
From Channels to Canonical Wnt Signaling: A Pathological Perspective

 , ,
, ,
Abstract
:1. Introduction
2. Molecular Interplay between Ions, Channels and Wnt Signaling
3. Channels-Wnt-Dependent Signaling Axes in Diseases
3.1. Channels, Wnt Signaling, and Cancer
3.2. Ion Channels and Wnt Signaling Regulation in Neurodegeneration and Neurological Diseases
3.3. Channels, Wnt Signaling in Other Diseases
4. Conclusions and Future Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Sharma, M.; Pruitt, K. Wnt Pathway: An Integral Hub for Developmental and Oncogenic Signaling Networks. Int. J. Mol. Sci. 2020, 21, 8018. [Google Scholar] [CrossRef]
- Wodarz, A.; Nusse, R. Mechanisms of Wnt signaling in development. Annu. Rev. Cell Dev. Biol. 1998, 14, 59–88. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, B.T.; He, X. Frizzled and LRp5/6 receptors for wnt/β-catenin signaling. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef]
- Fiedler, M.; Mendoza-Topaz, C.; Rutherford, T.J.; Mieszczanek, J.; Bienz, M. Dishevelled interacts with the DIX domain polymerization interface of Axin to interfere with its function in down-regulating β-catenin. Proc. Natl. Acad. Sci. USA 2011, 108, 1937–1942. [Google Scholar] [CrossRef] [Green Version]
- Taelman, V.F.; Dobrowolski, R.; Plouhinec, J.-L.; Fuentealba, L.C.; Vorwald, P.P.; Gumper, I.; Sabatini, D.D.; De Robertis, E.M. Wnt Signaling Requires Sequestration of Glycogen Synthase Kinase 3 inside Multivesicular Endosomes. Cell 2010, 143, 1136–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Kato, Y.; Zhang, Z.; Do, V.M.; Yankner, B.A.; He, X. -Trcp couples -catenin phosphorylation-degradation and regulates Xenopus axis formation. Proc. Natl. Acad. Sci. USA 1999, 96, 6273–6278. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, S.; Kishida, S.; Yamamoto, H.; Murai, H.; Koyama, S.; Kikuchi, A. Axin, a negative regulator of the Wnt signaling pathway, forms a complex with GSK-3β and β-catenin and promotes GSK-3β-dependent phosphorylation of β-catenin. EMBO J. 1998, 17. [Google Scholar] [CrossRef]
- Daniels, D.L.; I Weis, W. β-catenin directly displaces Groucho/TLE repressors from Tcf/Lef in Wnt-mediated transcription activation. Nat. Struct. Mol. Biol. 2005, 12, 364–371. [Google Scholar] [CrossRef]
- Cavallo, R.A.; Cox, R.T.; Moline, M.M.; Roose, J.; Polevoy, G.A.; Clevers, H.; Peifer, M.; Bejsovec, A. Drosophila Tcf and Groucho interact to repress Wingless signalling activity. Nat. Cell Biol. 1998, 395, 604–608. [Google Scholar] [CrossRef]
- Song, H.; Goetze, S.; Bischof, J.; Spichiger-Haeusermann, C.; Küster, M.; Brunner, E.; Basler, K. Coop functions as a corepressor of Pangolin and antagonizes Wingless signaling. Genes Dev. 2010, 24, 881–886. [Google Scholar] [CrossRef] [Green Version]
- Billin, A.N.; Thirlwell, H.; Ayer, D.E. β-Catenin–Histone Deacetylase Interactions Regulate the Transition of LEF1 from a Transcriptional Repressor to an Activator. Mol. Cell. Biol. 2000, 20, 6882–6890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Xu, X.; Hecht, A.; Boyer, T.G. Mediator Is a Transducer of Wnt/β-Catenin Signaling. J. Biol. Chem. 2006, 281, 14066–14075. [Google Scholar] [CrossRef] [Green Version]
- Che, M.; Kweon, S.-M.; Teo, J.-L.; Yuan, Y.-C.; Melstrom, L.G.; Waldron, R.T.; Lugea, A.; Urrutia, R.A.; Pandol, S.J.; Lai, K.K.Y. Targeting the CBP/β-Catenin Interaction to Suppress Activation of Cancer-Promoting Pancreatic Stellate Cells. Cancers 2020, 12, 1476. [Google Scholar] [CrossRef]
- Anthony, C.C.; Robbins, D.J.; Ahmed, Y.; Lee, E. Nuclear Regulation of Wnt/β-Catenin Signaling: It’s a Complex Situation. Genes 2020, 11, 886. [Google Scholar] [CrossRef]
- Majmundar, A.J.; Lee, D.S.M.; Skuli, N.; Mesquita, R.C.; Kim, M.N.; Yodh, A.G.; Nguyen-McCarty, M.; Li, B.; Simon, M.C. HIF modulation of Wnt signaling regulates skeletal myogenesis in vivo. Development 2015, 142. [Google Scholar] [CrossRef] [Green Version]
- Kaidi, A.; Williams, A.C.; Paraskeva, C. Interaction between β-catenin and HIF-1 promotes cellular adaptation to hypoxia. Nat. Cell Biol. 2007, 9, 210–217. [Google Scholar] [CrossRef]
- Sinner, D.; Kordich, J.J.; Spence, J.R.; Opoka, R.; A Rankin, S.; Lin, S.-C.J.; Jonatan, D.; Zorn, A.M.; Wells, J.M. Sox17 and Sox4 Differentially Regulate β-Catenin/T-Cell Factor Activity and Proliferation of Colon Carcinoma Cells. Mol. Cell. Biol. 2007, 27, 7802–7815. [Google Scholar] [CrossRef] [Green Version]
- Sorrenson, B.; Cognard, E.; Lee, K.L.; Dissanayake, W.C.; Fu, Y.; Han, W.; Hughes, W.E.; Shepherd, P.R. A Critical Role for β-Catenin in Modulating Levels of Insulin Secretion from β-Cells by Regulating Actin Cytoskeleton and Insulin Vesicle Localization. J. Biol. Chem. 2016, 291, 25888–25900. [Google Scholar] [CrossRef] [Green Version]
- Colozza, G.; Koo, B. Wnt/β-catenin signaling: Structure, assembly and endocytosis of the signalosome. Dev. Growth Differ. 2021, 63, 199–218. [Google Scholar] [CrossRef]
- Lorzadeh, S.; Kohan, L.; Ghavami, S.; Azarpira, N. Autophagy and the Wnt signaling pathway: A focus on Wnt/β-catenin signaling. Biochim. Biophys. Acta BBA Bioenerg. 2021, 1868, 118926. [Google Scholar] [CrossRef]
- Schneider, S.; Herrenknecht, K.; Butz, S.; Kemler, R.; Hausen, P. Catenins in Xenopus embryogenesis and their relation to the cadherin-mediated cell-cell adhesion system. Development 1993, 118, 629–640. [Google Scholar] [CrossRef]
- Kemler, R. From cadherins to catenins: Cytoplasmic protein interactions and regulation of cell adhesion. Trends Genet. 1993, 9, 317–321. [Google Scholar] [CrossRef]
- Huels, D.J.; Ridgway, R.A.; Radulescu, S.; Leushacke, M.; Campbell, A.D.; Biswas, S.; Leedham, S.; Serra, S.; Chetty, R.; Moreaux, G.; et al. E-cadherin can limit the transforming properties of activating β-catenin mutations. EMBO J. 2015, 34. [Google Scholar] [CrossRef]
- Martin-Orozco, E.; Sanchez-Fernandez, A.; Ortiz-Parra, I.; Nicolas, M.A.-S. WNT Signaling in Tumors: The Way to Evade Drugs and Immunity. Front. Immunol. 2019, 10, 2854. [Google Scholar] [CrossRef]
- Zancan, I.; Bellesso, S.; Costa, R.; Salvalaio, M.; Stroppiano, M.; Hammond, C.; Argenton, F.; Filocamo, M.; Moro, E. Glucocerebrosidase deficiency in zebrafish affects primary bone ossification through increased oxidative stress and reduced Wnt/β-catenin signaling. Hum. Mol. Genet. 2014, 24, 1280–1294. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, Y.P.; Paulson, C.; Shao, J.Z.; Zhang, X.; Wu, M.; Chen, W. Wnt and the Wnt signaling pathway in bone development and disease. Front. Biosci. Landmark 2014, 19, 379. [Google Scholar] [CrossRef] [Green Version]
- Berwick, D.C.; Harvey, K. The importance of Wnt signalling for neurodegeneration in Parkinson’s disease. Biochem. Soc. Trans. 2012, 40, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Peruzzo, R.; Costa, R.; Bachmann, M.; Leanza, L.; Szabò, I. Mitochondrial Metabolism, Contact Sites and Cellular Calcium Signaling: Implications for Tumorigenesis. Cancers 2020, 12, 2574. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.E.; Nourse, C.C.; Cooper, H.M. The Tangled Web of Non-Canonical Wnt Signalling in Neural Migration. Neurosignals 2012, 20, 202–220. [Google Scholar] [CrossRef]
- Yang, K.; Wang, X.; Zhang, H.; Wang, Z.; Nan, G.; Li, Y.; Zhang, F.; Mohammed, M.K.; Haydon, R.C.; Luu, H.H.; et al. The evolving roles of canonical WNT signaling in stem cells and tumorigenesis: Implications in targeted cancer therapies. Lab. Investig. 2016, 96, 116–136. [Google Scholar] [CrossRef] [Green Version]
- Herbst, A.; Jurinovic, V.; Krebs, S.; E Thieme, S.; Blum, H.; Göke, B.; Kolligs, F.T. Comprehensive analysis of β-catenin target genes in colorectal carcinoma cell lines with deregulated Wnt/β-catenin signaling. BMC Genom. 2014, 15, 74. [Google Scholar] [CrossRef] [Green Version]
- Montalto, F.I.; De Amicis, F. Cyclin D1 in Cancer: A Molecular Connection for Cell Cycle Control, Adhesion and Invasion in Tumor and Stroma. Cells 2020, 9, 2648. [Google Scholar] [CrossRef]
- Wong, M.; Joyson, S.; Hermeking, H.; Chiu, S. Transcription Factor AP4 Mediates Cell Fate Decisions: To Divide, Age, or Die. Cancers 2021, 13, 676. [Google Scholar] [CrossRef]
- Clapham, D.E. Calcium Signaling. Cell 2011, 131, 1047–1058. [Google Scholar] [CrossRef] [Green Version]
- Thrasivoulou, C.; Millar, M.; Ahmed, A. Activation of Intracellular Calcium by Multiple Wnt Ligands and Translocation of β-Catenin into the Nucleus. J. Biol. Chem. 2013, 288, 35651–35659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, R.; Peruzzo, R.; Bachmann, M.; Montà, G.D.; Vicario, M.; Santinon, G.; Mattarei, A.; Moro, E.; Quintana-Cabrera, R.; Scorrano, L.; et al. Impaired Mitochondrial ATP Production Downregulates Wnt Signaling via ER Stress Induction. Cell Rep. 2019, 28, 1949–1960.e6. [Google Scholar] [CrossRef] [Green Version]
- Horndasch, M.; Lienkamp, S.; Springer, E.; Schmitt, A.; Pavenstädt, H.; Walz, G.; Gloy, J. The C/EBP homologous protein CHOP (GADD153) is an inhibitor of Wnt/TCF signals. Oncogene 2006, 25. [Google Scholar] [CrossRef]
- Zhang, Q.-L.; Lian, D.-D.; Zhu, M.J.; Li, X.M.; Lee, J.K.; Yoon, T.-J.; Lee, J.-H.; Jiang, R.-H.; Kim, C.D. Antitumor Effect of Albendazole on Cutaneous Squamous Cell Carcinoma (SCC) Cells. BioMed Res. Int. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaccagnino, A.; Managò, A.; Leanza, L.; Gontarewitz, A.; Linder, B.; Azzolini, M.; Biasutto, L.; Zoratti, M.; Peruzzo, R.; Legler, K.; et al. Tumor-reducing effect of the clinically used drug clofazimine in a SCID mouse model of pancreatic ductal adenocarcinoma. Oncotarget 2016, 8, 38276–38293. [Google Scholar] [CrossRef] [PubMed]
- Koval, A.; Vlasov, P.; Shichkova, P.; Khunderyakova, S.; Markov, Y.; Panchenko, J.; Volodina, A.; Kondrashov, F.; Katanaev, V. Anti-leprosy drug clofazimine inhibits growth of triple-negative breast cancer cells via inhibition of canonical Wnt signaling. Biochem. Pharmacol. 2014, 87, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Breuer, E.-K.; Fukushiro-Lopes, D.; Dalheim, A.; Burnette, M.; Zartman, J.; Kaja, S.; Wells, C.; Campo, L.; Curtis, K.J.; Romero-Moreno, R.; et al. Potassium channel activity controls breast cancer metastasis by affecting β-catenin signaling. Cell Death Dis. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Manago, A.; Leanza, L.; Carraretto, L.; Sassi, N.; Grancara, S.; Quintanacabrera, R.; Trimarco, V.; Toninello, A.; Scorrano, L.; Trentin, L.; et al. Early effects of the antineoplastic agent salinomycin on mitochondrial function. Cell Death Dis. 2015, 6, e1930. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Choi, M.Y.; Yu, J.; Castro, J.E.; Kipps, T.J.; Carson, D.A. Salinomycin inhibits Wnt signaling and selectively induces apoptosis in chronic lymphocytic leukemia cells. Proc. Natl. Acad. Sci. USA 2011, 108, 13253–13257. [Google Scholar] [CrossRef] [Green Version]
- Hers, I.; Vincent, E.E.; Tavaré, J.M. Akt signalling in health and disease. Cell. Signal. 2011, 23, 1515–1527. [Google Scholar] [CrossRef]
- Divolis, G.; Mavroeidi, P.; Mavrofrydi, O.; Papazafiri, P. Differential effects of calcium on PI3K-Akt and HIF-1α survival pathways. Cell Biol. Toxicol. 2016, 32, 437–449. [Google Scholar] [CrossRef]
- Fang, D.; Hawke, D.; Zheng, Y.; Xia, Y.; Meisenhelder, J.; Nika, H.; Mills, G.B.; Kobayashi, R.; Hunter, T.; Lu, Z. Phosphorylation of β-Catenin by AKT Promotes β-Catenin Transcriptional Activity. J. Biol. Chem. 2007, 282, 11221–11229. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Weiling, W.; Sun, Y.; Lei, L.; Zhou, H.; Lei, T.; Xia, Y.; Verkman, A.S.; Yang, B. Aquaporin-1 retards renal cyst development in polycystic kidney disease by inhibition of Wnt signaling. FASEB J. 2015, 29, 1551–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meli, R.; Pirozzi, C.; Pelagalli, A. New Perspectives on the Potential Role of Aquaporins (AQPs) in the Physiology of Inflammation. Front. Physiol. 2018, 9, 101. [Google Scholar] [CrossRef] [Green Version]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Ion Channels in Cancer: Are Cancer Hallmarks Oncochannelopathies? Physiol. Rev. 2018, 98, 559–621. [Google Scholar] [CrossRef] [Green Version]
- Rapetti-Mauss, R.; Berenguier, C.; Allegrini, B.; Soriani, O. Interplay Between Ion Channels and the Wnt/β-Catenin Signaling Pathway in Cancers. Front. Pharmacol. 2020, 11, 525020. [Google Scholar] [CrossRef]
- Rapetti-Mauss, R.; Bustos, V.; Thomas, W.; McBryan, J.; Harvey, H.; Lajczak, N.; Madden, S.F.; Pellissier, B.; Borgese, F.; Soriani, O.; et al. Bidirectional KCNQ1: β-catenin interaction drives colorectal cancer cell differentiation. Proc. Natl. Acad. Sci. USA 2017, 114, 4159–4164. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Zhang, M.; Liu, W. Hypermethylated KCNQ1 acts as a tumor suppressor in hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2018, 503, 3100–3107. [Google Scholar] [CrossRef]
- Strubberg, A.M.; Liu, J.; Walker, N.M.; Stefanski, C.D.; MacLeod, R.J.; Magness, S.T.; Clarke, L.L. Cftr Modulates Wnt/β-Catenin Signaling and Stem Cell Proliferation in Murine Intestine. Cell. Mol. Gastroenterol. Hepatol. 2018, 5, 253–271. [Google Scholar] [CrossRef] [Green Version]
- Xie, R.; Xu, J.; Xiao, Y.; Wu, J.; Wan, H.; Tang, B.; Liu, J.; Fan, Y.; Wang, S.; Wu, Y.; et al. Calcium Promotes Human Gastric Cancer via a Novel Coupling of Calcium-Sensing Receptor and TRPV4 Channel. Cancer Res. 2017, 77, 6499–6512. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Chen, Z.; Zhu, Y.; Pan, Q.; Liu, Y.; Qi, X.; Jin, L.; Jin, J.; Ma, X.; Hua, D. Inhibition of Transient Receptor Potential Channel 5 Reverses 5-Fluorouracil Resistance in Human Colorectal Cancer Cells. J. Biol. Chem. 2015, 290, 448–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, A.M.G.; Kong, K.L.; Chen, L.; Liu, M.; Wong, A.M.G.; Zhu, C.; Tsang, J.W.-H.; Guan, X.-Y. Characterization ofCACNA2D3as a putative tumor suppressor gene in the development and progression of nasopharyngeal carcinoma. Int. J. Cancer 2013, 133, 2284–2295. [Google Scholar] [CrossRef]
- Jin, Y.; Cui, D.; Ren, J.; Wang, K.; Zeng, T.; Gao, L. CACNA2D3 is downregulated in gliomas and functions as a tumor suppressor. Mol. Carcinog. 2017, 56, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Than, B.L.N.; Linnekamp, J.F.; Starr, T.K.; Largaespada, D.A.; Rod, A.; Zhang, Y.; Bruner, V.; Abrahante, J.; Schumann, A.; Luczak, T.; et al. CFTR is a tumor suppressor gene in murine and human intestinal cancer. Oncogene 2016, 35. [Google Scholar] [CrossRef]
- Chen, Z.; Tang, C.; Zhu, Y.; Xie, M.; He, D.; Pan, Q.; Zhang, P.; Hua, D.; Wang, T.; Jin, L.; et al. TrpC5 regulates differentiation through the Ca2+/Wnt5a signalling pathway in colorectal cancer. Clin. Sci. 2017, 131, 227–237. [Google Scholar] [CrossRef]
- Sagredo, A.I.; Sagredo, E.A.; Cappelli, C.; Báez, P.; Andaur, R.E.; Blanco, C.; Tapia, J.C.; Echeverría, C.; Cerda, O.; Stutzin, A.; et al. TRPM4 regulates Akt/GSK3-β activity and enhances β-catenin signaling and cell proliferation in prostate cancer cells. Mol. Oncol. 2017, 12, 151–165. [Google Scholar] [CrossRef] [Green Version]
- Kudou, M.; Shiozaki, A.; Yamazato, Y.; Katsurahara, K.; Kosuga, T.; Shoda, K.; Arita, T.; Konishi, H.; Komatsu, S.; Kubota, T.; et al. The expression and role of TRPV2 in esophageal squamous cell carcinoma. Sci. Rep. 2019, 9, 16055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schickling, B.M.; England, S.K.; Aykin-Burns, N.; Norian, L.A.; Leslie, K.K.; Frieden-Korovkina, V.P. BKCa channel inhibitor modulates the tumorigenic ability of hormone-independent breast cancer cells via the Wnt pathway. Oncol. Rep. 2014, 33, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Boassa, D.; Stamer, W.D.; Yool, A.J. Ion Channel Function of Aquaporin-1 Natively Expressed in Choroid Plexus. J. Neurosci. 2006, 26, 7811–7819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Liu, L.; Zhang, Y.; Jing, H. Molecular mechanism of aquapontin (AQP3) in regulating differentiation and apoptosis of lung cancer stem cells through Wnt/GSK-3β/β-Catenin pathway. J. BUON Off. J. Balk. Union Oncol. 2020, 25, 828–834. [Google Scholar]
- Zhou, Y.; Wang, Y.; Wen, J.; Zhao, H.; Dong, X.; Zhang, Z.; Wang, S.; Shen, L. Aquaporin 3 promotes the stem-like properties of gastric cancer cells via Wnt/GSK-3β/β-catenin pathway. Oncotarget 2016, 7, 16529–16541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, S.; Chen, H.; Liu, M.; Gan, L.; Li, C.; Zhang, W.; Lv, L.; Mei, Z. Aquaporin 9 inhibits growth and metastasis of hepatocellular carcinoma cells via Wnt/β-catenin pathway. Aging 2020, 12, 1527–1544. [Google Scholar] [CrossRef]
- Tan, S.H.; Swathi, Y.; Tan, S.; Goh, J.; Seishima, R.; Murakami, K.; Oshima, M.; Tsuji, T.; Phuah, P.; Tan, L.T.; et al. AQP5 enriches for stem cells and cancer origins in the distal stomach. Nat. Cell Biol. 2020, 578, 437–443. [Google Scholar] [CrossRef]
- Leanza, L.; Zoratti, M.; Gulbins, E.; Szabo, I.L. Mitochondrial ion channels as oncological targets. Oncogene 2014, 33, 5569–5581. [Google Scholar] [CrossRef] [Green Version]
- Leanza, L.; Romio, M.; Becker, K.A.; Azzolini, M.; Trentin, L.; Manago, A.; Venturini, E.; Zaccagnino, A.; Mattarei, A.; Carraretto, L.; et al. Direct Pharmacological Targeting of a Mitochondrial Ion Channel Selectively Kills Tumor Cells In Vivo. Cancer Cell 2017, 31, 516–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Chen, Y.; Zhang, Q.; Li, C. Silencing of KCNA1 suppresses the cervical cancer development via mitochondria damage. Channels 2019, 13, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Suisse, A.; Treisman, J.E. Reduced SERCA Function Preferentially Affects Wnt Signaling by Retaining E-Cadherin in the Endoplasmic Reticulum. Cell Rep. 2019, 26, 322–329.e3. [Google Scholar] [CrossRef] [Green Version]
- Serafino, A.; Giovannini, D.; Rossi, S.; Cozzolino, M. Targeting the Wnt/β-catenin pathway in neurodegenerative diseases: Recent approaches and current challenges. Expert Opin. Drug Discov. 2020, 15, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Inestrosa, N.C.; Tapia-Rojas, C. Loss of canonical Wnt signaling is involved in the pathogenesis of Alzheimer’s disease. Neural Regen. Res. 2018, 13, 1705–1710. [Google Scholar] [CrossRef]
- Aghaizu, N.D.; Jin, H.; Whiting, P.J. Dysregulated Wnt Signalling in the Alzheimer’s Brain. Brain Sci. 2020, 10, 902. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.-Z.; Li, B.; Li, Y.-C.; Yang, X.-L.; Zhang, W.; Zhong, L.; Tang, S.-J. Activation of NMDA Receptors Upregulates A Disintegrin and Metalloproteinase 10 via a Wnt/MAPK Signaling Pathway. J. Neurosci. 2012, 32, 3910–3916. [Google Scholar] [CrossRef] [Green Version]
- Stephano, F.; Nolte, S.; Hoffmann, J.; El-Kholy, S.; Von Frieling, J.; Bruchhaus, I.; Fink, C.; Roeder, T. Impaired Wnt signaling in dopamine containing neurons is associated with pathogenesis in a rotenone triggered Drosophila Parkinson’s disease model. Sci. Rep. 2018, 8, 2372. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, B. Wnt/β-Catenin Signaling Pathway Governs a Full Program for Dopaminergic Neuron Survival, Neurorescue and Regeneration in the MPTP Mouse Model of Parkinson’s Disease. Int. J. Mol. Sci. 2018, 19, 3743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, C.; Medinas, D.B.; Fuentes-Villalobos, F.; Maripillán, J.; Castro, A.F.; Martínez, A.D.; Osses, N.; Hetz, C.; Henríquez, J.P. β-catenin aggregation in models of ALS motor neurons: GSK3β inhibition effect and neuronal differentiation. Neurobiol. Dis. 2019, 130, 104497. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, C.; González, P.; Rodríguez, F. New insights into Wnt signaling alterations in amyotrophic lateral sclerosis: A potential therapeutic target? Neural Regen. Res. 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Inestrosa, N.C.; Godoy, J.A.; Vargas, J.Y.; Arrázola, M.S.; Rios, J.A.; Carvajal, F.J.; Serrano, F.G.; Farías, G.G. Nicotine Prevents Synaptic Impairment Induced by Amyloid-β Oligomers Through α7-Nicotinic Acetylcholine Receptor Activation. NeuroMolecular Med. 2013, 15, 549–569. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Andreasen, J.T.; Arvaniti, M.; Kohlmeier, K.A. Nicotinic Acetylcholine Receptors in the Pathophysiology of Al zheimer’s Disease: The Role of Protein-Protein Interactions in Current and Future Treatment. Curr. Pharm. Des. 2016, 22, 2015–2034. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hao, S.; Yang, B.; Fan, Y.; Qin, X.; Chen, Y.; Hu, J. Wnt/β-catenin signaling plays an essential role in α7 nicotinic receptor-mediated neuroprotection of dopaminergic neurons in a mouse Parkinson’s disease model. Biochem. Pharmacol. 2017, 140, 115–123. [Google Scholar] [CrossRef] [PubMed]
- McQuate, A.; Latorre-Esteves, E.; Barria, A. A Wnt/Calcium Signaling Cascade Regulates Neuronal Excitability and Trafficking of NMDARs. Cell Rep. 2017, 21, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Zhang, J.; Li, H.; Sun, X. Silencing of aquaporin1 activates the Wnt signaling pathway to improve cognitive function in a mouse model of Alzheimer’s disease. Gene 2020, 755, 144904. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, R.; Liu, X.; Zhou, Y.; Qu, C.; Kikuiri, T.; Wang, S.; Zandi, E.; Du, J.; Ambudkar, I.S.; et al. Hydrogen Sulfide Maintains Mesenchymal Stem Cell Function and Bone Homeostasis via Regulation of Ca2+ Channel Sulfhydration. Cell Stem Cell 2014, 15, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Fei, D.; Zhang, Y.; Wu, J.; Zhang, H.; Liu, A.; He, X.; Wang, J.; Li, B.; Wang, Q.; Jin, Y. Cav 1.2 regulates osteogenesis of bone marrow-derived mesenchymal stem cells via canonical Wnt pathway in age-related osteoporosis. Aging Cell 2019, 18, e12967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.J.; Seo, E.; Kim, J.W.; Nam, S.A.; Lee, J.Y.; Jun, J.; Oh, S.; Park, M.; Jho, E.-H.; Yoo, K.H.; et al. TAZ/Wnt-β-catenin/c-MYC axis regulates cystogenesis in polycystic kidney disease. Proc. Natl. Acad. Sci. USA 2020, 117, 29001–29012. [Google Scholar] [CrossRef]
- Harris, P.C.; Torres, V.E. Polycystic Kidney Disease. Annu. Rev. Med. 2009, 60. [Google Scholar] [CrossRef]
- Shu, C.; Shu, Y.; Gao, Y.; Chi, H.; Han, J. Inhibitory effect of AQP1 silencing on adhesion and angiogenesis in ectopic endometrial cells of mice with endometriosis through activating the Wnt signaling pathway. Cell Cycle 2019, 18, 2026–2039. [Google Scholar] [CrossRef]
- Lu, J.; Xu, F.; Zhang, Y.; Lu, H.; Zhang, J. ClC-2 knockdown prevents cerebrovascular remodeling via inhibition of the Wnt/β-catenin signaling pathway. Cell. Mol. Biol. Lett. 2018, 23, 29. [Google Scholar] [CrossRef] [Green Version]
- Middelbeek, J.; Visser, D.; Henneman, L.; Kamermans, A.; Kuipers, A.J.; Hoogerbrugge, P.M.; Jalink, K.; Van Leeuwen, F.N. TRPM7 maintains progenitor-like features of neuroblastoma cells: Implications for metastasis formation. Oncotarget 2015, 6, 8760–8776. [Google Scholar] [CrossRef]
- Xu, X.; Dai, Y.; Feng, L.; Zhang, H.; Hu, Y.; Xu, L.; Zhu, X.; Jiang, Y. Knockdown of Nav1.5 inhibits cell proliferation, migration and invasion via Wnt/β-catenin signaling pathway in oral squamous cell carcinoma. Acta Biochim. Biophys. Sin. 2020, 52, 527–535. [Google Scholar] [CrossRef]
- Mu, H.; Mu, L.; Gao, J. Suppression of CLC-3 reduces the proliferation, invasion and migration of colorectal cancer through Wnt/β-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2020, 533, 1240–1246. [Google Scholar] [CrossRef]
- Li, X.; Hu, W.; Zhou, J.; Huang, Y.; Peng, J.; Yuan, Y.; Yu, J.; Zheng, S. CLCA1 suppresses colorectal cancer aggressiveness via inhibition of the Wnt/beta-catenin signaling pathway. Cell Commun. Signal. 2017, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
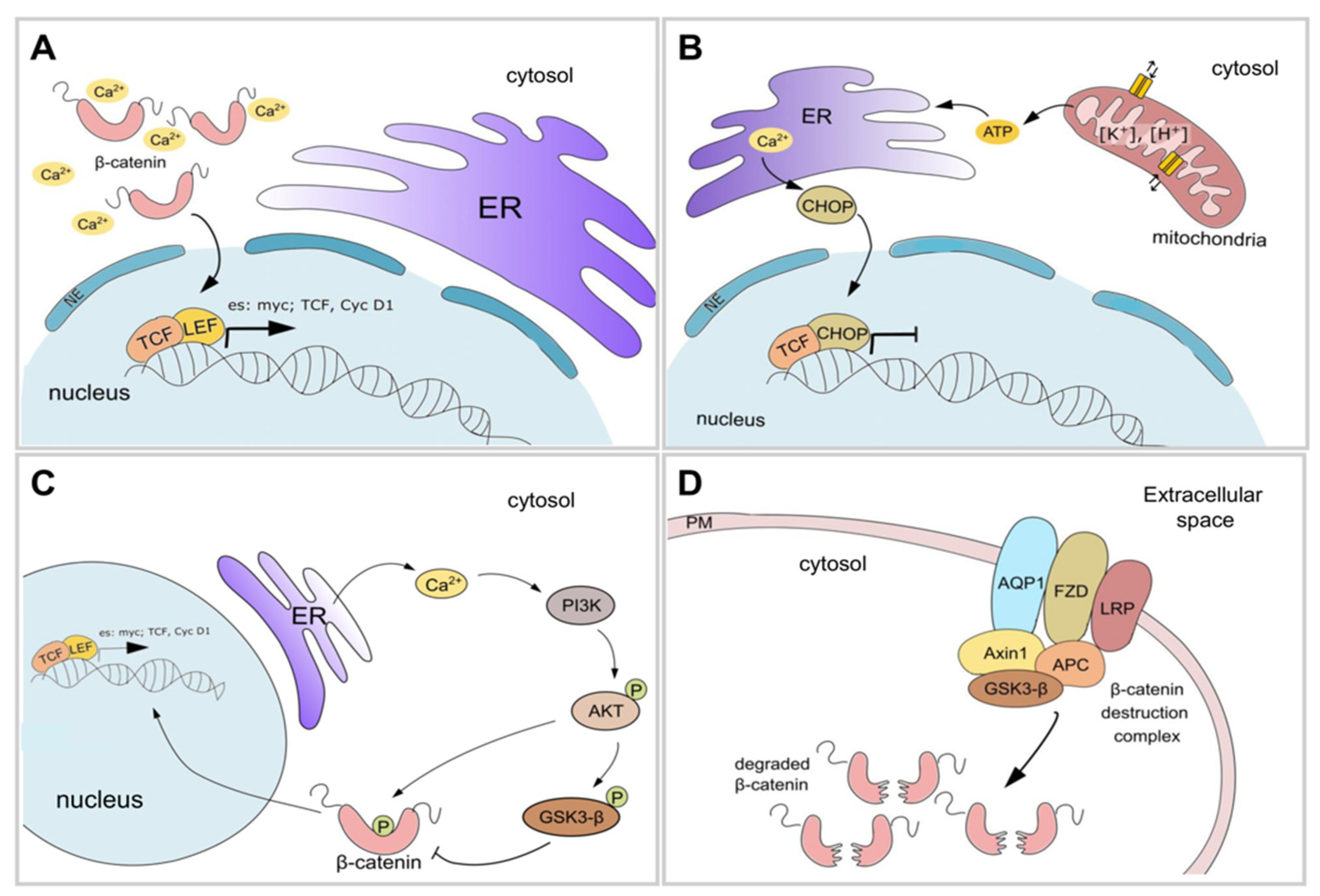{kind=link}
| Channels | Ions Gated | Effect | Cell Types and Diseases | Ref. |
|---|---|---|---|---|
| Kv7.1, | K+ | Tumor suppression by inactivation of the B catenin in the cytoplasm | Colon and Hepatic carcinoma | [51,52] |
| Kv11.1 | K+ | Channel activity inactivates β-catenin, preventing nuclear importation | Breast cancer | [41] |
| CFTR | Cl- | Tumor suppressor, CFTR activity contributes to the correct functionality of the complex FZD-LRP-Wnt. | Cystic fibrosis | [53] |
| TRPV4 | Ca2+ | TRPV4-mediated Ca2+ entry from the extracellular space promotes β-catenin activation by AKT modulation | Gastric cancer | [54] |
| TrpC5 | Ca2+ | TrpC5 overexpression favors the nuclear accumulation of β-catenin by Ca2+ | Colorectal cancer | [55] |
| Channels (ion flux) | Expression in Cancer | Cancer Type | Effects on Wnt Signaling | Mechanism of Action | Effect on Tumorigenesis | Ref. |
|---|---|---|---|---|---|---|
| CaCNA2D3 (Ca2+) | Low | Naso-pharyngeal carcinoma | ↓Wnt signaling targets (cyclin D1, c-myc); ↓MMP7 (invasion); ↓ SNAIL (EMT) | ↑ intracellular Ca2+ flux causes ↑ NLK protein kinase which antagonize canonical Wnt signaling | Overexpression induces mitochondrial-mediated apoptosis and repression of Wnt-dependent invasion, proliferation and EMT. | [56] |
| CaCNA2D3 (Ca2+) | Low | Glioma | ↓ Wnt signaling targets (CCND1, c-myc); ↓ MMP7 (invasion); ↓ SNAIL (EMT) | ↑ intracellular Ca2+ flux causes ↑ NLK protein kinase, which antagonize canonical Wnt signaling | Overexpression induces mitochondrial-mediated apoptosis and repression of Wnt-dependent invasion, proliferation and EMT. | [57] |
| CFTR (Cl−) | Low | Colorectal cancer; Gastric cancer | ↓ Wnt signaling targets | CFTR−/− Apcmin mice display ↑Wnt/β-catenin target genes (Ccnd1, CD44, Axin2, Lgr5, Mmp7, Wnt10A and Ptgs2) | CFTR−/− mice developed significantly more tumors in the colon and the entire small intestine and alteration in the intestinal stem cell compartment. | [58] |
| Kv7.1 (K+) | Low | Colorectal cancer | ↑β-catenin-E-cadherin interaction | Membrane depolarization prevents cytosolic β-catenin release through β-catenin-E-cadherin complex stabilization | Reduced EMT, cell proliferation, and tumorigenesis | [51] |
| Kv11.1 (K+) | Low | Breast cancer | ↑β-catenin-E-cadherin interaction | Membrane depolarization prevents cytosolic β-catenin release through β-catenin-E-cadherin complex stabilization GSK3-β-independent | Represses metastasis formation | [41] |
| Channels (ion flux) | Expression in Cancer | Cancer Type | Effects on Wnt Signaling | Mechanism of Action | Effect on Tumorigenesis | Ref. |
|---|---|---|---|---|---|---|
| Trp5 (non-selective Ca2+ cation channel) | High | Colorectal cancer | ↑Wnt5a, ↑ Wnt signaling targets (cyclin D1 and c-myc) | ↑ Trp5 increase Ca2+ influx in the cell, which induces ↑Wnt5a and ↑ nuclear β-catenin levels | Channel upregulation reduces colorectal cancer differentiation and stemness through Ca2+-Wnt5a | [59] |
| Trp5 (non-selective Ca2+ cation channel) | High | Colorectal cancer | β-catenin stabilization | Reduction in levels of the channels reduces Ca2+ influx lowering β-catenin stability | Silencing of the channel reduces colorectal cancer tumorigenesis and ameliorates 5-Fluorouracil chemoresistance | [55] |
| TRPM4 (non-selective Ca2+ cation channel) | High | Prostate cancer | ↑β-catenin stabilization | ↑ GSK3β and AKT1 phospho-inactive form through Ca2+ flux | Tumor proliferation and progression | [60] |
| TRPV2 (non-selective Ca2+ cation channel) | High | Esophageal squamous cell carcinoma | ↑ Wnt signaling | Increased Ca2+ flux results in ↑ Wnt signaling. The precise mechanism of action still to be determined. | Promotes proliferation, invasion, and angiogenesis | [61] |
| BKCa (K+) | High | Breast cancer | ↑β-catenin stabilization | Modulation of transmembrane depolarization regulates phospho-AKT and, in turn, β-catenin stability | Pharmacological inhibition of the channel (Ibtx) reduces anchorage-dependent growth and tumorigenesis | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muccioli, S.; Brillo, V.; Chieregato, L.; Leanza, L.; Checchetto, V.; Costa, R. From Channels to Canonical Wnt Signaling: A Pathological Perspective. Int. J. Mol. Sci. 2021, 22, 4613. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094613
Muccioli S, Brillo V, Chieregato L, Leanza L, Checchetto V, Costa R. From Channels to Canonical Wnt Signaling: A Pathological Perspective. International Journal of Molecular Sciences. 2021; 22(9):4613. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094613
Chicago/Turabian StyleMuccioli, Silvia, Valentina Brillo, Leonardo Chieregato, Luigi Leanza, Vanessa Checchetto, and Roberto Costa. 2021. "From Channels to Canonical Wnt Signaling: A Pathological Perspective" International Journal of Molecular Sciences 22, no. 9: 4613. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094613