
Endothelin ETB Receptor-Mediated Astrocytic Activation: Pathological Roles in Brain Disorders

Laboratory of Pharmacology, Kobe Pharmaceutical University, 4-19-1 Motoyama-Kita Higashinada, Kobe 668-8558, Japan
Int. J. Mol. Sci. 2021, 22(9), 4333; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094333
Submission received: 15 March 2021
/
Revised: 16 April 2021
/
Accepted: 19 April 2021
/
Published: 21 April 2021
(This article belongs to the Special Issue Molecular Mechanisms of Astrocytic Activation)
Abstract
:In brain disorders, reactive astrocytes, which are characterized by hypertrophy of the cell body and proliferative properties, are commonly observed. As reactive astrocytes are involved in the pathogenesis of several brain disorders, the control of astrocytic function has been proposed as a therapeutic strategy, and target molecules to effectively control astrocytic functions have been investigated. The production of brain endothelin-1 (ET-1), which increases in brain disorders, is involved in the pathophysiological response of the nervous system. Endothelin B (ETB) receptors are highly expressed in reactive astrocytes and are upregulated by brain injury. Activation of astrocyte ETB receptors promotes the induction of reactive astrocytes. In addition, the production of various astrocyte-derived factors, including neurotrophic factors and vascular permeability regulators, is regulated by ETB receptors. In animal models of Alzheimer’s disease, brain ischemia, neuropathic pain, and traumatic brain injury, ETB-receptor-mediated regulation of astrocytic activation has been reported to improve brain disorders. Therefore, the astrocytic ETB receptor is expected to be a promising drug target to improve several brain disorders. This article reviews the roles of ETB receptors in astrocytic activation and discusses its possible applications in the treatment of brain disorders.
1. Introduction
Astrocytes are the most abundant glial cells in the brain and play roles in supplying nutrients to nerve cells, in reinforcing synaptic structures at nerve endings, and in supporting the limited entry of blood components [1,2]. Under physiological conditions, astrocytes modulate neurotransmission through the clearance of neurotransmitters from the synaptic cleft; maintenance of extracellular fluid ion concentration; and release of gliotransmitters, such as ATP, D-serine, and L-glutamate. In brain disorders, astrocytes alter their phenotype to a reactive phenotype. Reactive astrocytes, which are characterized by hypertrophy of cell bodies with increased expression of glial fibrillary acidic protein (GFAP), are highly proliferative and often form glial scars at the damaged areas of nerve tissues. Reactive astrocytes produce various bioactive factors that regulate the pathophysiological responses of the injured nervous system [3,4]. Cytokines and chemokines produced by reactive astrocytes promote neuroinflammation and exacerbate nervous system damage [5,6]. Conversely, increased production of neurotrophic factors supports survival and synaptic formation of damaged neurons as well as neurogenesis from neural progenitor cells [7,8,9,10], which underlie the recovery of impaired nerve function. Thus, reactive astrocytes have both detrimental and beneficial effects on the injured nervous system. Therefore, it has been proposed that the regulation of astrocytic functions can lead to improvements in several brain disorders [11,12,13,14,15]. Given this background, the factors that regulate astrocytic activation have been investigated. Endothelin (ET) is a family of peptides that are also potent vasoconstrictors [16,17]. ETs are also present in nervous tissue. ET-1 production in the brain is increased during many neurological conditions such as ischemic stroke [18,19,20,21,22,23], traumatic brain injury (TBI) [24,25], Alzheimer’s disease (AD) [26], amyotrophic lateral sclerosis [27], multiple sclerosis [28], and viral infection [29,30]. Increased ET-1 in the brain causes various pathophysiological reactions in the injured nervous system, including exacerbation of neuroinflammation [31], vasospasm-mediated ischemic injury [32], angiogenesis [33], and neurogenesis [34,35,36]. Endothelin B (ETB) receptors are highly expressed in astrocytes [37,38]. Administration of a selective ETB agonist into rat brains promoted the induction of reactive astrocytes [39], whereas ETB antagonists reduced it in animal models of brain injury [40,41,42]. Therefore, the ETB receptor is expected to be a target molecule to control the functions of astrocytic activation. In addition, examinations to improve brain disorders by modulating ETB-receptor-mediated signal have being conducted. As such, this article reviews the roles of ET-1/ETB receptor signals in astrocytic activation and discusses the possibility of ETB receptor agonists and antagonists being beneficial for some brain disorders.
2. Overview of Endothelin
2.1. Endothelin Ligands
The ET peptide family is composed of three isoforms of 21 amino acid cyclic peptides that are the product of different genes. Since its discovery in 1989 as a novel family of peptides produced by the vascular endothelium [16], the function of ET has attracted attention due to its role in the circulatory system because of its strong vasoconstrictor effect. However, shortly after discovery, ET ligands were shown to be present in various tissues, including the central nervous system, and have a wide range of functions, such as cell proliferation, differentiation, and neurotransmission [17]. ET-1 was the subtype discovered first and is expressed in many tissue types, including nerve tissues. Some studies have shown that the expression of ET-2 is largely limited to the gastrointestinal tract, sex organs, and pituitary gland [43,44,45]. Chang et al. found that ET-2 is involved in energy homeostasis, thermoregulation, and the maintenance of lung morphology and function [46]. ET-3 is abundantly expressed in the intestine, pituitary gland, and brain [47]. Genetic deficiency of ET-3 function in the gastrointestinal tract is associated with the development of Hirschsprung disease [48,49].
ET-1 biosynthesis is regulated by transcription and processing after translation of the prepro-ET-1 gene, which is a precursor protein. Transcription of ET-1 is promoted by transcription factors including AP-1, GATA-2, Smad, HIF-1α, and NFkB [50], for which the binding sites are present in the 5′-flanking region of the gene. These transcription factors are activated not only by cytokines and hormones but also by pathological conditions such as hypoxia [51] and mechanical stress [16], which underlie the increase in ET-1 production under pathological conditions. The prepro-ET-1 protein is cleaved by neutral endopeptidase to form an inactive precursor called big-ET (Figure 1A). The conversion of big-ET to ET-1 is mediated by a family of endothelin-converting enzymes (ECEs). ECEs have three isoforms, ECE-1, ECE-2, and ECE-3, which differ in cell distribution, localization, and substrate specificity [17]. ECE-1 and ECE-2 are the prominent isoforms that cleave big-ET-1. These ECEs are involved in the production of ET-1 and the degradation of amyloid-β proteins [52,53], which is a causative factor of AD. Therefore, the role of ECEs in AD pathology has been investigated from the perspective of ET production and degradation of amyloid-β proteins [14,54].
2.2. Endothelin Receptors
ETs exhibit their actions through two G-protein-conjugated receptor subtypes: the ETA and ETB receptors. Among the endogenous ET ligands, the ETA receptor shows higher affinities for ET-1 and ET-2 than for ET-3. However, the ETB receptor has an equal affinity for these three ET ligands (Figure 1B) [17]. Both of these ET receptors are linked to the Gq protein and increase intracellular Ca2+ by activating phospholipase C (PLC) [55,56,57]. For adenylate cyclase-mediated signals, the ETA and ETB receptors have different regulatory mechanisms, wherein the ETA type is Gs-linked to increase cAMP, whereas ETB is linked to Gi and suppresses the signal [58,59]. In addition, the ETA and ETB receptors are linked to the G12/13 protein. Signals triggered by the G12/13 protein activate the Rho protein, a low molecular weight G protein, and stimulate Rho-associated protein kinase (ROCK) [60], which regulates cellular proliferation, Ca2+, and cytoskeletal actin reorganization [61,62,63,64]. Similar to other G-protein-coupled receptors, ET receptors form dimers. Evance et al. showed that the ETA/ETB heterodimer induces a long-lasting intracellular Ca2+ increase in response to ET-1, which was not observed by the activation of ETA or ETB receptor homodimers [65,66].
ET receptors are also expressed in astrocytes. Ligand binding experiments [17] and measurement of mRNA expression levels [16] showed that ETB receptors are highly expressed in astrocytes over ETA receptors. Activation of the astrocytic ETB receptor causes increased intracellular Ca2+ and activation of protein kinase C/extracellular signal-regulated kinase signals via Gq-type proteins [17,64,67,68], which are intracellular signals involved in ET-induced astrocytic proliferation [64,69,70] and the production of some bioactive substances [71,72,73]. The astrocytic ETB receptor was shown to be coupled with Gi-type proteins. The astrocytic ETB-receptor-mediated Gi signal was reported to be involved in ET-induced reduction in intercellular communication through gap junctions [74]. Rho protein-mediated signals can be activated by astrocytic ETB receptors [63]. Activation of the ETB-receptor-mediated Rho signal in astrocytes involves cytoskeletal reorganization [63,75] and cell-adhesion-dependent proliferation [64,76]. As astrocytic proliferation and morphological alteration occur with phenotype conversion to reactive astrocytes [3,4], these signal mechanisms triggered by ETB receptors are thought to underlie the induction of reactive astrocytes.
2.3. Endothelin Agonists and Antagonists
Due to the potent vasoconstricting action of ET-1, studies on ET receptor agonists and antagonists have been directed toward application in medications for cardiovascular diseases [17] (Table 1). BQ123 [77] and FR139317 [78] are peptide-based ETA-selective antagonists that are generated shortly after the discovery of ET-1. However, these ETA antagonists are unsuitable for oral administration because of the low gastrointestinal absorption due to the peptide structure and have not been clinically applied. Conversely, bosentan (ETA/ETB antagonist) [79], ambrisentan (ETA selective antagonist) [80], and macisentan (ETA/ETB antagonist) [81], which are orally administered, are approved in many countries for the treatment of pulmonary arterial hypertension. As an ET receptor agonist, IRL-1620, which shows high selectivity for the ETB receptor, was developed [82]. IRL-1620 was reported to ameliorate ischemic brain injury and is expected to be a novel neuroprotective drug [83,84]. BQ788 [85], A192621 [86], and IRL-2500 [87] are selective ETB receptor antagonists. Nagiri et al. suggested that IRL-2500 acts as an inverse agonist by analyzing the structure of the IRL-2500/ETB receptor complex [88]. However, to date, only a few studies have attempted to examine the roles of ETB receptors in the treatment of diseases.
3. ET System in the Brain
Increases in brain ET-1 have been observed in animal models of nerve injury [18,19,24] and in patients with stroke, head trauma, and neurodegenerative diseases [20,21,22,23,25]. Immunohistochemical observations of a damaged brain have shown that ET-1 is produced by brain microvascular endothelial cells and reactive astrocytes. Factors such as tumor necrosis factor-α, interleukin-1β and thrombin, as well as hypoxia induce ET-1 production in brain microvascular endothelial cells [89] and astrocytes [90,91,92]. ET-1 stimulates astrocytic ET-1 production. We found that the stimulation of ETB receptors in mouse cultured astrocytes increased prepro-ET-1 mRNA levels [24]. Furthermore, prepro-ET-1 expression in the mouse TBI model was reduced by the antagonism of ETB receptors [24]. These results indicate that astrocytic ET-1 production in brain disorders is enhanced by an autocrine mechanism using ET-1.
Both ETA and ETB receptors are present in the brain with different cellular distributions. Brain ETA receptors are expressed in vascular smooth muscle. The activation of brain ETA receptors causes vasospasm in several brain disorders, which aggravates ischemic brain damage [93,94]. In the brain, ETB receptors are highly expressed, especially in astrocytes. Astrocytic ETB receptors are upregulated and accompanied by the conversion to reactive astrocytes [41,95,96,97]. Together with an increase in ET-1, the upregulation of astrocytic ETB receptors suggests that ETB-mediated regulation of astrocytic function becomes more pronounced in brain disorders.
4. ETB-receptors-mediated Astrocytic Activation
Phenotypic conversion of resting astrocytes to reactive astrocytes is commonly observed in various brain disorders. Since reactive astrocytes affect brain damage and/or the recovery of the damaged nervous system, mechanisms to induce reactive phenotype conversion have been investigated. Administration of ETB agonists into the rat brain increased the number of GFAP-positive hypertrophic astrocytes without neuronal degeneration or microglial activation [39]. In animal models of stab wound injury [40] and brain ischemia [41], the induction of activated astrocytes was reduced by administration of the ETB antagonist BQ788. With the conversion of the activated phenotype, astrocytes become hypertrophic and proliferative [3,4]. Stimulation of ETB receptors in cultured astrocytes causes morphological alterations accompanied by cytoskeletal actin reorganization [63,98] and stimulated cellular proliferation [64,69,70], which is consistent with the action of ETB receptors in vivo. These findings indicate that the activation of astrocytic ETB receptors promotes their conversion to activated astrocytes. The conversion to reactive astrocytes is induced by several signaling factors in the damaged nerve tissues [3]. Among these factors, the ETB-receptor-mediated mechanism is characterized by maintaining or enhancing the activated state of astrocytes through the autocrine mechanism of ET-1, suggesting a pivotal role for ETB-mediated induction of reactive astrocytes.
Stat3, a transcription factor, is activated in response to brain injury, where reactive astrocytes have high levels of activated Stat3 [99,100,101]. Increased production of GFAP, which underlies the hypertrophy of activated astrocytes, is stimulated by gene transcription through Stat3. In addition, cyclin D1 and S-phase-regulated kinase-2, which are proteins that promote cell cycle G1/S phase transition and are upregulated in activated astrocytes [102,103,104], have a Stat3 binding site on their gene promoter. Studies have shown that the inhibition of Stat3 prevents the induction and proliferation of reactive astrocytes in animal models of brain injury [105,106]. We showed that the activation of ETB receptors in cultured astrocytes activated Stat3 and stimulated the transcription of cyclin D1 and Skp2 [69]. Administration of BQ788 reduced the activated form of Stat3 in a mouse model of TBI [69]. This result indicates that the generation of activated astrocytes by ETB receptor stimulation is Stat3-mediated. LeComte et al. showed that the transcription of ETB receptors was promoted by Stat3, suggesting that this mechanism underlies the upregulation of astrocytic ETB receptors in brain ischemia [95]. Thus, it can be concluded that ETB-receptor-mediated Stat3 activation is an intracellular signal involved not only in astrocyte phenotype conversion but also in the positive feedback mechanism due to ETB receptor upregulation (Figure 2).
5. Roles of Astrocytic ETB Receptors in Brain Disorders
As reactive astrocytes are involved in the pathogenesis of many brain disorders, examinations have been conducted to improve brain disorders by controlling the astrocytic functions. Studies using animal models of brain disorders suggest that ETB-receptor-mediated alterations of astrocytic functions have a beneficial effect on AD, brain ischemia, neuropathic pain, and TBI.
5.1. Alzheimer’s Disease
Reactive astrocytes are observed in the brains of patients with AD and contribute to the pathology of AD [14]. In AD, the accumulation of amyloid-β proteins in the brain is the cause of nerve cell degeneration and cognitive impairment. In addition, ischemic neurodegeneration associated with decreased cerebral blood flow is involved in the progression of cognitive injury due to AD. In the brains of AD patients, ET-1 content and ECE-2 expression were increased [26]. Hung et al. showed that astrocyte-specific overexpression of ET-1 promoted amyloid-β production [107]. Conversely, in cultured neurons, amyloid-β proteins increase ECE expression and ET-1 production [108]. These observations suggest that ET-1 and amyloid-β proteins promote their expression and aggravate AD pathology. It has been suggested that ECE also cleaves amyloid-β proteins and removes them from the brain [109]. Therefore, further investigation into the pathological significance of increased ECE expression in AD is necessary. It was shown that the dysfunction of brain blood vessels and reduction in cerebral blood flow are observed in the brains of patients with AD. Since reduction in cerebral blood flow correlates with the degree of cognitive symptoms, it is considered a factor aggravating AD pathology. In addition to direct action on neuronal cells, the accumulation of amyloid-β proteins causes cerebrovascular injury and reduction in cerebral blood flow. Elesber et al. showed that the non-selective ET receptor antagonist bosentan improves cerebrovascular dysfunction in AD model mice overexpressing amyloid precursor protein [110]. In the pathological brain, ET-1 production occurs in activated astrocytes, which causes a decrease in cerebral blood flow. Therefore, suppression of astrocyte activation in the AD brain is expected to produce beneficial effects by improving cerebral blood flow and by reducing amyloid-β production.
5.2. Brain Ischemia
Due to its potent vasoconstricting action, the role of brain ET-1 was examined in relation to the vasospasm observed during cerebral ischemia. Since ETA receptors mediate the vasoconstricting action of ET-1, the effects of ETA antagonists on nerve damage in animal models of brain ischemia have been examined [93,94]. Although these animal experiments indicate the protective effect of ETA antagonists, these drugs have not been clinically applied for cerebral ischemic injury at the time of writing. In contrast, Leonard et al. reported that the ETB receptor agonist IRL-1620 had a neuroprotective effect and promoted the recovery of nerve function in a rat brain ischemia model using middle cerebral artery occlusion [84]. IRL-1620 administration was accompanied by angiogenesis and neurogenesis, which in turn were accompanied by increased vascular endothelial growth factor (VEGF) and nerve growth factor (NGF) production [83]. Activation of astrocytic ETB receptors stimulates the production of several growth factors to induce angiogenesis and neurogenesis, including brain-derived neurotrophic factor (BDNF), glial cell line-derived neurotrophic factor (GDNF), VEGF, and NGF (Figure 3) [72,111,112,113]. Although it was not examined if the protective action of IRL-1620 against cerebral ischemia is caused by astrocyte-derived growth factors, these observations suggest that the activation of astrocytic ETB receptors is beneficial in promoting the recovery of nerve functions after brain ischemia.
5.3. Neuropathic Pain
ET-1 is known to modulate nociceptive pain by affecting neurotransmission in the ascending pain pathway [114]. In the spinal cord, ET-1 induces hyperalgesia via the activation of ETA receptors. ET-1 also has an anti-nociceptive effect, which was reported to be mediated through ETB receptors [114]. The modulation of transmission in the pain pathway by ET-1 is mediated by the regulation of ion channel activity and neurotransmitter release in the spinal cord and brain. Neuropathic pain is an intractable pain that occurs without tissue damage.
The induction of neuropathic pain is caused by hyperexcitation of neural networks in the spinal pain pathway. Although the mechanisms underlying hyperexcitation in the pain pathway have not been fully clarified, previous studies showed the involvement of reactive astrocytes in the spinal cord. Increased numbers of reactive astrocytes were observed in the spinal cord of experimental animals with neuropathic pain [115]. Tsuda et al. showed that administration of a Stat3 inhibitor to a rat neuropathic pain model resulted in the suppression of the development of reactive astrocytes in the spinal cord and recovery from the established hyperalgesia [116]. Yamasaki et al. found that, in neuropathic pain induced by allergic inflammation, astrocytic ETB receptors were upregulated with the induction of reactive astrocytes in the spinal cord [117]. The suppression of reactive astrocytes by BQ788 also reduced hyperalgesia [117]. These observations suggest that the ETB-receptor-mediated induction of reactive astrocytes causes neuropathic pain.
5.4. Traumatic Brain Injury (TBI)
TBI is often caused by a physical blow to the brain in a traffic accident or a fall. In the acute phase of TBI, disruption of the blood–brain barrier (BBB) occurs around the impact core region. Disruption of the BBB allows entry of blood components and blood cells into the brain parenchyma, which causes brain edema and neuroinflammation that aggravates brain damage due to TBI. Therefore, BBB protection in the acute phase is critical for the recovery of patients with TBI. A large part of the BBB is formed between the brain microvascular endothelial cells and astrocytes. The permeability of brain microvessels is not static and is not regulated by various factors produced by astrocytes (Figure 3) [118]. In the acute phase of TBI, reactive astrocytes release excessive amounts of vascular permeability regulators, such as VEGF and matrix metalloproteinases (MMPs), to brain microvessels and disrupt the barrier function [13]. In a mouse fluid percussion injury (FPI)-induced TBI model, we found that BQ788 ameliorated the disruption of BBB and brain edema accompanied by a decrease in reactive astrocytes [36]. Amelioration of BBB disruption was also achieved by the administration of bosentan (ETA/ETB antagonist) after FPI to the mouse brain [24]. These findings indicate that ETB antagonism is effective in reducing TBI-induced BBB disruption. Activation of astrocytic ETB receptors stimulates VEGF and MMP9 production, while decreasing angiopoietin-1, a factor that stabilizes the BBB (Table 2) [119,120]. In the mouse TBI model, BQ788 normalized the altered expressions of VEGF, MMP9, and angiopoietin-1 in the injured region [42,121]. The actions of these vascular permeability regulators are thought to underlie protection of the BBB by ETB receptor antagonism. BBB disruption by TBI involves several factors that are produced by reactive astrocytes. The antagonism of ETB receptors simultaneously improves the production of these factors by reducing astrocytic activation, which suggests that ETB antagonists effectively protect BBB function in the acute phase of TBI.
6. Conclusions
As the pathological roles of astrocytes have been clarified, therapeutic strategies targeting reactive astrocytes are being studied in several brain disorders. Astrocytic activation is triggered by the release of several signaling molecules from damaged nerve tissues (Table 2). Among these factors, the ET-1/ETB receptor system maintains and enhances the activated state of astrocytes through the autocrine pathway of ET-1 and upregulation of ETB receptor expression. This positive feedback mechanism indicates a pivotal role of the ET-1/ETB receptor system in the functional regulation of reactive astrocytes. In addition, many ET receptor agonists and antagonists have been developed, some of which are clinically applied. In the future, ET receptor agonists and antagonists are expected to be clinically applied in the treatment of several brain disorders.
Funding
This work was supported by a Grant-in-Aid for Scientific Research (C) (grant number: 18K06695) from the Japan Society for the Promotion of Science (JSPS).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol. Rev. 2018, 98, 239–389. [Google Scholar] [CrossRef]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 2009, 2, 638–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liddelow, S.A.; Barres, B.A. Reactive astrocytes: Production, function, and therapeutic potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hostenbach, S.; Cambron, M.; D’haeseleer, M.; Kooijman, R.; De Keyser, J. Astrocyte loss and astrogliosis in neuroinflammatory disorders. Neurosci. Lett. 2014, 565, 39–41. [Google Scholar] [CrossRef]
- Colombo, E.; Farina, C. Astrocytes: Key regulators of neuroinflammation. Trends Immunol. 2016, 37, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Cassé, F.; Richetin, K.; Toni, N. Astrocytes’ contribution to adult neurogenesis in physiology and Alzheimer’s disease. Front. Cell Neurosci. 2018, 12, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lium, Z.; Chopp, M. Astrocytes, therapeutic targets for neuroprotection and neurorestoration in ischemic stroke. Prog. Neurobiol. 2016, 144, 103–120. [Google Scholar]
- Magnusson, J.P.; Göritz, C.; Tatarishvili, J.; Dias, D.O.; Smith, E.M.; Lindvall, O.; Kokaia, Z.; Frisén, J. A latent neurogenic program in astrocytes regulated by Notch signaling in the mouse. Science 2014, 346, 237–241. [Google Scholar] [CrossRef]
- Lo, F.S.; Zhao, S.; Erzurumlu, R.S. Astrocytes promote peripheral nerve injury-induced reactive synaptogenesis in the neonatal CNS. J. Neurophysiol. 2011, 106, 2876–2887. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Li, J.; Zheng, J.; Qin, S. Reactive astrocytes in neurodegenerative diseases. Aging Dis. 2019, 10, 664–675. [Google Scholar] [CrossRef] [Green Version]
- Izrael, M.; Slutsky, S.G.; Revel, M. Rising stars: Astrocytes as a therapeutic target for ALS disease. Front. Neurosci. 2020, 14, 824. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Shao, A.; Yao, Y.; Tu, S.; Deng, Y.; Zhang, J. Dual roles of astrocytes in plasticity and reconstruction after traumatic brain injury. Cell Commun. Signal. 2020, 18, 62. [Google Scholar] [CrossRef] [Green Version]
- Acosta, C.; Anderson, H.D.; Anderson, C.M. Astrocyte dysfunction in Alzheimer disease. J. Neurosci. Res. 2017, 95, 2430–2447. [Google Scholar] [CrossRef] [PubMed]
- Pekny, M.; Wilhelmsson, U.; Tatlisumak, T.; Pekna, M. Astrocyte activation and reactive gliosis-A new target in stroke? Neurosci. Lett. 2019, 689, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, M.; Kurihara, H.; Kimura, S.; Tomobe, Y.; Kobayashi, M.; Mitsui, Y.; Yazaki, Y.; Goto, K.; Masaki, T. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 1988, 332, 411–415. [Google Scholar] [CrossRef] [Green Version]
- Davenport, A.P.; Hyndman, K.A.; Dhaun, N.; Southan, C.; Kohan, D.E.; Pollock, J.S.; Pollock, D.M.; Webb, D.J.; Maguire, J.J. Endothelin. Pharmacol. Rev. 2016, 68, 357–418. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, Y.; Mihara, S.; Ninomiya, M.; Fujimoto, M. Protective effect of endothelin type A receptor antagonist on brain edema and injury after transient middle cerebral artery occlusion in rats. Stroke 2001, 32, 2143–2148. [Google Scholar] [CrossRef] [PubMed]
- Barone, F.C.; Globus, M.Y.; Price, W.J.; White, R.F.; Storer, B.L.; Feuerstein, G.Z.; Busto, R.; Ohlstein, E.H. Endothelin levels increase in rat focal and global ischemia. J. Cereb. Blood Flow Metab. 1994, 14, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Lampl, Y.; Fleminger, G.; Gilad, R.; Galron, R.; Sarova-Pinhas, I.; Sokolovsky, M. Endothelin in cerebrospinal fluid and plasma of patients in the early stage of ischemic stroke. Stroke 1997, 28, 1951–1955. [Google Scholar] [CrossRef] [PubMed]
- Ziv, I.; Fleminger, G.; Djaldetti, R.; Achiron, A.; Melamed, E.; Sokolovsky, M. Increased plasma endothelin-1 in acute ischemic stroke. Stroke 1992, 23, 1014–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, H.; Ide, H.; Ishii, H.; Kabuto, M.; Handa, Y.; Kubota, T. Endothelin-1 levels in plasma and cerebrospinal fluid following subarachnoid haemorrhage. J. Clin. Neurosci. 1995, 2, 252–256. [Google Scholar] [CrossRef]
- Suzuki, R.; Masaoka, H.; Hirata, Y.; Marumo, F.; Isotani, E.; Hirakawa, K. The role of endotheln-1 in the origin of cerebral vasospasm in patients with aneurysmal subarachnoid hemorrhage. J. Neurosurg. 1992, 77, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Michinaga, S.; Inoue, A.; Yamamoto, H.; Ryu, R.; Inoue, A.; Mizuguchi, H.; Koyama, Y. Endothelin receptor antagonists al-leviate blood-brain barrier disruption and cerebral edema in a mouse model of traumatic brain injury: A comparison between bosentan and ambrisentan. Neuropharmacology 2020, 175, 108182. [Google Scholar] [CrossRef] [PubMed]
- Maier, B.; Lehnert, M.; Laurer, H.L.; Marzi, I. Biphasic elevation in cerebrospinal fluid and plasma concentrations of endothelin 1 after traumatic brain injury in human patients. Shock 2007, 27, 610–614. [Google Scholar] [CrossRef]
- Palmer, J.C.; Barker, R.; Kehoe, P.G.; Love, S. Endothelin-1 is elevated in Alzheimer’s disease and upregulated by amyloid-beta. J. Alzheimers Dis. 2012, 29, 853–861. [Google Scholar] [CrossRef] [Green Version]
- Ranno, E.; D’Antoni, S.; Spatuzza, M.; Berretta, A.; Laureanti, F.; Bonaccorso, C.M.; Pellitteri, R.; Longone, P.; Spalloni, A.; Iyer, A.M.; et al. Endothelin-1 is over-expressed in amyotrophic lateral sclerosis and induces motor neuron cell death. Neurobiol. Dis. 2014, 65, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Haufschild, T.; Shaw, S.G.; Kesselring, J.; Flammer, J. Increased endothelin-1 plasma levels in patients with multiple sclerosis. J. Neuroophthalmol. 2001, 21, 37–38. [Google Scholar] [CrossRef] [Green Version]
- Rolinski, B.; Heigermoser, A.; Lederer, E.; Bogner, J.R.; Loch, O.; Goebel, F.D. Endothelin-1 is elevated in the cerebrospinal fluid of HIV-infected patients with encephalopathy. Infection 1999, 27, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Hahn, S.; Gartner, S.; Pardo, C.A.; Netesan, S.K.; McArthur, J.; Nath, A. Molecular programming of endothelin-1 in HIV-infected brain: Role of Tat in up-regulation of ET-1 and its inhibition by statins. FASEB J. 2007, 21, 777–789. [Google Scholar] [CrossRef] [Green Version]
- D’Orléans-Juste, P.; Akide Ndunge, O.B.; Desbiens, L.; Tanowitz, H.B.; Desruisseaux, M.S. Endothelins in inflammatory neurological diseases. Pharmacol. Ther. 2019, 94, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Armstead, W.M. Endothelins and the role of endothelin antagonists in the management of posttraumatic vasospasm. Curr. Pharm. Des. 2004, 10, 2185–2192. [Google Scholar] [CrossRef] [PubMed]
- Milan, J.; Charalambous, C.; Elhag, R.; Chen, T.C.; Li, W.; Guan, S.; Hofman, F.M.; Zidovetzki, R. Multiple signaling pathways are involved in endothelin-1-induced brain endothelial cell migration. Am. J. Physiol. Cell Physiol. 2006, 291, C155–C164. [Google Scholar] [CrossRef] [Green Version]
- Briyal, S.; Nguyen, C.; Leonard, M.; Gulati, A. Stimulation of endothelin B receptors by IRL-1620 decreases the progression of Alzheimer’s disease. Neuroscience 2015, 301, 1–11. [Google Scholar] [CrossRef]
- Adams, K.L.; Riparini, G.; Banerjee, P.; Breur, M.; Bugiani, M.; Gallo, V. Endothelin-1 signaling maintains glial progenitor proliferation in the postnatal subventricular zone. Nat. Commun. 2020, 11, 2138. [Google Scholar] [CrossRef]
- Nishikawa, K.; Ayukawa, K.; Hara, Y.; Wada, K.; Aoki, S. Endothelin/endothelin-B receptor signals regulate ventricle-directed interkinetic nuclear migration of cerebral cortical neural progenitors. Neurochem. Int. 2011, 58, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Ukita, A.; Abe, K.; Iwamae, K.; Tokuyama, S.; Tanaka, K.; Kotake, Y. Dexamethasone downregulates endothelin receptors and reduces endothelin-induced production of matrix metalloproteinases in cultured rat astrocytes. Mol. Pharmacol. 2017, 92, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Lazarini, F.; Strosberg, A.D.; Couraud, P.O.; Cazaubon, S.M. Coupling of ETB endothelin receptor to mitogen-activated protein kinase stimulation and DNA synthesis in primary cultures of rat astrocytes. J. Neurochem. 1996, 66, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, N.; Takemura, M.; Koyama, Y.; Shigenaga, Y.; Okada, T.; Baba, A. Endothelins promote the activation of astrocytes in rat neostriatum through ETB receptors. Eur. J. Neurosci. 1997, 9, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Takemura, M.; Fujiki, K.; Ishikawa, N.; Shigenaga, Y.; Baba, A. BQ788, an endothelin ETB receptor antagonist attenuates stab wound injury-induced reactive astrocytes in rat brain. Glia 1999, 26, 268–271. [Google Scholar] [CrossRef]
- Gadea, A.; Schinelli, S.; Gallo, V. Endothelin-1 regulates astrocyte proliferation and reactive gliosis via a JNK/c-Jun signaling pathway. J. Neurosci. 2008, 28, 2394–2408. [Google Scholar] [CrossRef]
- Michinaga, S.; Kimura, A.; Hatanaka, S.; Minami, S.; Asano, A.; Ikushima, Y.; Matsui, S.; Toriyama, Y.; Fujii, M.; Koyama, Y. Delayed administration of BQ788, an ETB antagonist, after experimental traumatic brain Injury promotes recovery of blood-brain barrier function and a reduction of cerebral edema in mice. J. Neurotrauma. 2018, 35, 1481–1494. [Google Scholar] [CrossRef]
- Saida, K.; Mitsui, Y.; Ishida, N. A novel peptide, vasoactive intestinal contractor, of a new (endothelin) peptide family. Molecular cloning, expression, and biological activity. J. Biol. Chem. 1989, 264, 14613–14616. [Google Scholar] [CrossRef]
- Masuo, Y.; Ishikawa, Y.; Kozakai, T.; Uchide, T.; Komatsu, Y.; Saida, K. Vasoactive intestinal contractor/endothelin-2 gene expression in the murine central nervous system. Biochem. Biophys. Res. Commun. 2003, 300, 661–668. [Google Scholar] [CrossRef]
- Uchide, T.; Masuda, H.; Lee, Y.S.; Makiyama, Y.; Mitsui, Y.; Saida, K. Fluctuating gene expression and localized cellular distribution of vasoactive intestinal contractor (VIC) in mouse uterus. J. Histochem. Cytochem. 2000, 48, 699–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, I.; Bramall, A.N.; Baynash, A.G.; Rattner, A.; Rakheja, D.; Post, M.; Joza, S.; McKerlie, C.; Stewart, D.J.; McInnes, R.R.; et al. Endothelin-2 deficiency causes growth retardation, hypothermia, and emphysema in mice. J. Clin. Investig. 2013, 123, 2643–2653. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, H.; Suzuki, N.; Onda, H.; Fujino, M. Abundance of endothelin-3 in rat intestine, pituitary gland and brain. Biochem. Biophys. Res. Commun. 1989, 164, 74–80. [Google Scholar] [CrossRef]
- Baynash, A.G.; Hosoda, K.; Giaid, A.; Richardson, J.A.; Emoto, N.; Hammer, R.E.; Yanagisawa, M. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell 1994, 79, 1277–1285. [Google Scholar] [CrossRef]
- Hofstra, R.M.; Osinga, J.; Tan-Sindhunata, G.; Wu, Y.; Kamsteeg, E.J.; Stulp, R.P.; van Ravenswaaij-Arts, C.; Majoor-Krakauer, D.; Angrist, M.; Chakravarti, A.; et al. A homozygous mutation in the endothelin-3 gene associated with a combined Waardenburg type 2 and Hirschsprung phenotype (Shah-Waardenburg syndrome). Nat. Genet. 1996, 12, 445–447. [Google Scholar] [CrossRef]
- Stow, L.R.; Jacobs, M.E.; Wingo, C.S.; Cain, B.D. Endothelin-1 gene regulation. FASEB J. 2011, 25, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Discher, D.J.; Bishopric, N.H.; Webster, K.A. Hypoxia regulates expression of the endothelin-1 gene through a proximal hypoxia-inducible factor-1 binding site on the antisense strand. Biochem. Biophys. Res. Commun. 1998, 245, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Eckman, E.A.; Reed, D.K.; Eckman, C.B. Degradation of the Alzheimer’s amyloid beta peptide by endothelin-converting enzyme. J. Biol. Chem. 2001, 276, 24540–24548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, J.C.; Kehoe, P.G.; Love, S. Endothelin-converting enzyme-1 in Alzheimer’s disease and vascular dementia. Neuropathol. Appl. Neurobiol. 2010, 36, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.; Love, S. Endothelin receptor antagonists: Potential in Alzheimer’s disease. Pharmacol. Res. 2011, 63, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Kawanabe, Y.; Okamoto, Y.; Miwa, S.; Hashimoto, N.; Masaki, T. Molecular mechanisms for the activation of voltage-independent Ca2+ channels by endothelin-1 in chinese hamster ovary cells stably expressing human endothelin-A receptors. Mol. Pharmacol. 2002, 62, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Kawanabe, Y.; Nozaki, K.; Hashimoto, N.; Masaki, T. Characterization of Ca2+ channels and G proteins involved in arachidonic acid release by endothelin-1/endothelin-A receptor. Mol. Pharmacol. 2003, 64, 689–695. [Google Scholar] [CrossRef] [Green Version]
- Jouneaux, C.; Mallat, A.; Serradeil-Le Gal, C.; Goldsmith, P.; Hanoune, J.; Lotersztajn, S. Coupling of endothelin B receptors to the calcium pump and phospholipase C via Gs and Gq in rat liver. J. Biol. Chem. 1994, 269, 1845–1851. [Google Scholar] [CrossRef]
- Eguchi, S.; Hirata, Y.; Imai, T.; Marumo, F. Endothelin receptor subtypes are coupled to adenylate cyclase via different guanyl nucleotide-binding proteins in vasculature. Endocrinology 1993, 132, 524–529. [Google Scholar] [CrossRef]
- Rauh, A.; Windischhofer, W.; Kovacevic, A.; DeVaney, T.; Huber, E.; Semlitsch, M.; Leis, H.J.; Sattler, W.; Malle, E. Endothelin (ET)-1 and ET-3 promote expression of c-fos and c-jun in human choriocarcinoma via ETB receptor-mediated Gi- and Gq-pathways and MAP kinase activation. Br. J. Pharmacol. 2008, 154, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Horinouchi, T.; Asano, H.; Higa, T.; Nishimoto, A.; Nishiya, T.; Muramatsu, I.; Miwa, S. Differential coupling of human endothelin type A receptor to Gq/11 and G12 proteins: The functional significance of receptor expression level in generating multiple receptor signaling. J. Pharmacol. Sci. 2009, 111, 338–351. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, S.; Ridley, A.J.; Lutz, S. The function of Rho-associated kinases ROCK1 and ROCK2 in the pathogenesis of cardiovascular disease. Front. Pharmacol. 2015, 6, 276. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, H.; Rashid, M. Development of Rho-kinase inhibitors for cardiovascular medicine. Trends Pharmacol. Sci. 2007, 28, 296–302. [Google Scholar] [CrossRef]
- Koyama, Y.; Baba, A. Endothelin-induced cytoskeletal actin re-organization in cultured astrocytes: Inhibition by C3 ADP-ribosyltransferase. Glia 1996, 16, 342–350. [Google Scholar] [CrossRef]
- Koyama, Y.; Yoshioka, Y.; Shinde, M.; Matsuda, T.; Baba, A. Focal adhesion kinase mediates endothelin-induced cyclin D3 expression in rat cultured astrocytes. J. Neurochem. 2004, 90, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Evans, N.J.; Walker, J.W. Endothelin receptor dimers evaluated by FRET, ligand binding, and calcium mobilization. Biophys. J. 2008, 95, 483–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, N.J.; Walker, J.W. Sustained Ca2+ signaling and delayed internalization associated with endothelin receptor heterodimers linked through a PDZ finger. Can. J. Physiol. Pharmacol. 2008, 86, 526–535. [Google Scholar] [CrossRef]
- Stanimirovic, D.B.; Ball, R.; Mealing, G.; Morley, P.; Durkin, J.P. The role of intracellular calcium and protein kinase C in endothelin-stimulated proliferation of rat type I astrocytes. Glia 1995, 15, 119–130. [Google Scholar] [CrossRef]
- Holzwarth, J.A.; Glaum, S.R.; Miller, R.J. Activation of endothelin receptors by sarafotoxin regulates Ca2+ homeostasis in cerebellar astrocytes. Glia 1992, 5, 239–250. [Google Scholar] [CrossRef]
- Koyama, Y.; Sumie, S.; Nakano, Y.; Nagao, T.; Tokumaru, S.; Michinaga, S. Endothelin-1 stimulates expression of cyclin D1 and S-phase kinase-associated protein 2 by activating the transcription factor STAT3 in cultured rat astrocytes. J. Biol. Chem. 2019, 294, 3920–3933. [Google Scholar] [CrossRef]
- Tabernero, A.; Sánchez-Alvarez, R.; Medina, J.M. Increased levels of cyclins D1 and D3 after inhibition of gap junctional communication in astrocytes. J. Neurochem. 2006, 96, 973–982. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Prasanna, G.; Yorio, T. Endothelin-1-mediated signaling in the expression of matrix metalloproteinases and tissue inhibitors of metalloproteinases in astrocytes. Investig. Ophthalmol. Vis. Sci. 2007, 48, 3737–3745. [Google Scholar] [CrossRef] [Green Version]
- Koyama, Y.; Tsujikawa, K.; Matsuda, T.; Baba, A. Endothelin-1 stimulates glial cell line-derived neurotrophic factor expression in cultured rat astrocytes. Biochem. Biophys. Res. Commun. 2003, 303, 1101–1105. [Google Scholar] [CrossRef]
- Koyama, Y.; Tanaka, K. Endothelins stimulate the production of stromelysin-1 in cultured rat astrocytes. Biochem. Biophys. Res. Commun. 2008, 371, 659–663. [Google Scholar] [CrossRef]
- Tencé, M.; Ezan, P.; Amigou, E.; Giaume, C. Increased interaction of connexin43 with zonula occludens-1 during inhibition of gap junctions by G protein-coupled receptor agonists. Cell Signal. 2012, 24, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Hama, H.; Sakurai, T.; Kasuya, Y.; Fujiki, M.; Masaki, T.; Goto, K. Action of endothelin-1 on rat astrocytes through the ETB receptor. Biochem. Biophys. Res. Commun. 1992, 186, 355–362. [Google Scholar] [CrossRef]
- Cazaubon, S.; Chaverot, N.; Romero, I.A.; Girault, J.A.; Adamson, P.; Strosberg, A.D.; Couraud, P.O. Growth factor activity of endothelin-1 in primary astrocytes mediated by adhesion-dependent and -independent pathways. J. Neurosci. 1997, 17, 6203–6212. [Google Scholar] [CrossRef] [Green Version]
- Ihara, M.; Noguchi, K.; Saeki, T.; Fukuroda, T.; Tsuchida, S.; Kimura, S.; Fukami, T.; Ishikawa, K.; Nishikibe, M.; Yano, M. Biological profiles of highly potent novel endothelin antagonists selective for the ETA receptor. Life Sci. 1992, 50, 247–255. [Google Scholar] [CrossRef]
- Sogabe, K.; Nirei, H.; Shoubo, M.; Nomoto, A.; Ao, S.; Notsu, Y.; Ono, T. Pharmacological profile of FR139317, a novel, potent endothelin ETA receptor antagonist. J. Pharmacol. Exp. Ther. 1993, 264, 1040–1046. [Google Scholar]
- Clozel, M.; Breu, V.; Gray, G.A.; Kalina, B.; Löffler, B.M.; Burri, K.; Cassal, J.M.; Hirth, G.; Müller, M.; Neidhart, W. Pharmacological characterization of bosentan, a new potent orally active nonpeptide endothelin receptor antagonist. J. Pharmacol. Exp. Ther. 1994, 270, 228–235. [Google Scholar]
- Vatter, H.; Seifert, V. Ambrisentan, a non-peptide endothelin receptor antagonist. Cardiovasc. Drug Rev. 2006, 24, 63–76. [Google Scholar] [CrossRef]
- Bolli, M.H.; Boss, C.; Binkert, C.; Buchmann, S.; Bur, D.; Hess, P.; Iglarz, M.; Meyer, S.; Rein, J.; Rey, M.; et al. The discovery of N-[5-(4-bromophenyl)-6-[2-[(5-bromo-2-pyrimidinyl)oxy]ethoxy]-4-pyrimidinyl]-N′-p ropylsulfamide (Macitentan), an orally active, potent dual endothelin receptor antagonist. J. Med. Chem. 2012, 55, 7849–7861. [Google Scholar] [CrossRef]
- Takai, M.; Umemura, I.; Yamasaki, K.; Watakabe, T.; Fujitani, Y.; Oda, K.; Urade, Y.; Inui, T.; Yamamura, T.; Okada, T. A potent and specific agonist, Suc-[Glu9,Ala11,15]-endothelin-1(8-21), IRL 1620, for the ETB receptor. Biochem. Biophys. Res. Commun. 1992, 184, 953–959. [Google Scholar] [CrossRef]
- Leonard, M.G.; Gulati, A. Endothelin B receptor agonist, IRL-1620, enhances angiogenesis and neurogenesis following cerebral ischemia in rats. Brain Res. 2013, 1528, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Leonard, M.G.; Briyal, S.; Gulati, A. Endothelin B receptor agonist, IRL-1620, provides long-term neuroprotection in cerebral ischemia in rats. Brain Res. 2012, 1464, 14–23. [Google Scholar] [CrossRef]
- Ishikawa, K.; Ihara, M.; Noguchi, K.; Mase, T.; Mino, N.; Saeki, T.; Fukuroda, T.; Fukami, T.; Ozaki, S.; Nagase, T. Biochemical and pharmacological profile of a potent and selective endothelin B-receptor antagonist, BQ-788. Proc. Natl. Acad. Sci. USA 1994, 91, 4892–4896. [Google Scholar] [CrossRef] [Green Version]
- Von Geldern, T.W.; Tasker, A.S.; Sorensen, B.K.; Winn, M.; Szczepankiewicz, B.G.; Dixon, D.B.; Chiou, W.J.; Wang, L.; Wessale, J.L.; Adler, A.; et al. Pyrrolidine-3-carboxylic acids as endothelin antagonists. 4. Side chain conformational restriction leads to ETB selectivity. J. Med. Chem. 1999, 42, 3668–3678. [Google Scholar] [CrossRef] [PubMed]
- Balwierczak, J.L.; Bruseo, C.W.; DelGrande, D.; Jeng, A.Y.; Savage, P.; Shetty, S.S. Characterization of a potent and selective endothelin-B receptor antagonist, IRL 2500. J. Cardiovasc. Pharmacol. 1995, 26 (Suppl. 3), S393–S396. [Google Scholar] [CrossRef]
- Nagiri, C.; Shihoya, W.; Inoue, A.; Kadji, F.M.N.; Aoki, J.; Nureki, O. Crystal structure of human endothelin ETB receptor in complex with peptide inverse agonist IRL2500. Commun. Biol. 2019, 2, 236. [Google Scholar] [CrossRef]
- Skopál, J.; Turbucz, P.; Vastag, M.; Bori, Z.; Pék, M.; deChâtel, R.; Nagy, Z.; Tóth, M.; Karádi, I. Regulation of endothelin release from human brain microvessel endothelial cells. J. Cardiovasc. Pharmacol. 1998, 31 (Suppl. 1), S370–S372. [Google Scholar] [CrossRef]
- Desai, D.; He, S.; Yorio, T.; Krishnamoorthy, R.R.; Prasanna, G. Hypoxia augments TNF-alpha-mediated endothelin-1 release and cell proliferation in human optic nerve head astrocytes. Biochem. Biophys. Res. Commun. 2004, 318, 642–648. [Google Scholar] [CrossRef]
- Ehrenreich, H.; Costa, T.; Clouse, K.A.; Pluta, R.M.; Ogino, Y.; Coligan, J.E.; Burd, P.R. Thrombin is a regulator of astrocytic endothelin-1. Brain Res. 1993, 600, 201–207. [Google Scholar] [CrossRef]
- Didier, N.; Romero, I.A.; Créminon, C.; Wijkhuisen, A.; Grassi, J.; Mabondzo, A. Secretion of interleukin-1 by astrocytes mediates endothelin-1 and tumor necrosis factor- effects on human brain microvascular endothelial cell permeability. J. Neurochem. 2003, 86, 246–254. [Google Scholar] [CrossRef]
- Sagher, O.; Jin, Y.; Thai, Q.A.; Fergus, A.; Kassell, N.F.; Lee, K.S. Cerebral microvascular responses to endothelins: The role of ETA receptors. Brain Res. 1994, 658, 179–184. [Google Scholar] [CrossRef]
- Yu, J.C.; Pickard, J.D.; Davenport, A.P. Endothelin ETA receptor expression in human cerebrovascular smooth muscle cells. Br. J. Pharmacol. 1995, 116, 2441–2446. [Google Scholar] [CrossRef] [Green Version]
- LeComte, M.D.; Shimada, I.S.; Sherwin, C.; Spees, J.L. Notch1-STAT3-ETBR signaling axis controls reactive astrocyte proliferation after brain injury. Proc. Natl. Acad. Sci. USA 2015, 112, 8726–8731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, C.M.; Rogers, S.D.; Pomonis, J.D.; Egnaczyk, G.F.; Keyser, C.P.; Schmidt, J.A.; Ghilardi, J.R.; Maggio, J.E.; Mantyh, P.W. Endothelin receptor expression in the normal and injured spinal cord: Potential involvement in injury-induced ischemia and gliosis. Exp. Neurol. 2003, 180, 1–13. [Google Scholar] [CrossRef]
- Wilhelmsson, U.; Li, L.; Pekna, M.; Berthold, C.H.; Blom, S.; Eliasson, C.; Renner, O.; Bushong, E.; Ellisman, M.; Morgan, T.E.; et al. Absence of glial fibrillary acidic protein and vimentin prevents hypertrophy of astrocytic processes and improves post-traumatic regeneration. J. Neurosci. 2004, 24, 5016–5021. [Google Scholar] [CrossRef] [Green Version]
- Koyama, Y.; Baba, A. Endothelin-induced protein tyrosine phosphorylation of cultured astrocytes: Its relationship to cytoskeletal actin organization. Glia 1999, 26, 324–332. [Google Scholar] [CrossRef]
- Choi, J.S.; Kim, S.Y.; Cha, J.H.; Choi, Y.S.; Sung, K.W.; Oh, S.T.; Kim, O.N.; Chung, J.W.; Chun, M.H.; Lee, S.B.; et al. Upregulation of gp130 and STAT3 activation in the rat hippocampus following transient forebrain ischemia. Glia 2003, 41, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Justicia, C.; Gabriel, C.; Planas, A.M. Activation of the JAK/STAT pathway following transient focal cerebral ischemia: Signaling through Jak1 and Stat3 in astrocytes. Glia 2000, 30, 253–270. [Google Scholar] [CrossRef] [Green Version]
- Oliva, A.A., Jr.; Kang, Y.; Sanchez-Molano, J.; Furones, C.; Atkins, C.M. STAT3 signaling after traumatic brain injury. J. Neurochem. 2012, 120, 710–720. [Google Scholar] [CrossRef]
- Di Giovanni, S.; Movsesyan, V.; Ahmed, F.; Cernak, I.; Schinelli, S.; Stoica, B.; Faden, A.I. Cell cycle inhibition provides neuroprotection and reduces glial proliferation and scar formation after traumatic brain injury. Proc. Natl. Acad. Sci. USA 2005, 102, 8333–8338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, Y.; Cheng, C.; Chen, Y.; Shi, S.; Qin, J.; Xiao, F.; Zhou, D.; Lu, M.; Lu, Q.; et al. A relationship between p27(kip1) and Skp2 after adult brain injury: Implications for glial proliferation. J. Neurotrauma. 2010, 27, 361–371. [Google Scholar] [CrossRef]
- Wu, J.; Pajoohesh-Ganji, A.; Stoica, B.A.; Dinizo, M.; Guanciale, K.; Faden, A.I. Delayed expression of cell cycle proteins contributes to astroglial scar formation and chronic inflammation after rat spinal cord contusion. J. Neuroinflammation. 2012, 9, 169. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, J.E.; Imura, T.; Song, B.; Qi, J.; Ao, Y.; Nguyen, T.K.; Korsak, R.A.; Takeda, K.; Akira, S.; Sofroniew, M.V. STAT3 is a critical regulator of astrogliosis and scar formation after spinal cord injury. J. Neurosci. 2008, 28, 7231–7243. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Nakamura, M.; Katoh, H.; Miyao, T.; Shimazaki, T.; Ishii, K.; Yamane, J.; Yoshimura, A.; Iwamoto, Y.; Toyama, Y.; et al. Conditional ablation of Stat3 or Socs3 discloses a dual role for reactive astrocytes after spinal cord injury. Nat. Med. 2006, 12, 829–834. [Google Scholar] [CrossRef]
- Hung, V.K.; Yeung, P.K.; Lai, A.K.; Ho, M.C.; Lo, A.C.; Chan, K.C.; Wu, E.X.; Chung, S.S.; Cheung, C.W.; Chung, S.K. Selective astrocytic endothelin-1 overexpression contributes to dementia associated with ischemic stroke by exaggerating astrocyte-derived amyloid secretion. J. Cereb. Blood Flow Metab. 2015, 35, 1687–1696. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.C.; Baig, S.; Kehoe, P.G.; Love, S. Endothelin-converting enzyme-2 is increased in Alzheimer’s disease and up-regulated by A. Am. J. Pathol. 2009, 175, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Pacheco-Quinto, J.; Eckman, E.A. Endothelin-converting enzymes degrade intracellular beta-amyloid produced within the endosomal/lysosomal pathway and autophagosomes. J. Biol. Chem. 2013, 88, 5606–5615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elesber, A.A.; Bonetti, P.O.; Woodrum, J.E.; Zhu, X.Y.; Lerman, L.O.; Younkin, S.G.; Lerman, A. Bosentan preserves endothelial function in mice overexpressing app. Neurobiol. Aging 2006, 27, 446–450. [Google Scholar] [CrossRef]
- Koyama, Y.; Tsujikawa, K.; Matsuda, T.; Baba, A. Endothelin increases expression of exon III- and exon IV-containing brain-derived neurotrophic factor transcripts in cultured astrocytes and rat brain. J. Neurosci. Res. 2005, 80, 809–816. [Google Scholar] [CrossRef]
- Ladenheim, R.G.; Lacroix, I.; Foignant-Chaverot, N.; Strosberg, A.D.; Couraud, P.O. Endothelins stimulate c-fos and nerve growth factor expression in astrocytes and astrocytoma. J. Neurochem. 1993, 60, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Tsujikawa, K.; Matsuda, T.; Baba, A. Intracerebroventricular administration of an endothelin ETB receptor agonist increases expressions of GDNF and BDNF in rat brain. Eur. J. Neurosci. 2003, 18, 887–894. [Google Scholar] [CrossRef]
- Khodorova, A.; Montmayeur, J.P.; Strichartz, G. Endothelin receptors and pain. J. Pain. 2009, 10, 4–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwak, Y.S.; Kang, J.; Unabia, G.C.; Hulsebosch, C.E. Spatial and temporal activation of spinal glial cells: Role of gliopathy in central neuropathic pain following spinal cord injury in rats. Exp. Neurol. 2012, 234, 362–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuda, M.; Kohro, Y.; Yano, T.; Tsujikawa, T.; Kitano, J.; Tozaki-Saitoh, H.; Koyanagi, S.; Ohdo, S.; Ji, R.R.; Salter, M.W.; et al. JAK-STAT3 pathway regulates spinal astrocyte proliferation and neuropathic pain maintenance in rats. Brain 2011, 134 Pt 4, 1127–1139. [Google Scholar] [CrossRef]
- Yamasaki, R.; Fujii, T.; Wang, B.; Masaki, K.; Kido, M.A.; Yoshida, M.; Matsushita, T.; Kira, J.I. Allergic inflammation leads to neuropathic pain via glial cell activation. J. Neurosci. 2016, 36, 11929–11945. [Google Scholar] [CrossRef] [Green Version]
- Michinaga, S.; Koyama, Y. Dual roles of astrocyte-derived factors in regulation of blood-brain barrier function after brain damage. Int. J. Mol. Sci. 2019, 20, 571. [Google Scholar] [CrossRef] [Green Version]
- Koyama, Y.; Maebara, Y.; Hayashi, M.; Nagae, R.; Tokuyama, S.; Michinaga, S. Endothelins reciprocally regulate VEGF-A and angiopoietin-1 production in cultured rat astrocytes: Implications on astrocytic proliferation. Glia 2012, 60, 1954–1963. [Google Scholar] [CrossRef]
- Koyama, Y.; Tanaka, K. Intracerebroventricular administration of an endothelin ETB receptor agonist increases expression of matrix metalloproteinase-2 and -9 in rat brain. J. Pharmacol. Sci. 2010, 114, 433–443. [Google Scholar] [CrossRef] [Green Version]
- Michinaga, S.; Tanabe, A.; Nakaya, R.; Fukutome, C.; Inoue, A.; Iwane, A.; Minato, Y.; Tujiuchi, Y.; Miyake, D.; Mizuguchi, H.; et al. Angiopoietin-1/Tie-2 signal after focal traumatic brain injury is potentiated by BQ788, an ETB receptor antagonist, in the mouse cerebrum: Involvement in recovery of blood-brain barrier function. J. Neurochem. 2020, 154, 330–348. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Kotani, M.; Sawamura, T.; Kuribayashi, M.; Konishi, R.; Michinaga, S. Different actions of endothelin-1 on chemokine production in rat cultured astrocytes: Reduction of CX3CL1/fractalkine and an increase in CCL2/MCP-1 and CXCL1/CINC-1. J. Neuroinflamm. 2013, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Michinaga, S.; Inoue, A.; Sonoda, K.; Mizuguchi, H.; Koyama, Y. Down-regulation of astrocytic sonic hedgehog by activation of endothelin ETB receptors: Involvement in traumatic brain injury-induced disruption of blood brain barrier in a mouse model. Neurochem. Int. 2021, 7, 105042. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Tsuboi, S.; Mizogui, F. Endothelin-1 decreases the expression of ephrin-A and B subtypes in cultured rat astrocytes through ETB receptors. Neurosci. Lett. 2021, 741, 135393. [Google Scholar] [CrossRef] [PubMed]
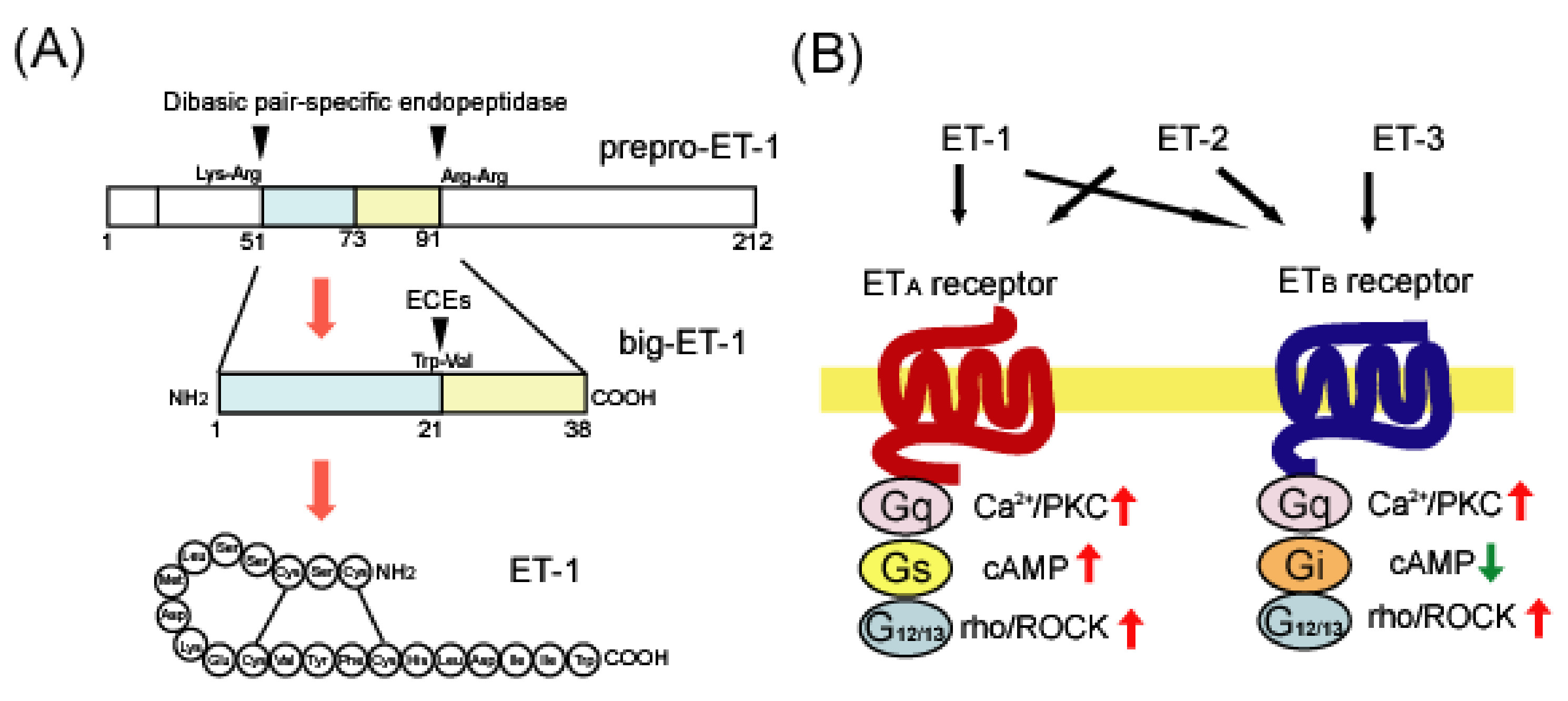
Figure 1.
(A) Biosynthesis of human endothelin-1 (ET-1) from prepro-ET-1. ET-1 is translated as an inactive precursor protein called prepro-ET-1. Prepro-ET-1 is cleaved by dibasic pair-specific endopeptidases and converted to big-ET-1. Specific processing of big-ET-1 by endothelin-converting enzymes (ECEs) results in the production of mature ET-1. (B) Ligand preference and signal transduction of ETA and ETB receptors. There are three distinct ET family peptides: ET-1, ET-2, and ET-3. The ETA receptor has a ligand preference for ET-1 and ET-2, whereas the ETB receptor binds these three ET ligands with a similar affinity. Both ETA and ETB receptors are linked to Gq- and G12/13-type G proteins, which activate Ca2+/protein kinase C (PKC) and rho/Rho-associated protein kinase (ROCK), respectively. ETA receptors are also linked to the Gs protein to trigger cAMP-mediated signals, whereas ETB receptors are linked to Gi to suppress them.
Figure 1.
(A) Biosynthesis of human endothelin-1 (ET-1) from prepro-ET-1. ET-1 is translated as an inactive precursor protein called prepro-ET-1. Prepro-ET-1 is cleaved by dibasic pair-specific endopeptidases and converted to big-ET-1. Specific processing of big-ET-1 by endothelin-converting enzymes (ECEs) results in the production of mature ET-1. (B) Ligand preference and signal transduction of ETA and ETB receptors. There are three distinct ET family peptides: ET-1, ET-2, and ET-3. The ETA receptor has a ligand preference for ET-1 and ET-2, whereas the ETB receptor binds these three ET ligands with a similar affinity. Both ETA and ETB receptors are linked to Gq- and G12/13-type G proteins, which activate Ca2+/protein kinase C (PKC) and rho/Rho-associated protein kinase (ROCK), respectively. ETA receptors are also linked to the Gs protein to trigger cAMP-mediated signals, whereas ETB receptors are linked to Gi to suppress them.

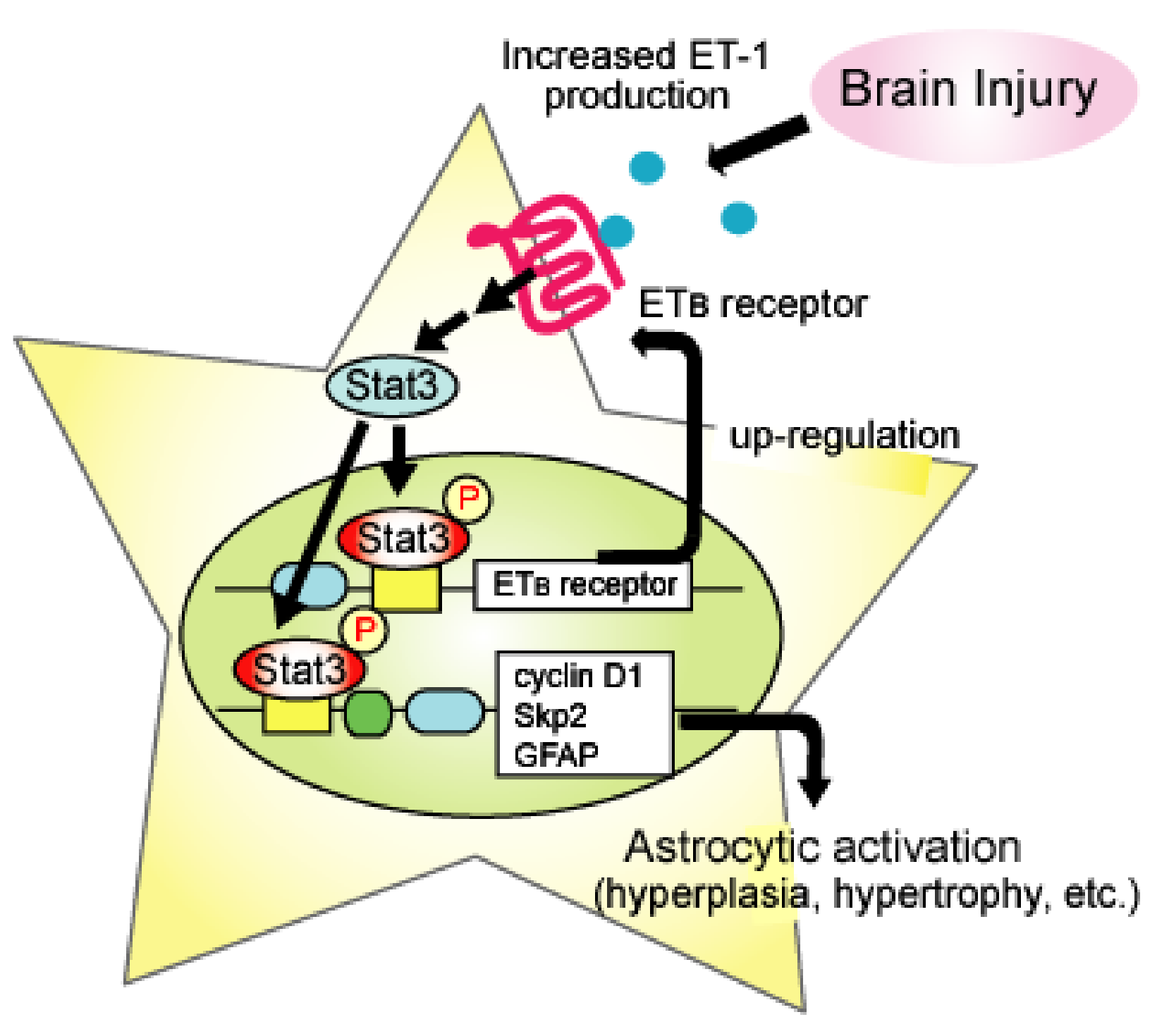
Figure 2.
Positive feedback mechanism of the astrocytic ETB receptor signal through activation of Stat3. Activation of astrocytic ETB receptors stimulates the transcription of cyclin D1, S-phase kinase-associated protein 2 (Skp2), and GFAP through Stat3. Increases in cyclin D1, skp2m, and GFAP proteins are involved in proliferation and hypertrophy associated with phenotype conversion to reactive astrocytes. Activated Stat3 also promotes transcription of ETB receptors. The upregulation of ETB receptors results in enhancement in Stat3-mediated gene expression in astrocytes.
Figure 2.
Positive feedback mechanism of the astrocytic ETB receptor signal through activation of Stat3. Activation of astrocytic ETB receptors stimulates the transcription of cyclin D1, S-phase kinase-associated protein 2 (Skp2), and GFAP through Stat3. Increases in cyclin D1, skp2m, and GFAP proteins are involved in proliferation and hypertrophy associated with phenotype conversion to reactive astrocytes. Activated Stat3 also promotes transcription of ETB receptors. The upregulation of ETB receptors results in enhancement in Stat3-mediated gene expression in astrocytes.

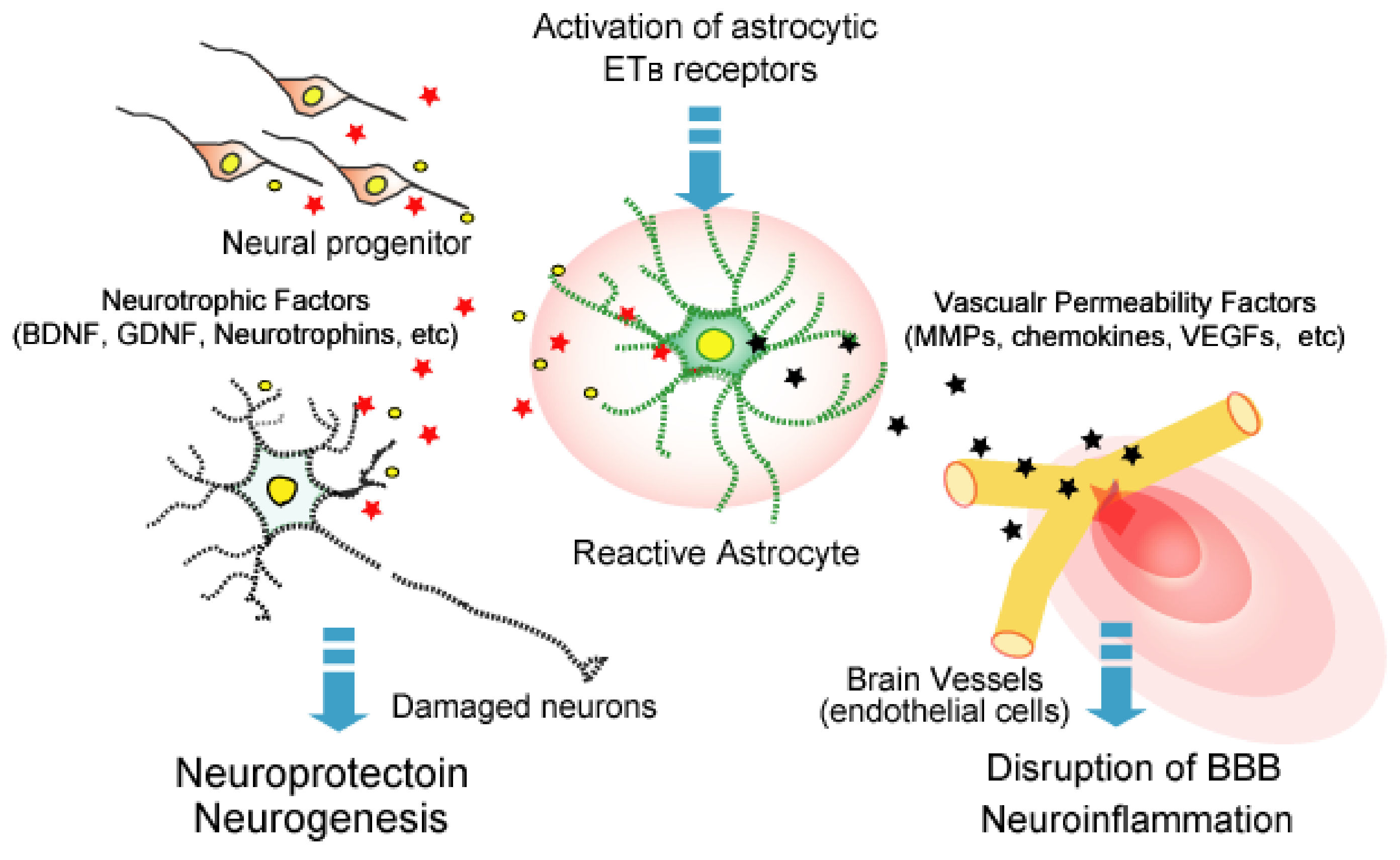
Figure 3.
Roles of ETB receptor-mediated bioactive factor production from reactive astrocytes. Increased ET-1 in brain disorders activates astrocytic ETB receptors and induces reactive astrocytes. Reactive astrocytes release various types of bioactive factors. These include factors that increase the permeability of brain microvessels to promote blood–brain barrier (BBB) disruption and neuroinflammation, and factors that promote nerve protection and neurogenesis.
Figure 3.
Roles of ETB receptor-mediated bioactive factor production from reactive astrocytes. Increased ET-1 in brain disorders activates astrocytic ETB receptors and induces reactive astrocytes. Reactive astrocytes release various types of bioactive factors. These include factors that increase the permeability of brain microvessels to promote blood–brain barrier (BBB) disruption and neuroinflammation, and factors that promote nerve protection and neurogenesis.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Agonists and antagonists for ET receptors.
| Agonist | Antagonist | |
|---|---|---|
| ET receptor non-selective | ET-1 | Bosentan, Macitentan |
| ETA selective | sarafotoxin 6b | Ambrisentan, Sitaxsentan, Atrasentan, Clazosentan, Zibotentan, S-0139, SB234551, Ro-61-1790 |
| ETB selective | sarafotoxin 6c, IRL-1620, BQ3020, Ala1,3,11,15-ET-1 | BQ788, IRL-2500, A192621, RES-701-1 |
Table 2.
Regulations of astrocytic bioactive factors by ETB receptors.
| Neurotrophic Factors | Vascular Permeability Regulators | Others | |
|---|---|---|---|
| Up-regulation | GDNF [72], BDNF [111], NGF [112] | VEGF [119], MMP2 [120], MMP3 [73], MMP9 [71,120] ET-1 [24] | CCL2/MCP-1 [122], CXCL1/CINC-1 [122] |
| Down-regulation | angiopoietin-1 [119,121] sonic hedgehog [123] | CX3CL1/fractalkine [122], ephrin-A2, -A4, -B2, -B3 [124] |
GDNF, glial cell line-derived neurotrophic factor; BDNF, brain-derived neurotrophic factor; NGF, nerve growth factor; VEGF, vascular endothelial growth factor; MMP, matrix metalloproteinases; ET-1, endothelin-1; MCP-1, monocyte chemotactic protein-1; CINC-1, cytokine-induced neutrophil chemoattractant-1.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Koyama, Y. Endothelin ETB Receptor-Mediated Astrocytic Activation: Pathological Roles in Brain Disorders. Int. J. Mol. Sci. 2021, 22, 4333. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094333
AMA Style
Koyama Y. Endothelin ETB Receptor-Mediated Astrocytic Activation: Pathological Roles in Brain Disorders. International Journal of Molecular Sciences. 2021; 22(9):4333. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094333
Chicago/Turabian StyleKoyama, Yutaka. 2021. "Endothelin ETB Receptor-Mediated Astrocytic Activation: Pathological Roles in Brain Disorders" International Journal of Molecular Sciences 22, no. 9: 4333. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094333
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.