
Orexin A Enhances Pro-Opiomelanocortin Transcription Regulated by BMP-4 in Mouse Corticotrope AtT20 Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
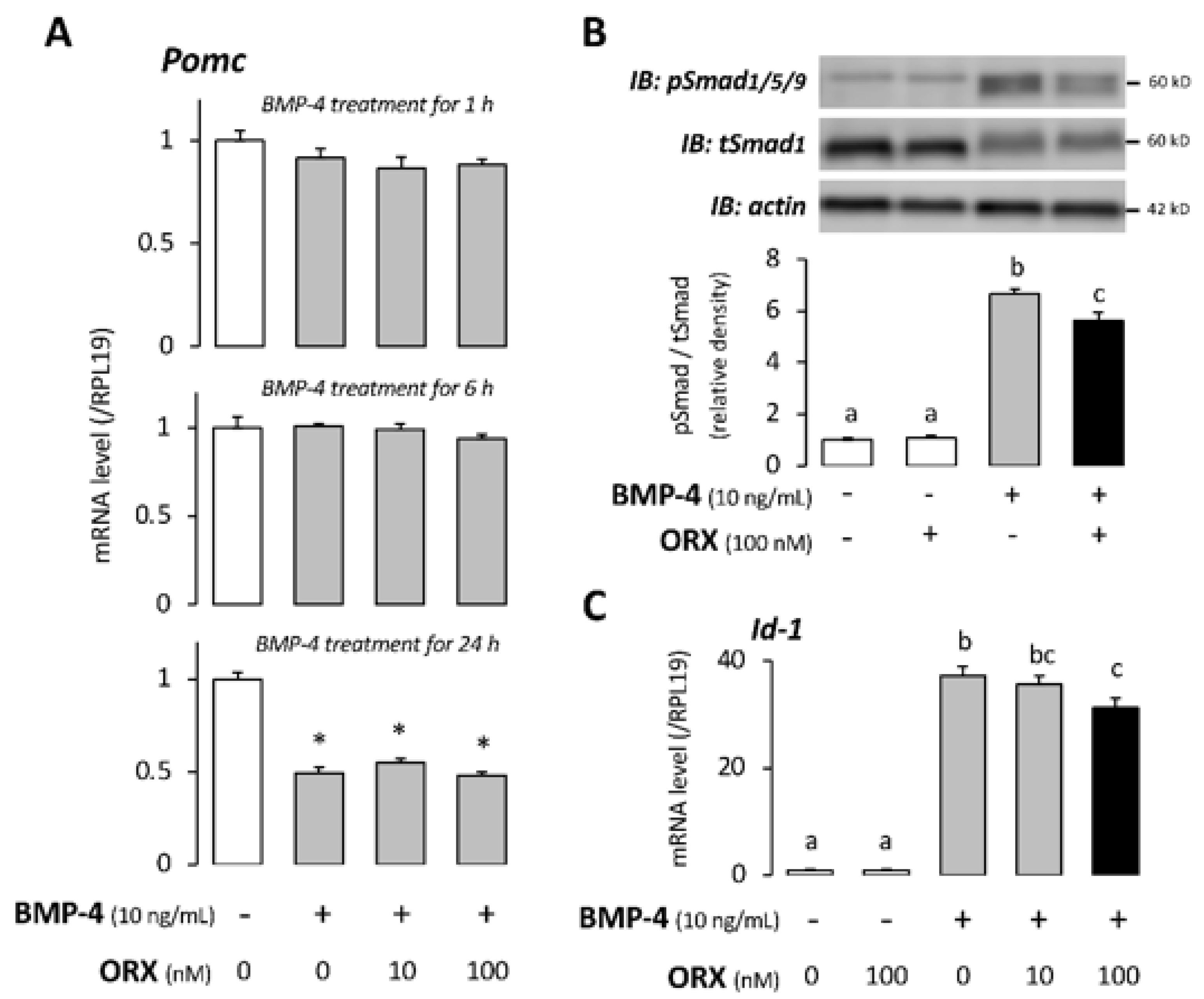
2. Results
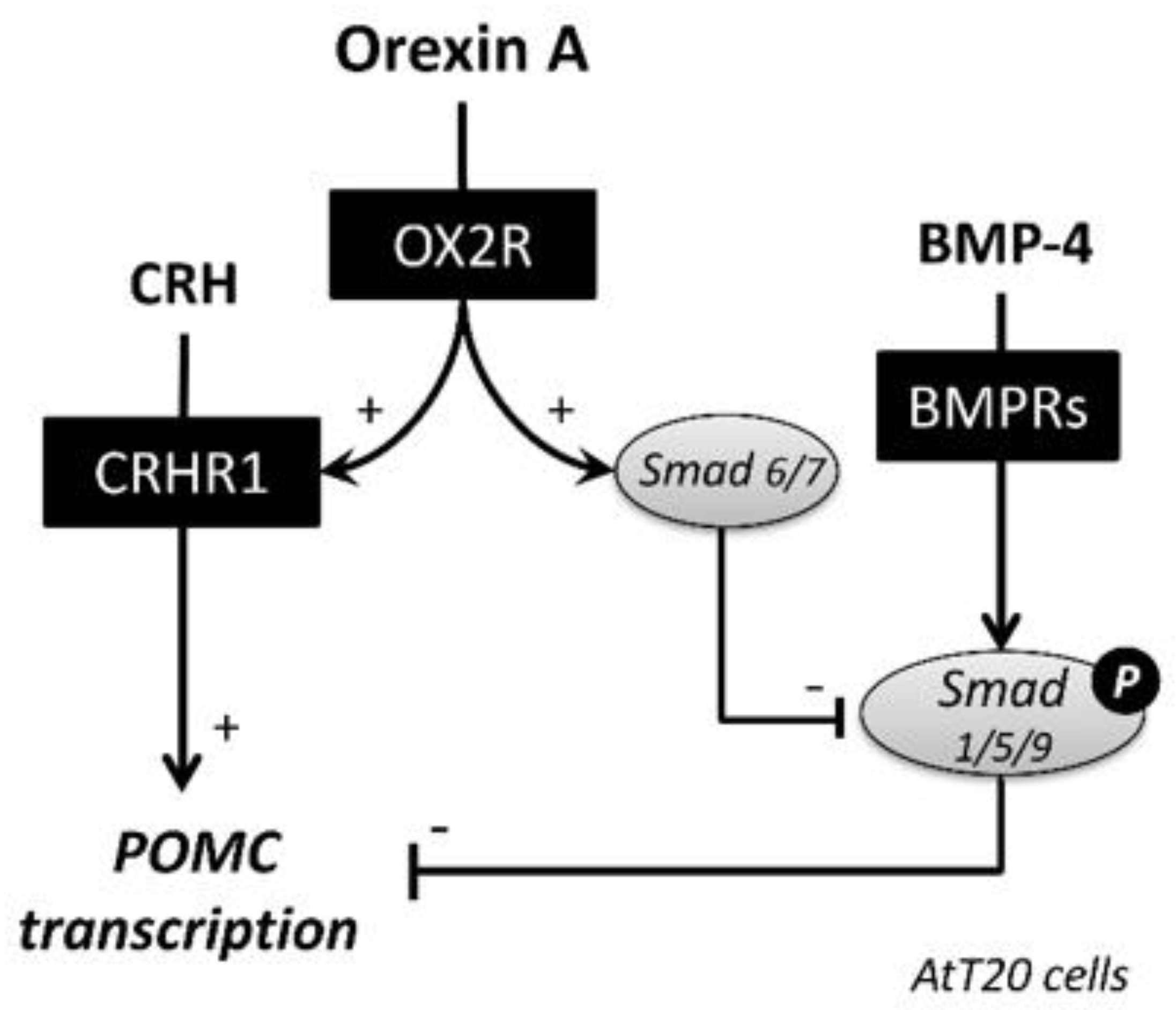
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. RNA Extraction and Quantitative PCR
4.4. Western Immunoblotting
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACTH | adrenocorticotropin |
| ActRII | activin type II receptor |
| ALK | activin receptor-like kinase |
| AVP | arginine vasopressin |
| BMP | bone morphogenetic protein |
| BMPRII | BMP type II receptor |
| CRH | corticotropin-releasing hormone |
| CRHR | CRH receptor |
| HPA | hypothalamic-pituitary-adrenal |
| ORX | orexin A |
| OX1R | orexin type 1 receptor |
| OX2R | orexin type 2 receptor |
| POMC | pro-opiomelanocortin |
| PVN | paraventricular nucleus |
| (TGF)β | transforming growth factor (TGF)β |
References
- Sakurai, T. The role of orexin in motivated behaviours. Nat. Rev. Neurosci. 2014, 15, 719–731. [Google Scholar] [CrossRef]
- Xu, T.-R.; Yang, Y.; Ward, R.; Gao, L.; Liu, Y. Orexin receptors: Multi-functional therapeutic targets for sleeping disorders, eating disorders, drug addiction, cancers and other physiological disorders. Cell. Signal. 2013, 25, 2413–2423. [Google Scholar] [CrossRef]
- De Lecea, L.; Kilduff, T.S.; Peyron, C.; Gao, X.; Foye, P.E.; Danielson, P.E.; Fukuhara, C.; Battenberg, E.L.; Gautvik, V.T.; Bartlett, F.S., 2nd; et al. The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity. Proc. Natl. Acad. Sci. USA 1998, 95, 322–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, T.; Amemiya, A.; Ishii, M.; Matsuzaki, I.; Chemelli, R.M.; Tanaka, H.; Williams, S.C.; Richardson, J.A.; Kozlowski, G.P.; Wilson, S.; et al. Orexins and orexin receptors: A family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 1998, 92, 573–585. [Google Scholar] [CrossRef] [Green Version]
- Heinonen, M.V.; Purhonen, A.K.; Mäkelä, K.A.; Herzig, K.H. Functions of orexins in peripheral tissues. Acta Physiol. 2008, 192, 471–485. [Google Scholar] [CrossRef]
- López, M.; Tena-Sempere, M.; Diéguez, C. Cross-talk between orexins (hypocretins) and the neuroendocrine axes (hypothalamic–pituitary axes). Front. Neuroendocr. 2010, 31, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Kagerer, S.M.; Jöhren, O. Interactions of orexins/hypocretins with adrenocortical functions. Acta Physiol. 2010, 198, 361–371. [Google Scholar] [CrossRef]
- Winsky-Sommerer, R.; Yamanaka, A.; Diano, S.; Borok, E.; Roberts, A.J.; Sakurai, T.; Kilduff, T.S.; Horvath, T.L.; De Lecea, L. Interaction between the Corticotropin-Releasing Factor System and Hypocretins (Orexins): A Novel Circuit Mediating Stress Response. J. Neurosci. 2004, 24, 11439–11448. [Google Scholar] [CrossRef]
- Jászberényi, M.; Bujdosó, E.; Pataki, I.; Telegdy, G. Effects of Orexins on the Hypothalamic-Pituitary-Adrenal System. J. Neuroendocr. 2001, 12, 1174–1178. [Google Scholar] [CrossRef]
- Kuru, M.; Ueta, Y.; Serino, R.; Nakazato, M.; Yamamoto, Y.; Shibuya, I.; Yamashita, H. Centrally administered orexin/hypocretin activates HPA axis in rats. NeuroReport 2000, 11, 1977–1980. [Google Scholar] [CrossRef] [PubMed]
- Samson, W.K.; Taylor, M.M.; Follwell, M.; Ferguson, A.V. Orexin actions in hypothalamic paraventricular nucleus: Physiological consequences and cellular correlates. Regul. Pept. 2002, 104, 97–103. [Google Scholar] [CrossRef]
- Al-Barazanji, K.A.; Wilson, S.; Baker, J.; Jessop, D.S.; Harbuz, M.S. Central Orexin-A Activates Hypothalamic-Pituitary-Adrenal Axis and Stimulates Hypothalamic Corticotropin Releasing Factor and Arginine Vasopressin Neurones in Conscious Rats. J. Neuroendocr. 2001, 13, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Mazzocchi, G.; Malendowicz, L.K.; Gottardo, L.; Aragona, F.; Nussdorfer, G.G. Orexin A Stimulates Cortisol Secretion from Human Adrenocortical Cells through Activation of the Adenylate Cyclase-Dependent Signaling Cascade. J. Clin. Endocrinol. Metab. 2001, 86, 778–782. [Google Scholar] [CrossRef] [PubMed]
- Malendowicz, L.K.; Tortorella, C.; Nussdorfer, G.G. Orexins stimulate corticosterone secretion of rat adrenocortical cells, through the activation of the adenylate cyclase-dependent signaling cascade. J. Steroid Biochem. Mol. Biol. 1999, 70, 185–188. [Google Scholar] [CrossRef]
- Inutsuka, A.; Yamanaka, A. The physiological role of orexin/hypocretin neurons in the regulation of sleep/wakefulness and neuroendocrine functions. Front. Endocrinol. 2013, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, M.; Lopez, M.; Garcia-Caballero, T.; Gallego, R.; Vazquez-Boquete, A.; Morel, G.; Senaris, R.; Casanueva, F.; Dieguez, C.; Beiras, A. Cellular localization of orexin receptors in human pituitary. J. Clin. Endocrinol. Metab. 2001, 86, 3444–3447. [Google Scholar] [CrossRef]
- Otsuka, F. Multiple Endocrine Regulation by Bone Morphogenetic Protein System. Endocr. J. 2010, 57, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Labeur, M.; Paez-Pereda, M.; Haedo, M.; Arzt, E.; Stalla, G.K. Pituitary tumors: Cell type-specific roles for BMP-4. Mol. Cell. Endocrinol. 2010, 326, 85–88. [Google Scholar] [CrossRef]
- Otsuka, F.; Tsukamoto, N.; Miyoshi, T.; Iwasaki, Y.; Makino, H. BMP action in the pituitary: Its possible role in modulating somatostatin sensitivity in pituitary tumor cells. Mol. Cell. Endocrinol. 2012, 349, 105–110. [Google Scholar] [CrossRef]
- Tsukamoto, N.; Otsuka, F.; Miyoshi, T.; Inagaki, K.; Nakamura, E.; Terasaka, T.; Takeda, M.; Ogura, T.; Iwasaki, Y.; Makino, H. Functional interaction of bone morphogenetic protein and growth hormone releasing peptide in adrenocorticotropin regulation by corticotrope cells. Mol. Cell. Endocrinol. 2011, 344, 41–50. [Google Scholar] [CrossRef]
- Tsukamoto, N.; Otsuka, F.; Miyoshi, T.; Yamanaka, R.; Inagaki, K.; Yamashita, M.; Otani, H.; Takeda, M.; Suzuki, J.; Ogura, T.; et al. Effects of Bone Morphogenetic Protein (BMP) on Adrenocorticotropin Production by Pituitary Corticotrope Cells: Involvement of Up-Regulation of BMP Receptor Signaling by Somatostatin Analogs. Endocrinology 2010, 151, 1129–1141. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, S.; Komatsubara, M.; Ogura-Ochi, K.; Tsukamoto-Yamauchi, N.; Toma, K.; Inagaki, K.; Wada, J.; Otsuka, F. Orexin A modulates prolactin production by regulating BMP-4 activity in rat pituitary lactotorope cells. Peptides 2019, 113, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcus, J.N.; Aschkenasi, C.J.; Lee, C.E.; Chemelli, R.M.; Saper, C.B.; Yanagisawa, M.; Elmquist, J.K. Differential expression of orexin receptors 1 and 2 in the rat brain. J. Comp. Neurol. 2001, 435, 6–25. [Google Scholar] [CrossRef]
- Chang, H.; Saito, T.; Ohiwa, N.; Tateoka, M.; Deocaris, C.C.; Fujikawa, T.; Soya, H. Inhibitory effects of an orexin-2 receptor antagonist on orexin A- and stress-induced ACTH responses in conscious rats. Neurosci. Res. 2007, 57, 462–466. [Google Scholar] [CrossRef]
- Grafe, L.A.; Eacret, D.; Luz, S.; Gotter, A.L.; Renger, J.J.; Winrow, C.J.; Bhatnagar, S. Orexin 2 receptor regulation of the hypothalamic-pituitary-adrenal (HPA) response to acute and repeated stress. Neuroscience 2017, 348, 313–323. [Google Scholar] [CrossRef]
- Heydendael, W.; Sengupta, A.; Bhatnagar, S. Putative genes mediating the effects of orexins in the posterior paraventricular thalamus on neuroendocrine and behavioral adaptations to repeated stress. Brain Res. Bull. 2012, 89, 203–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prajapati, S.K.; Krishnamurthy, S. Non-selective orexin-receptor antagonist attenuates stress-re-stress-induced core PTSD-like symptoms in rats: Behavioural and neurochemical analyses. Behav. Brain Res. 2021, 399, 113015. [Google Scholar] [CrossRef]
- Samson, W.K.; Taylor, M.M. Hypocretin/orexin suppresses corticotroph responsiveness in vitro. Am. J. Physiol. Integr. Comp. Physiol. 2001, 281, R1140–R1145. [Google Scholar] [CrossRef]
- Kamiński, T.; Smolinska, N. Expression of Orexin Receptors in the Pituitary. Vitam. Horm. 2012, 89, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Aoki, Y.; Iwasaki, Y.; Katahira, M.; Oiso, Y.; Saito, H. Regulation of the Rat Proopiomelanocortin Gene Expression in AtT-20 Cells. I: Effects of the Common Secretagogues. Endocrinology 1997, 138, 1923–1929. [Google Scholar] [CrossRef]
- Fujita, S.; Hasegawa, T.; Nishiyama, Y.; Fujisawa, S.; Nakano, Y.; Nada, T.; Iwata, N.; Kamada, Y.; Masuyama, H.; Otsuka, F. Interaction between orexin A and bone morphogenetic protein system on progesterone biosynthesis by rat granulosa cells. J. Steroid Biochem. Mol. Biol. 2018, 181, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Kodadek, T.; Cai, D. Chemistry and biology of orexin signaling. Mol. BioSyst. 2010, 6, 1366–1375. [Google Scholar] [CrossRef]
- Otsuka, F. Interaction of Melatonin and BMP-6 in Ovarian Steroidogenesis. Vitam. Horm. 2018, 107, 137–153. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto-Yamauchi, N.; Terasaka, T.; Iwasaki, Y.; Otsuka, F. Interaction of pituitary hormones and expression of clock genes modulated by bone morphogenetic protein-4 and melatonin. Biochem. Biophys. Res. Commun. 2015, 459, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Nagamine, T. Neuroendocrine, Autonomic, and Metabolic Responses to an Orexin Antagonist, Suvorexant, in Psychiatric Patients with Insomnia. Innov. Clin. Neurosci. 2017, 14, 30–37. [Google Scholar]
- Beuckmann, C.T.; Ueno, T.; Nakagawa, M.; Suzuki, M.; Akasofu, S. Preclinical in vivo characterization of lemborexant (E2006), a novel dual orexin receptor antagonist for sleep/wake regulation. Sleep 2019, 42, zsz076. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujisawa, S.; Komatsubara, M.; Tsukamoto-Yamauchi, N.; Iwata, N.; Nada, T.; Wada, J.; Otsuka, F. Orexin A Enhances Pro-Opiomelanocortin Transcription Regulated by BMP-4 in Mouse Corticotrope AtT20 Cells. Int. J. Mol. Sci. 2021, 22, 4553. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094553
Fujisawa S, Komatsubara M, Tsukamoto-Yamauchi N, Iwata N, Nada T, Wada J, Otsuka F. Orexin A Enhances Pro-Opiomelanocortin Transcription Regulated by BMP-4 in Mouse Corticotrope AtT20 Cells. International Journal of Molecular Sciences. 2021; 22(9):4553. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094553
Chicago/Turabian StyleFujisawa, Satoshi, Motoshi Komatsubara, Naoko Tsukamoto-Yamauchi, Nahoko Iwata, Takahiro Nada, Jun Wada, and Fumio Otsuka. 2021. "Orexin A Enhances Pro-Opiomelanocortin Transcription Regulated by BMP-4 in Mouse Corticotrope AtT20 Cells" International Journal of Molecular Sciences 22, no. 9: 4553. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094553