
Novel Quercetin Derivative of 3,7-Dioleylquercetin Shows Less Toxicity and Highly Potent Tyrosinase Inhibition Activity

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
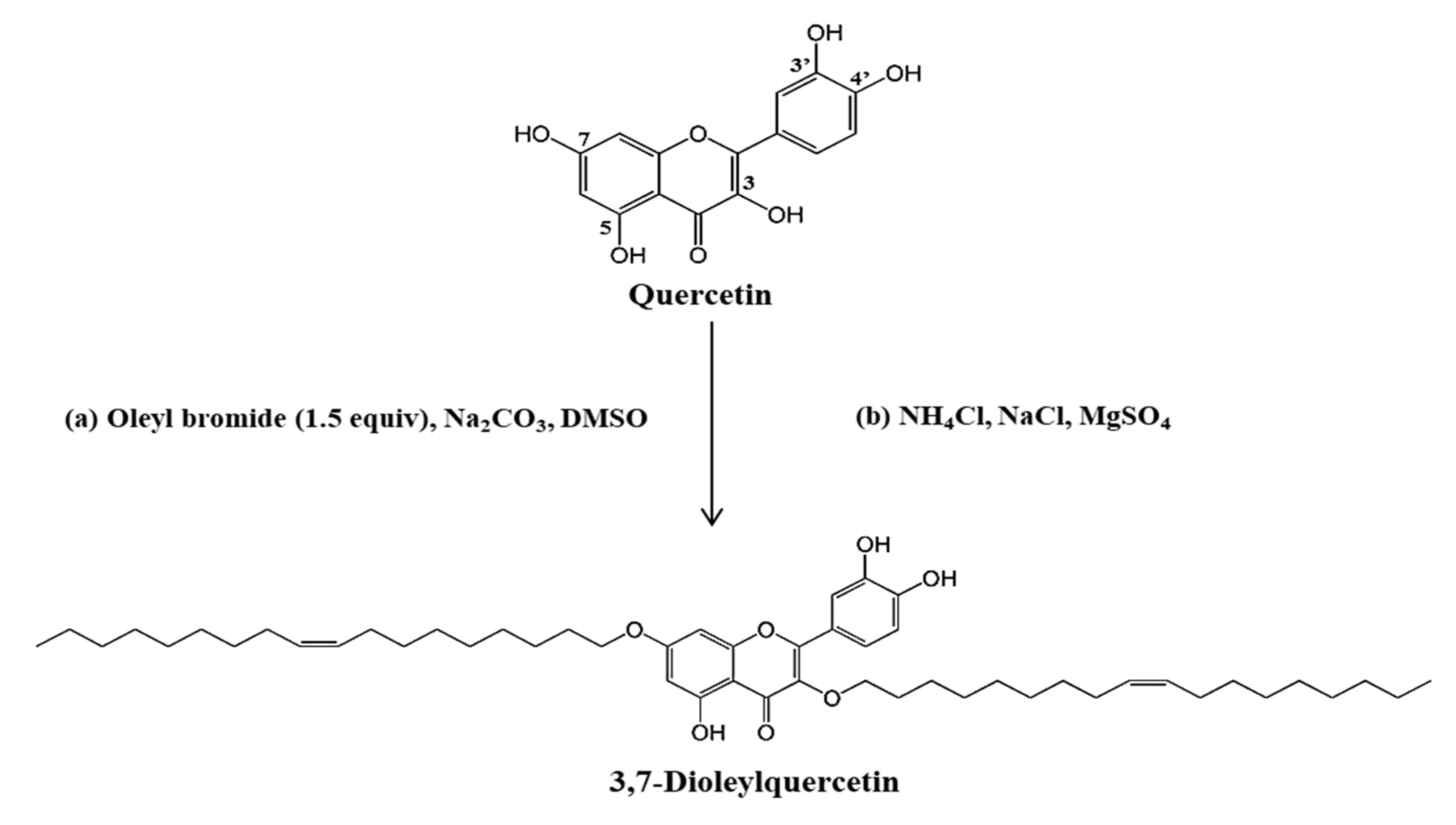
2.1. Synthesis and Identification of Novel Compounds
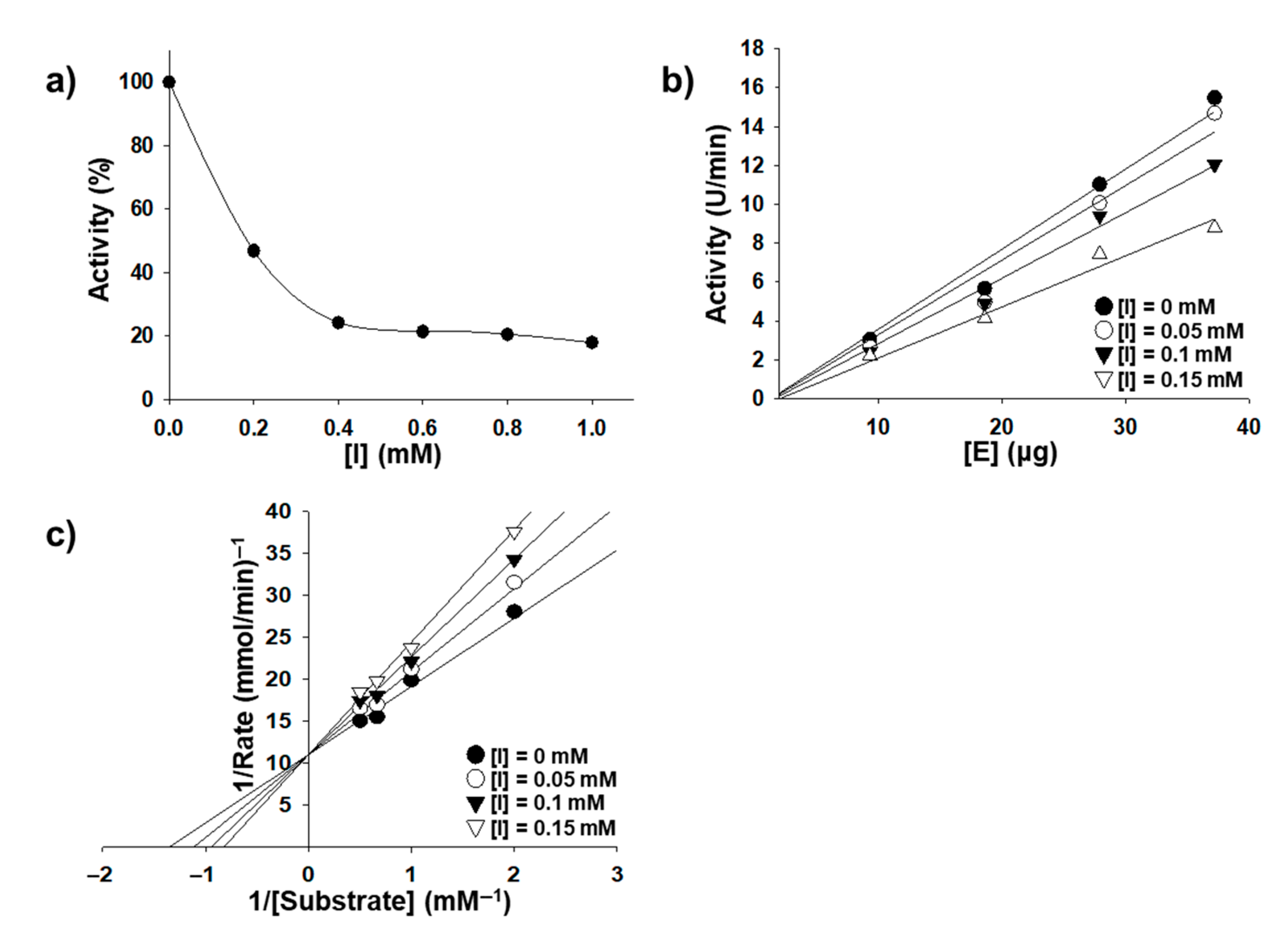
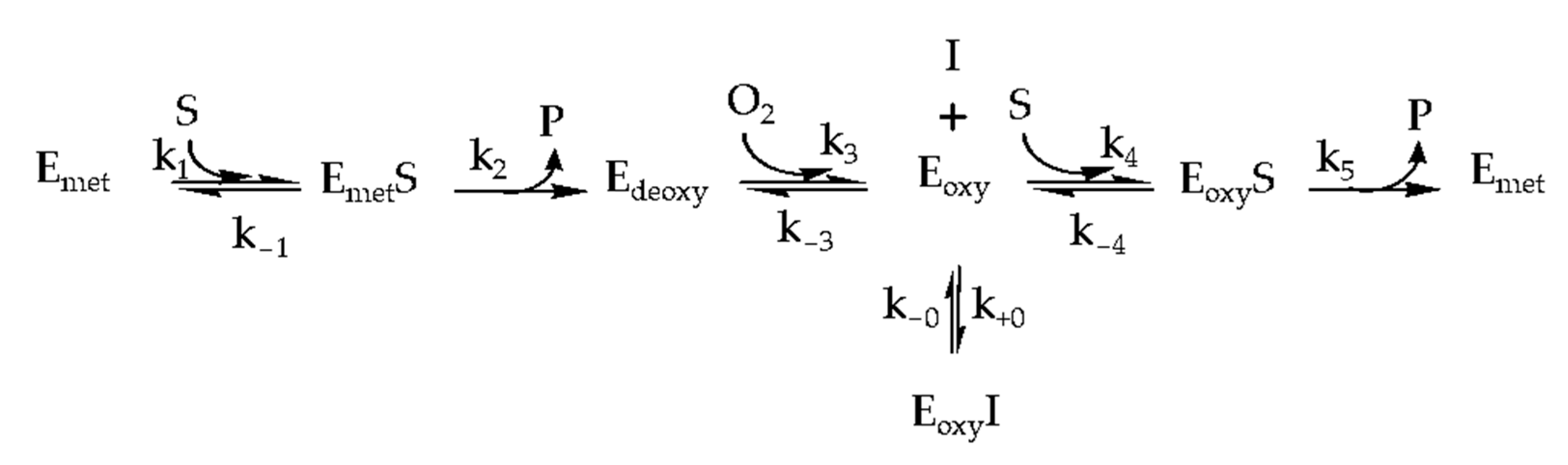
2.2. Inhibition Kinetics
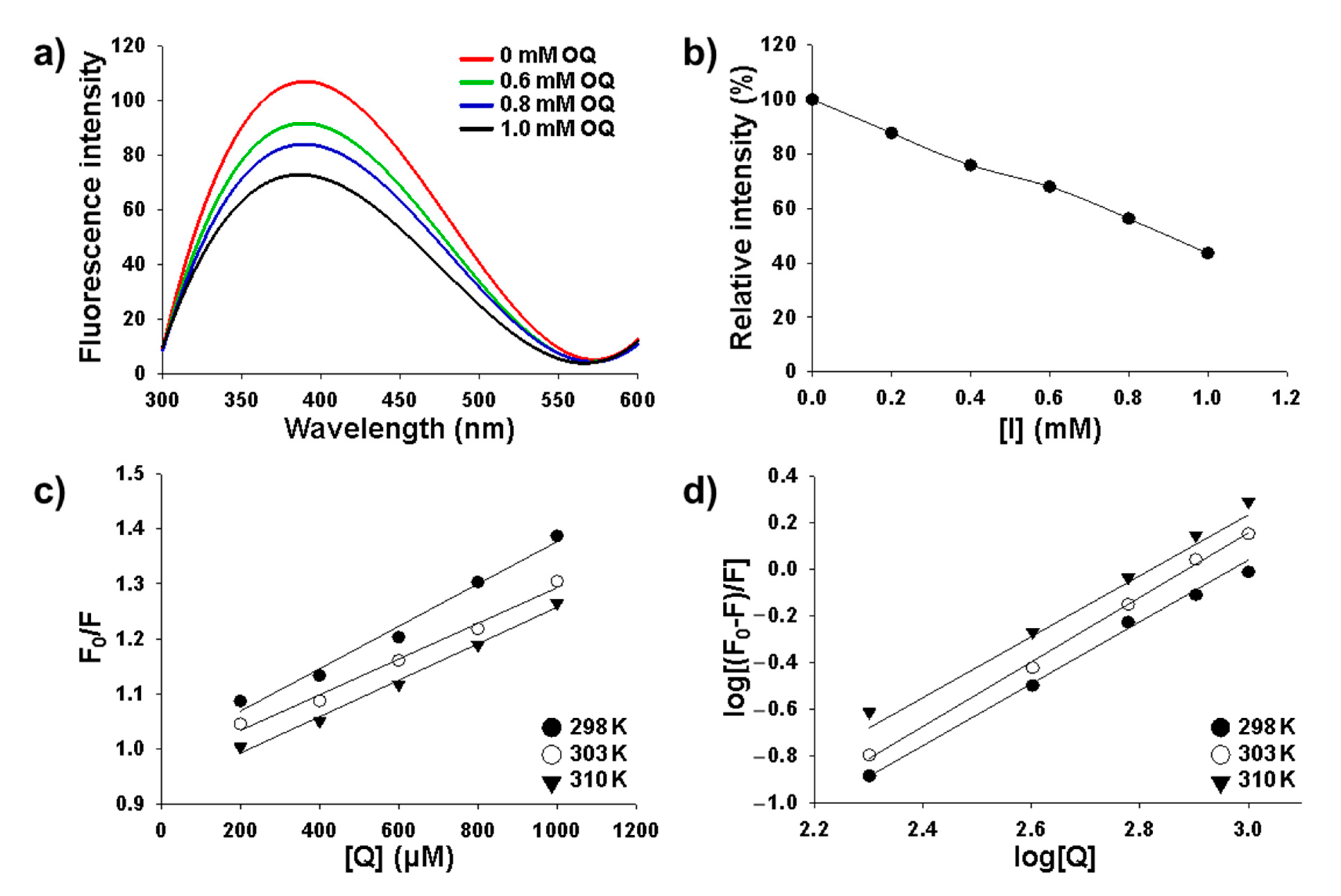
2.3. Intrinsic Fluorescence Quenching Analysis
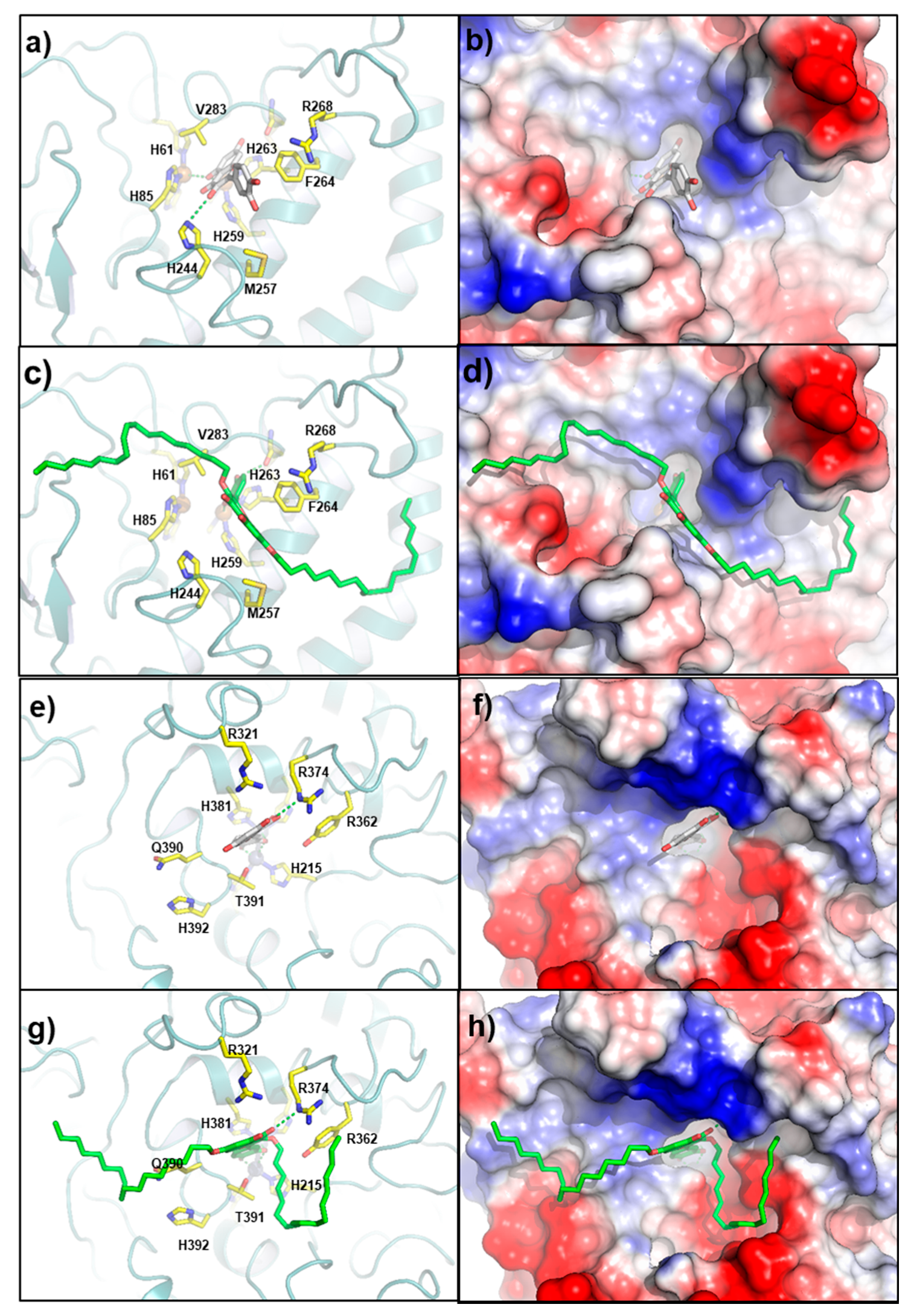
2.4. Molecular Docking Study
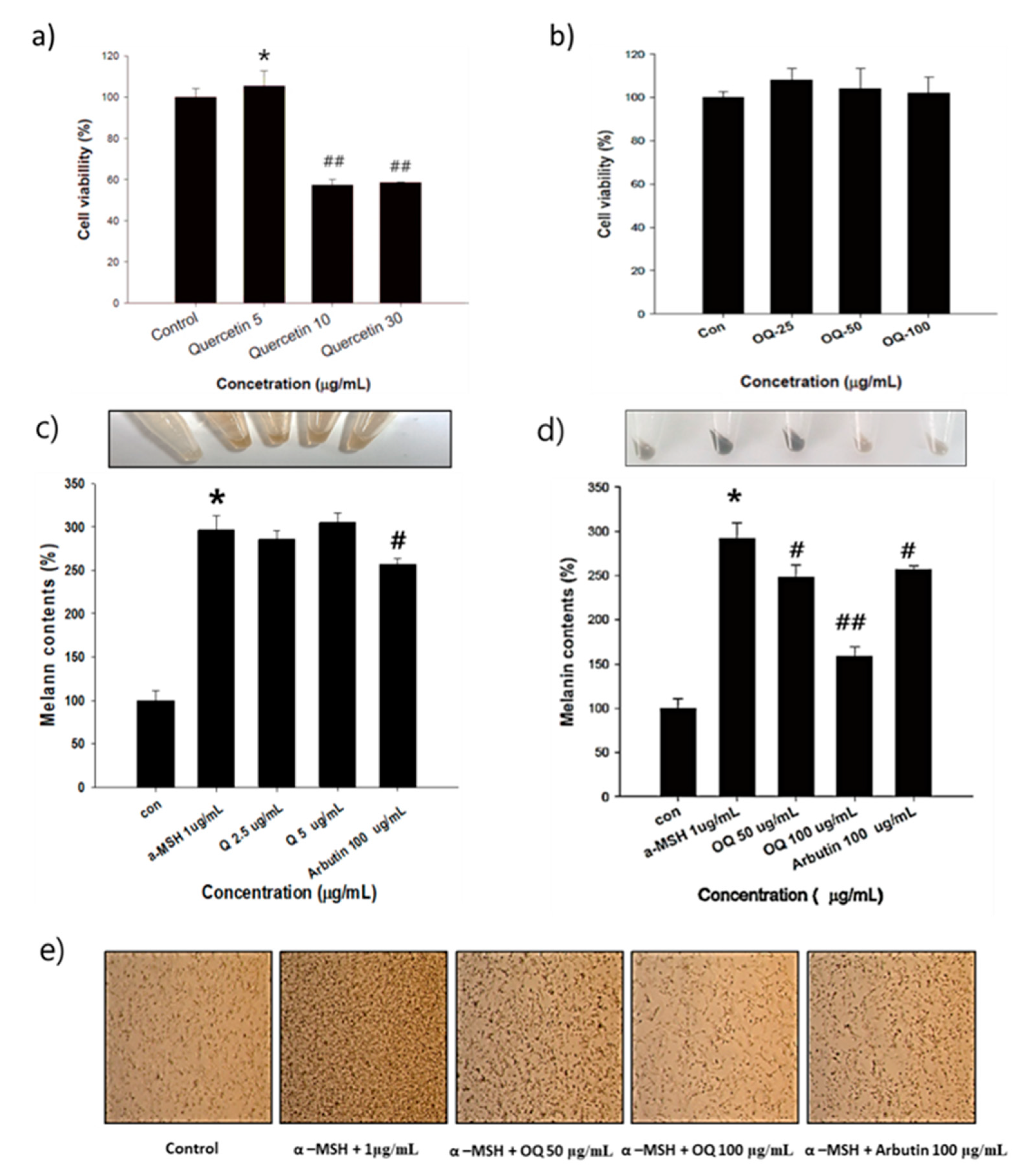
2.5. Cell Viability and Melanin Contents of Native Quercetin and OQ
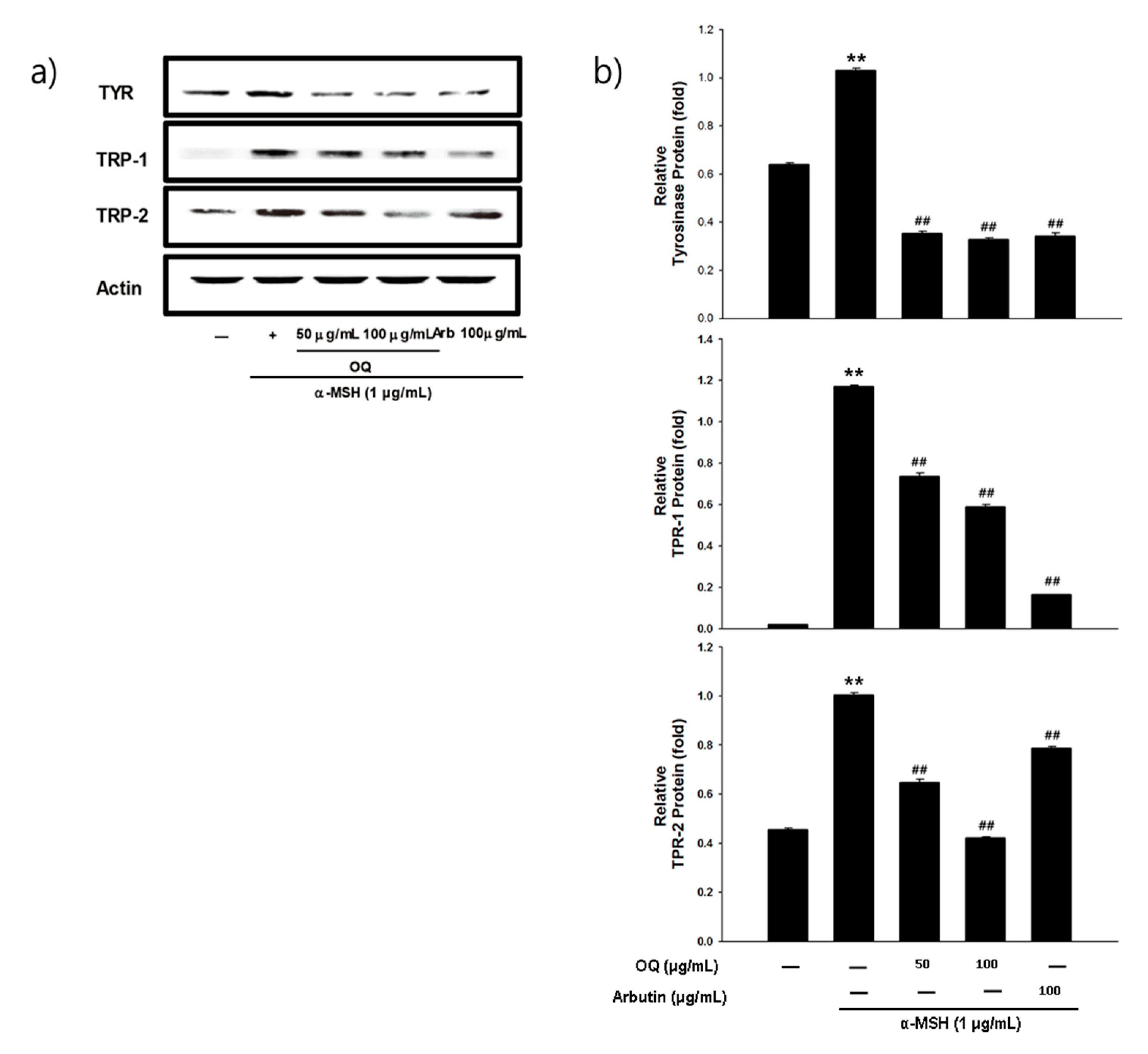
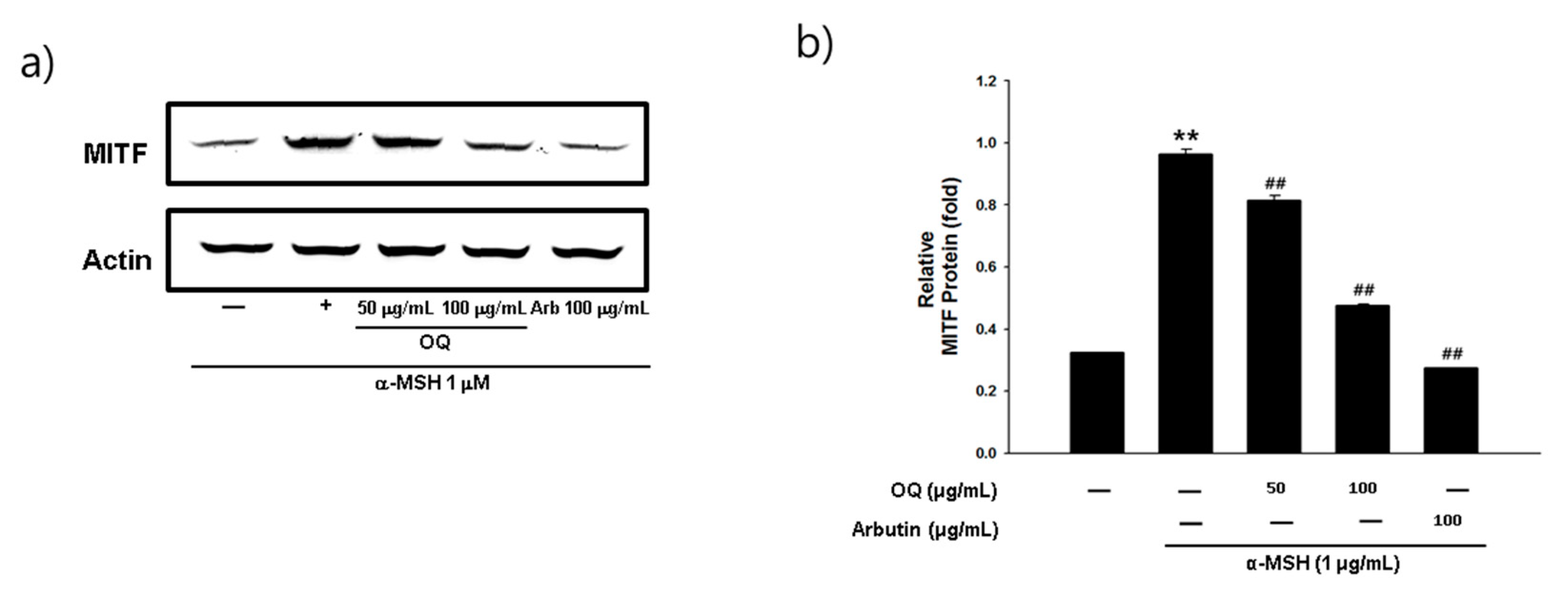
2.6. Effet of OQ on Anti-Melanogenesis-Elated Proteinsin B16F10 Cell Line
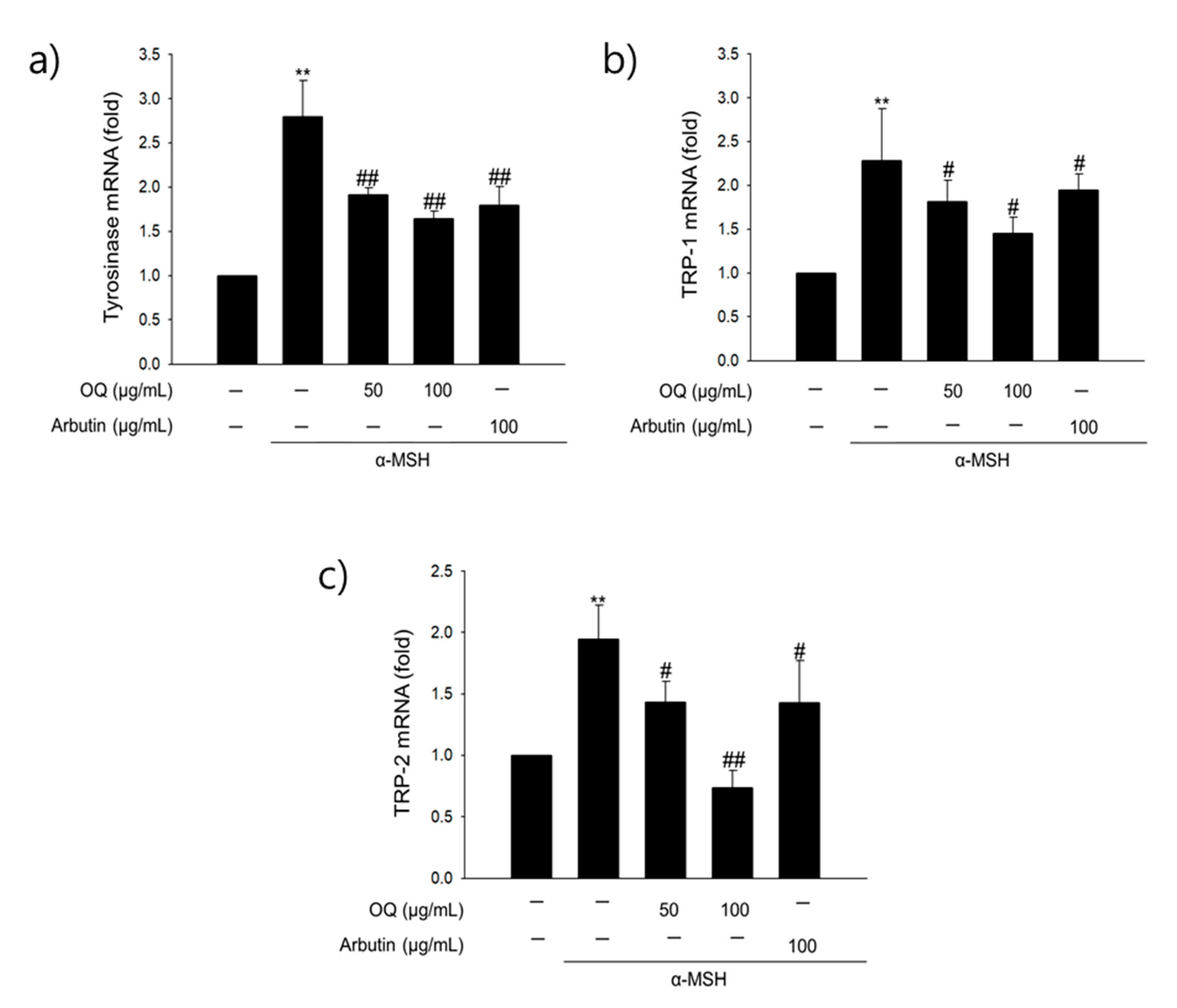
2.7. Effect of OQ on Anti-Melanogenesis-Related Genes in B16F10 Cells
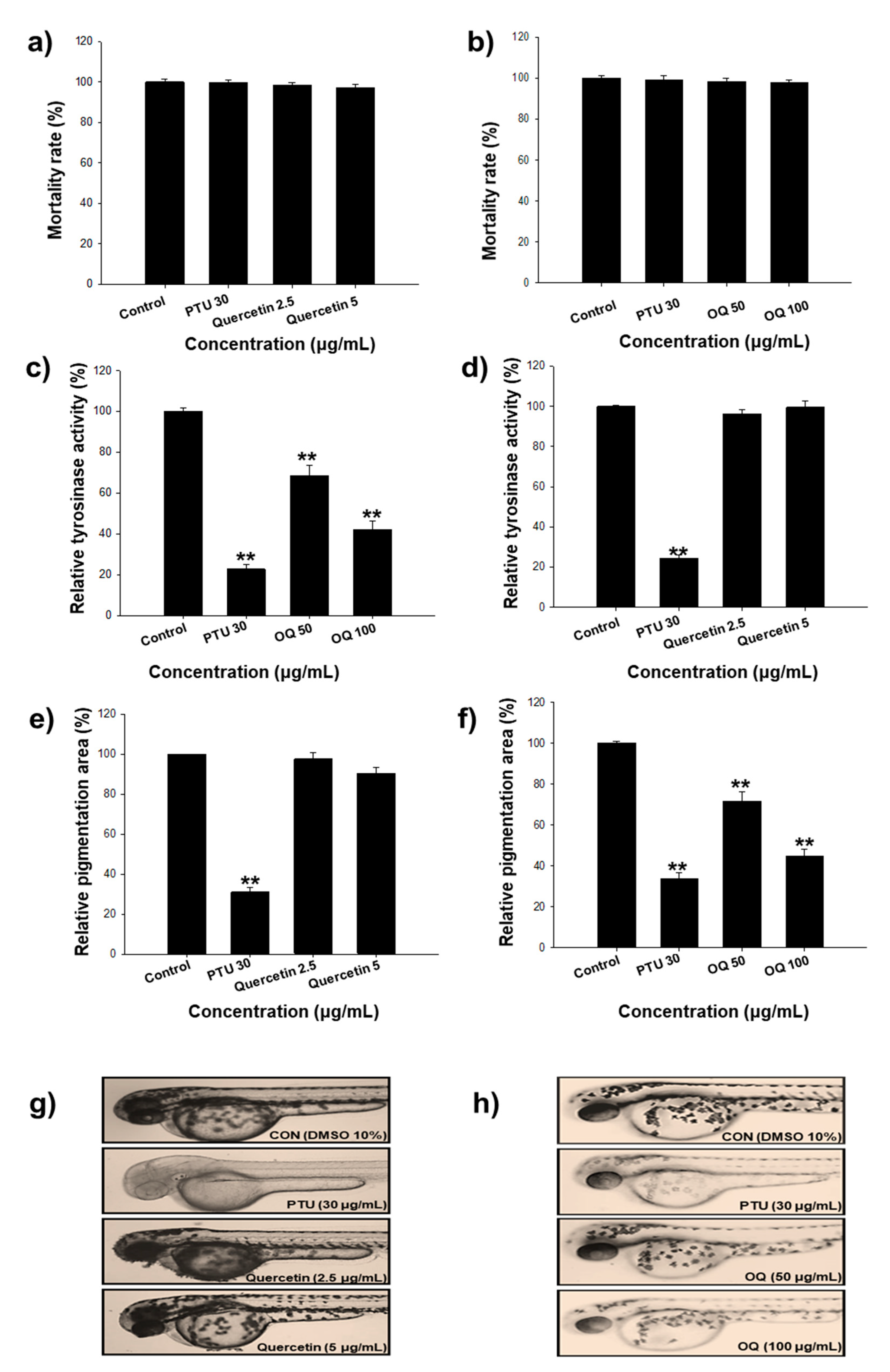
2.8. Zebrafish Model Study
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Synthesis of 3,7-dioleylquercetin (OQ)
4.3. In Vitro Tyrosinase Inhibition Assay and Enzyme Kinetics
4.4. Quenching Study
4.5. Molecular Docking Procedure
4.6. Cell Culture
4.7. MTT Cell Viability Assay
4.8. Measurement of Melanin Content
4.9. RNA Isolation and Reverse Transcription Polymerase Chanin Reaction (RT-PCR)
4.10. Zebrafish Study
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| α-MSH | α-Melanocyte-stimulating hormone |
| Arg | arginine |
| CREB | cAMP response element-binding protein |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMSO | dimethyl sulfoxide |
| EtOAc | ethyl acetate |
| FBS | fetal bovine serum |
| Gln | glutamine |
| His | histidine |
| HPLC | high performance liquid chromatography |
| hTRP-1 | human tyrosinase-related protein-1 |
| IC50 | inhibitory concentration 50 |
| L-DOPA | L-3,4-dihydroxyphenylalanine |
| Met | methionine |
| MITF MPLC | microphthalmia-associated transcription factor medium pressure liquid chromatography |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| OQ | 3,7-dioleylquercetin |
| PBS | phosphate-buffered saline |
| p-CREB | phosphorylation of cAMP response element-binding protein |
| Phe | phenylalanine |
| PKA | protein kinase A |
| p-PKA | phosphorylation of protein kinase A |
| PTU | 1-phenyl-2-thiourea |
| Q | quercetin |
| RT-PCR | reverse transcription polymerase chain reaction |
| Thr | threonine |
| TRP-1 | tyrosinase-related protein-1 |
| TRP-2 | tyrosinase-related protein-2 |
| Val | valine |
References
- Taieb, A.; Cario-Andre, M.; Briganti, S.; Picardo, M. Inhibitors and enhancers of melanogenesis. In Melanins and Melanosomes: Biosynthesis, Biogenesis, Physiological, and Pathological Functions; Borovansky, J., Riley, P.A., Eds.; Wiley-Blackwell: Weinheim, Germany, 2011; Volume 5, pp. 117–166. [Google Scholar]
- Solano, F.; Briganti, S.; Picardo, M.; Ghanem, G. Hypopigmenting agents: An updated review on biological, chemical and clinical aspects. Pigment Cell Res. 2006, 19, 550–571. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.H.; Shin, H.J. Anti-melanogenesis effect of quercetin. Cosmetics 2016, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- Couteau, C.; Coiffard, L. Overview of skin whitening agents: Drugs and cosmetic products. Cosmetics 2016, 3, 27. [Google Scholar] [CrossRef]
- Sugimoto, K.; Nomura, K.; Nishimura, T.; Kiso, T.; Sugimoto, K.; Kuriki, T. Syntheses of α-arbutin-α-glycosides and their inhibitory effects on human tyrosinase. J. Biosci. Bioeng. 2005, 99, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Nagata, H.; Takekoshi, S.; Takeyama, R.; Homma, T.; Osamura, R.Y. Quercetin enhances melanogenesis by increasing the activity and synthesis of tyrosinase in human melanoma cells and normal human melanocytes. Pigment Cell Res. 2004, 17, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Takekoshi, S.; Matsuzaki, K.; Kitatani, K. Quercetin stimulates melanogenesis in hair follicle melanocyte of the mouse. Tokai J. Exp. Clin. Med. 2013, 38, 129–134. [Google Scholar]
- Takekoshi, S.; Nagata, H.; Kitatani, K. Flavonoids enhance melanogenesis in human melanoma cells. Tokai J. Exp. Clin. Med. 2014, 39, 116–121. [Google Scholar] [PubMed]
- Arung, E.T.; Kusuma, I.W.; Shimizu, K.; Kondo, R. Tyrosinase inhibitory effect of quercetin 4′-O-b-D-glucopyranoside from dried skin of red onion (Allium cepa). Nat. Prod. Res. 2011, 25, 256–263. [Google Scholar] [CrossRef]
- Strzepek-Gomolka, M.; Gawel-Beben, K.; Angelis, A.; Antosiewicz, B.; Sakipova, Z.; Kozhanova, K.; Glowniak, K.; Kukula-Koch, W. Identification of mushroom and murine tyrosinase inhibitors from Achillea biebersteinii Afan. extract. Molecules 2021, 26, 964. [Google Scholar] [CrossRef]
- Chao, H.C.; Najjaa, H.; Villareal, M.O.; Ksouri, R.; Han, J.; Neffati, M.; Isoda, H. Arthrophytumscoparium inhibits melanogenesis through the down-regulation of tyrosinase and melanogenic gene expressions in B16 melanoma cells. Exp. Dermatol. 2013, 22, 131–136. [Google Scholar] [CrossRef]
- Taira, J.; Tsuchida, E.; Uehara, M.; Ohhama, N.; Ohmine, W.; Ogi, T. The leaf extract of Mallotus japonicas and its major active constituent, rutin, suppressed on melanin production in murine B16F1 melanoma. Asian Pac. J. Trop. Biomed. 2015, 5, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Kato, K.; Ninomiya, M.; Tanaka, K.; Koketsu, M. Effects of functional groups and sugar composition of quercetin derivatives on their radical scavenging properties. J. Nat. Prod. 2016, 79, 1808–1814. [Google Scholar] [CrossRef]
- Lee, M.K.; Moon, S.S.; Lee, S.E.; Bok, S.H.; Jeong, T.S.; Park, Y.B.; Choi, M.S. Naringenin 7-O-cetyl ether as inhibitor of HMG-CoA reductase and modulator of plasma and hepatic lipids in high cholesterol-fed rats. Bioorg. Med. Chem. 2003, 11, 393–398. [Google Scholar] [CrossRef]
- Chen, Q.X.; Kubo, I. Kinetics of mushroom tyrosinase inhibition by quercetin. J. Agric. Food Chem. 2002, 50, 4108–4112. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, G.; Hu, Y.; Ma, Y. Effect of luteolin on xanthine oxidase: Inhibition kinetics and interaction mechanism merging with docking simulation. Food Chem. 2013, 141, 3766–3773. [Google Scholar] [CrossRef] [PubMed]
- Tsou, C.L. Kinetics of substrate reaction during irreversible modification of enzyme activity. In Advances in Enzymology and Related Areas of Molecular Biology; Meister, A., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1988; Volume 61, pp. 381–436. [Google Scholar]
- Vaezi, M.; Behbehani, G.R.; Gheibi, N.; Farasat, A. Thermodynamic, kinetic and docking studies of some unsaturated fatty acids-quercetin derivatives as inhibitors of mushroom tyrosinase. AIMS Biophys. 2020, 7, 393–410. [Google Scholar] [CrossRef]
- Kim, D.H.; Park, J.Y.; Kim, J.H.; Han, C.K.; Yoon, J.H.; Kim, N.D.; Seo, J.H.; Lee, C.H. Flavonoids as mushroom tyrosinase inhibitors: A fluorescence quenching study. J. Agric. Food Chem. 2006, 54, 935–941. [Google Scholar] [CrossRef]
- Chai, W.M.; Lin, M.Z.; Song, F.Z.; Wang, Y.X.; Xu, K.L.; Huang, J.X.; Fu, J.P.; Peng, Y.Y. Rifampicin as a novel tyrosinase inhibitor. Int. J. Biol. Macromol. 2017, 102, 425–430. [Google Scholar] [CrossRef]
- Yu, Q.; Fan, L.; Duan, Z. Five individual polyphenols as tyrosinase inhibitors: Inhibitory activity, synergistic effect, action mechanism, and molecular docking. Food Chem. 2019, 297, 124910. [Google Scholar] [CrossRef]
- Oh, E.J.; Park, J.I.; Lee, J.E.; Myung, C.H.; Kim, S.Y.; Chang, S.E.; Hwang, J.S. A novel role of serotonin receptor 2B agonist as an anti-melanogenesis agent. Int. J. Mol. Sci. 2016, 17, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trouillas, P.; Marsal, P.; Siri, D.; Lazzaroni, R.; Duroux, J.L. A DFT study of the reactivity of OH groups in quercetin and taxifolin antioxidants: The specificity of the 3-OH site. Food Chem. 2006, 97, 679–688. [Google Scholar] [CrossRef]
- Musialik, M.; Kuzmicz, R.; Pawłowski, T.S.; Litwinienko, G. Acidity of hydroxyl groups: An overlooked influence on antiradical properties of flavonoids. J. Org. Chem. 2009, 74, 2699–2709. [Google Scholar] [CrossRef]
- Lee, H.J.; Kerr, R.A.; Korshavn, K.J.; Lee, J.; Kang, J.; Ramamoorthy, A.; Ruotolo, B.T.; Lim, M.H. Effects of hydroxyl group variations on a flavonoid backbone toward modulation of metal-free and metal-induced amyloid-β aggregation. Inorg. Chem. Front. 2016, 3, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Rao, K.V.; Owoyale, J.A. Partial methylation of quercetin: Direct synthesis of tamarixetin, ombuin and ayanin. J. Heterocycl. Chem. 1976, 13, 1293–1295. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Fang, Z.; Jin, H.; Chen, Y.; He, L. Selective monomethylation of quercetin. Synthesis 2010, 23, 3980–3986. [Google Scholar]
- Bao, X.R.; Liao, H.; Qu, J.; Sun, Y.; Guo, X.; Wang, E.X.; Zhen, Y.H. Synthesis, characterization and cytotoxicity of alkylated quercetin derivatives. Iran. J. Pharm. Res. 2016, 15, 329. [Google Scholar]
- Al-Jabban, S.M.; Zhang, X.; Chen, G.; Mekuria, E.A.; Rakotondraibe, L.H.; Chen, Q.H. Synthesis and anti-proliferative effects of quercetin derivatives. Nat. Prod. Commun. 2015, 10, 2113–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Park, Y.; Cho, S.; Burapan, S.; Han, J. Synthesis of alkyl quercetin derivatives. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 343–348. [Google Scholar] [CrossRef]
- Wilcox, D.E.; Porras, A.G.; Hwang, Y.T.; Lerch, K.; Winkler, M.E.; Solomon, E.I. Substrate analog binding to the coupled binuclear copper active site in tyrosinase. J. Am. Chem. Soc. 1985, 107, 4015–4027. [Google Scholar] [CrossRef]
- Fuller, B.B.; Lunsford, J.B.; Iman, D.S. Alpha-melanocyte-stimulating hormone regulation of tyrosinase in Cloudman S-91 mouse melanoma cell cultures. J. Biol. Chem. 1987, 262, 4024–4033. [Google Scholar] [CrossRef]
- Park, H.Y.; Russakovsky, V.; Ao, Y.; Fernandez, E.; Gilchrest, B.A. α-melanocyte stimulating hormone-induced pigmentation is blocked by depletion of protein kinase C. Exp. Cell Res. 1996, 227, 70–79. [Google Scholar] [CrossRef]
- Kuzumaki, T.; Matsuda, A.; Wakamatsu, K.; Ito, S.; Ishikawa, K. Eumelanin biosynthesis is regulated by coordinate expression of tyrosinase and tyrosinase-related protein-1 genes. Exp. Cell Res. 1993, 207, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Tseng, T.S.; Hsiao, N.W.; Lin, Y.L.; Wen, Z.H.; Tsai, C.C.; Lee, Y.C.; Lin, H.H.; Tsai, K.C. Discovery of highly potent tyrosinase inhibitor, T1, with significant anti-melanogenesis ability by zebrafish in vivo assay and computational molecular modeling. Sci. Rep. 2015, 5, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, D.K.; Jung, S.H.; Jeong, J.H.; Yoo, H.M.; Lee, I.S.; Shin, H.S. The antimelanogenic effect of inularin isolated from flowers of Inula britannica on B16F10 melanoma cells and zebrafish embryos. J. Microbiol. Biotechnol. 2020, 30, 749–752. [Google Scholar] [CrossRef]
- Chung, S.; Lim, G.J.; Lee, J.Y. Quantitative analysis of melanin content in a three-dimensional melanoma cell culture. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Zhang, G.; Hu, X.; Xu, X.; Gong, D. Quercetin as a tyrosinase inhibitor: Inhibitory activity, conformational change and mechanism. Food Res. Int. 2017, 100, 226–233. [Google Scholar] [CrossRef]
- Chai, W.M.; Lin, M.Z.; Wang, Y.X.; Xu, K.L.; Huang, W.Y.; Pan, D.D.; Zou, Z.R.; Peng, Y.Y. Inhibition of tyrosinase by cherimoya pericarp proanthocyanidins: Structural characterization, inhibitory activity and mechanism. Food Res. Int. 2017, 100, 731–739. [Google Scholar] [CrossRef]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Matthew, P.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; et al. Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- Halgren, T.A.; Murphy, R.B.; Friesner, R.A.; Beard, H.S.; Frye, L.L.; Pollard, W.T.; Banks, J.L. Glide: A new approach for rapid, accurate docking and scoring. 2. Enrichment factors in database screening. J. Med. Chem. 2004, 47, 1750–1759. [Google Scholar] [CrossRef]
- Jacobson, M.P.; Pincus, D.L.; Rapp, C.S.; Day, T.J.; Honig, B.; Shaw, D.E.; Friesner, R.A. A hierarchical approach to all-atom protein loop prediction. Proteins. 2004, 55, 351–367. [Google Scholar] [CrossRef] [Green Version]
- Roos, K.; Wu, C.; Damm, W.; Reboul, M.; Stevenson, J.M.; Lu, C.; Dahlgren, M.K.; Mondal, S.; Chen, W.; Wang, L.; et al. OPLS3e: Extending Force Field Coverage for Drug-Like Small Molecules. J. Chem. Theory Comput. 2019, 15, 1863–1874. [Google Scholar] [CrossRef]
- Lai, X.; Wichers, H.J.; Soler-Lopez, M.; Dijkstra, B.W. Structure of human tyrosinase related protein 1 reveals a binuclear zinc active site important for melanogenesis. Angew. Chem. Int. Ed. 2017, 56, 9812–9815. [Google Scholar] [CrossRef] [PubMed]
- Hosoi, J.; Abe, E.; Suda, T.; Kuroki, T. Regulation of melanin synthesis of B16 mouse melanoma cells by 1α, 25-dihydroxyvitamin D3 and retinoic acid. Cancer Res. 1985, 45, 1474–1478. [Google Scholar]
- Choi, M.H.; Jo, H.G.; Yang, J.H.; Ki, S.H.; Shin, H.J. Antioxidative and anti-melanogenic activities of bamboo stems (Phyllostachys nigra variety henosis) via PKA/CREB-mediated MITF downregulation in B16F10 melanoma cells. Int. J. Mol. Sci. 2018, 19, 409. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liu, K.; Wang, X.; He, Q.; Chen, X. Toxic effects of celastrol on embryonic development of zebrafish (Danio rerio). Drug Chem. Toxicol. 2011, 34, 61–65. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Native Quercetin a,b | 3,7-dioleylquercetin a | Quercetin-7-oleate c | |
|---|---|---|---|
| IC50 | 0.4714 mM (0.13 mM) | 0.0987 mM | 0.71 mM |
| Km | 0.7524 mM (0.84 mM) | 0.736 mM | 1.11 mM |
| Vm | 75.6 U/min (122 U/min) | 30.2 U/min | 4.98 mM |
| Inhibition | Reversible (reversible) | Reversible | Reversible |
| Inhibition type | Competitive (competitive) | Competitive | Competitive |
| Ki | 0.2459 mM (0.0386mM) | 0.232 mM | 0.43 mM |
| k+0 | 0.0217 mM−1s−1 (0.0216 mM−1s−1) | 0.0298 mM−1s−1 | - |
| 0.0235 mM−1s−1 (0.0219 mM−1s−1) | 0.0272 mM−1s−1 | - | |
| k–0 | 0.49 × 10−2 s−1 (0.832 × 10−3s−1) | 0.43 × 10−2 s−1 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, M.-H.; Yang, S.-H.; Kim, D.-S.; Kim, N.D.; Shin, H.-J.; Liu, K. Novel Quercetin Derivative of 3,7-Dioleylquercetin Shows Less Toxicity and Highly Potent Tyrosinase Inhibition Activity. Int. J. Mol. Sci. 2021, 22, 4264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084264
Choi M-H, Yang S-H, Kim D-S, Kim ND, Shin H-J, Liu K. Novel Quercetin Derivative of 3,7-Dioleylquercetin Shows Less Toxicity and Highly Potent Tyrosinase Inhibition Activity. International Journal of Molecular Sciences. 2021; 22(8):4264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084264
Chicago/Turabian StyleChoi, Moon-Hee, Seung-Hwa Yang, Da-Song Kim, Nam Doo Kim, Hyun-Jae Shin, and Kechun Liu. 2021. "Novel Quercetin Derivative of 3,7-Dioleylquercetin Shows Less Toxicity and Highly Potent Tyrosinase Inhibition Activity" International Journal of Molecular Sciences 22, no. 8: 4264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084264