci. Signal. 2019, 12. [Google Scholar] [CrossRef]
- Liu, J.; Guo, K.; Hu, L.; Luo, T.; Ma, Y.; Zhang, Y.; Lai, W.; Guo, Z. ZAP70 deficiency promotes reverse cholesterol transport through MAPK/ERK pathway in Jurkat cell. Mol. Immunol. 2019, 107, 21–28. [Google Scholar] [CrossRef]


{kind=link}
| Compound | Isoform Specificity | Diseases Targeted | Identifier |
|---|---|---|---|
| AZD7624 | p8α, p38β | Endotoxin-induced inflammation, COPD | NCT01937338 NCT02238483 |
| LY2228820 (Ralimetinib) | p38 pan-inhibition | Ovarian cancer, glioblastoma (both concomitant), metastatic breast cancer | NCT02322853 NCT02364206 NCT01663857 NCT01393990 |
| LY3007113 | p38 pan-inhibition | Metastatic cancer | NCT01463631 |
| VX-745 (Neflamapimod) | p38α | Alzheimer’s disease, Huntington disease, Lewy body dementia | NCT03980938 NCT04001517 NCT03402659 NCT03435861 |
| VX-702 | p38α | Rheumatoid arthritis | NCT00395577 NCT00205478 |
| PH-797804 | p38α | Rheumatoid arthritis, COPD | NCT01321463 NCT00559910 NCT01589614 |
| SB681323 (Dilmapimod) | p38α | Neuropathic pain, COPD, ALI/ARDS, Coronary heart disease | NCT00134693 NCT00564746 NCT00390845 NCT00144859 NCT00320450 NCT00996840 NCT00291902 |
| Losmapimod GW856553X or GSK-AHAB (Losmapimod) | p38 pan-inhibition | Acute coronary syndrome, COPD, neuropathic pain, SARS-CoV-2, atherosclerosis, acute coronary syndrome, focal segmental glomerulosclerosis, facioscapulohumeral muscular dystrophy | NCT04264442 NCT04511819 NCT02000440 NCT02299375 NCT04003974 NCT01541852 NCT01756495 NCT02145468 NCT01218126 NCT00633022 |
| BMS-582949 | p38 pan-inhibition | Arterial inflammation, atherosclerosis | NCT00162292 NCT00399906 |
| ARRY-371797 | p38α | LMNA-related dilated cardiomyopathy, rheumatoid arthritis, osteoarthritis of the knee, ankylosing spondylitis | NCT02351856 NCT03439514 NCT00729209 NCT01366014 NCT00811499 |
| PF-03715455 | p38α | Asthma, COPD | NCT02219048 NCT02366637 |
| BIRB 796 (Doramapimod) | p38 pan-inhibition | Crohn’s disease, plaque-type psoriasis, rheumatoid arthritis, endotoxin-induced inflammation | NCT02214888 NCT02209753 NCT02209792 NCT02209779 NCT02211170 |
| SCIO-469 (Talapimod) | p38α | Rheumatoid arthritis, multiple myeloma | NCT00095680 NCT00087867 NCT00043732 NCT00508768 |
| Pirfenidone | p38γ | Idiopathic pulmonary fibrosis | NCT03208933 |
| BCT-197 (Acumapimod) | p38α | COPD | NCT01332097 NCT02700919 |
| Disease | Pathological Outcome | References | |
|---|---|---|---|
| Cardiovascular | Myocardial infarction/Ischemia reperfusion | Induces overexpression of pro-inflammatory cytokines like IL-6, TNF-α, and IL-1β, and elevates intracellular calcium (Ca2+i) levels, inflammation, and apoptosis | [70,83,84,101,102,103,104,105,106,107,108] |
| Diabetic cardiomyopathy | Overexpression of pro-inflammatory cytokines induces cardiomyocyte apoptosis | [109,110] | |
| Atherosclerosis | Promotes ANG-II-dependent MerTK shedding in macrophages resulting in defective efferocytosis and, in turn, induces plaque progression | [111,112,113,114,115] | |
| Pulmonary | Chronic obstructive pulmonary disease (COPD) | Activates transcription factors and induces overexpression of pro-inflammatory cytokines and chemokines, amplifying lung inflammation | [116,117,118,119,120,121] |
| Acute respiratory distress syndrome (ARDS) | Induces decreased corticosteroid responsiveness, alveolar macrophage-induced impairment of respiratory function, and overexpression of pro-inflammatory cytokines like IL-6, IL-8, TNF-α and IL-1β | [63,101,122,123] | |
| Acute lung injury (ALI) | Induces overexpression of pro-inflammatory cytokines like IL-6, TNF-α, and IL-1β, and cell apoptosis | [60,123,124,125,126] | |
| Viral infections and SARS-CoV-2 | Induction of type 1 interferons, expression of IL-12, promotion of viral replication, expression of pro-inflammatory cytokines resulting in inflammation, thrombosis, and vasoconstriction in SARS-CoV-2 | [60,127,128,129] | |
| Oncology | Non-small cell lung cancer (NSCLC) | Enhances proliferation, migration, chemoresistance, and inflammatory cytokine expression | [130,131,132,133,134,135,136] |
| Head and neck small cell carcinoma (HNSCC) | Inhibition of p38 increases HNSCC sensitivity to cisplatin, cannabinoids promote progressive HNSCC via p38 [42], increases mRNA stability via MK2, p38 isoforms as a diagnostic of HNSCC, and regulates angiogenesis and lymphangiogenesis | [137,138,139,140] | |
| Breast cancer | Elevated p38δ levels promote cell detachment, migration, invasion, and increased metastatic lesions, and inhibition of p38 triggers DNA damage and tumor cell death | [133,141,142] | |
| Bladder cancer | Induces cell invasion and metastasis by increasing MMP-2 and MMP-9 activity | [135,143] | |
| Neurodegenerative | Alzheimer’s disease | Elevated p-p38 levels progress neuroinflammation tau phosphorylation, neurotoxicity, and synaptic dysfunction | [144,145,146,147] |
| Parkinson’s disease | p-p38 overload induces a COX-2-mediated inflammation and subsequent dopaminergic neuron degeneration | [148,149,150] | |
| Amyotrophic lateral sclerosis (ALS) | Induces defects in axonal retrograde transport of signaling endosomes | [151,152,153] | |
| Spinal muscular atrophy | Induces p38 MAPK-dependent p53 phosphorylation leading to selective degeneration of motor neurons | [154] | |
| Ocular | Age-related macular degeneration (AMD) | Induces VEGF expression and angiogenesis, regulates Ang-II-mediated MMP-2 and MMP-14, basigin expression, and extracellular matrix accumulation in AMD | [155,156] |
| Diabetic retinopathy | ASK/p38 NLRP3 inflammasome signaling, retinal angiogenesis, retinal endothelial cell dysfunction, inner-blood-retinal-barrier leakage | [110,157,158,159,160,161] | |
| Glaucoma | Induces anterograde transport degradation and axon degeneration in the optic nerve | [162,163] |
| Disease | Mechanism of p38 Autophosphorylation | Model | Specific Cell or Animal Line |
|---|---|---|---|
| Cardiovascular ischemia and reperfusion | TAB1-mediated | Murine in vivo [70,80,83,104,105] | MKK3−/− [80,105]; C57BL/6 [80,104]; Sprague Dawley [80]; Wistar [83]; TAB1 KI [70] |
| Murine in vitro [70,72,83,104,105,183] | H9c2 [105]; Sprague Dawley [83,104,183]; Wistar [83]; C57BL/6 [70,72] | ||
| Human in vitro [70,83,84,104,108] | HEK293 [70,83,104,108] | ||
| Structural modeling [66] | |||
| Myocardial infarction, amyloidosis, and cardiomyopathy | TAB1-mediated | Murine in vivo [107] | Sprague Dawley |
| Murine in vitro [81,107,184] | H9c2 [107]; Wistar [184] | ||
| Human in vitro [184] | Patient heart | ||
| Zebrafish in vivo [185] | |||
| General inflammation and cancer | TAB1-mediated | Murine in vivo [31,82,186] | BALB/c [186]; CD1/CD1 [31]; C57BL/6, Itch−/− [82] |
| Murine in vitro [31,82,186] | Vβ8.1, OT-II [186]; TAB1−/− [31]; C57BL/6, Itch−/− [82] | ||
| Human in vitro [31,32,87] | HUVEC [31,32,87]; HEK293 [31]; HDMEC [87] | ||
| Structural modeling [89,187] | |||
| Parasitic infection | TAB1-mediated | Murine in vivo [188] | BALB/c |
| Murine in vitro [188,189,190] | RAW264.9 [188]; MKK3−/− [189]; BALB/c [190] | ||
| Viral infection | TAB1-mediated | Murine in vitro [128] | C57BL/6, BC-1 |
| Human in vitro [127] | Huh7.5.1, HEK293, patient liver | ||
| Bacterial infection | TAB1-mediated | Human in vitro [191] | HPMEC |
| Shrimp [192] | |||
| Diabetes | TAB1-mediated | Murine in vitro [193,194] | β-TC6 [193,194]; Sprague Dawley, NMRI [194] |
| Human in vitro [194] | Islet | ||
| Leukocyte dysfunction | TAB1-mediated | Murine in vivo [195] | Vβ8.1 |
| Murine in vitro [195] | 2B4 | ||
| Human in vitro [196,197,198] | Patient blood | ||
| Pregnancy complications | TAB1-mediated | Murine in vitro [199] | CD-1 |
| Human in vitro [79,199] | Patient placenta | ||
| Other | TAB1-mediated | Murine in vitro [200,201] | MKK3−/−/6−/− [200]; MKK3−/− [201] |
| Human in vitro [73,78,85,202] | HEK293 [73,78,85,202]; MDA231 [202] | ||
| Structural modeling [203] | |||
| Immune system (T-Cell) modulation | Zap70-mediated | Murine in vivo [88] | P116 |
| Murine in vitro [204,205,206,207] | Gadd45a−/− [204]; CD4SP [205]; C57BL/6 [206,207] | ||
| Human in vitro [88,208,209,210,211] | Jurkat, P116 | ||
| Chicken in vitro [210] | DT40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burton, J.C.; Antoniades, W.; Okalova, J.; Roos, M.M.; Grimsey, N.J. Atypical p38 Signaling, Activation, and Implications for Disease. Int. J. Mol. Sci. 2021, 22, 4183. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084183
Burton JC, Antoniades W, Okalova J, Roos MM, Grimsey NJ. Atypical p38 Signaling, Activation, and Implications for Disease. International Journal of Molecular Sciences. 2021; 22(8):4183. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084183
Chicago/Turabian StyleBurton, Jeremy C., William Antoniades, Jennifer Okalova, Morgan M. Roos, and Neil J. Grimsey. 2021. "Atypical p38 Signaling, Activation, and Implications for Disease" International Journal of Molecular Sciences 22, no. 8: 4183. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084183