
Altered Composition of Microbiota in Women with Ovarian Endometrioma: Microbiome Analyses of Extracellular Vesicles in the Peritoneal Fluid

 , ,
, ,
Abstract
:1. Introduction
2. Results
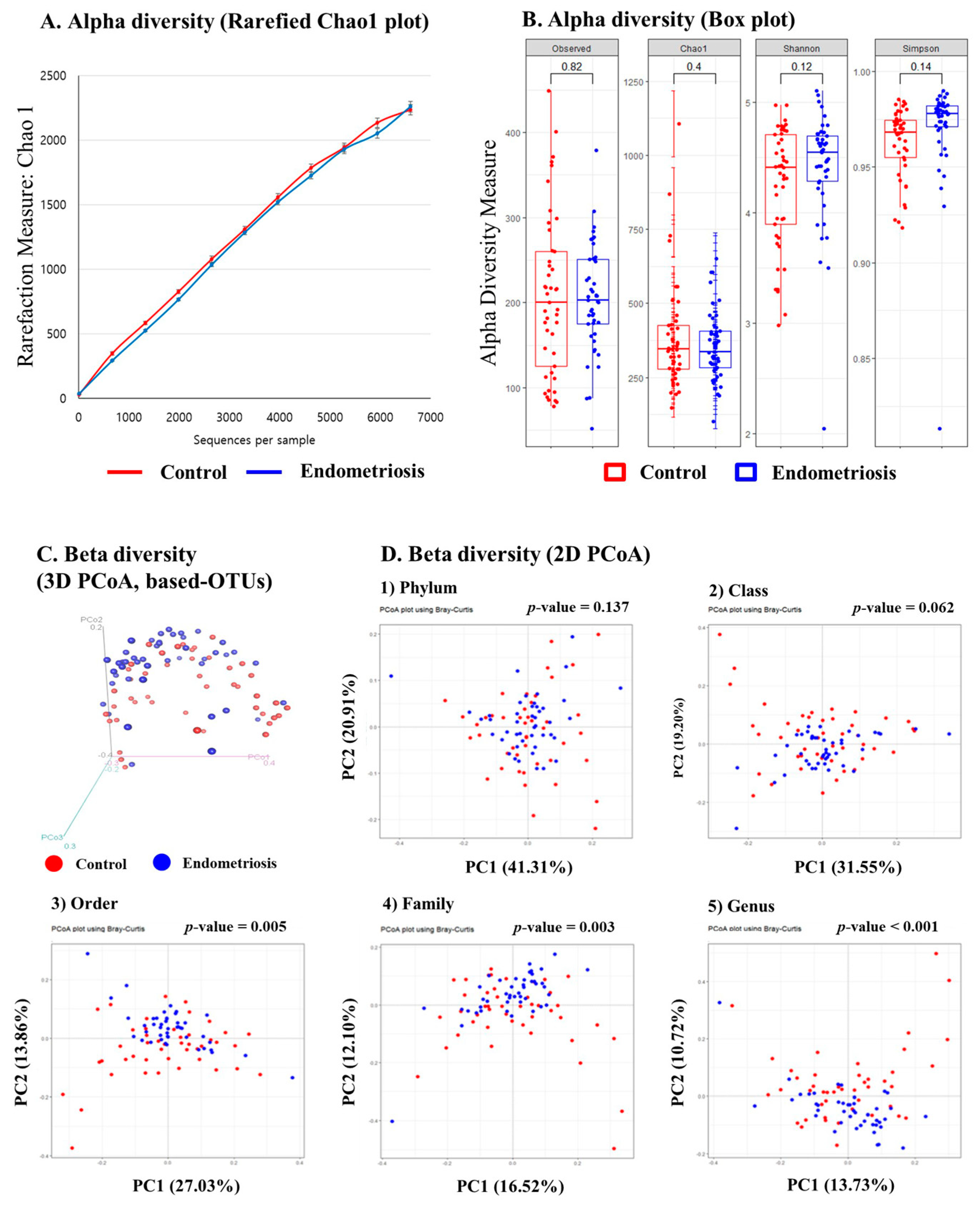
2.1. Microbiome Analysis of EVs in PF: Alpha Diversity
2.2. Microbiome Analysis of EVs in PF: Beta Diversity
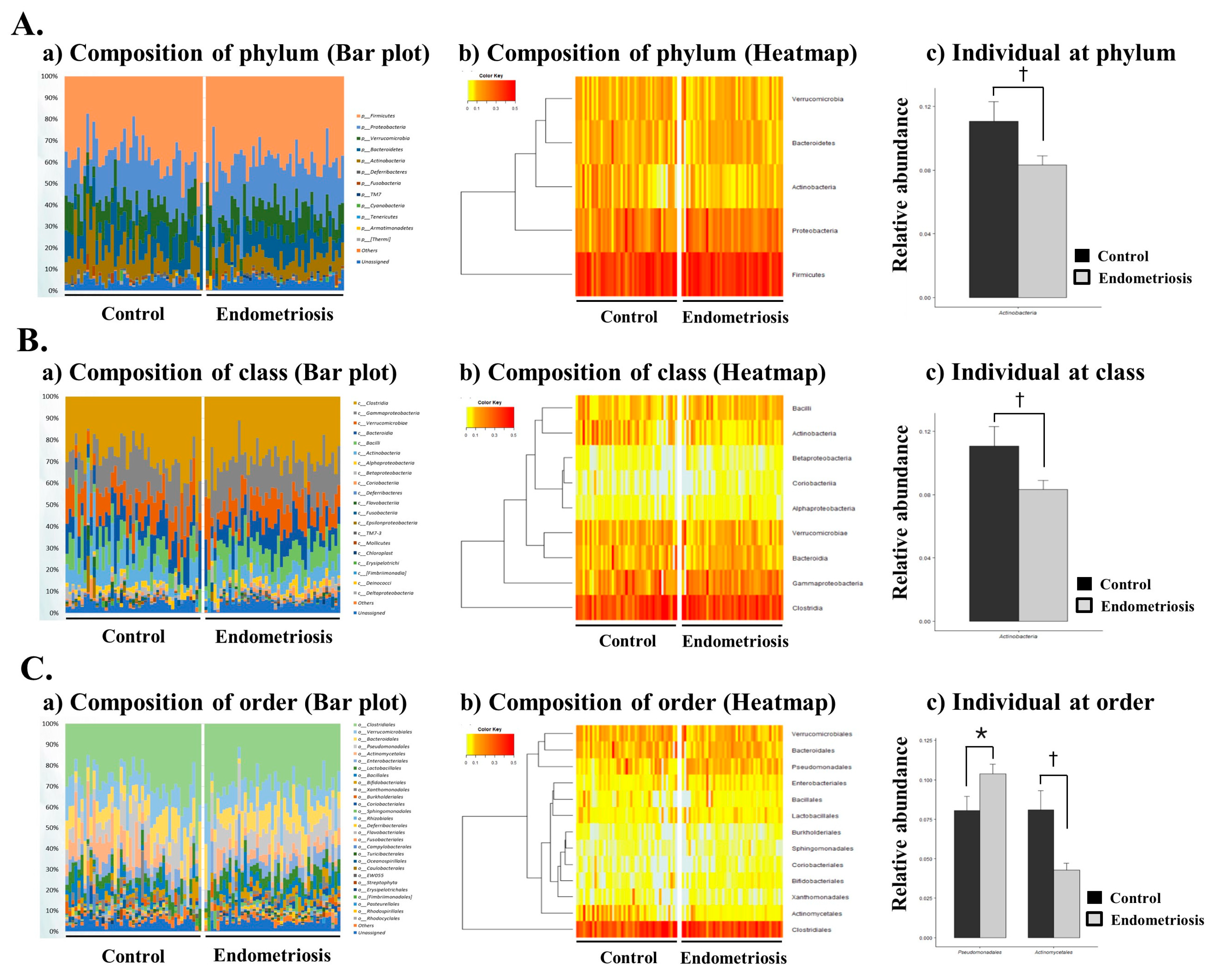
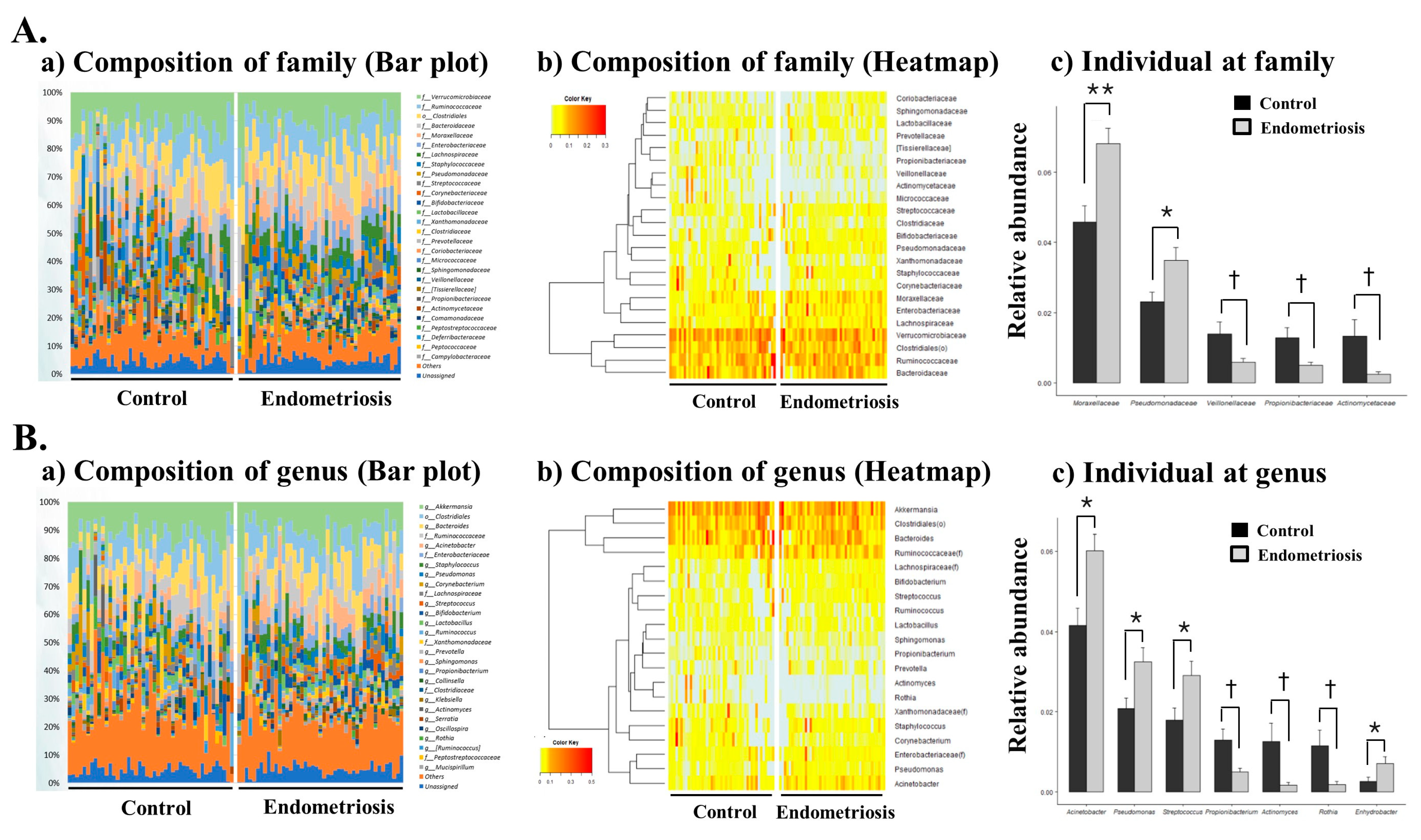
2.3. Microbiome Composition of EVs in PF in Women with and without Endometriosis
3. Discussion
4. Materials and Methods
4.1. Subjects, Collection, and Preparation of PF Samples
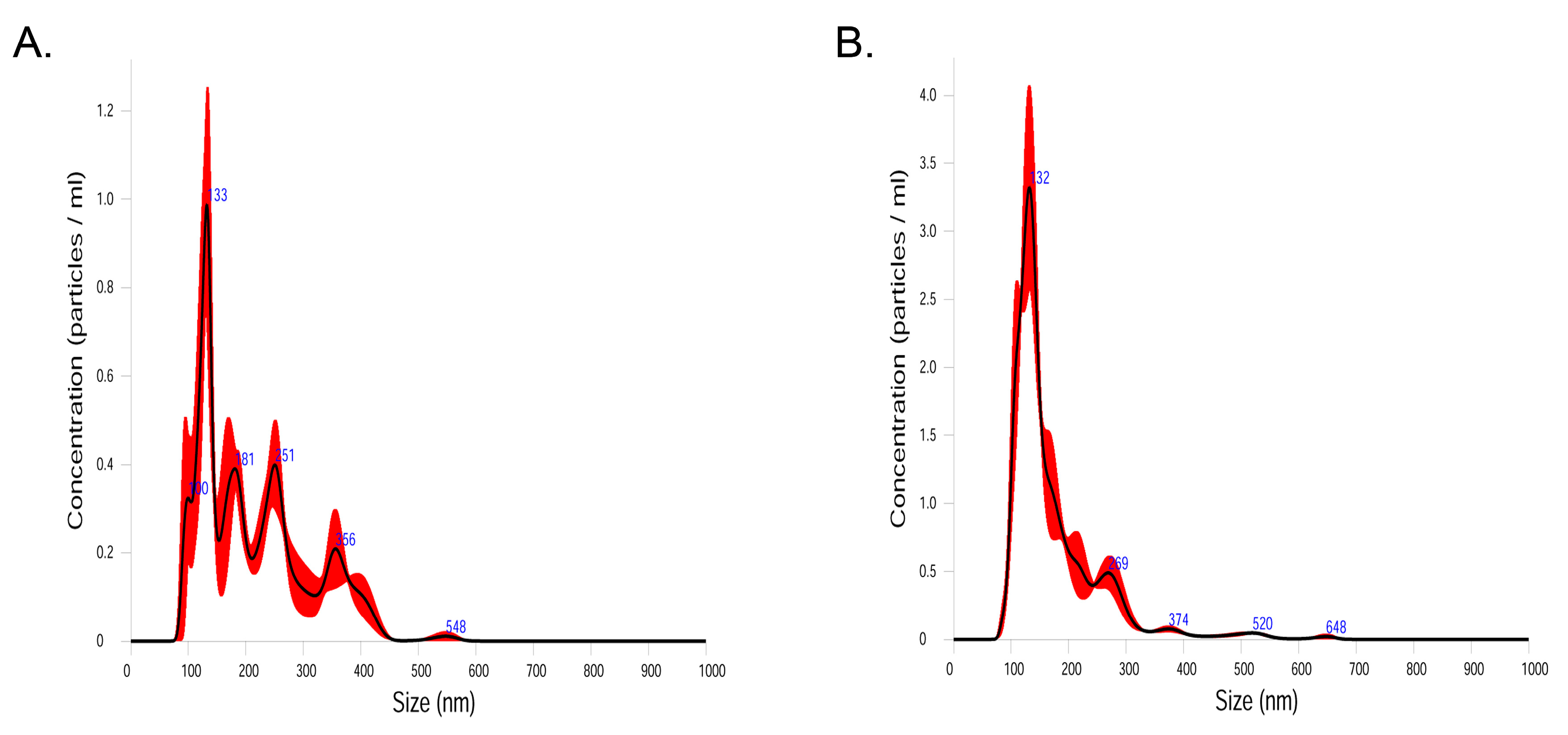
4.2. Isolation of EV and DNA Extraction
4.3. PCR Amplification and Pyrosequencing
4.4. Metagenomic Data Analysis of Human PF Samples
4.5. Analysis of Microbial Composition
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Baldi, A.; Campioni, M.; Signorile, P.G. Endometriosis: Pathogenesis, diagnosis, therapy and association with cancer. Oncol. Rep. 2008, 19, 843–846. [Google Scholar] [CrossRef]
- Bulun, S.E. Endometriosis. N. Engl. J. Med. 2009, 360, 268–279. [Google Scholar] [CrossRef]
- Giudice, L.C.; Kao, L.C. Endometriosis. Lancet 2004, 364, 1789–1799. [Google Scholar] [CrossRef]
- Haas, D.; Chvatal, R.; Reichert, B.; Renner, S.; Shebl, O.; Binder, H.; Wurm, P.; Oppelt, P. Endometriosis: A premenopausal disease? Age pattern in 42,079 patients with endometriosis. Arch. Gynecol. Obstet. 2012, 286, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.; Touloupidis, S.; Papatsoris, A.G.; Triantafyllidis, A.; Kollias, A.; Schweppe, K.W. Endometriosis of the urinary tract in women of reproductive age. Int. J. Urol. 2006, 13, 902–904. [Google Scholar] [CrossRef] [PubMed]
- Practice Committee of the American Society for Reproductive Medicine. Endometriosis and infertility. Fertil. Steril. 2004, 82 (Suppl. 1), S40–S45. [Google Scholar] [CrossRef] [Green Version]
- Burney, R.O.; Giudice, L.C. Pathogenesis and pathophysiology of endometriosis. Fertil. Steril. 2012, 98, 511–519. [Google Scholar] [CrossRef] [Green Version]
- Attar, E.; Bulun, S.E. Aromatase and other steroidogenic genes in endometriosis: Translational aspects. Hum. Reprod Update 2006, 12, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.N.; Kitajima, M.; Hiraki, K.; Fujishita, A.; Sekine, I.; Ishimaru, T.; Masuzaki, H. Immunopathogenesis of pelvic endometriosis: Role of hepatocyte growth factor, macrophages and ovarian steroids. Am. J. Reprod. Immunol. 2008, 60, 383–404. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishna, B.S. Role of the gut microbiota in human nutrition and metabolism. J. Gastroenterol. Hepatol. 2013, 28 (Suppl. 4), 9–17. [Google Scholar] [CrossRef]
- Schirbel, A.; Kessler, S.; Rieder, F.; West, G.; Rebert, N.; Asosingh, K.; McDonald, C.; Fiocchi, C. Pro-angiogenic activity of TLRs and NLRs: A novel link between gut microbiota and intestinal angiogenesis. Gastroenterology 2013, 144, 613–623.e9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.J.; Li, S.; Gan, R.Y.; Zhou, T.; Xu, D.P.; Li, H.B. Impacts of gut bacteria on human health and diseases. Int. J. Mol. Sci. 2015, 16, 7493–7519. [Google Scholar] [CrossRef]
- Costello, M.E.; Robinson, P.C.; Benham, H.; Brown, M.A. The intestinal microbiome in human disease and how it relates to arthritis and spondyloarthritis. Best Pract. Res. Clin. Rheumatol. 2015, 29, 202–212. [Google Scholar] [CrossRef]
- Khan, K.N.; Fujishita, A.; Hiraki, K.; Kitajima, M.; Nakashima, M.; Fushiki, S.; Kitawaki, J. Bacterial contamination hypothesis: A new concept in endometriosis. Reprod. Med. Biol. 2018, 17, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Khan, K.N.; Fujishita, A.; Kitajima, M.; Hiraki, K.; Nakashima, M.; Masuzaki, H. Intra-uterine microbial colonization and occurrence of endometritis in women with endometriosis. Hum. Reprod. 2014, 29, 2446–2456. [Google Scholar] [CrossRef] [Green Version]
- Khan, K.N.; Fujishita, A.; Masumoto, H.; Muto, H.; Kitajima, M.; Masuzaki, H.; Kitawaki, J. Molecular detection of intrauterine microbial colonization in women with endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 199, 69–75. [Google Scholar] [CrossRef]
- Khan, K.N.; Kitajima, M.; Hiraki, K.; Yamaguchi, N.; Katamine, S.; Matsuyama, T.; Nakashima, M.; Fujishita, A.; Ishimaru, T.; Masuzaki, H. Escherichia coli contamination of menstrual blood and effect of bacterial endotoxin on endometriosis. Fertil. Steril. 2010, 94, 2860–2863.e1-3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, M.T.; Coe, C.L. Endometriosis is associated with an altered profile of intestinal microflora in female rhesus monkeys. Hum. Reprod. 2002, 17, 1704–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ata, B.; Yildiz, S.; Turkgeldi, E.; Brocal, V.P.; Dinleyici, E.C.; Moya, A.; Urman, B. The Endobiota Study: Comparison of vaginal, cervical and gut microbiota between women with stage 3/4 endometriosis and healthy controls. Sci. Rep. 2019, 9, 2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyofuku, M.; Nomura, N.; Eberl, L. Types and origins of bacterial membrane vesicles. Nat. Rev. Microbiol. 2019, 17, 13–24. [Google Scholar] [CrossRef]
- Macia, L.; Nanan, R.; Hosseini-Beheshti, E.; Grau, G.E. Host- and microbiota-derived extracellular vesicles, immune function, and disease development. Int. J. Mol. Sci. 2020, 21, 107. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the wall: Extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat. Rev. Microbiol. 2015, 13, 620–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Park, H.; Park, H.S.; Kim, Y.K. Extracellular vesicles, a key mediator to link environmental microbiota to airway immunity. Allergy Asthma Immunol. Res. 2017, 9, 101–106. [Google Scholar] [CrossRef]
- Dagnelie, M.A.; Corvec, S.; Khammari, A.; Dréno, B. Bacterial extracellular vesicles: A new way to decipher host-microbiota communications in inflammatory dermatoses. Exp. Dermatol. 2020, 29, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Kwon, Y.; Kim, D.K.; Jeon, J.; Jang, S.C.; Wang, T.; Ban, M.; Kim, M.H.; Jeon, S.G.; Kim, M.S.; et al. Gut microbe-derived extracellular vesicles induce insulin resistance, thereby impairing glucose metabolism in skeletal muscle. Sci. Rep. 2015, 5, 15878. [Google Scholar] [CrossRef] [Green Version]
- Khalaj, K.; Miller, J.E.; Lingegowda, H.; Fazleabas, A.T.; Young, S.L.; Lessey, B.A.; Koti, M.; Tayade, C. Extracellular vesicles from endometriosis patients are characterized by a unique miRNA-lncRNA signature. JCI Insight 2019, 4, e128846. [Google Scholar] [CrossRef]
- Nazri, H.M.; Imran, M.; Fischer, R.; Heilig, R.; Manek, S.; Dragovic, R.A.; Kessler, B.M.; Zondervan, K.T.; Tapmeier, T.T.; Becker, C.M. Characterization of exosomes in peritoneal fluid of endometriosis patients. Fertil. Steril. 2020, 113, 364–373.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Song, X.; Wei, W.; Zhong, H.; Dai, J.; Lan, Z.; Li, F.; Yu, X.; Feng, Q.; Wang, Z.; et al. The microbiota continuum along the female reproductive tract and its relation to uterine-related diseases. Nat. Commun. 2017, 8, 875. [Google Scholar] [CrossRef] [Green Version]
- Winters, A.D.; Romero, R.; Gervasi, M.T.; Gomez-Lopez, N.; Tran, M.R.; Flores, V.G.; Pacora, P.; Jung, E.; Hassan, S.S.; Hsu, C.D.; et al. Does the endometrial cavity have a molecular microbial signature? Sci. Rep. 2019, 9, 9905. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Zhang, X.; Tang, H.; Zeng, L.; Wu, R. Microbiota composition and distribution along the female reproductive tract of women with endometriosis. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandes, C.; Silveira, P.; Rodrigues Sereia, A.F.; Christoff, A.P.; Mendes, H.; Valter de Oliveira, L.F.; Podgaec, S. Microbiome profile of deep endometriosis patients: Comparison of vaginal fluid, endometrium and lesion. Diagnostics 2020, 10, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.N.; Kitajima, M.; Yamaguchi, N.; Fujishita, A.; Nakashima, M.; Ishimaru, T.; Masuzaki, H. Role of prostaglandin E2 in bacterial growth in women with endometriosis. Hum. Reprod. 2012, 27, 3417–3424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revised American Society for Reproductive Medicine classification of endometriosis: 1996. Fertil. Steril. 1997, 67, 817–821. [CrossRef]
- Navas-Molina, J.A.; Peralta-Sánchez, J.M.; González, A.; McMurdie, P.J.; Vázquez-Baeza, Y.; Xu, Z.; Ursell, L.K.; Lauber, C.; Zhou, H.; Song, S.J.; et al. Advancing our understanding of the human microbiome using QIIME. Methods Enzymol. 2013, 531, 371–444. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Endometriosis | t-test | |||
|---|---|---|---|---|---|
| Taxon | Mean | SD | Mean | SD | p-Value |
| Phylum | |||||
| Actinobacteria | 0.1107 | 0.0815 | 0.0831 | 0.0394 | 0.0477 |
| Class | |||||
| Actinobacteria | 0.1004 | 0.0784 | 0.0705 | 0.0368 | 0.0255 |
| Order | |||||
| Pseudomonadales | 0.0805 | 0.0606 | 0.1037 | 0.0411 | 0.0387 |
| Actinomycetales | 0.0811 | 0.0792 | 0.0429 | 0.0284 | 0.0039 |
| Family | |||||
| Moraxellaceae | 0.0459 | 0.0306 | 0.0682 | 0.0298 | 0.0008 |
| Pseudomonadaceae | 0.0230 | 0.0189 | 0.0348 | 0.0244 | 0.0128 |
| Veillonellaceae | 0.0139 | 0.0227 | 0.0058 | 0.0069 | 0.0282 |
| Propionibacteriaceae | 0.0129 | 0.0190 | 0.0049 | 0.0062 | 0.0110 |
| Actinomycetaceae | 0.0133 | 0.0309 | 0.0023 | 0.0058 | 0.0243 |
| Genus | |||||
| Acinetobacter | 0.0416 | 0.0289 | 0.0602 | 0.0265 | 0.0022 |
| Pseudomonas | 0.0208 | 0.0175 | 0.0325 | 0.0235 | 0.0097 |
| Propionibacterium | 0.0129 | 0.0190 | 0.0049 | 0.0062 | 0.0111 |
| Streptococcus | 0.0179 | 0.0204 | 0.0291 | 0.0231 | 0.0183 |
| Rothia | 0.0115 | 0.0261 | 0.0018 | 0.0045 | 0.0190 |
| Actinomyces | 0.0126 | 0.0305 | 0.0017 | 0.0043 | 0.0233 |
| Enhydrobacter | 0.0026 | 0.0072 | 0.0071 | 0.0114 | 0.0300 |
| Endometriosis Group (n = 45) | Control Group (n = 45) | p-Value | |
|---|---|---|---|
| Age | 36.2 ± 1.3 | 39.4 ± 1.1 | 0.06 a |
| No. of deliveries | 0.7 ± 0.1 | 1.3 ± 0.1 | <0.01 a |
| Body mass index (kg/m2) Married women (%) Infertility (%) AFS classification of endometriosis | 21.0 ± 0.5 25 (55.6%) 2 (4.4%) | 22.2 ± 0.5 32 (71%) 0 | 0.18 a 0.16 b 0.15 b |
| Stage III | 34 | ||
| Stage IV | 11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-R.; Lee, J.-C.; Kim, S.-H.; Oh, Y.-S.; Chae, H.-D.; Seo, H.; Kang, C.-S.; Shin, T.-S. Altered Composition of Microbiota in Women with Ovarian Endometrioma: Microbiome Analyses of Extracellular Vesicles in the Peritoneal Fluid. Int. J. Mol. Sci. 2021, 22, 4608. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094608
Lee S-R, Lee J-C, Kim S-H, Oh Y-S, Chae H-D, Seo H, Kang C-S, Shin T-S. Altered Composition of Microbiota in Women with Ovarian Endometrioma: Microbiome Analyses of Extracellular Vesicles in the Peritoneal Fluid. International Journal of Molecular Sciences. 2021; 22(9):4608. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094608
Chicago/Turabian StyleLee, Sa-Ra, Jae-Chul Lee, Sung-Hoon Kim, Young-Sang Oh, Hee-Dong Chae, Hochan Seo, Chil-Sung Kang, and Tae-Seop Shin. 2021. "Altered Composition of Microbiota in Women with Ovarian Endometrioma: Microbiome Analyses of Extracellular Vesicles in the Peritoneal Fluid" International Journal of Molecular Sciences 22, no. 9: 4608. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094608