Platyphyllenone Induces Autophagy and Apoptosis by Modulating the AKT and JNK Mitogen-Activated Protein Kinase Pathways in Oral Cancer Cells

 , and
, and 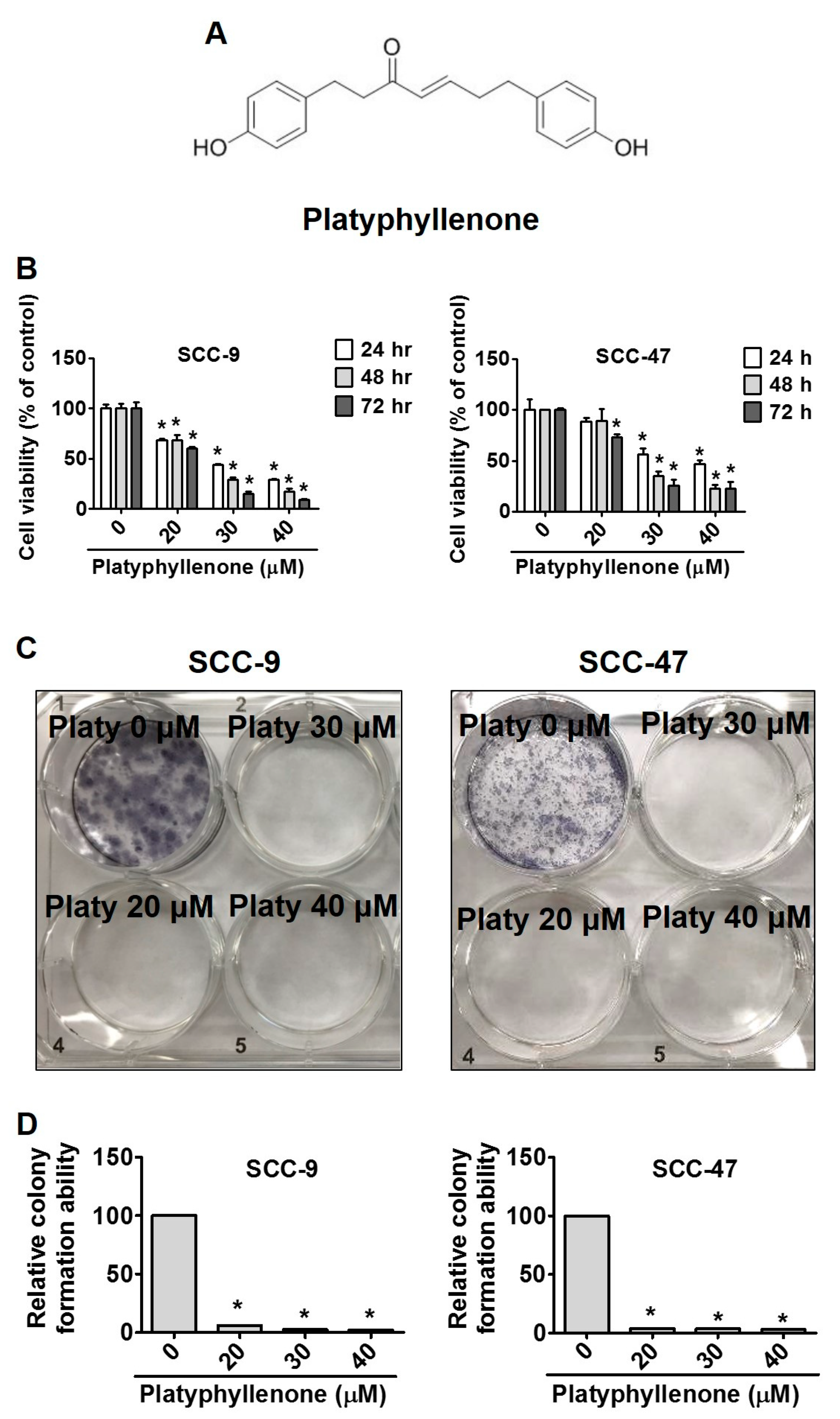{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Platyphyllenone Induces Cytotoxicity and Inhibits Colony Formation in Oral Cancer Cells
2.2. Effect of Platyphyllenone on Cell Cycle Progression in Oral Cancer Cells
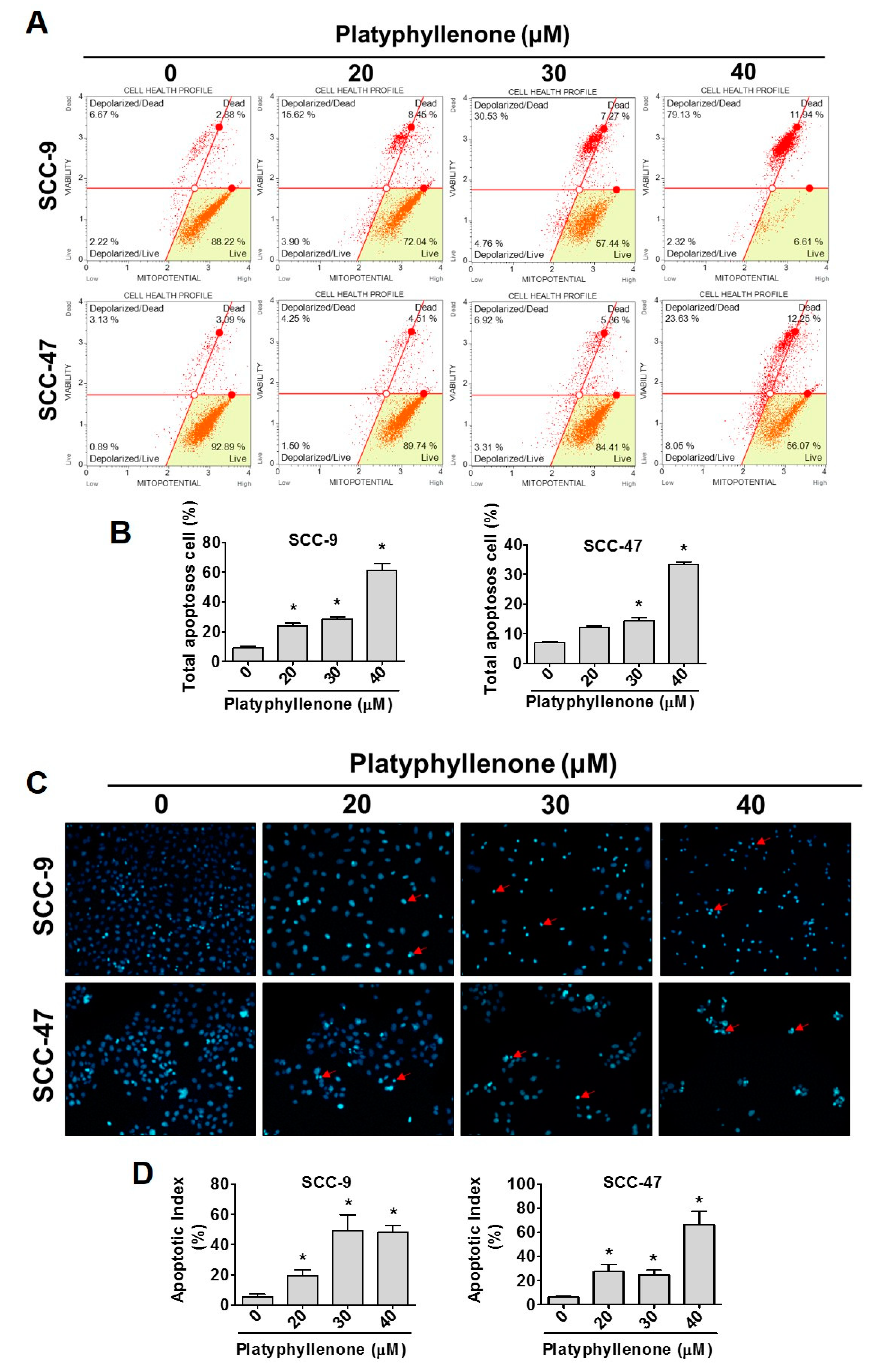
2.3. Platyphyllenone Induces Cell Apoptosis in Oral Cancer Cells
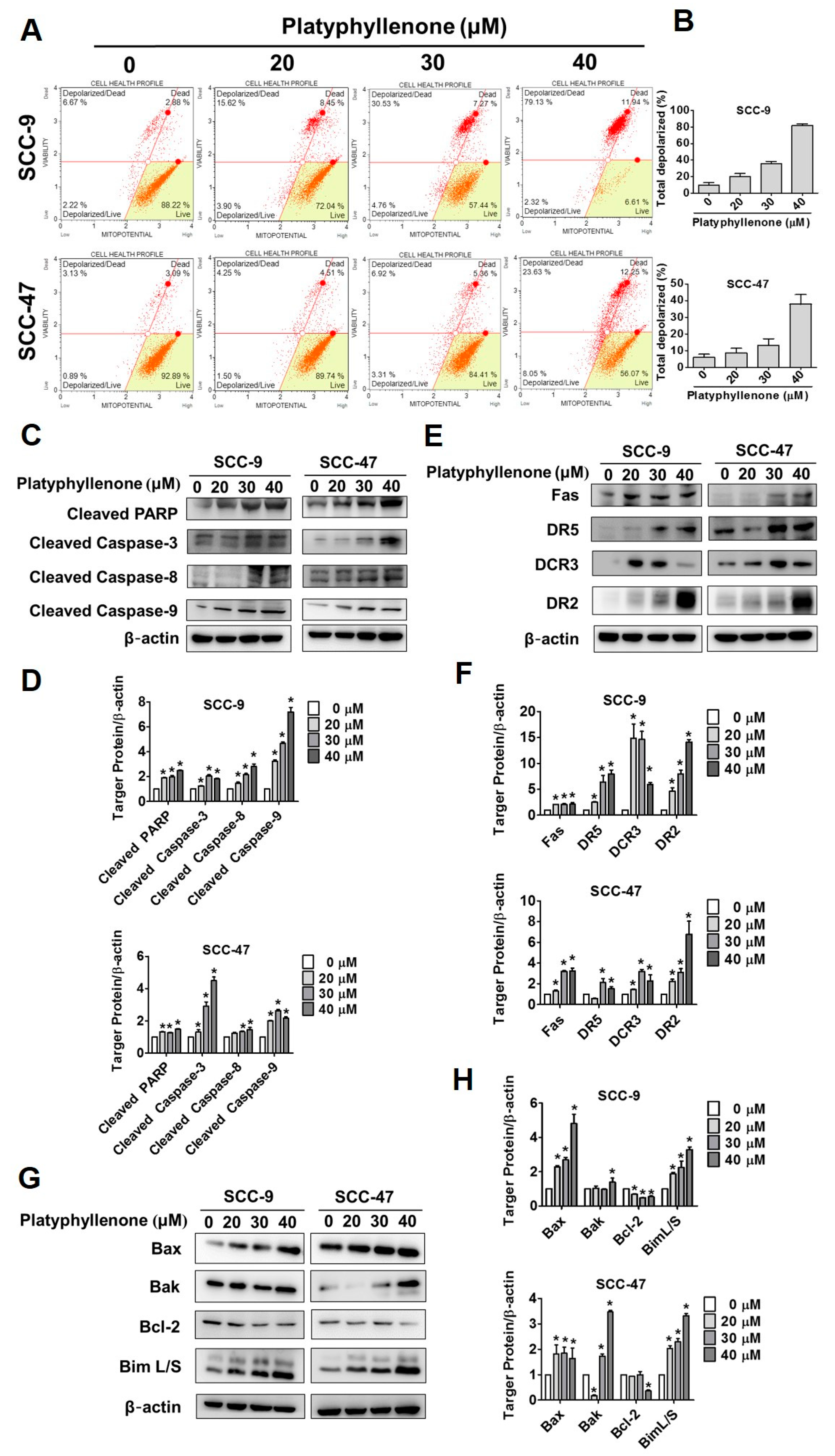
2.4. Platyphyllenone Activates Apoptosis via the Mitochondrial and Death Receptor Pathways and Regulators of Apoptosis-Related Proteins in Oral Cancer Cells
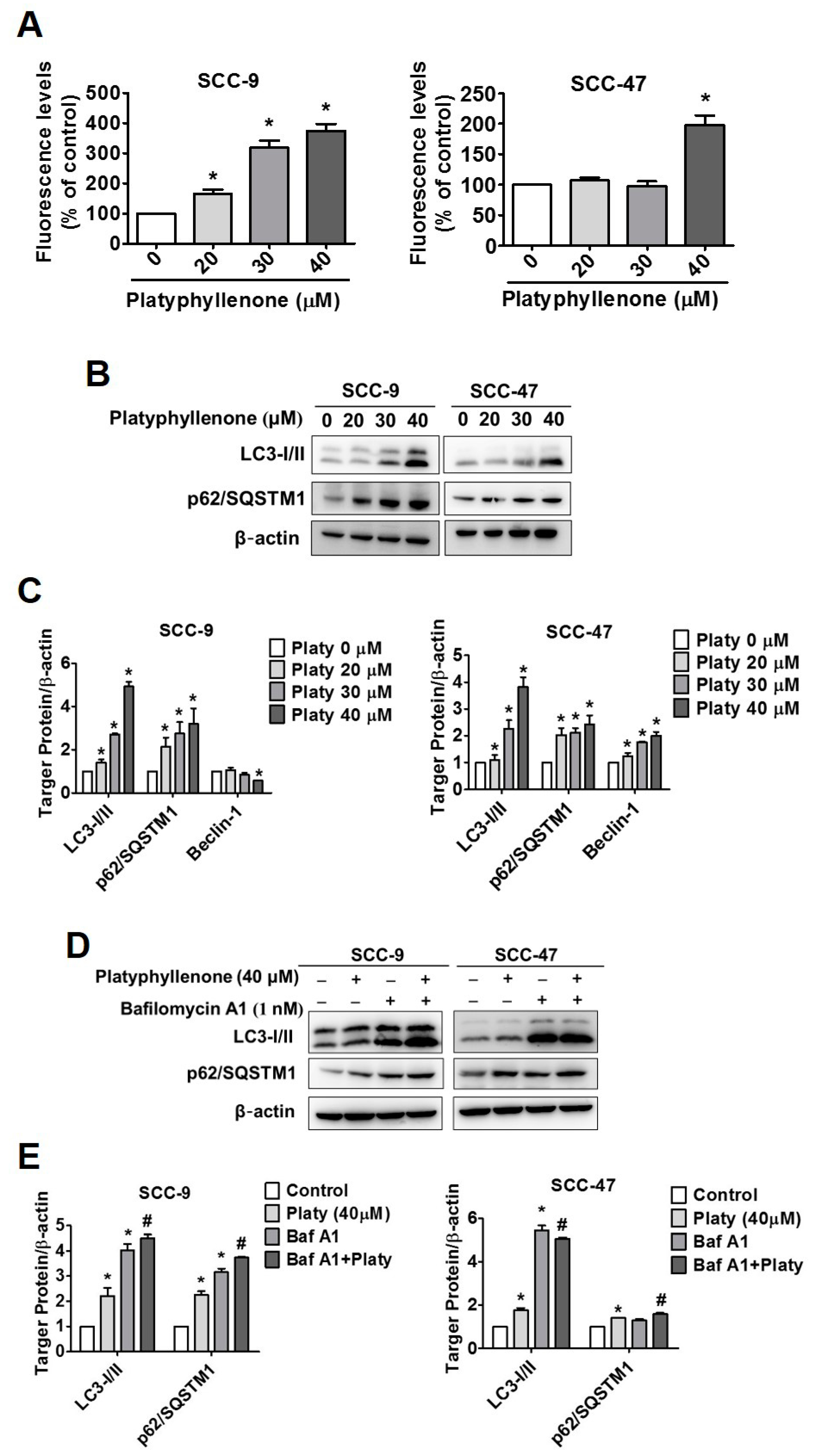
2.5. Platyphyllenone Induces Autophagy in SCC-9 and SCC-47 Oral Cancer Cells
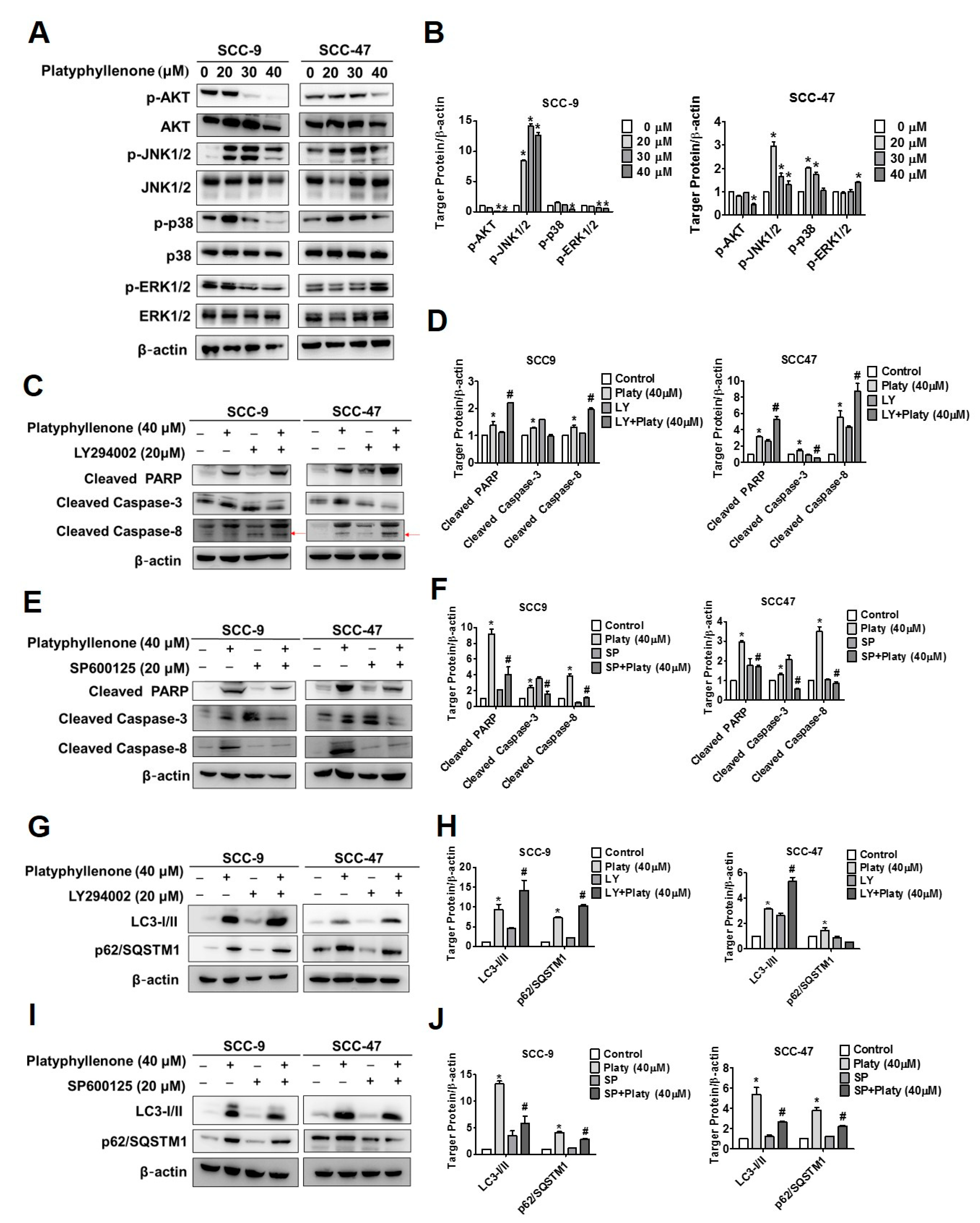
2.6. The Induction of Apoptosis and Autophagy by Platyphyllenone Is Dependent on the Regulation of AKT and JNK Signaling Pathways in SCC-9 and SCC-47 Oral Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Platyphyllenone Treatment and Chemicals
4.3. Cell Cytotoxicity Assay
4.4. Colony Formation Assay
4.5. Cell Cycle Analysis
4.6. Annexin V/PI Double Staining Analysis
4.7. Mitochondrial Membrane Potential Analysis
4.8. DAPI Staining Assay
4.9. Western Blot Analysis
4.10. Autohphagy Fluorescence Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gholizadeh, P.; Eslami, H.; Yousefi, M.; Asgharzadeh, M.; Aghazadeh, M.; Kafil, H.S. Role of oral microbiome on oral cancers, a review. Biomed. Pharmacother. 2016, 84, 552–558. [Google Scholar] [CrossRef]
- Huang, C.C.; Ou, C.Y.; Lee, W.T.; Hsiao, J.R.; Tsai, S.T.; Wang, J.D. Life expectancy and expected years of life lost to oral cancer in Taiwan: A nation-wide analysis of 22,024 cases followed for 10 years. Oral Oncol. 2015, 51, 349–354. [Google Scholar] [CrossRef]
- Jitender, S.; Sarika, G.; Varada, H.R.; Omprakash, Y.; Mohsin, K. Screening for oral cancer. J. Exp. Ther. Oncol. 2016, 11, 303–307. [Google Scholar] [PubMed]
- Yadav, D.; Singh, S.C.; Verma, R.K.; Saxena, K.; Verma, R.; Murthy, P.K.; Gupta, M.M. Antifilarial diarylheptanoids from Alnus nepalensis leaves growing in high altitude areas of Uttarakhand, India. Phytomedicine 2013, 20, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.Z.; Jeong, J.H.; Lee, Y.I.; Lee, S.Y.; Zhao, H.Y.; Jeon, R.; Lee, H.J.; Ryu, J.H. Diarylheptanoids suppress proliferation of pancreatic cancer PANC-1 cells through modulating shh-Gli-FoxM1 pathway. Arch. Pharm. Res. 2017, 40, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.; Sung, C.K.; Im, Y.J.; Chun, C. Activation of JNK and p38 in MCF-7 Cells and the In Vitro Anticancer Activity of Alnus hirsuta Extract. Molecules 2020, 25, 1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novakovic, M.; Stankovic, M.; Vuckovic, I.; Todorovic, N.; Trifunovic, S.; Tesevic, V.; Vajs, V.; Milosavljevic, S. Diarylheptanoids from Alnus glutinosa bark and their chemoprotective effect on human lymphocytes DNA. Planta Med. 2013, 79, 499–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, S.R.; Mohamed, G.A.; Khedr, A.I.; Aljaeid, B.M. Anti-oxidant and Anti-Inflammatory Cyclic Diarylheptanoids from Alnus japonica Stem Bark. Iran J. Pharm. Res. 2017, 16, 83–91. [Google Scholar]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Majno, G.; Joris, I. Apoptosis, oncosis, and necrosis. An overview of cell death. Am. J. Pathol. 1995, 146, 3–15. [Google Scholar]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.J.; Chien, S.Y.; Lin, J.T.; Yang, S.F.; Chen, M.K. Polyphyllin G induces apoptosis and autophagy cell death in human oral cancer cells. Phytomedicine 2016, 23, 1545–1554. [Google Scholar] [CrossRef]
- Bonam, S.R.; Wang, F.; Muller, S. Lysosomes as a therapeutic target. Nat. Rev. Drug Discov. 2019, 18, 923–948. [Google Scholar] [CrossRef] [Green Version]
- Bonam, S.R.; Bayry, J.; Tschan, M.P.; Muller, S. Progress and Challenges in The Use of MAP1LC3 as a Legitimate Marker for Measuring Dynamic Autophagy In Vivo. Cells 2020, 9, 1321. [Google Scholar] [CrossRef] [PubMed]
- Delou, J.M.; Biasoli, D.; Borges, H.L. The Complex Link between Apoptosis and Autophagy: A Promising New Role for RB. An. Acad. Bras. Cienc. 2016, 88, 2257–2275. [Google Scholar] [CrossRef] [Green Version]
- Su, M.; Mei, Y.; Sinha, S. Role of the Crosstalk between Autophagy and Apoptosis in Cancer. J. Oncol. 2013, 2013, 102735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yorimitsu, T.; Klionsky, D.J. Autophagy: Molecular machinery for self-eating. Cell Death Differ. 2005, 12 (Suppl. S2), 1542–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsin, M.C.; Hsieh, Y.H.; Wang, P.H.; Ko, J.L.; Hsin, I.L.; Yang, S.F. Hispolon suppresses metastasis via autophagic degradation of cathepsin S in cervical cancer cells. Cell Death Dis. 2017, 8, e3089. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.Y.; Li, Y.; Jiang, W.Q.; Zhou, L.F. MAPK/JNK signalling: A potential autophagy regulation pathway. Biosci. Rep. 2015, 35, e00199. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Wang, Y.; Jiang, D.; Wang, T.; Zhang, Y.; Ma, H.; Kang, Y. Cyclic Compressive Stress Regulates Apoptosis in Rat Osteoblasts: Involvement of PI3K/Akt and JNK MAPK Signaling Pathways. PLoS ONE 2016, 11, e0165845. [Google Scholar] [CrossRef]
- Tsou, H.H.; Hu, C.H.; Liu, J.H.; Liu, C.J.; Lee, C.H.; Liu, T.Y.; Wang, H.T. Acrolein Is Involved in the Synergistic Potential of Cigarette Smoking- and Betel Quid Chewing-Related Human Oral Cancer. Cancer Epidemiol. Biomark. Prev. 2019, 28, 954–962. [Google Scholar] [PubMed] [Green Version]
- Hung, L.C.; Kung, P.T.; Lung, C.H.; Tsai, M.H.; Liu, S.A.; Chiu, L.T.; Huang, K.H.; Tsai, W.C. Assessment of the Risk of Oral Cancer Incidence in A High-Risk Population and Establishment of A Predictive Model for Oral Cancer Incidence Using A Population-Based Cohort in Taiwan. Int. J. Environ. Res. Public Health 2020, 17, 665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Donnelly, C.R.; Gong, W.; Heath, B.R.; Hao, Y.; Donnelly, L.A.; Moghbeli, T.; Tan, Y.S.; Lin, X.; Bellile, E.; et al. HPV16 drives cancer immune escape via NLRX1-mediated degradation of STING. J. Clin. Investig. 2020, 130, 1635–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yete, S.; D’Souza, W.; Saranath, D. High-Risk Human Papillomavirus in Oral Cancer: Clinical Implications. Oncology 2018, 94, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Nasman, A.; Du, J.; Dalianis, T. A global epidemic increase of an HPV-induced tonsil and tongue base cancer—potential benefit from a pan-gender use of HPV vaccine. J. Intern. Med. 2020, 287, 134–152. [Google Scholar] [CrossRef] [Green Version]
- Tumban, E. A Current Update on Human Papillomavirus-Associated Head and Neck Cancers. Viruses 2019, 11, 922. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.P.; Chen, T.C.; Chen, H.H.; Hsu, W.L.; Chang, Y.L. Prevalence of current oral HPV infection among 100 betel nut chewers or cigarette smokers in Northern Taiwan. J. Formos Med. Assoc. 2019, 118 Pt 1, 203–208. [Google Scholar] [CrossRef]
- Zhai, L.; Tumban, E. Gardasil-9: A global survey of projected efficacy. Antivir. Res. 2016, 130, 101–109. [Google Scholar] [CrossRef]
- Tabata, K.; Yamazaki, Y.; Okada, M.; Fukumura, K.; Shimada, A.; Sun, Y.; Yasukawa, K.; Suzuki, T. Diarylheptanoids derived from Alpinia officinarum induce apoptosis, S-phase arrest and differentiation in human neuroblastoma cells. Anticancer Res. 2009, 29, 4981–4988. [Google Scholar]
- Patel, V.; Jakus, J.; Harris, C.M.; Ensley, J.F.; Robbins, K.C.; Yeudall, W.A. Altered expression and activity of G1/S cyclins and cyclin-dependent kinases characterize squamous cell carcinomas of the head and neck. Int. J. Cancer 1997, 73, 551–555. [Google Scholar] [CrossRef]
- Bhutia, S.K.; Panda, P.K.; Sinha, N.; Praharaj, P.P.; Bhol, C.S.; Panigrahi, D.P.; Mahapatra, K.K.; Saha, S.; Patra, S.; Mishra, S.R.; et al. Plant lectins in cancer therapeutics: Targeting apoptosis and autophagy-dependent cell death. Pharmacol. Res. 2019, 144, 8–18. [Google Scholar] [CrossRef]
- Tilija Pun, N.; Jang, W.J.; Jeong, C.H. Role of autophagy in regulation of cancer cell death/apoptosis during anti-cancer therapy: Focus on autophagy flux blockade. Arch. Pharm. Res. 2020, 43, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Ciechomska, I.A. The role of autophagy in cancer—characterization of crosstalk between apoptosis and autophagy; autophagy as a new therapeutic strategy in glioblastoma. Postepy Biochem. 2018, 64, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, Y.T.; Kuo, C.L.; Lin, J.J.; Huang, W.W.; Peng, S.F.; Chueh, F.S.; Bau, D.T.; Chung, J.G. Curcuminoids combined with gefitinib mediated apoptosis and autophagy of human oral cancer SAS cells in vitro and reduced tumor of SAS cell xenograft mice in vivo. Environ. Toxicol. 2018, 33, 821–832. [Google Scholar] [CrossRef]
- Lin, C.W.; Chin, H.K.; Lee, S.L.; Chiu, C.F.; Chung, J.G.; Lin, Z.Y.; Wu, C.Y.; Liu, Y.C.; Hsiao, Y.T.; Feng, C.H.; et al. Ursolic acid induces apoptosis and autophagy in oral cancer cells. Environ. Toxicol. 2019, 34, 983–991. [Google Scholar] [CrossRef]
- Ho, H.Y.; Lin, C.C.; Chuang, Y.C.; Lo, Y.S.; Hsieh, M.J.; Chen, M.K. Apoptotic effects of dehydrocrenatidine via JNK and ERK pathway regulation in oral squamous cell carcinoma. Biomed. Pharmacother. 2021, 137, 111362. [Google Scholar] [CrossRef]
- Chen, J.M.; Chen, P.Y.; Lin, C.C.; Hsieh, M.C.; Lin, J.T. Antimetastatic Effects of Sesamin on Human Head and Neck Squamous Cell Carcinoma through Regulation of Matrix Metalloproteinase-2. Molecules 2020, 25, 2248. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.J.; Tsai, T.L.; Hsieh, Y.S.; Wang, C.J.; Chiou, H.L. Erratum to: Dioscin-induced autophagy mitigates cell apoptosis through modulation of PI3K/Akt and ERK and JNK signaling pathways in human lung cancer cell lines. Arch. Toxicol. 2017, 91, 2495–2496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, E.F.; Nebreda, A.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, W.Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.F. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J. Recept Signal Transduct Res. 2015, 35, 600–604. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.-T.; Ho, H.-Y.; Lin, C.-C.; Chuang, Y.-C.; Lo, Y.-S.; Hsieh, M.-J.; Chen, M.-K. Platyphyllenone Induces Autophagy and Apoptosis by Modulating the AKT and JNK Mitogen-Activated Protein Kinase Pathways in Oral Cancer Cells. Int. J. Mol. Sci. 2021, 22, 4211. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084211
Liu Y-T, Ho H-Y, Lin C-C, Chuang Y-C, Lo Y-S, Hsieh M-J, Chen M-K. Platyphyllenone Induces Autophagy and Apoptosis by Modulating the AKT and JNK Mitogen-Activated Protein Kinase Pathways in Oral Cancer Cells. International Journal of Molecular Sciences. 2021; 22(8):4211. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084211
Chicago/Turabian StyleLiu, Yen-Tze, Hsin-Yu Ho, Chia-Chieh Lin, Yi-Ching Chuang, Yu-Sheng Lo, Ming-Ju Hsieh, and Mu-Kuan Chen. 2021. "Platyphyllenone Induces Autophagy and Apoptosis by Modulating the AKT and JNK Mitogen-Activated Protein Kinase Pathways in Oral Cancer Cells" International Journal of Molecular Sciences 22, no. 8: 4211. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084211