
Genetic Variants in Transcription Factor Binding Sites in Humans: Triggered by Natural Selection and Triggers of Diseases

 , ,
, ,  and
and
Abstract
:1. Introduction
- (1)
- TFBS polymorphisms comprise only 8% of the genome polymorphisms but 31% of the trait-associated polymorphisms identified by GWAS [8].
- (2)
- Up to 21.6% of variants affecting gene expression overlap annotated TFBSs [9].
- (3)
- Polymorphisms leading to the differential binding of transcription factors are highly enriched in the set of causal variants reported for traits across several independent studies [10].
- (4)
- Chromatin immunoprecipitation followed by high-throughput sequencing (ChIP-seq) has demonstrated the extensive contributions of genetic variations to transcription factor binding and a significant correlation between nucleotide changes affecting transcription factor binding and gene expression [11].
2. Genomic Features of TFBSs and Genetic Variants of TFBSs
3. Mechanisms Linking TFBS Variations and Differential Gene Expression
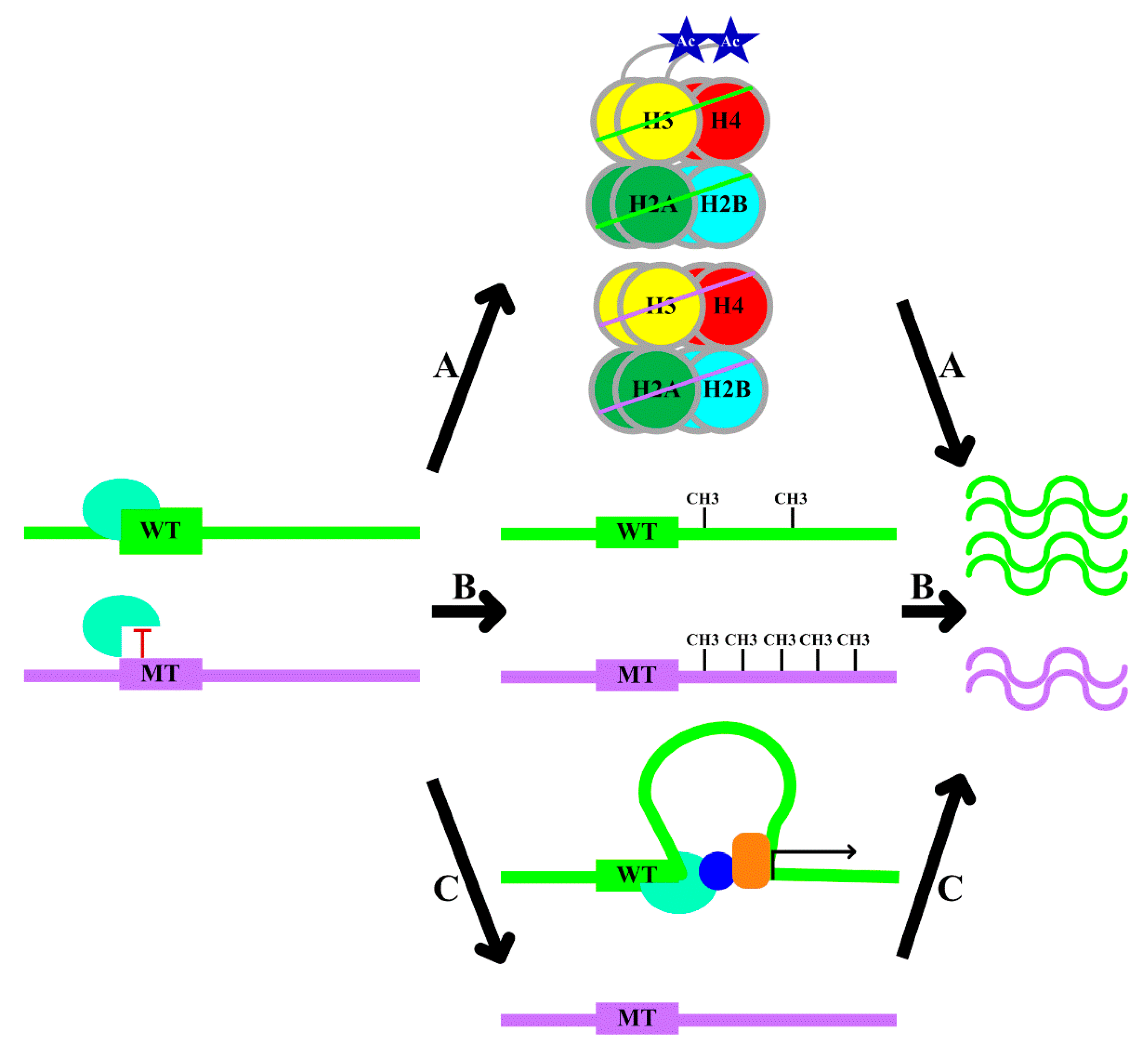
- A.
- B.
- Local DNA methylation: transcription factor–DNA binding leads to an altered local DNA methylation profile (Figure 1, Pathway B) [20]. Through modulating DNA methylation, transcription factor binding exerts downstream effects on genome regulation. Thus, the consideration of DNA methylation data in the interpretation of the functional role of variants is recommended [20].
- C.
- Changes in chromatin conformation: several studies have utilized chromosome conformation capture (Hi-C) datasets to demonstrate that transcription factors might drive topological genome reorganization and change the structure of enhancer-promoter loops and recruiting other co-factors, thereby contributing to gene regulation (Figure 1, Pathway C) [21].
- A.
- rs2886870 disrupts a nuclear factor-κB (NF-κB) binding site and is associated with H3K27ac levels and C3orf59 mRNA expression (Table 1) [22]. rs4784227 is a breast cancer risk-associated polymorphism. The risk allele of rs4784227 enhances FOXA1 binding, decreases H3K9Ac levels, inhibits the expression of TOX3, and therefore, promotes the proliferation of breast cancer cells (Table 1) [23]. rs6983267 is associated with numerous malignancies. The risk allele of rs6983267 is associated with enhanced TCF4 binding and more prominent histone modifications and drives elevated c-MYC expression (Table 1) [24].
- B.
- The rs2240032 allele specifically binds SMAD3, affects the methylation of the promoter region, and influences RAD50 and IL4 expression (Table 1) [25]. Similarly, the rs612529 risk allele decreases binding of YY1 and PU.1, is associated with the hypermethylation of the promoter, specifically downregulates SIRL-1 expression, and increases the risk of atopic dermatitis (Table 1) [26]. A rare variant at chr22:24,059,610 disrupts the UA4 binding motif, increases the methylation levels at the promoter of the nearby GUSBP11 gene, and reduces the expression of GUSBP11 (Table 1) [20].
- C.
- The rs12913832 risk allele increases the binding of HLTF, LEF1, and MITF to the enhancer region and enhances chromatin loop formation, and increases OCA2 expression and, thus, pigmentation (Table 1) [27]. The C allele of rs13228237 causes increased binding of ZNF143, leads to an increase in chromatin loop formation between the first intron of the ZC3HAV1 gene and two distal regulatory elements, and increases ZC3HAV1 expression (Table 1) [28]. The presence of the G allele of rs2802292 creates an HSF1 binding site, which induces promoter–enhancer interaction via chromatin looping, thereby fostering FOXO3 expression (Table 1) [29].
4. Challenges of Investigating Genetic Variants in TFBSs
5. Origin of TFBS Genetic Variants: Natural Selection
- A.
- Infection: rs139999735 is associated with APAF1-interacting protein (APIP), which inhibits pyroptosis and apoptosis, both of which are responses to Salmonella infection (Table 2). Individuals homozygous for rs139999735 show decreased APIP expression and, therefore, might generate a better response to Salmonella infection. Interestingly, rs139999735 displays a higher allelic frequency in Africans (0.34) than in Asians (0.11) and Europeans (0.12), suggesting the natural selection of rs139999735 in Africans [67]. The ACKR1-null polymorphism rs2814778 located in ACKR1, which disrupts the binding of the transcription factor GATA binding protein 1 (GATA1), is associated with reduced susceptibility to malaria infections caused by Plasmodium vivax (Table 2). The associated protective effects may explain the spread of the ACKR1-null polymorphism by natural selection in areas of relatively high malaria transmission, such as central, western, and southeastern Africa, in which the prevalence reaches almost 100% [68]. Another well-studied example is IFN-γ + 874. This risk allele fails to provide a binding site for the transcription factor NF-κB. As NF-κB induces IFN-γ expression, the risk allele correlates with reduced IFN-γ expression and susceptibility to tuberculosis (Table 2). Because only the more resistant individuals survived and reproduced, over successive generations of selective pressure from tuberculosis, the frequency of the risk genotype decreased, and eventually, the cases of tuberculosis in the white population decreased. Consistent with these observations, the frequency of the risk genotype is much higher in South African populations (47%) than in Sicilian (26%) and Spanish populations (28%) [69].
- B.
- Radiation: rs201097793 and rs2279744 both illustrate the molecular adaptation of modern human populations to ultraviolet radiation. rs201097793 is located in a TFBS and is associated with MC1R (Table 2). Interestingly, rs201097793 has a higher allelic frequency in Africans (0.70) and Asians (0.64) than in Europeans (0.17) [67]. MC1R is known to be associated with pigmentation in humans and is maintained by purifying selection in low-latitude, high-ultraviolet-radiation regions, protecting against folate photolysis [70]. In line with this idea, the rs201097793 allele associated with darker skin pigmentation exhibits a high frequency in Africans and Asians. As regards rs2279744 (SNP309) in MDM2, MDM2 counteracts p53 in a “yin and yang” fashion to regulate embryo implantation [71]. A single-nucleotide change from T to G in rs2279744 creates a binding site for the transcription factor SP1 [71]. Consistent with this observation, homozygotes for the G allele express more MDM2 than homozygotes for the T allele [72]. Modern humans migrating northwards to regions with lower ultraviolet radiation required less p53 to avert the adverse effects of p53 hyperactivity, such as embryonic death. Correspondingly, the population data in both East Asia and Europe show that MDM2 rs2279744 G homozygotes are selected for by low ultraviolet radiation exposure (Table 2) [71].
- C.
- Taste: Taste perception has been critical in evolution, especially for the detection of toxins. rs139938620 in TAS1R3, a sweet receptor, shows a high allelic frequency in Asians (0.79) compared with other populations (Table 2). TAS1R3 is a component of the dimeric protein TAS1R1/TAS1R3, which is the umami taste receptor, and the umami taste is a common feature of many foods in Asia. As a result, it is reasonable to speculate that this variant is beneficial for toxin detection in Asians and is, thus, selected for [67].
- D.
- Water conservation: A well-studied example is rs16846053. The minor allele of rs16846053 in SLC4A10 that predisposes individuals to increased plasma osmolality—the reduced central sensing of water loss and/or renal water conservation—is underrepresented in the African population (minor allele frequency 0.02) compared with the European population (minor allele frequency 0.10) (Table 2) [73].
6. Consequences of TFBS Genetic Variants: Disease Susceptibility
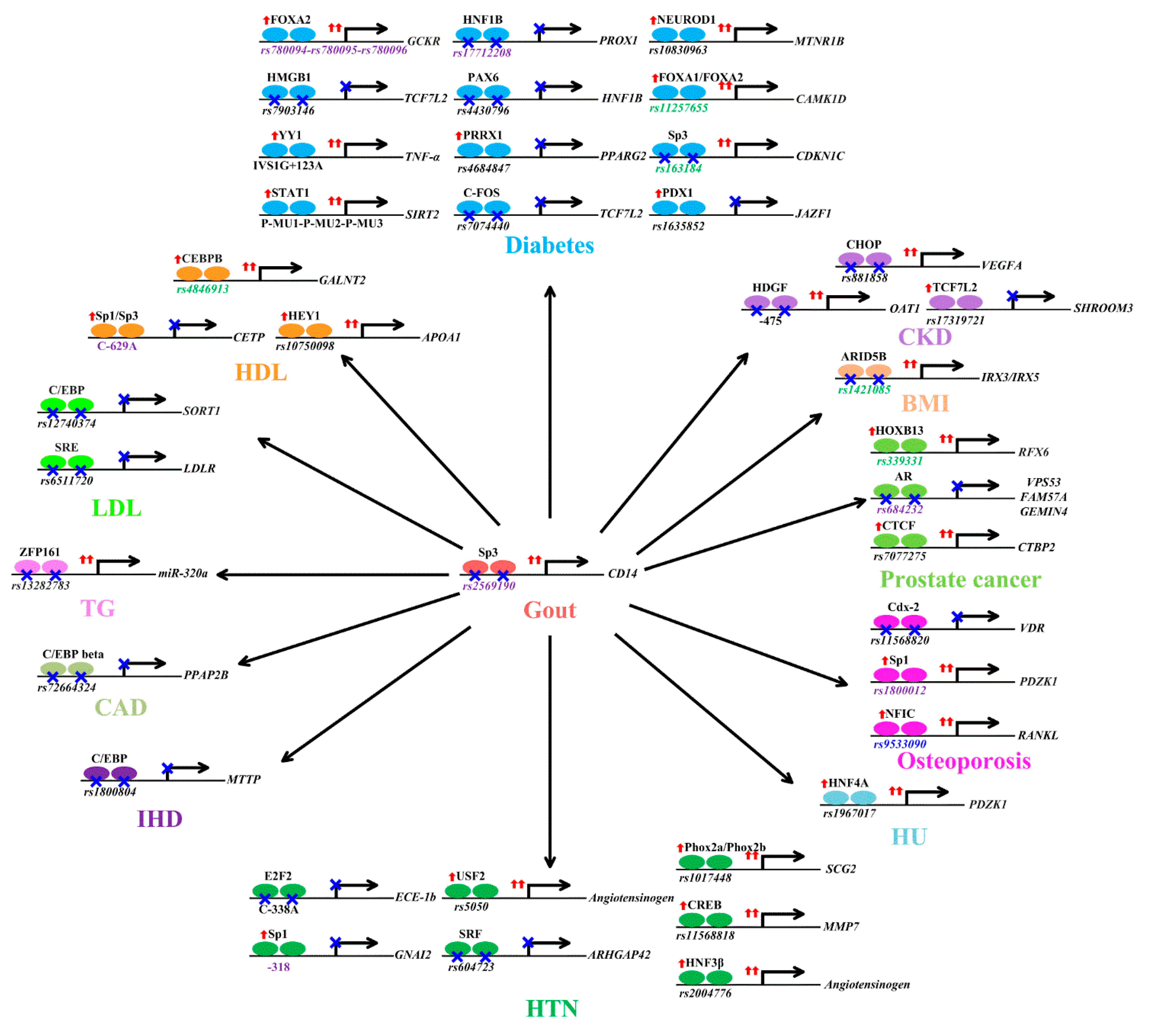
6.1. Gout
6.2. Body Mass Index
6.3. Chronic Kidney Disease
6.4. Diabetes
6.5. Dyslipidemia
6.6. Heart Disease
6.7. Hypertension
6.8. Hyperuricemia
6.9. Osteoporosis
6.10. Prostate Cancer
7. Consequences of TFBS Genetic Variants: Treatment Response
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hindorff, L.A.; Sethupathy, P.; Junkins, H.A.; Ramos, E.M.; Mehta, J.P.; Collins, F.S.; Manolio, T.A. Potential etiologic and functional implications of genome-wide association loci for human diseases and traits. Proc. Natl. Acad. Sci. USA 2009, 106, 9362–9367. [Google Scholar] [CrossRef] [Green Version]
- Ponomarenko, M.; Rasskazov, D.; Chadaeva, I.; Sharypova, E.; Drachkova, I.; Oshchepkov, D.; Ponomarenko, P.; Savinkova, L.; Oshchepkova, E.; Nazarenko, M.; et al. Candidate SNP Markers of Atherogenesis Significantly Shifting the Affinity of TATA-Binding Protein for Human Gene Promoters show stabilizing Natural Selection as a Sum of Neutral Drift Accelerating Atherogenesis and Directional Natural Selection Slowing It. Int. J. Mol. Sci. 2020, 21, 1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhou, T.; Wunderlich, Z.; Maurano, M.T.; DePace, A.H.; Nuzhdin, S.V.; Rohs, R. Analysis of Genetic Variation Indicates DNA Shape Involvement in Purifying Selection. Mol. Biol. Evol. 2018, 35, 1958–1967. [Google Scholar] [CrossRef]
- Suter, D.M. Transcription Factors and DNA Play Hide and Seek. Trends. Cell. Biol. 2020, 30, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Johnston, A.D.; Simões-Pires, C.A.; Thompson, T.V.; Suzuki, M.; Greally, J.M. Functional genetic variants can mediate their regulatory effects through alteration of transcription factor binding. Nat. Commun. 2019, 10, 3472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graae, L.; Paddock, S.; Belin, A.C. ReMo-SNPs: A new software tool for identification of polymorphisms in regions and motifs genome-wide. Genet. Res. (Camb.) 2015, 97, e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuğrul, M.; Paixão, T.; Barton, N.H.; Tkačik, G. Dynamics of Transcription Factor Binding Site Evolution. Plos. Genet. 2015, 11, e1005639. [Google Scholar] [CrossRef] [Green Version]
- Nishizaki, S.S.; Ng, N.; Dong, S.; Porter, R.S.; Morterud, C.; Williams, C.; Asman, C.; Switzenberg, J.A.; Boyle, A.P. Predicting the effects of SNPs on transcription factor binding affinity. Bioinformatics 2020, 36, 364–372. [Google Scholar] [CrossRef]
- Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; Abecasis, G.R.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar]
- Yan, J.; Qiu, Y.; Ribeiro Dos Santos, A.M.; Yin, Y.; Li, Y.E.; Vinckier, N.; Nariai, N.; Benaglio, P.; Raman, A.; Li, X.; et al. Systematic analysis of binding of transcription factors to noncoding variants. Nature 2021, 591, 147–151. [Google Scholar] [CrossRef]
- Kasowski, M.; Grubert, F.; Heffelfinger, C.; Hariharan, M.; Asabere, A.; Waszak, S.M.; Habegger, L.; Rozowsky, J.; Shi, M.; Urban, A.E.; et al. Variation in transcription factor binding among humans. Science 2010, 328, 232–235. [Google Scholar] [CrossRef] [Green Version]
- Consortium, E.P. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Samee, M.A.H.; Bruneau, B.G.; Pollard, K.S. A De Novo Shape Motif Discovery Algorithm Reveals Preferences of Transcription Factors for DNA Shape Beyond Sequence Motifs. Cell. Syst. 2019, 8, 27–42.e26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, B.; Rohs, R. Relationship between histone modifications and transcription factor binding is protein family specific. Genome. Res. 2018, 28, 321–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eickhardt, E.; Damm, A.T.; Grove, J.; Boerglum, A.D.; Lescai, F. Estimating the functional impact of INDELs in transcription factor binding sites: A genome-wide landscape. bioRxiv. 2016. [Google Scholar] [CrossRef] [Green Version]
- de Santiago, I.; Liu, W.; Yuan, K.; O’Reilly, M.; Chilamakuri, C.S.; Ponder, B.A.; Meyer, K.B.; Markowetz, F. BaalChIP: Bayesian analysis of allele-specific transcription factor binding in cancer genomes. Genome. Biol. 2017, 18, 39. [Google Scholar] [CrossRef] [Green Version]
- Kasowski, M.; Kyriazopoulou-Panagiotopoulou, S.; Grubert, F.; Zaugg, J.B.; Kundaje, A.; Liu, Y.; Boyle, A.P.; Zhang, Q.C.; Zakharia, F.; Spacek, D.V.; et al. Extensive variation in chromatin states across humans. Science 2013, 342, 750–752. [Google Scholar] [CrossRef] [Green Version]
- Benveniste, D.; Sonntag, H.J.; Sanguinetti, G.; Sproul, D. Transcription factor binding predicts histone modifications in human cell lines. Proc. Natl. Acad. Sci. USA 2014, 111, 13367–13372. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, M.; Daujat, S.; Schneider, R. Lateral Thinking: How Histone Modifications Regulate Gene Expression. Trends. Genet. 2016, 32, 42–56. [Google Scholar] [CrossRef] [Green Version]
- Martin-Trujillo, A.; Patel, N.; Richter, F.; Jadhav, B.; Garg, P.; Morton, S.U.; McKean, D.M.; DePalma, S.R.; Goldmuntz, E.; Gruber, D.; et al. Rare genetic variation at transcription factor binding sites modulates local DNA methylation profiles. PLoS Genet. 2020, 16, e1009189. [Google Scholar] [CrossRef] [PubMed]
- Di Giammartino, D.C.; Polyzos, A.; Apostolou, E. Transcription factors: Building hubs in the 3D space. Cell. Cycle 2020, 19, 2395–2410. [Google Scholar] [CrossRef] [PubMed]
- McVicker, G.; van de Geijn, B.; Degner, J.F.; Cain, C.E.; Banovich, N.E.; Raj, A.; Lewellen, N.; Myrthil, M.; Gilad, Y.; Pritchard, J.K. Identification of genetic variants that affect histone modifications in human cells. Science 2013, 342, 747–749. [Google Scholar] [CrossRef] [Green Version]
- Cowper-Sal lari, R.; Zhang, X.; Wright, J.B.; Bailey, S.D.; Cole, M.D.; Eeckhoute, J.; Moore, J.H.; Lupien, M. Breast cancer risk-associated SNPs modulate the affinity of chromatin for FOXA1 and alter gene expression. Nat. Genet. 2012, 44, 1191–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, J.B.; Brown, S.J.; Cole, M.D. Upregulation of c-MYC in cis through a large chromatin loop linked to a cancer risk-associated single-nucleotide polymorphism in colorectal cancer cells. Mol. Cell. Biol. 2010, 30, 1411–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schieck, M.; Sharma, V.; Michel, S.; Toncheva, A.A.; Worth, L.; Potaczek, D.P.; Genuneit, J.; Kretschmer, A.; Depner, M.; Dalphin, J.C.; et al. A polymorphism in the TH 2 locus control region is associated with changes in DNA methylation and gene expression. Allergy 2014, 69, 1171–1180. [Google Scholar] [CrossRef]
- Kumar, D.; Puan, K.J.; Andiappan, A.K.; Lee, B.; Westerlaken, G.H.; Haase, D.; Melchiotti, R.; Li, Z.; Yusof, N.; Lum, J.; et al. A functional SNP associated with atopic dermatitis controls cell type-specific methylation of the VSTM1 gene locus. Genome Med. 2017, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Visser, M.; Kayser, M.; Palstra, R.J. HERC2 rs12913832 modulates human pigmentation by attenuating chromatin-loop formation between a long-range enhancer and the OCA2 promoter. Genome Res. 2012, 22, 446–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, S.D.; Zhang, X.; Desai, K.; Aid, M.; Corradin, O.; Cowper-Sal Lari, R.; Akhtar-Zaidi, B.; Scacheri, P.C.; Haibe-Kains, B.; Lupien, M. ZNF143 provides sequence specificity to secure chromatin interactions at gene promoters. Nat. Commun. 2015, 2, 6186. [Google Scholar] [CrossRef]
- Sanese, P.; Forte, G.; Disciglio, V.; Grossi, V.; Simone, C. FOXO3 on the Road to Longevity: Lessons from SNPs and Chromatin Hubs. Comput. Struct. Biotechnol. J. 2019, 17, 737–745. [Google Scholar] [CrossRef]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The Human Transcription Factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Ye, Z.; Wang, L. Genome Wide Approaches to Identify Protein-DNA Interactions. Curr. Med. Chem. 2019, 26, 7641–7654. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Kudron, M.M.; Victorsen, A.; Gao, J.; Ammouri, H.N.; Navarro, F.C.P.; Gevirtzman, L.; Waterston, R.H.; White, K.P.; Reinke, V.; et al. To mock or not: A comprehensive comparison of mock IP and DNA input for ChIP-seq. Nucleic. Acids. Res. 2021, 49, e17. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.; Singh, R.S.; Hogan, D.; Vidhyasagar, V.; Yang, S.; Yeuk Wah Chung, I.; Kusalik, A.; Dmitriev, O.Y.; Cygler, M.; Wu, Y. The KH domain facilitates the substrate specificity and unwinding processivity of DDX43 helicase. J. Biol. Chem. 2020. [Google Scholar] [CrossRef]
- Shokri, L.; Inukai, S.; Hafner, A.; Weinand, K.; Hens, K.; Vedenko, A.; Gisselbrecht, S.S.; Dainese, R.; Bischof, J.; Furger, E.; et al. A Comprehensive Drosophila melanogaster Transcription Factor Interactome. Cell. Rep. 2019, 27, 955–970.e957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, A.E.; Kugel, J.F.; Goodrich, J.A. Single molecule microscopy reveals mechanistic insight into RNA polymerase II preinitiation complex assembly and transcriptional activity. Nucleic. Acids. Res. 2016, 44, 7132–7143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, N.; Suyama, M. An integrated analysis of public genomic data unveils a possible functional mechanism of psoriasis risk via a long-range ERRFI1 enhancer. BMC. Med. Genom. 2020, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Moradifard, S.; Saghiri, R.; Ehsani, P.; Mirkhani, F.; Ebrahimi-Rad, M. A preliminary computational outputs versus experimental results: Application of sTRAP, a biophysical tool for the analysis of SNPs of transcription factor-binding sites. Mol. Genet. Genomic. Med. 2020, 8, e1219. [Google Scholar] [CrossRef] [Green Version]
- Stalke, A.; Pfister, E.D.; Baumann, U.; Illig, T.; Reischl, E.; Sandbothe, M.; Vajen, B.; Huge, N.; Schlegelberger, B.; von Neuhoff, N.; et al. MTF1 binds to metal-responsive element e within the ATP7B promoter and is a strong candidate in regulating the ATP7B expression. Ann. Hum. Genet. 2020, 84, 195–200. [Google Scholar] [CrossRef] [Green Version]
- de Smith, A.J.; Walsh, K.M.; Francis, S.S.; Zhang, C.; Hansen, H.M.; Smirnov, I.; Morimoto, L.; Whitehead, T.P.; Kang, A.; Shao, X.; et al. BMI1 enhancer polymorphism underlies chromosome 10p12.31 association with childhood acute lymphoblastic leukemia. Int. J. Cancer. 2018, 143, 2647–2658. [Google Scholar] [CrossRef] [Green Version]
- Santana-Garcia, W.; Rocha-Acevedo, M.; Ramirez-Navarro, L.; Mbouamboua, Y.; Thieffry, D.; Thomas-Chollier, M.; Contreras-Moreira, B.; van Helden, J.; Medina-Rivera, A. RSAT variation-tools: An accessible and flexible framework to predict the impact of regulatory variants on transcription factor binding. Comput. Struct. Biotechnol. J. 2019, 17, 1415–1428. [Google Scholar] [CrossRef]
- Siggers, T.; Gordân, R. Protein-DNA binding: Complexities and multi-protein codes. Nucleic. Acids. Res. 2014, 42, 2099–2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, Z.; Roy, B.; Chang, Y.K.; Granas, D.; Stormo, G.D. Measuring quantitative effects of methylation on transcription factor-DNA binding affinity. Sci. Adv. 2017, 3, eaao1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, A.A.; Pritchard, J.K.; Gilad, Y. The genetic and mechanistic basis for variation in gene regulation. PLoS Genet. 2015, 11, e1004857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledo, N.; Ko, Y.A.; Park, A.S.; Kang, H.M.; Han, S.Y.; Choi, P.; Susztak, K. Functional genomic annotation of genetic risk loci highlights inflammation and epithelial biology networks in CKD. J. Am. Soc. Nephrol. 2015, 26, 692–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurano, M.T.; Haugen, E.; Sandstrom, R.; Vierstra, J.; Shafer, A.; Kaul, R.; Stamatoyannopoulos, J.A. Large-scale identification of sequence variants influencing human transcription factor occupancy in vivo. Nat. Genet. 2015, 47, 1393–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polavarapu, N.; Mariño-Ramírez, L.; Landsman, D.; McDonald, J.F.; Jordan, I.K. Evolutionary rates and patterns for human transcription factor binding sites derived from repetitive DNA. BMC Genom. 2008, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, H.M.; Kundaje, A.; Boyle, A.P. The ENCODE Blacklist: Identification of Problematic Regions of the Genome. Sci. Rep. 2019, 9, 9354. [Google Scholar] [CrossRef] [Green Version]
- Cusanovich, D.A.; Pavlovic, B.; Pritchard, J.K.; Gilad, Y. The functional consequences of variation in transcription factor binding. PLoS. Genet. 2014, 10, e1004226. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, X.; Zheng, Z.; Ma, T.; Long, H.; Cheng, H.; Fang, M.; Gong, J.; Li, X.; Zhao, S.; et al. Genome-wide analysis of expression QTL (eQTL) and allele-specific expression (ASE) in pig muscle identifies candidate genes for meat quality traits. Genet. Sel. Evol. 2020, 52, 59. [Google Scholar] [CrossRef]
- Mumbach, M.R.; Satpathy, A.T.; Boyle, E.A.; Dai, C.; Gowen, B.G.; Cho, S.W.; Nguyen, M.L.; Rubin, A.J.; Granja, J.M.; Kazane, K.R.; et al. Enhancer connectome in primary human cells identifies target genes of disease-associated DNA elements. Nat. Genet. 2017, 49, 1602–1612. [Google Scholar] [CrossRef] [Green Version]
- Miguel-Escalada, I.; Bonàs-Guarch, S.; Cebola, I.; Ponsa-Cobas, J.; Mendieta-Esteban, J.; Atla, G.; Javierre, B.M.; Rolando, D.M.Y.; Farabella, I.; Morgan, C.C.; et al. Human pancreatic islet three-dimensional chromatin architecture provides insights into the genetics of type 2 diabetes. Nat. Genet. 2019, 51, 1137–1148. [Google Scholar] [CrossRef] [PubMed]
- Ballester, B.; Medina-Rivera, A.; Schmidt, D.; Gonzàlez-Porta, M.; Carlucci, M.; Chen, X.; Chessman, K.; Faure, A.J.; Funnell, A.P.; Goncalves, A.; et al. Multi-species, multi-transcription factor binding highlights conserved control of tissue-specific biological pathways. Elife 2014, 3, e02626. [Google Scholar] [CrossRef] [PubMed]
- Splinter, E.; Heath, H.; Kooren, J.; Palstra, R.J.; Klous, P.; Grosveld, F.; Galjart, N.; de Laat, W. CTCF mediates long-range chromatin looping and local histone modification in the beta-globin locus. Genes Dev. 2006, 20, 2349–2354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golov, A.K.; Kondratyev, N.V.; Kostyuk, G.P.; Golimbet, A.V.E. Novel Approaches for Identifying the Molecular Background of Schizophrenia. Cells 2020, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Cherkaoui, I.; Misra, S.; Rutter, G.A. Functional Genomics in Pancreatic β Cells: Recent Advances in Gene Deletion and Genome Editing Technologies for Diabetes Research. Front. Endocrinol. (Lausanne.) 2020, 11, 576632. [Google Scholar] [CrossRef]
- Lillian, J.; Tessema, L.; Gessner, R.; Wilson, C.; Kandl, K. Utilizing Knockdowns, Overexpression and Tagging to Study the SUMO Proteases Ulp1 and Ulp2 in Tetrahymena thermophila. FASEB J. 2020. [Google Scholar] [CrossRef]
- Kwok, A.; Raulf, N.; Habib, N. Developing small activating RNA as a therapeutic: Current challenges and promises. Ther. Deliv. 2019, 10, 151–164. [Google Scholar] [CrossRef]
- Lu, Z.R.; Laney, V.E.A.; Hall, R.; Ayat, N. Environment-Responsive Lipid/siRNA Nanoparticles for Cancer Therapy. Adv. Healthc. Mater. 2021, 10, e2001294. [Google Scholar] [CrossRef]
- Liu, H.; Pizzano, S.; Li, R.; Zhao, W.; Veling, M.W.; Hu, Y.; Yang, L.; Ye, B. isoTarget: A Genetic Method for Analyzing the Functional Diversity of Splicing Isoforms in Vivo. Cell. Rep. 2020, 33, 108361. [Google Scholar] [CrossRef]
- Wang, K.; Wang, J.; Domeniconi, C.; Zhang, X.; Yu, G. Differentiating isoform functions with collaborative matrix factorization. Bioinformatics 2020, 36, 1864–1871. [Google Scholar] [CrossRef]
- Ferrer-Bonsoms, J.A.; Cassol, I.; Fernández-Acín, P.; Castilla, C.; Carazo, F.; Rubio, A. ISOGO: Functional annotation of protein-coding splice variants. Sci. Rep. 2020, 10, 1069. [Google Scholar] [CrossRef]
- Derks, M.F.L.; Gjuvsland, A.B.; Bosse, M.; Lopes, M.S.; van Son, M.; Harlizius, B.; Tan, B.F.; Hamland, H.; Grindflek, E.; Groenen, M.A.M.; et al. Loss of function mutations in essential genes cause embryonic lethality in pigs. PLoS Genet. 2019, 15, e1008055. [Google Scholar] [CrossRef] [PubMed]
- Le, D.D.; Shimko, T.C.; Aditham, A.K.; Keys, A.M.; Longwell, S.A.; Orenstein, Y.; Fordyce, P.M. Comprehensive, high-resolution binding energy landscapes reveal context dependencies of transcription factor binding. Proc. Natl. Acad. Sci. USA 2018, 115, E3702–E3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.F.; Cushman, S.A.; He, Y.X.; Li, Y. Genome sequencing and population genomics modeling provide insights into the local adaptation of weeping forsythia. Hortic. Res. 2020, 7, 130. [Google Scholar] [CrossRef] [PubMed]
- Arbiza, L.; Gronau, I.; Aksoy, B.A.; Hubisz, M.J.; Gulko, B.; Keinan, A.; Siepel, A. Genome-wide inference of natural selection on human transcription factor binding sites. Nat. Genet. 2013, 45, 723–729. [Google Scholar] [CrossRef] [Green Version]
- Moyerbrailean, G.A.; Kalita, C.A.; Harvey, C.T.; Wen, X.; Luca, F.; Pique-Regi, R. Which Genetics Variants in DNase-Seq Footprints Are More Likely to Alter Binding? PLoS Genet. 2016, 12, e1005875. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro-dos-Santos, A.M.; da Silva, V.L.; de Souza, J.E.; de Souza, S.J. Populational landscape of INDELs affecting transcription factor-binding sites in humans. BMC Genom. 2015, 16, 536. [Google Scholar] [CrossRef] [Green Version]
- Rappoport, N.; Simon, A.J.; Amariglio, N.; Rechavi, G. The Duffy antigen receptor for chemokines, ACKR1,- ‘Jeanne DARC’ of benign neutropenia. Br. J. Haematol. 2019, 184, 497–507. [Google Scholar] [CrossRef] [Green Version]
- Tso, H.W.; Ip, W.K.; Chong, W.P.; Tam, C.M.; Chiang, A.K.; Lau, Y.L. Association of interferon gamma and interleukin 10 genes with tuberculosis in Hong Kong Chinese. Genes Immun. 2005, 6, 358–363. [Google Scholar] [CrossRef] [Green Version]
- Jablonski, N.G.; Chaplin, G. Colloquium paper: Human skin pigmentation as an adaptation to UV radiation. Proc. Natl. Acad. Sci. USA 2010, 107, 8962–8968. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Su, B. Molecular adaptation of modern human populations. Int. J. Evol. Biol. 2010, 2011, 484769. [Google Scholar] [CrossRef] [Green Version]
- Bond, G.L.; Hu, W.; Bond, E.E.; Robins, H.; Lutzker, S.G.; Arva, N.C.; Bargonetti, J.; Bartel, F.; Taubert, H.; Wuerl, P.; et al. A single nucleotide polymorphism in the MDM2 promoter attenuates the p53 tumor suppressor pathway and accelerates tumor formation in humans. Cell 2004, 119, 591–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böger, C.A.; Gorski, M.; McMahon, G.M.; Xu, H.; Chang, Y.C.; van der Most, P.J.; Navis, G.; Nolte, I.M.; de Borst, M.H.; Zhang, W.; et al. NFAT5 and SLC4A10 Loci Associate with Plasma Osmolality. J. Am. Soc. Nephrol. 2017, 28, 2311–2321. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Momozawa, Y.; Sakaue, S.; Kanai, M.; Ishigaki, K.; Akiyama, M.; Kishikawa, T.; Arai, Y.; Sasaki, T.; Kosaki, K.; et al. Deep whole-genome sequencing reveals recent selection signatures linked to evolution and disease risk of Japanese. Nat. Commun. 2018, 9, 1631. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Tseng, C.C.; Yen, J.H.; Chang, J.G.; Chou, W.C.; Chu, H.W.; Chang, S.J.; Liao, W.T. ABCG2 contributes to the development of gout and hyperuricemia in a genome-wide association study. Sci. Rep. 2018, 8, 3137. [Google Scholar] [CrossRef] [PubMed]
- Roughley, M.; Sultan, A.A.; Clarson, L.; Muller, S.; Whittle, R.; Belcher, J.; Mallen, C.D.; Roddy, E. Risk of chronic kidney disease in patients with gout and the impact of urate lowering therapy: A population-based cohort study. Arthritis. Res. Ther. 2018, 20, 243. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.S.; Lin, C.L.; Tsai, C.H.; Chang, K.H. Association of gout with CAD and effect of antigout therapy on CVD risk among gout patients. J. Investig. Med. 2020, 68, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Thanassoulis, G.; Brophy, J.M.; Richard, H.; Pilote, L. Gout, allopurinol use, and heart failure outcomes. Arch. Intern. Med. 2010, 170, 1358–1364. [Google Scholar] [CrossRef] [Green Version]
- Borghi, C.; Virdis, A. Serum Urate, Uricase, and Blood Pressure Control in Gout. Hypertension 2019, 74, 23–25. [Google Scholar] [CrossRef]
- Kok, V.C.; Horng, J.T.; Wang, M.N.; Chen, Z.Y.; Kuo, J.T.; Hung, G.D. Gout as a risk factor for osteoporosis: Epidemiologic evidence from a population-based longitudinal study involving 108,060 individuals. Osteoporos. Int. 2018, 29, 973–985. [Google Scholar] [CrossRef]
- Shih, H.J.; Kao, M.C.; Tsai, P.S.; Fan, Y.C.; Huang, C.J. Long-term allopurinol use decreases the risk of prostate cancer in patients with gout: A population-based study. Prostate. Cancer. Prostatic. Dis. 2017, 20, 328–333. [Google Scholar] [CrossRef]
- Sanjak, J.S.; Sidorenko, J.; Robinson, M.R.; Thornton, K.R.; Visscher, P.M. Evidence of directional and stabilizing selection in contemporary humans. Proc. Natl. Acad. Sci. USA 2018, 115, 151–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devuyst, O.; Pattaro, C. The UMOD Locus: Insights into the Pathogenesis and Prognosis of Kidney Disease. J. Am. Soc. Nephrol. 2018, 29, 713–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, A.; Wu, Y.; Zhu, Z.; Zhang, F.; Kemper, K.E.; Zheng, Z.; Yengo, L.; Lloyd-Jones, L.R.; Sidorenko, J.; McRae, A.F.; et al. Genome-wide association analyses identify 143 risk variants and putative regulatory mechanisms for type 2 diabetes. Nat. Commun. 2018, 9, 2941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobon, B.; Montanucci, L.; Peretó, J.; Bertranpetit, J.; Laayouni, H. Gene connectivity and enzyme evolution in the human metabolic network. Biol. Direct. 2019, 14, 17. [Google Scholar] [CrossRef] [Green Version]
- Byars, S.G.; Huang, Q.Q.; Gray, L.A.; Bakshi, A.; Ripatti, S.; Abraham, G.; Stearns, S.C.; Inouye, M. Genetic loci associated with coronary artery disease harbor evidence of selection and antagonistic pleiotropy. PLoS Genet. 2017, 13, e1006328. [Google Scholar] [CrossRef] [Green Version]
- Hoh, B.P.; Abdul Rahman, T.; Yusoff, K. Natural selection and local adaptation of blood pressure regulation and their perspectives on precision medicine in hypertension. Hereditas 2019, 156, 1. [Google Scholar] [CrossRef]
- Mullin, B.H.; Walsh, J.P.; Zheng, H.F.; Brown, S.J.; Surdulescu, G.L.; Curtis, C.; Breen, G.; Dudbridge, F.; Richards, J.B.; Spector, T.D.; et al. Genome-wide association study using family-based cohorts identifies the WLS and CCDC170/ESR1 loci as associated with bone mineral density. BMC Genom. 2016, 17, 136. [Google Scholar] [CrossRef] [Green Version]
- Bányai, L.; Trexler, M.; Kerekes, K.; Csuka, O.; Patthy, L. Use of signals of positive and negative selection to distinguish cancer genes and passenger genes. Elife 2021. [Google Scholar] [CrossRef]
- Wu, M.; Tian, Y.; Wang, Q.; Guo, C. Gout: A disease involved with complicated immunoinflammatory responses: A narrative review. Clin. Rheumatol. 2020, 39, 2849–2859. [Google Scholar] [CrossRef]
- McKinney, C.; Stamp, L.K.; Dalbeth, N.; Topless, R.K.; Day, R.O.; Kannangara, D.R.; Williams, K.M.; Janssen, M.; Jansen, T.L.; Joosten, L.A.; et al. Multiplicative interaction of functional inflammasome genetic variants in determining the risk of gout. Arthritis. Res. Ther. 2015, 17, 288. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Deng, Y.; Zhou, X. Multiple Membrane Transporters and Some Immune Regulatory Genes are Major Genetic Factors to Gout. Open. Rheumatol. J. 2018, 12, 94–113. [Google Scholar] [CrossRef] [PubMed]
- Munthe-Kaas, M.C.; Torjussen, T.M.; Gervin, K.; Lødrup Carlsen, K.C.; Carlsen, K.H.; Granum, B.; Hjorthaug, H.S.; Undlien, D.; Lyle, R. CD14 polymorphisms and serum CD14 levels through childhood: A role for gene methylation? J. Allergy. Clin. Immunol. 2010, 125, 1361–1368. [Google Scholar] [CrossRef] [PubMed]
- Frisdal, E.; Klerkx, A.H.; Le Goff, W.; Tanck, M.W.; Lagarde, J.P.; Jukema, J.W.; Kastelein, J.J.; Chapman, M.J.; Guerin, M. Functional interaction between -629C/A, -971G/A and -1337C/T polymorphisms in the CETP gene is a major determinant of promoter activity and plasma CETP concentration in the REGRESS Study. Hum. Mol. Genet. 2005, 14, 2607–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claussnitzer, M.; Dankel, S.N.; Kim, K.H.; Quon, G.; Meuleman, W.; Haugen, C.; Glunk, V.; Sousa, I.S.; Beaudry, J.L.; Puviindran, V.; et al. FTO Obesity Variant Circuitry and Adipocyte Browning in Humans. N. Engl. J. Med. 2015, 373, 895–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, C.G.; Finer, S.; Lindgren, C.M.; Wilson, G.A.; Rakyan, V.K.; Teschendorff, A.E.; Akan, P.; Stupka, E.; Down, T.A.; Prokopenko, I.; et al. Integrated genetic and epigenetic analysis identifies haplotype-specific methylation in the FTO type 2 diabetes and obesity susceptibility locus. PLoS ONE 2010, 5, e14040. [Google Scholar] [CrossRef] [PubMed]
- Köttgen, A.; Glazer, N.L.; Dehghan, A.; Hwang, S.J.; Katz, R.; Li, M.; Yang, Q.; Gudnason, V.; Launer, L.J.; Harris, T.B.; et al. Multiple loci associated with indices of renal function and chronic kidney disease. Nat. Genet. 2009, 41, 712–717. [Google Scholar] [CrossRef] [Green Version]
- Prokop, J.W.; Yeo, N.C.; Ottmann, C.; Chhetri, S.B.; Florus, K.L.; Ross, E.J.; Sosonkina, N.; Link, B.A.; Freedman, B.I.; Coppola, C.J.; et al. Characterization of Coding/Noncoding Variants for SHROOM3 in Patients with CKD. J. Am. Soc. Nephrol. 2018, 29, 1525–1535. [Google Scholar] [CrossRef] [Green Version]
- Köttgen, A.; Pattaro, C.; Böger, C.A.; Fuchsberger, C.; Olden, M.; Glazer, N.L.; Parsa, A.; Gao, X.; Yang, Q.; Smith, A.V.; et al. New loci associated with kidney function and chronic kidney disease. Nat. Genet. 2010, 42, 376–384. [Google Scholar] [CrossRef] [Green Version]
- Ahluwalia, T.S.; Troelsen, J.T.; Balslev-Harder, M.; Bork-Jensen, J.; Thuesen, B.H.; Cerqueira, C.; Linneberg, A.; Grarup, N.; Pedersen, O.; Hansen, T.; et al. Carriers of a VEGFA enhancer polymorphism selectively binding CHOP/DDIT3 are predisposed to increased circulating levels of thyroid-stimulating hormone. J. Med. Genet. 2017, 54, 166–175. [Google Scholar] [CrossRef]
- Sun, C.Y.; Wu, M.S.; Lee, C.C.; Chen, S.H.; Lo, K.C.; Chen, Y.H. A novel SNP in the 5’ regulatory region of organic anion transporter 1 is associated with chronic kidney disease. Sci. Rep. 2018, 8, 8085. [Google Scholar] [CrossRef]
- Gaulton, K.J.; Ferreira, T.; Lee, Y.; Raimondo, A.; Mägi, R.; Reschen, M.E.; Mahajan, A.; Locke, A.; Rayner, N.W.; Robertson, N.; et al. Genetic fine mapping and genomic annotation defines causal mechanisms at type 2 diabetes susceptibility loci. Nat. Genet. 2015, 47, 1415–1425. [Google Scholar] [CrossRef] [PubMed]
- Lyssenko, V.; Nagorny, C.L.; Erdos, M.R.; Wierup, N.; Jonsson, A.; Spégel, P.; Bugliani, M.; Saxena, R.; Fex, M.; Pulizzi, N.; et al. Common variant in MTNR1B associated with increased risk of type 2 diabetes and impaired early insulin secretion. Nat. Genet. 2009, 41, 82–88. [Google Scholar] [CrossRef]
- Fogarty, M.P.; Cannon, M.E.; Vadlamudi, S.; Gaulton, K.J.; Mohlke, K.L. Identification of a regulatory variant that binds FOXA1 and FOXA2 at the CDC123/CAMK1D type 2 diabetes GWAS locus. PLoS Genet. 2014, 10, e1004633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fromont, C.; Atzori, A.; Kaur, D.; Hashmi, L.; Greco, G.; Cabanillas, A.; Nguyen, H.V.; Jones, D.H.; Garzón, M.; Varela, A.; et al. Discovery of Highly Selective Inhibitors of Calmodulin-Dependent Kinases That Restore Insulin Sensitivity in the Diet-Induced Obesity in Vivo Mouse Model. J. Med. Chem. 2020, 63, 6784–6801. [Google Scholar] [CrossRef]
- Hiramoto, M.; Udagawa, H.; Ishibashi, N.; Takahashi, E.; Kaburagi, Y.; Miyazawa, K.; Funahashi, N.; Nammo, T.; Yasuda, K. A type 2 diabetes-associated SNP in KCNQ1 (rs163184) modulates the binding activity of the locus for Sp3 and Lsd1/Kdm1a, potentially affecting CDKN1C expression. Int. J. Mol. Med. 2018, 41, 717–728. [Google Scholar] [CrossRef]
- Elliott, H.R.; Shihab, H.A.; Lockett, G.A.; Holloway, J.W.; McRae, A.F.; Smith, G.D.; Ring, S.M.; Gaunt, T.R.; Relton, C.L. Role of DNA Methylation in Type 2 Diabetes Etiology: Using Genotype as a Causal Anchor. Diabetes 2017, 66, 1713–1722. [Google Scholar] [CrossRef] [Green Version]
- Fogarty, M.P.; Panhuis, T.M.; Vadlamudi, S.; Buchkovich, M.L.; Mohlke, K.L. Allele-specific transcriptional activity at type 2 diabetes-associated single nucleotide polymorphisms in regions of pancreatic islet open chromatin at the JAZF1 locus. Diabetes 2013, 62, 1756–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wesolowska-Andersen, A.; Zhuo Yu, G.; Nylander, V.; Abaitua, F.; Thurner, M.; Torres, J.M.; Mahajan, A.; Gloyn, A.L.; McCarthy, M.I. Deep learning models predict regulatory variants in pancreatic islets and refine type 2 diabetes association signals. Elife 2020, 9, e51503. [Google Scholar] [CrossRef]
- Adamska-Patruno, E.; Godzien, J.; Ciborowski, M.; Samczuk, P.; Bauer, W.; Siewko, K.; Gorska, M.; Barbas, C.; Kretowski, A. The Type 2 Diabetes Susceptibility PROX1 Gene Variants Are Associated with Postprandial Plasma Metabolites Profile in Non-Diabetic Men. Nutrients 2019, 11, 882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.; Huang, X.; Zhang, M.; Huang, Q. Computational and functional analyses of T2D GWAS SNPs for transcription factor binding. Biochem. Biophys. Res. Commun. 2020, 523, 658–665. [Google Scholar] [CrossRef]
- Claussnitzer, M.; Dankel, S.N.; Klocke, B.; Grallert, H.; Glunk, V.; Berulava, T.; Lee, H.; Oskolkov, N.; Fadista, J.; Ehlers, K.; et al. Leveraging cross-species transcription factor binding site patterns: From diabetes risk loci to disease mechanisms. Cell 2014, 156, 343–358. [Google Scholar] [CrossRef] [Green Version]
- Piao, X.; Yahagi, N.; Takeuchi, Y.; Aita, Y.; Murayama, Y.; Sawada, Y.; Shikama, A.; Masuda, Y.; Nishi-Tatsumi, M.; Kubota, M.; et al. A candidate functional SNP rs7074440 in TCF7L2 alters gene expression through C-FOS in hepatocytes. FEBS Lett. 2018, 592, 422–433. [Google Scholar] [CrossRef] [Green Version]
- López Rodríguez, M.; Kaminska, D.; Lappalainen, K.; Pihlajamäki, J.; Kaikkonen, M.U.; Laakso, M. Identification and characterization of a FOXA2-regulated transcriptional enhancer at a type 2 diabetes intronic locus that controls GCKR expression in liver cells. Genome. Med. 2017, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Oskolkov, N.; Shcherbina, L.; Ratti, J.; Kock, K.H.; Su, J.; Martin, B.; Oskolkova, M.Z.; Göransson, O.; Bacon, J.; et al. HMGB1 binds to the rs7903146 locus in TCF7L2 in human pancreatic islets. Mol. Cell. Endocrinol. 2016, 430, 138–145. [Google Scholar] [CrossRef]
- Susa, S.; Daimon, M.; Sakabe, J.; Sato, H.; Oizumi, T.; Karasawa, S.; Wada, K.; Jimbu, Y.; Kameda, W.; Emi, M.; et al. A functional polymorphism of the TNF-alpha gene that is associated with type 2 DM. Biochem. Biophys. Res. Commun. 2008, 369, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Li, J.; Sheng, J.; Dai, Y.; Wang, Y.; Liu, J.; Xu, Y. Haplotypes of the Mutated SIRT2 Promoter Contributing to Transcription Factor Binding and Type 2 Diabetes Susceptibility. Genes 2020, 11, 569. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Tian, M.; Hu, S.; Wang, Y.; Wang, D.W. Identifying functional non-coding variants in APOA5/A4/C3/A1 gene cluster associated with coronary heart disease. J. Mol. Cell. Cardiol. 2020, 144, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Roman, T.S.; Marvelle, A.F.; Fogarty, M.P.; Vadlamudi, S.; Gonzalez, A.J.; Buchkovich, M.L.; Huyghe, J.R.; Fuchsberger, C.; Jackson, A.U.; Wu, Y.; et al. Multiple Hepatic Regulatory Variants at the GALNT2 GWAS Locus Associated with High-Density Lipoprotein Cholesterol. Am. J. Hum. Genet. 2015, 97, 801–815. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, Y.; Zhang, M.; Wang, Y. GALNT2 regulates ANGPTL3 cleavage in cells and in vivo of mice. Sci. Rep. 2020, 10, 16168. [Google Scholar] [CrossRef]
- Pfeiffer, L.; Wahl, S.; Pilling, L.C.; Reischl, E.; Sandling, J.K.; Kunze, S.; Holdt, L.M.; Kretschmer, A.; Schramm, K.; Adamski, J.; et al. DNA methylation of lipid-related genes affects blood lipid levels. Circ. Cardiovasc. Genet. 2015, 8, 334–342. [Google Scholar] [CrossRef]
- Lin, S.; Dai, R.; Lin, R. A meta-analytic evaluation of cholesteryl ester transfer protein (CETP) C-629A polymorphism in association with coronary heart disease risk and lipid changes. Oncotarget 2017, 8, 2153–2163. [Google Scholar] [CrossRef] [Green Version]
- Dachet, C.; Poirier, O.; Cambien, F.; Chapman, J.; Rouis, M. New functional promoter polymorphism, CETP/-629, in cholesteryl ester transfer protein (CETP) gene related to CETP mass and high density lipoprotein cholesterol levels: Role of Sp1/Sp3 in transcriptional regulation. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Musunuru, K.; Strong, A.; Frank-Kamenetsky, M.; Lee, N.E.; Ahfeldt, T.; Sachs, K.V.; Li, X.; Li, H.; Kuperwasser, N.; Ruda, V.M.; et al. From noncoding variant to phenotype via SORT1 at the 1p13 cholesterol locus. Nature 2010, 466, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Gurdasani, D.; Carstensen, T.; Fatumo, S.; Chen, G.; Franklin, C.S.; Prado-Martinez, J.; Bouman, H.; Abascal, F.; Haber, M.; Tachmazidou, I.; et al. Uganda Genome Resource Enables Insights into Population History and Genomic Discovery in Africa. Cell 2019, 179, 984–1002.e1036. [Google Scholar] [CrossRef] [PubMed]
- Fairoozy, R.H.; White, J.; Palmen, J.; Kalea, A.Z.; Humphries, S.E. Identification of the Functional Variant(s) that Explain the Low-Density Lipoprotein Receptor (LDLR) GWAS SNP rs6511720 Association with Lower LDL-C and Risk of CHD. PLoS ONE 2016, 11, e0167676. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Zhao, Y.; Du, H.; Nie, X.; Li, H.; Fan, J.; He, M.; Dai, B.; Zhang, X.; Yuan, S.; et al. A Key GWAS-Identified Genetic Variant Contributes to Hyperlipidemia by Upregulating miR-320a. iScience 2020, 23, 101788. [Google Scholar] [CrossRef] [PubMed]
- Reschen, M.E.; Gaulton, K.J.; Lin, D.; Soilleux, E.J.; Morris, A.J.; Smyth, S.S.; O’Callaghan, C.A. Lipid-induced epigenomic changes in human macrophages identify a coronary artery disease-associated variant that regulates PPAP2B Expression through Altered C/EBP-beta binding. PLoS Genet. 2015, 11, e1005061. [Google Scholar] [CrossRef]
- Aminoff, A.; Ledmyr, H.; Thulin, P.; Lundell, K.; Nunez, L.; Strandhagen, E.; Murphy, C.; Lidberg, U.; Westerbacka, J.; Franco-Cereceda, A.; et al. Allele-specific regulation of MTTP expression influences the risk of ischemic heart disease. J. Lipid. Res. 2010, 51, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Wen, G.; Wessel, J.; Zhou, W.; Ehret, G.B.; Rao, F.; Stridsberg, M.; Mahata, S.K.; Gent, P.M.; Das, M.; Cooper, R.S.; et al. An ancestral variant of Secretogranin II confers regulation by PHOX2 transcription factors and association with hypertension. Hum. Mol. Genet. 2007, 16, 1752–1764. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, Y.; Wen, G.; Mahata, M.; Rao, F.; Fung, M.M.; Vaingankar, S.; Biswas, N.; Gayen, J.R.; Friese, R.S.; et al. Catecholamine storage vesicles: Role of core protein genetic polymorphisms in hypertension. Curr. Hypertens. Rep. 2011, 13, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, L.; Maghajothi, S.; Singh, M.; Kesh, K.; Kalyani, A.; Sharma, S.; Khullar, M.; Victor, S.M.; Swarnakar, S.; Asthana, S.; et al. A Common Tag Nucleotide Variant in MMP7 Promoter Increases Risk for Hypertension via Enhanced Interactions With CREB (Cyclic AMP Response Element-Binding Protein) Transcription Factor. Hypertension 2019, 74, 1448–1459. [Google Scholar] [CrossRef]
- Mopidevi, B.; Kaw, M.K.; Sivankutty, I.; Jain, S.; Perla, S.K.; Kumar, A. A polymorphism in intron I of the human angiotensinogen gene (hAGT) affects binding by HNF3 and hAGT expression and increases blood pressure in mice. J. Biol. Chem. 2019, 294, 11829–11839. [Google Scholar] [CrossRef]
- Park, S.; Lu, K.T.; Liu, X.; Chatterjee, T.K.; Rudich, S.M.; Weintraub, N.L.; Kwitek, A.E.; Sigmund, C.D. Allele-specific expression of angiotensinogen in human subcutaneous adipose tissue. Hypertension 2013, 62, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Purkait, P.; Halder, K.; Thakur, S.; Ghosh Roy, A.; Raychaudhuri, P.; Bhattacharya, S.; Sarkar, B.N.; Naidu, J.M. Association of angiotensinogen gene SNPs and haplotypes with risk of hypertension in eastern Indian population. Clin. Hypertens. 2017, 23, 12. [Google Scholar] [CrossRef] [Green Version]
- Fjorder, A.S.; Rasmussen, M.B.; Mehrjouy, M.M.; Nazaryan-Petersen, L.; Hansen, C.; Bak, M.; Grarup, N.; Nørremølle, A.; Larsen, L.A.; Vestergaard, H.; et al. Haploinsufficiency of ARHGAP42 is associated with hypertension. Eur. J. Hum. Genet. 2019, 27, 1296–1303. [Google Scholar] [CrossRef]
- Bai, X.; Mangum, K.D.; Dee, R.A.; Stouffer, G.A.; Lee, C.R.; Oni-Orisan, A.; Patterson, C.; Schisler, J.C.; Viera, A.J.; Taylor, J.M.; et al. Blood pressure-associated polymorphism controls ARHGAP42 expression via serum response factor DNA binding. J. Clin. Invest. 2017, 127, 670–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Zhu, Y.; Cheng, M.; Dinesh, D.; Thorne, T.; Poh, K.K.; Liu, D.; Botros, C.; Tang, Y.L.; Reisdorph, N.; et al. Regulation of vascular contractility and blood pressure by the E2F2 transcription factor. Circulation 2009, 120, 1213–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzaghi, C.; Paroni, G.; De Bonis, C.; Soccio, T.; Marucci, A.; Bacci, S.; Trischitta, V. The -318 C>G single-nucleotide polymorphism in GNAI2 gene promoter region impairs transcriptional activity through specific binding of Sp1 transcription factor and is associated with high blood pressure in Caucasians from Italy. J. Am. Soc. Nephrol. 2006, 17, S115–S119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wainford, R.D.; Carmichael, C.Y.; Pascale, C.L.; Kuwabara, J.T. Gαi2-protein-mediated signal transduction: Central nervous system molecular mechanism countering the development of sodium-dependent hypertension. Hypertension 2015, 65, 178–186. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.A.; Cleveland, J.D. Gout and the risk of age-related macular degeneration in the elderly. PLoS ONE 2018, 13, e0199562. [Google Scholar] [CrossRef]
- Ketharnathan, S.; Leask, M.; Boocock, J.; Phipps-Green, A.J.; Antony, J.; O’Sullivan, J.M.; Merriman, T.R.; Horsfield, J.A. A non-coding genetic variant maximally associated with serum urate levels is functionally linked to HNF4A-dependent PDZK1 expression. Hum. Mol. Genet. 2018, 27, 3964–3973. [Google Scholar] [CrossRef]
- Chen, M.; Lu, X.; Lu, C.; Shen, N.; Jiang, Y.; Wu, H. Soluble uric acid increases PDZK1 and ABCG2 expression in human intestinal cell lines via the TLR4-NLRP3 inflammasome and PI3K/Akt signaling pathway. Arthritis. Res. Ther. 2018, 20, 20. [Google Scholar] [CrossRef] [Green Version]
- Ziablitsev, D.S.; Larin, O.S. Influence of single nucleotide polymorphisms of vitamin D receptor-gene on the level of osteoassociated hormones linkage with postmenopausal osteoporosis. Fiziol. Zh. 2015, 61, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, H.; Miyamoto, K.I.; Yoshida, M.; Yamamoto, H.; Taketani, Y.; Morita, K.; Kubota, M.; Yoshida, S.; Ikeda, M.; Watabe, F.; et al. The polymorphism in the caudal-related homeodomain protein Cdx-2 binding element in the human vitamin D receptor gene. J. Bone. Miner. Res. 2001, 16, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Marozik, P.; Rudenka, A.; Kobets, K.; Rudenka, E. Vitamin D Status, Bone Mineral Density, and VDR Gene Polymorphism in a Cohort of Belarusian Postmenopausal Women. Nutrients 2021, 13, 837. [Google Scholar] [CrossRef] [PubMed]
- Mann, V.; Hobson, E.E.; Li, B.; Stewart, T.L.; Grant, S.F.; Robins, S.P.; Aspden, R.M.; Ralston, S.H. A COL1A1 Sp1 binding site polymorphism predisposes to osteoporotic fracture by affecting bone density and quality. J. Clin. Invest. 2001, 107, 899–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, P.; Liu, B.; Zhang, L.; Chen, R.; Yang, B.; Dong, J.; Rong, L. Association of COL1A1 polymorphisms with osteoporosis: A meta-analysis of clinical studies. Int. J. Clin. Exp. Med. 2015, 8, 14764–14781. [Google Scholar] [PubMed]
- Zhu, D.L.; Chen, X.F.; Hu, W.X.; Dong, S.S.; Lu, B.J.; Rong, Y.; Chen, Y.X.; Chen, H.; Thynn, H.N.; Wang, N.N.; et al. Multiple Functional Variants at 13q14 Risk Locus for Osteoporosis Regulate RANKL Expression Through Long-Range Super-Enhancer. J. Bone. Miner. Res. 2018, 33, 1335–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lill, C.M.; Liu, T.; Norman, K.; Meyer, A.; Steinhagen-Thiessen, E.; Demuth, I.; Bertram, L. Genetic Burden Analyses of Phenotypes Relevant to Aging in the Berlin Aging Study II (BASE-II). Gerontology 2016, 62, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Whitington, T.; Gao, P.; Lindberg, J.F.; Yang, Y.; Sun, J.; Väisänen, M.R.; Szulkin, R.; Annala, M.; Yan, J.; et al. A prostate cancer susceptibility allele at 6q22 increases RFX6 expression by modulating HOXB13 chromatin binding. Nat. Genet. 2014, 46, 126–135. [Google Scholar] [CrossRef]
- Houlahan, K.E.; Shiah, Y.J.; Gusev, A.; Yuan, J.; Ahmed, M.; Shetty, A.; Ramanand, S.G.; Yao, C.Q.; Bell, C.; O’Connor, E.; et al. Genome-wide germline correlates of the epigenetic landscape of prostate cancer. Nat. Med. 2019, 25, 1615–1626. [Google Scholar] [CrossRef] [PubMed]
- Ramanand, S.G.; Chen, Y.; Yuan, J.; Daescu, K.; Lambros, M.B.; Houlahan, K.E.; Carreira, S.; Yuan, W.; Baek, G.; Sharp, A.; et al. The landscape of RNA polymerase II-associated chromatin interactions in prostate cancer. J. Clin. Invest. 2020, 130, 3987–4005. [Google Scholar] [CrossRef]
- Emami, N.C.; Cavazos, T.B.; Rashkin, S.R.; Graff, R.E.; Tai, C.G.; Mefford, J.A.; Kachuri, L.; Cario, C.L.; Wan, E.; Wong, S.; et al. A large-scale association study detects novel rare variants, risk genes, functional elements, and polygenic architecture of prostate cancer susceptibility. Cancer Res. 2020. [Google Scholar] [CrossRef]
- Whitington, T.; Gao, P.; Song, W.; Ross-Adams, H.; Lamb, A.D.; Yang, Y.; Svezia, I.; Klevebring, D.; Mills, I.G.; Karlsson, R.; et al. Gene regulatory mechanisms underpinning prostate cancer susceptibility. Nat. Genet. 2016, 48, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Tillmans, L.S.; Thibodeau, S.N.; Wang, L. Single-Nucleotide Polymorphisms Sequencing Identifies Candidate Functional Variants at Prostate Cancer Risk Loci. Genes 2019, 10, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Feng, Z.; Li, J.; Han, Y.; Su, L.; Wang, F.; Yang, Y.; Zhang, Y. Functional Variant rs4442975 Modulating FOXA1 Binding Affinity Can Influence Bone Marrow Suppression during Neoadjuvant Chemotherapy for Luminal A Type Breast Cancer. Biomed. Res. Int. 2019, 2019, 7073498. [Google Scholar] [CrossRef] [PubMed]
- Patti, G.; Micieli, G.; Cimminiello, C.; Bolognese, L. The Role of Clopidogrel in 2020: A Reappraisal. Cardiovasc. Ther. 2020, 2020, 8703627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depta, J.P.; Lenzini, P.A.; Lanfear, D.E.; Wang, T.Y.; Spertus, J.A.; Bach, R.G.; Cresci, S. Clinical outcomes associated with proton pump inhibitor use among clopidogrel-treated patients within CYP2C19 genotype groups following acute myocardial infarction. Pharm. J. 2015, 15, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Frére, C.; Cuisset, T.; Gaborit, B.; Alessi, M.C.; Hulot, J.S. The CYP2C19*17 allele is associated with better platelet response to clopidogrel in patients admitted for non-ST acute coronary syndrome. J. Thromb. Haemost. 2009, 7, 1409–1411. [Google Scholar] [CrossRef] [PubMed]
- Frey, U.H.; Hauner, H.; Jöckel, K.H.; Manthey, I.; Brockmeyer, N.; Siffert, W. A novel promoter polymorphism in the human gene GNAS affects binding of transcription factor upstream stimulatory factor 1, Galphas protein expression and body weight regulation. Pharm. Genom. 2008, 18, 141–151. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Variants | Transcription Factors | Target Genes | References |
|---|---|---|---|
| Alter histone modification | |||
| rs2886870 | NF-κB | C3orf59 | [22] |
| rs4784227 | FOXA1 | TOX3 | [23] |
| rs6983267 | TCF4 | c-MYC | [24] |
| Alter DNA methylation | |||
| rs2240032 | SMAD3 | RAD50 and IL4 | [25] |
| rs612529 | YY1 and PU.1 | SIRL-1 | [26] |
| chr22:24,059,610 | UA4 | GUSBP11 | [20] |
| Alter chromatin conformation | |||
| rs12913832 | HLTF, LEF1, and MITF | OCA2 | [27] |
| rs13228237 | ZNF143 | ZC3HAV1 | [28] |
| rs2802292 | HSF1 | FOXO3 | [29] |
| Category | Variant | Gene | Biological Function | Reference |
|---|---|---|---|---|
| Infection | rs139999735 | APIP | Response to Salmonella | [67] |
| rs281477 8 | ACKR1 | Protection against malaria infection | [68] | |
| IFN-γ + 874 | IFN-γ | Tuberculosis susceptibility | [69] | |
| Radiation | rs201097793 | MC1R | Pigmentation | [67] |
| rs2279744 | MDM2 | Embryo implantation | [71,72] | |
| Taste | rs139938620 | TAS1R3 | Umami taste | [67] |
| Water conservation | rs16846053 | SLC4A10 | Increased plasma osmolality | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tseng, C.-C.; Wong, M.-C.; Liao, W.-T.; Chen, C.-J.; Lee, S.-C.; Yen, J.-H.; Chang, S.-J. Genetic Variants in Transcription Factor Binding Sites in Humans: Triggered by Natural Selection and Triggers of Diseases. Int. J. Mol. Sci. 2021, 22, 4187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084187
Tseng C-C, Wong M-C, Liao W-T, Chen C-J, Lee S-C, Yen J-H, Chang S-J. Genetic Variants in Transcription Factor Binding Sites in Humans: Triggered by Natural Selection and Triggers of Diseases. International Journal of Molecular Sciences. 2021; 22(8):4187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084187
Chicago/Turabian StyleTseng, Chia-Chun, Man-Chun Wong, Wei-Ting Liao, Chung-Jen Chen, Su-Chen Lee, Jeng-Hsien Yen, and Shun-Jen Chang. 2021. "Genetic Variants in Transcription Factor Binding Sites in Humans: Triggered by Natural Selection and Triggers of Diseases" International Journal of Molecular Sciences 22, no. 8: 4187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084187