
Functional Ambivalence of Dendritic Cells: Tolerogenicity and Immunogenicity

 , , , and
, , , and 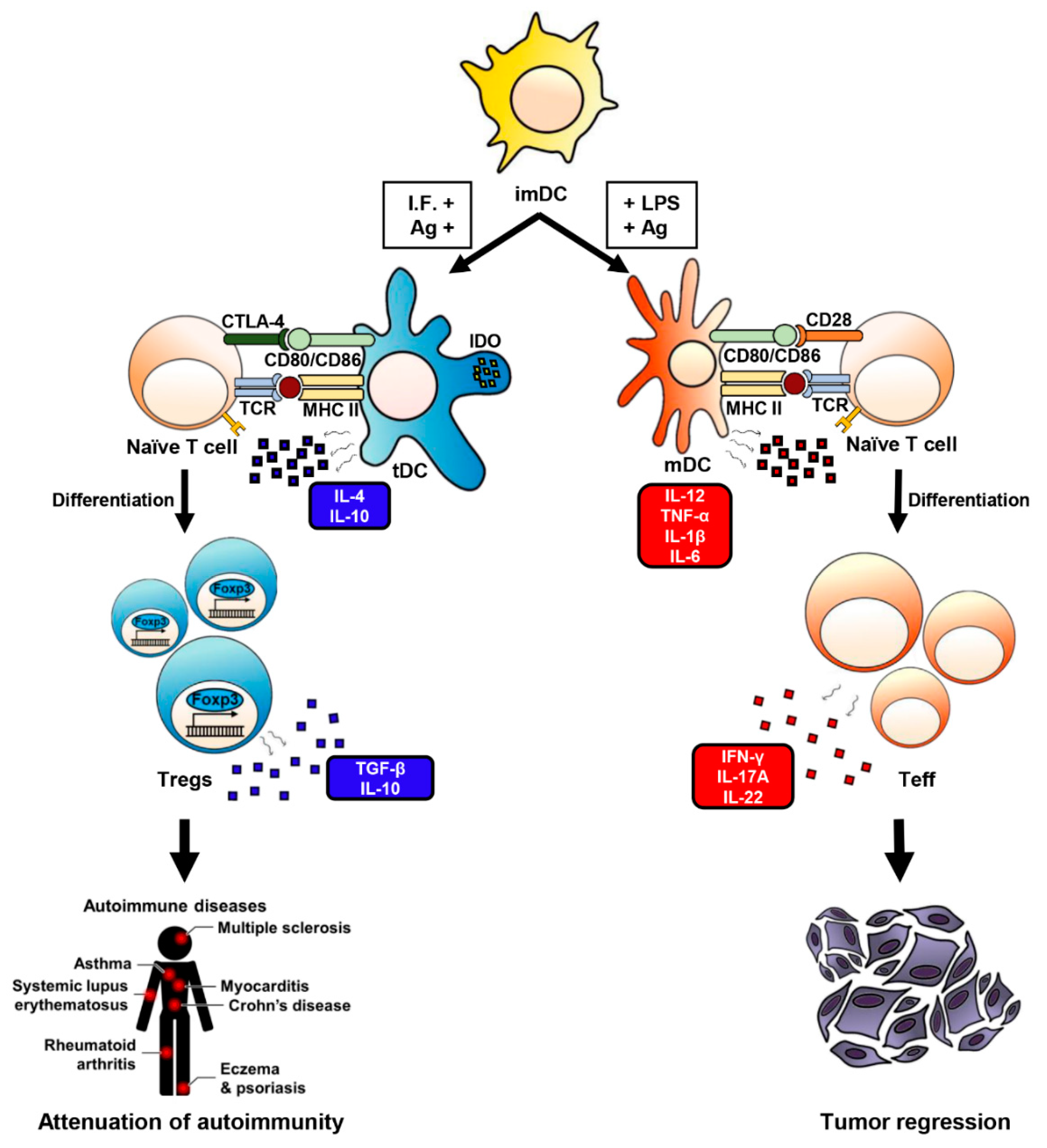{kind=link}
Abstract
:1. Introduction
2. DCs Play a Central Role in Inducing Anti-Tumor Immune Responses
3. Tolerogenicity of DCs
4. DCs Are Responsible for Peripheral Tolerance, But This Depends on the Method of Generation and/or the Presence of Immunosuppressive Agents
5. tDCs Induce Naïve T Cells Differentiation into Treg Cells
6. Identifying Reliable Markers of tDCs Is Important
7. tDCs Have Therapeutic Effects in Autoimmune and Inflammatory Diseases
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DCs | Dendritic cells |
| APCs | Antigen-presenting cells |
| imDCs | Immature DCs |
| tDCs | Tolerogenic DCs |
| mDCs | Fully-matured DCs |
| Treg cells | Regulatory T cells |
| RA | Rheumatoid arthritis |
| EAM | Experimental autoimmune myocarditis |
| AMI | Acute myocardial infarction |
| IDO | Indoleamine 2,3-dioxygenase |
| IL | Interleukin |
| Th1 cells | Type 1 T helper cells |
| Th2 cells | Type 2 T helper T cells |
| Th17 cells | Type 17 T helper T cells |
| CTL | Cytotoxic T lymphocyte |
| PMN | Polymorphonuclear |
| NK cells | Natural killer cells |
| TNF-α | Tumor necrosis factor- α |
| TGF-β | Transforming growth factor-β |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| PPAR-γ | Peroxisome proliferator-activated receptor-γ |
| NF-κB | Nuclear factor kappa light chain enhancer of activated B cells |
| Erk1/2 | Extracellular-signal-regulated kinase |
| PD-1 | Programmed death receptor-1 |
| CTLA-4 | Cytotoxic T lymphocyte-associated protein-4 |
References
- Vyas, J.M. The dendritic cell: The general of the army. Virulence 2012, 3, 601–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merad, M.; Manz, M.G. Dendritic cell homeostasis. Blood 2009, 113, 3418–3427. [Google Scholar] [CrossRef] [Green Version]
- Steinman, R.M. Linking innate to adaptive immunity through dendritic cells. Novartis Found Symp. 2006, 279, 101–109. [Google Scholar]
- Sabado, R.L.; Balan, S.; Bhardwaj, N. Dendritic cell-based immunotherapy. Cell Res. 2017, 27, 74–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, A.D.; Coulie, P.G.; van den Eynde, B.J.; Agostinis, P. Integrating next-generation dendritic cell vaccines into the current cancer immunotherapy landscape. Trends Immunol. 2017, 38, 577–593. [Google Scholar] [CrossRef] [PubMed]
- Guermonprez, P.; Valladeau, J.; Zitvogel, L.; Thery, C.; Amigorena, S. Antigen presentation and T cell stimulation by dendritic cells. Annu. Rev. Immunol. 2002, 20, 621–667. [Google Scholar] [CrossRef] [PubMed]
- Palucka, K.; Banchereau, J. Dendritic-cell-based therapeutic cancer vaccines. Immunity 2013, 39, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.; Singh, A.; Yu, G.; Bucayu, R.; Longbottom, T.; Nadeau, K. CCR9+ and CD103+ tolerogenic dendritic cell populations in food allergy patients undergoing oral immunotherapy. Clin. Transl. Allergy 2011, O51. [Google Scholar] [CrossRef] [Green Version]
- Malmström, V.; Shipton, D.; Singh, B.; Al-Shamkhani, A.; Puklavec, M.J.; Barclay, A.N.; Powrie, F. CD134L expression on dendritic cells in the mesenteric lymph nodes drives colitis in T cell-restored SCID mice. J. Immunol. 2001, 166, 6972–6981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, A.L.; Al-Hassi, H.O.; Rigby, R.J.; Bell, S.J.; Emmanuel, A.V.; Knight, S.C.; Kamm, M.A.; Stagg, A.J. Characteristics of intestinal dendritic cells in inflammatory bowel diseases. Gastroenterology 2005, 129, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Ruane, D.T.; Lavelle, E.C. The role of CD103+ dendritic cells in the intestinal mucosal immune system. Front. Immunol. 2011, 2, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlig, H.H.; McKenzie, B.S.; Hue, S.; Thompson, C.; Joyce-Shaikh, B.; Stepankova, R.; Robinson, N.; Buonocore, S.; Tlaskalova-Hogenova, H.; Cua, D.J.; et al. Differential activity of IL-12 and IL-23 in mucosal and systemic innate immune pathology. Immunity 2006, 25, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Byun, S.H.; Lee, J.H.; Jung, N.C.; Choi, H.J.; Song, J.Y.; Seo, H.G.; Choi, J.; Jung, S.Y.; Kang, S.; Choi, Y.S.; et al. Rosiglitazone-mediated dendritic cells ameliorate collagen-induced arthritis in mice. Biochem. Pharmcol. 2016, 115, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, T.H.; Park, H.E.; Lee, E.G.; Jung, N.C.; Song, J.Y.; Seo, H.G.; Seung, K.B.; Chang, K.; Lim, D.S. Myosin-primed tolerogenic dendritic cells ameliorate experimental autoimmune myocarditis. Cardiovasc. Res. 2014, 101, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Choo, E.H.; Lee, J.H.; Park, E.H.; Park, H.E.; Jung, N.C.; Kim, T.H.; Koh, Y.S.; Kim, E.; Seung, K.B.; Park, C.; et al. Infarcted myocardium-primed dendritic cells improve remodeling and cardiac function after myocardial infarction by modulating the regulatory T cell and macrophage polarization. Circulation 2017, 135, 1444–1457. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Jung, H.H.; Lee, C.K. Generation, Characteristics and Clinical Trials of Ex Vivo Generated Tolerogenic Dendritic cells. Yonsei Med. J. 2018, 59, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, H.; Matsumoto, T. Mechanisms of tolerance induction by dendritic cells in vivo. Front Immunol. 2018, 9, 350. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.G.; Jung, N.C.; Lee, J.H.; Song, J.Y.; Ryu, S.Y.; Seo, H.G.; Han, S.G.; Ahn, K.J.; Hong, K.S.; Choi, J.; et al. Tolerogenic dendritic cells show gene expression profiles that are different from those of immunogenic dendritic cells in DBA/1 mice. Autoimmunity 2016, 49, 90–101. [Google Scholar] [CrossRef]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef]
- Schuler, G.; Schuler-Thurner, B.; Steinman, R.M. The use of dendritic cells in cancer immunotherapy. Curr. Opin. Immunol. 2003, 15, 138–147. [Google Scholar] [CrossRef]
- Schuler, G.; Steinman, R.M. Dendritic cells as adjuvants for immune-mediated resistance to tumors. J. Exp. Med. 1997, 186, 1183–1187. [Google Scholar] [CrossRef] [PubMed]
- Kaliński, P.; Hilkens, C.M.; Wierenga, E.A.; Kapsenberg, M.L. T-cell priming by type-1 and type-2 polarized dendritic cells: The concept of a third signal. Immunol. Today 1999, 20, 561–567. [Google Scholar] [CrossRef]
- Gerson, J.N.; Ramamurthy, C.; Borghaei, H. Managing adverse effects of immunotherapy. Clin. Adv. Hematol. Oncol. 2018, 16, 364–374. [Google Scholar] [PubMed]
- Plaks, V.; Birnberg, T.; Berkutzki, T.; Sela, S.; BenYashar, A.; Kalchenko, V.; Mor, G.; Keshet, E.; Dekel, N.; Neeman, M.; et al. Uterine DCs are crucial for decidua formation during embryo implantation in mice. J. Clin. Investig. 2008, 118, 3954–3965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krey, G.; Frank, P.; Shaikly, V.; Barrientos, G.; Cordo-Russo, R.; Ringel, F.; Moschansky, P.; Chernukhin, I.V.; Metodiev, M.; Fernández, N.; et al. In vivo dendritic cell depletion reduces breeding efficiency, affecting implantation and early placental development in mice. J. Mol. Med. 2008, 86, 999–1011. [Google Scholar] [CrossRef] [PubMed]
- Panel, V.; Bachya, D.J.; Williamsb, M.; Ibrahim, A.A. Altered dendritic cell function in normal pregnancy. J. Reprod. Immunol. 2008, 78, 11–21. [Google Scholar]
- Shigeru, S.; Akitoshi, N.; Tomoko, S.; Mika, I. Th1/Th2/Th17 and regulatory T-cell paradigm in pregnancy. Am. J. Reprod. Immunol. 2010, 63, 601–610. [Google Scholar]
- Amodio, G.; Cichy, J.; Conde, P.; Matteoli, G.; Moreau, A.; Ochando, J.; Oral, B.H.; Pekarova, M.; Ryan, E.J.; Roth, J.; et al. Role of myeloid regulatory cells (MRCs) in maintaining tissue homeostasis and promoting tolerance in autoimmunity, inflammatory disease and transplantation. Cancer Immunol. Immunother. 2019, 68, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Majewski, P.; Majchrzak-Gorecka, M.; Grygier, B.; Skrzeczynska-Moncznik, J.; Osiecka, O.; Cichy, J. Inhibitors of serine proteases in regulating the production and function of neutrophil extracellular traps. Front. Immunol. 2016, 7, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vono, M.; Lin, A.; Norrby-Teglund, A.; Koup, R.A.; Liang, F.; Lore, K. Neutrophils acquire the capacity for antigen presentation to memory CD4(+) T cells in vitro and ex vivo. Blood 2017, 129, 1991–2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmona-Rivera, C.; Kaplan, M.J. Low-density granulocytes: A distinct class of neutrophils in systemic autoimmunity. Semin. Immunopathol. 2013, 35, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Morell, M.; Varela, N.; Maranon, C. Myeloid populations in systemic autoimmune diseases. Clin. Rev. Allergy Immunol. 2017, 53, 198–218. [Google Scholar] [CrossRef]
- Amoruso, A.; Sola, D.; Rossi, L.; Obeng, J.A.; Fresu, L.G.; Sainaghi, P.P.; Pirisi, M.; Brunelleschi, S. Relation among anti-rheumatic drug therapy, CD14(+)CD16(+) blood monocytes and disease activity markers (DAS28 and US7 scores) in rheumatoid arthritis: A pilot study. Pharmacol. Res. 2016, 107, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Amodio, G.; Gregori, S. Dendritic cells: A doubleedge sword in autoimmune responses. Front. Immunol. 2012, 3, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, J.L.; Leytze, G.M.; Emswiler, J.; Peach, R.; Bajorath, J.; Cosand, W.; Linsley, P.S. Covalent Dimerization of CD28/CTLA-4 and Oligomerization of CD80/CD86 Regulate T Cell Costimulatory Interactions. J. Biol. Chem. 1996, 271, 26762–26771. [Google Scholar] [CrossRef] [Green Version]
- Rudulier, C.D.; McKinstry, K.K.; Al-Yassin, G.A.; Kroeger, D.R.; Bretscher, P.A. The number of responding CD4 T cells and the dose of antigen conjointly determine the TH1/TH2 phenotype by modulating B7/CD28 interactions. J. Immunol. 2014, 192, 5140–5150. [Google Scholar] [CrossRef] [Green Version]
- Lucy, S.K.W. Treg and CTLA-4: Two intertwining pathways to immune tolerance. J. Autoimmun. 2013, 45, 49–57. [Google Scholar]
- Giannoukakis, N.; Phillips, B.; Finegold, D.; Harnaha, J.; Trucco, M. Phase I (safety) study of autologous tolerogenic dendritic cells in type 1 diabetic patients. Diabetes Care 2011, 34, 2026–2032. [Google Scholar] [CrossRef] [Green Version]
- Harry, R.A.; Anderson, A.E.; Isaacs, J.D.; Hilkens, C.M. Generation and characterisation of therapeutic tolerogenic dendritic cells for rheumatoid arthritis. Ann. Rheum. Dis. 2010, 69, 2042–2050. [Google Scholar] [CrossRef] [Green Version]
- Schreiner, B.; Mitsdoerffer, M.; Kieseier, B.C.; Chen, L.; Hartung, H.P.; Weller, M.; Wiendl, H. Interferon-beta enhances monocyte and dendritic cell expression of B7-H1 (PD-L1), a strong inhibitor of autologous T-cell activation: Relevance for the immune modulatory effect in multiple sclerosis. J. Neuroimmunol. 2004, 155, 172–182. [Google Scholar] [CrossRef]
- Lim, D.S.; Kang, M.S.; Jeong, J.A.; Bae, Y.S. Semi-mature DC are immunogenic and not tolerogenic when inoculated at a high dose in collagen-induced arthritis mice. Eur. J. Immunol. 2009, 39, 1334–1343. [Google Scholar] [CrossRef]
- Torres-Aguilar, H.; Aguilar-Ruiz, S.R.; González-Pérez, G.; Munguía, R.; Bajaña, S.; Meraz-Ríos, M.A.; Sánchez-Torres, C. Tolerogenic dendritic cells generated with different immunosuppressive cytokines induce antigen-specific anergy and regulatory properties in memory CD4+ T cells. J. Immunol. 2010, 184, 1765–1775. [Google Scholar] [CrossRef] [Green Version]
- Gregori, S.; Tomasoni, D.; Pacciani, V.; Scirpoli, M.; Battaglia, M.; Magnani, C.F.; Hauben, E.; Roncarolo, M.-G. Differentiation of type 1 T regulatory cells (Tr1) by tolerogenic DC-10 requires the IL-10-dependent ILT4/HLA-G pathway. Blood 2010, 116, 935–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.G.; Wang, J.; Wang, P.; Gray, J.D.; Horwitz, D.A. IL-2 is essential for TGF-beta to convert naive CD4+CD25- cells to CD25+Foxp3+ regulatory T cells and for expansion of these cells. J. Immunol. 2007, 178, 2018–2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.L.; Pittet, M.J.; Gorelik, L.; Flavell, R.A.; Weissleder, R.; Von Boehmer, H.; Khazaie, K. Regulatory T cells suppress tumor-specific CD8 T cell cytotoxicity through TGF-β signals in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 419–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalev, I.; Schmelzle, M.; Robsonb, S.C.; Levy, G. Making sense of regulatory T cell suppressive function. Semin. Immunol. 2011, 23, 282–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Facciabene, A.; Motz, G.T.; Coukos, G. T-regulatory cells: Key players in tumor immune escape and angiogenesis. Cancer Res. 2012, 72, 2162–2171. [Google Scholar] [CrossRef] [Green Version]
- Prendergast, G.C. Immune escape as a fundamental trait of cancer: Focus on IDO. Oncogene 2008, 27, 3889–3900. [Google Scholar] [CrossRef] [Green Version]
- Gianchecchi, E.; Fierabracci, A. Inhibitory Receptors and Pathways of Lymphocytes: The Role of PD-1 in Treg Development and Their Involvement in Autoimmunity Onset and Cancer Progression. Front. Immunol. 2018, 9, 2374. [Google Scholar] [CrossRef] [Green Version]
- Dannull, J.; Su, Z.; Rizzieri, D.; Yang, B.K.; Coleman, D.; Yancey, D.; Zhang, A.; Dahm, P.; Chao, N.; Gilboa, E.; et al. Enhancement of vaccine-mediated antitumor immunity in cancer patients after depletion of regulatory T cells. J. Clin. Investig. 2005, 115, 3623–3633. [Google Scholar] [CrossRef]
- Taylor, N.A.; Vick, S.C.; Iglesia, M.D.; Brickey, W.J.; Midkiff, B.R.; McKinnon, K.P.; Reisdorf, S.; Anders, C.K.; Carey, L.A.; Parker, J.S.; et al. Treg depletion potentiates checkpoint inhibition in claudin-low breast cancer. J. Clin. Investig. 2017, 127, 3472–3483. [Google Scholar] [CrossRef] [PubMed]
- Cvetanovich, G.L.; Hafler, D.A. Human Regulatory T Cells in Autoimmune Diseases. Curr. Opin. Immunol. 2010, 22, 753–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afzali, B.; Lombardi, G.; Lechler, R.I.; Lord, G.M. The role of T helper 17 (Th17) and regulatory T cells (Treg) in human organ transplantation and autoimmune disease. Clin. Exp. Immunol. 2007, 148, 32–46. [Google Scholar] [CrossRef] [Green Version]
- Kitz, A.; de Marcken, M.; Gautron, A.; Mitrovic, M.; Hafler, D.A.; Dominguez-Villar, M. AKT isoforms modulate Th1-like Treg generation and function in human autoimmune disease. EMBO Rep. 2016, 17, 1169–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.; Bluestone, J.A. The Foxp3+ regulatory T cell: A jack of all trades, master of regulation. Nat. Immunol. 2008, 9, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Matteoli, G.; Mazzini, E.; Iliev, I.D.; Mileti, E.; Fallarino, F.; Puccetti, P.; Chieppa, M.; Rescigno, M. Gut CD103+ dendritic cells express indoleamine 2,3-dioxygenase which influences T regulatory/T effector cell balance and oral tolerance induction. Gut 2009, 59, 595–604. [Google Scholar] [CrossRef]
- Scott, C.L.; Aumeunier, A.M.; Mowat, A.M. Intestinal CD103+ dendritic cells: Master regulators of tolerance? Trends Immunol. 2011, 32, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Mascarell, L.; Airouche, S.; Berjont, N.; Gary, C.; Gueguen, C.; Fourcade, G.; Bellier, B.; Togbe, D.; Ryffel, B.; Klatzmann, D.; et al. The regulatory dendritic cell marker C1q is a potent inhibitor of allergic inflammation. Mucosal. Immunol. 2017, 10, 695–704. [Google Scholar] [CrossRef]
- Son, M.; Santiago-Schwarz, F.; Al-Abed, Y.; Diamond, B. C1q limits dendritic cell differentiation and activation by engaging LAIR-1. Proc. Natl. Acad. Sci. USA 2012, 109, E3160–E3167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairweather, D.; Root-Bernstein, R. Autoimmune Disease: Mechanisms. In Els; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Takada, K.; Takahama, Y. Positive-selection-inducing self-peptides displayed by cortical thymic epithelial cells. Adv. Immunol. 2015, 125, 87–110. [Google Scholar]
- Anderson, G.; Takahama, Y. Thymic epithelial cells: Working class heroes for T cell development and repertoire selection. Trends Immunol. 2012, 33, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Takahama, Y.; Takada, K.; Murata, S.; Tanaka, K. β5t-containing thymoproteasome: Specific expression in thymic cortical epithelial cells and role in positive selection of CD8+ T cells. Curr. Opin. Immunol. 2012, 24, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Klein, L.; Kyewski, B.; Allen, P.M.; Hogquist, K.A. Positive and negative selection of the T cell repertoire: What thymocytes see (and don’t see). Nat. Rev. Immunol. 2014, 14, 377–391. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, F.S.; Gershwin, M.E. Human autoimmune diseases: A comprehensive update. J. Intern. Med. 2015, 278, 369–395. [Google Scholar] [CrossRef]
- Marrack, P.; Kappler, J.; Kotzin, B.L. Autoimmune disease: Why and where it occurs. Nat. Med. 2001, 7, 899–905. [Google Scholar] [CrossRef]
- Fairweather, D.; Petri, M.A.; Coronado, M.J.; Cooper, L.T. Autoimmune heart disease: Role of sex hormones and autoantibodies in disease pathogenesis. Expert Rev. Clin. Immunol. 2012, 8, 269–284. [Google Scholar] [CrossRef]
- Kuchroo, V.K.; Das, M.P.; Brown, J.A.; Ranger, A.M.; Zamvil, S.S.; Sobel, R.A.; Weiner, H.L.; Nabavi, N.; Glimcher, L.H. B7-1 and B7-2 Costimulatory Molecules Activate Differentially the Thl/Th2 Developmental Pathways: Application to Autoimmune Disease Therapy. Cell 1996, 80, 707–718. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Chen, H.M.; Subudhi, S.K.; Chen, J.; Koka, R.; Chen, L.; Fu, Y.X. Costimulatory molecule-targeted antibody therapy of a spontaneous autoimmune disease. Nat. Med. 2002, 8, 1405–1413. [Google Scholar] [CrossRef]
- Yasunaga, M. Antibody therapeutics and immunoregulation in cancer and autoimmune disease. Semin. Cancer Biol. 2020, 64, 1–12. [Google Scholar] [CrossRef]
- Bonam, S.R.; Wang, F.; Muller, S. Autophagy: A new concept in autoimmunity regulation and a novel therapeutic option. J. Autoimmun. 2018, 94, 16–32. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, J.-H.; Lee, J.-H.; Choi, S.-Y.; Jung, N.-C.; Song, J.-Y.; Seo, H.-G.; Lim, D.-S. Functional Ambivalence of Dendritic Cells: Tolerogenicity and Immunogenicity. Int. J. Mol. Sci. 2021, 22, 4430. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094430
Nam J-H, Lee J-H, Choi S-Y, Jung N-C, Song J-Y, Seo H-G, Lim D-S. Functional Ambivalence of Dendritic Cells: Tolerogenicity and Immunogenicity. International Journal of Molecular Sciences. 2021; 22(9):4430. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094430
Chicago/Turabian StyleNam, Ji-Hee, Jun-Ho Lee, So-Yeon Choi, Nam-Chul Jung, Jie-Young Song, Han-Geuk Seo, and Dae-Seog Lim. 2021. "Functional Ambivalence of Dendritic Cells: Tolerogenicity and Immunogenicity" International Journal of Molecular Sciences 22, no. 9: 4430. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094430