
Phenotypic and Functional Consequences of PLT Binding to Monocytes and Its Association with Clinical Features in SLE

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
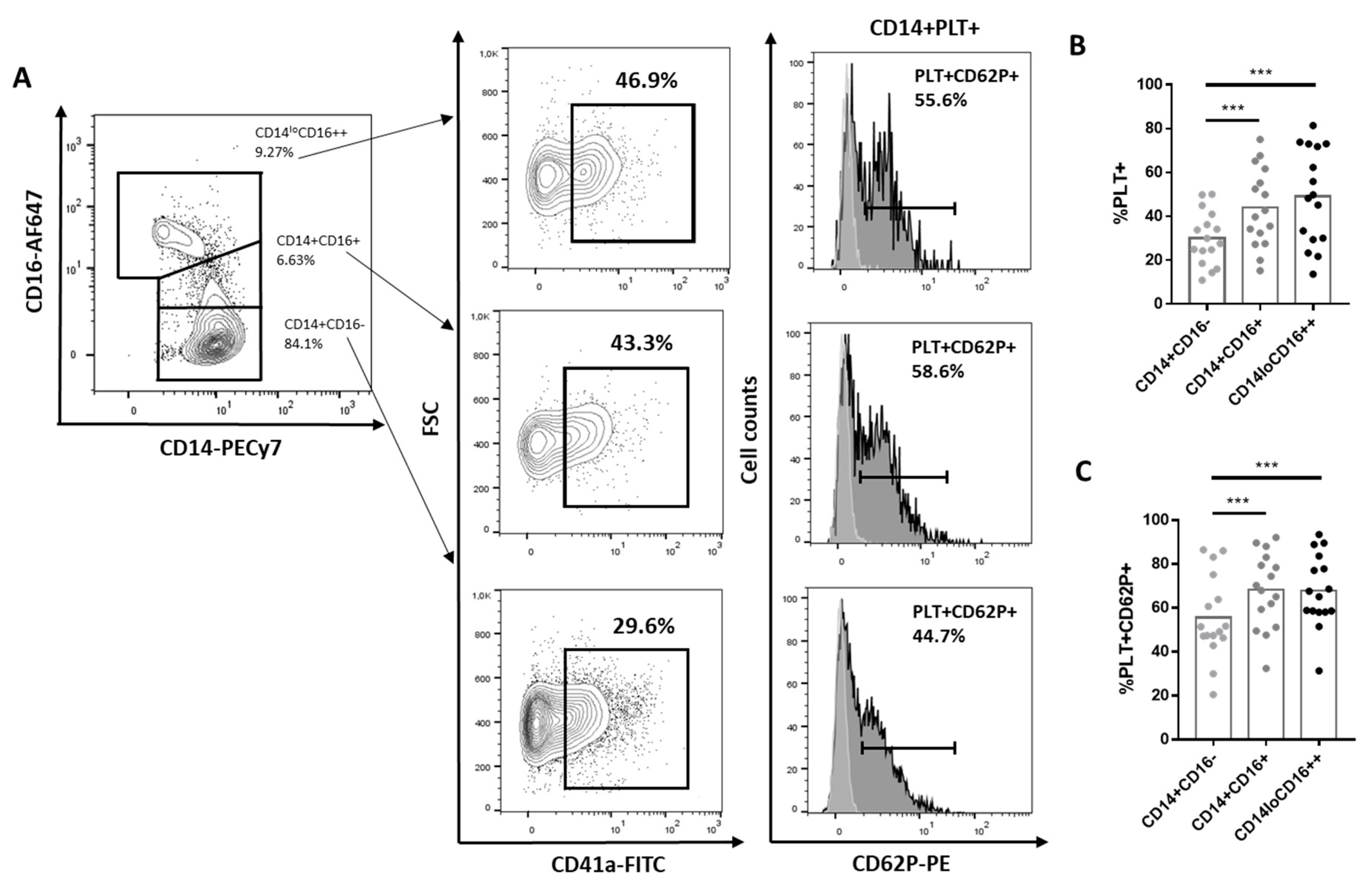
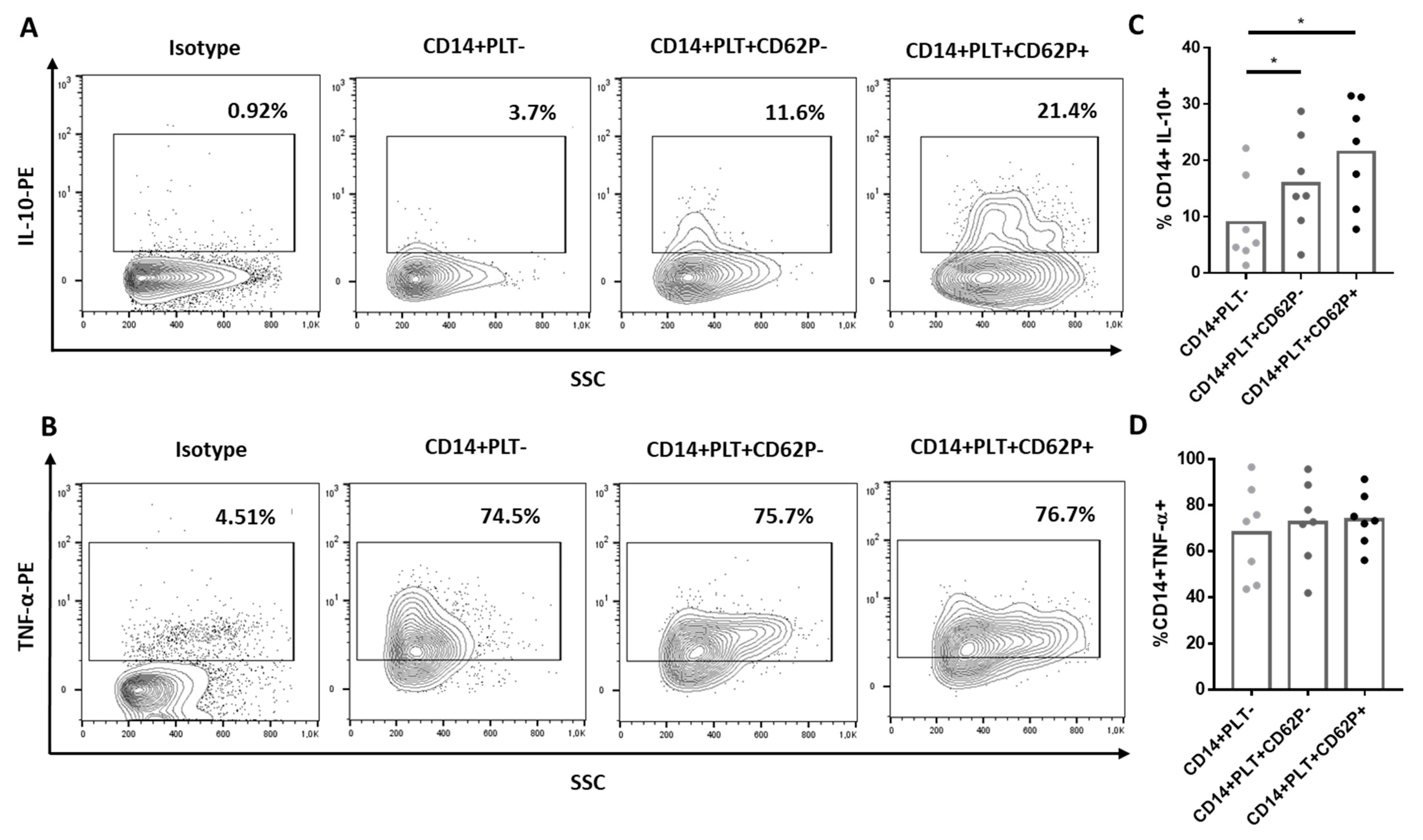
2.1. Comparison of Phenotype and Function of Monocytes with and without Bound PLTs in HD
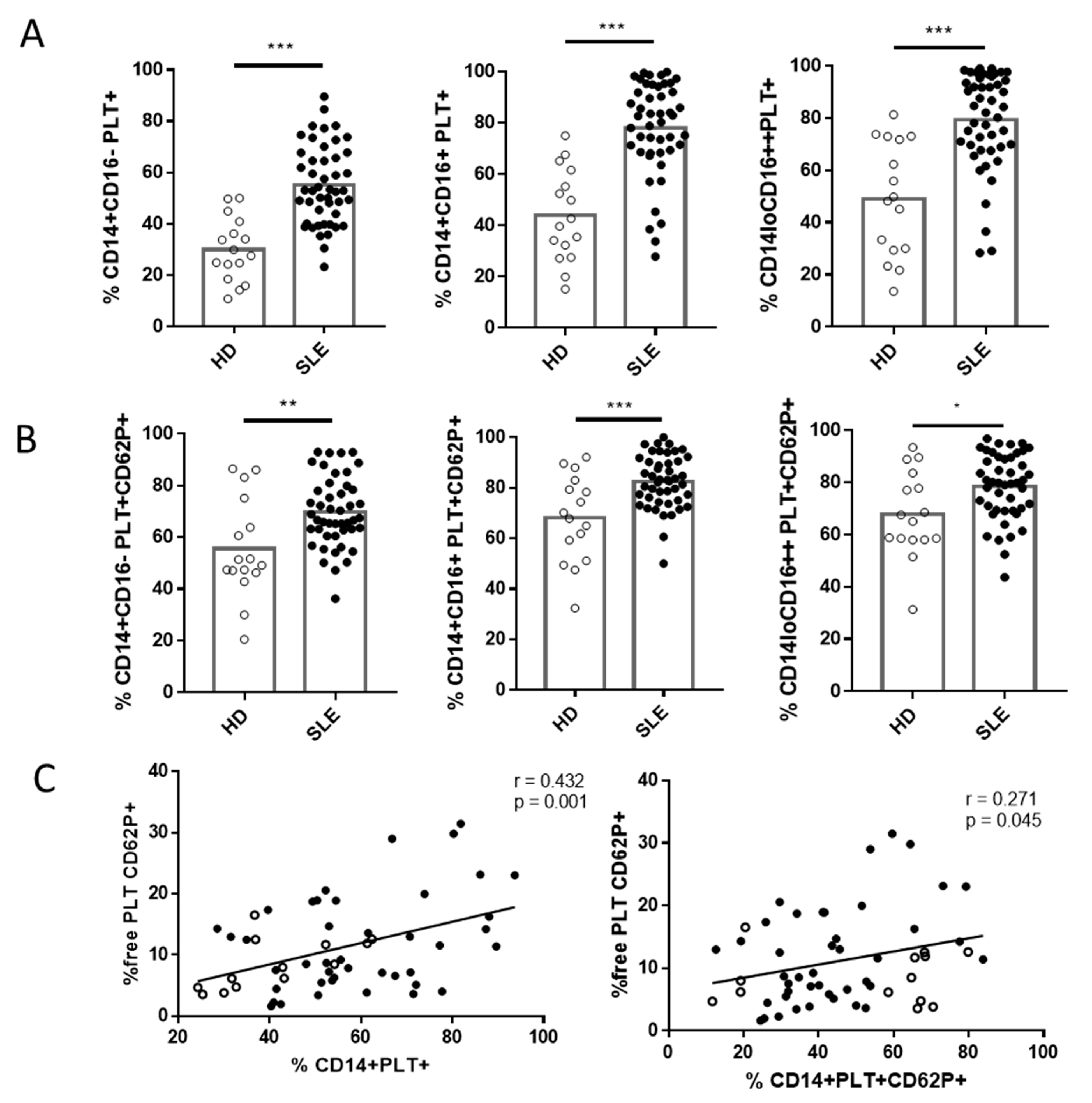
2.2. Comparison of Phenotype and Function of Monocytes with Bound PLTs in HD and SLE Patients
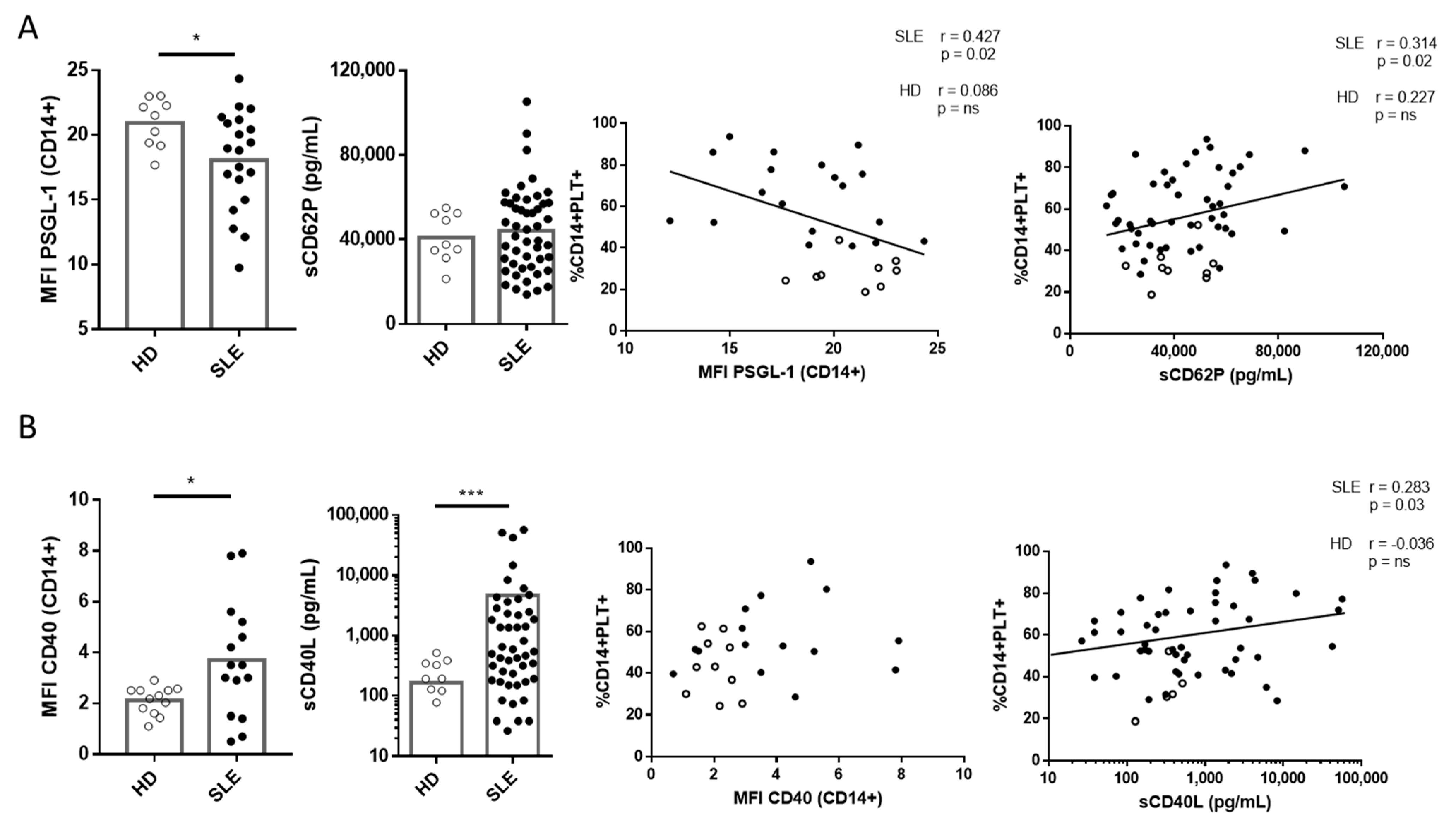
2.3. Correlation of PSGL-1 and CD40 and Their Ligands sCD62P and sCD40L with CD14+PLT+ in SLE Patients and HD
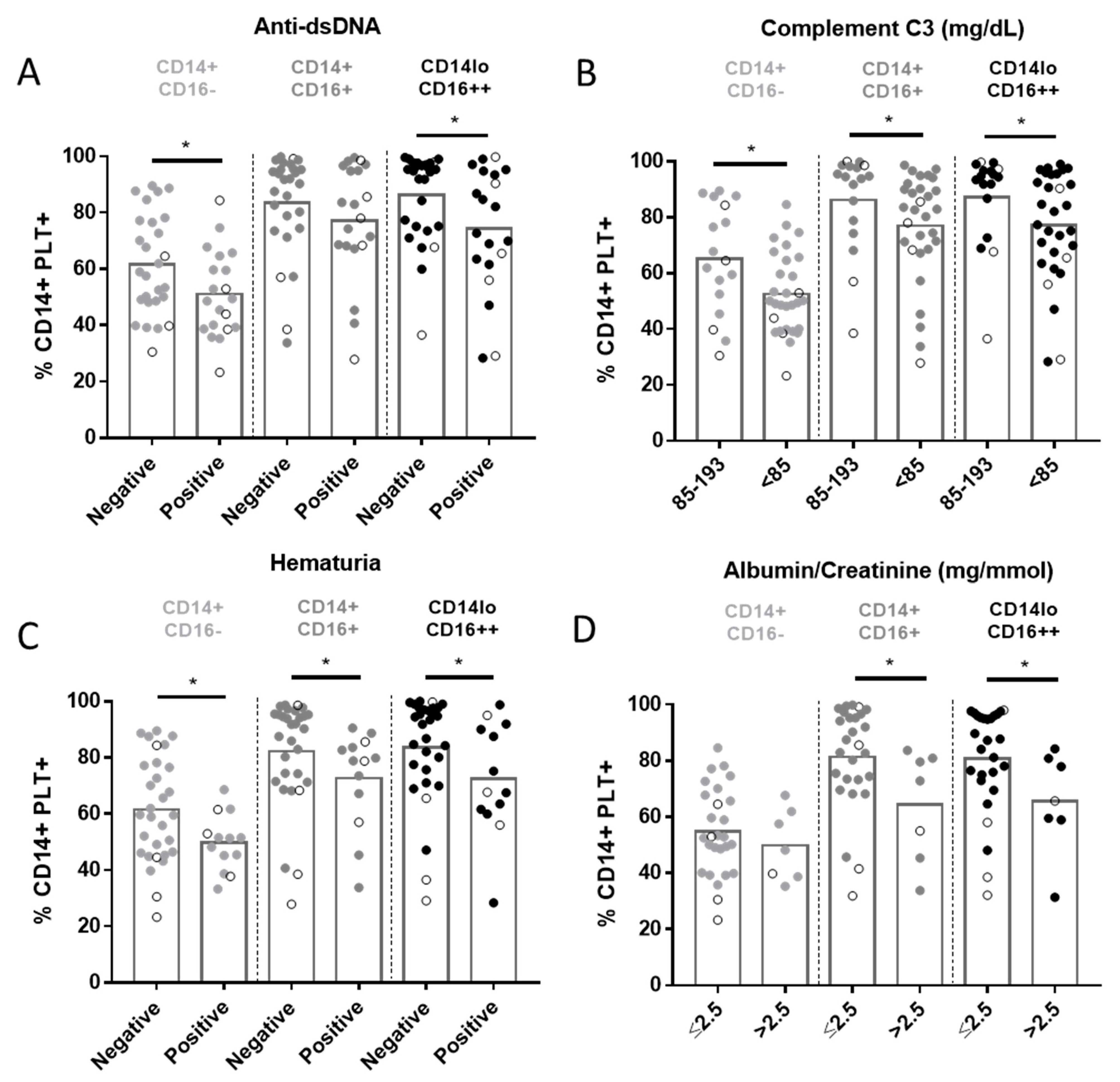
2.4. Relationship between CD14+PLT+ and Clinical Features in SLE Patients
3. Discussion
4. Materials and Methods
4.1. Study Subjects and Sample Collection
4.2. Staining of Whole Blood Cells and Flow Cytometry Analysis
4.3. IL-10 and TNF-α Secretion Assays
4.4. Phagocytosis Assays
4.5. Apoptosis Analysis of Neutrophils and Efferocytosis Assays
4.6. Determination of IFN-α, IL-6, IL-10, sCD62P and sCD40L Levels
4.7. Quantification of Autoantibodies, C3, Albumin/Creatinine and Hematuria
4.8. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Semple, J.W.; Italiano, J.E.; Freedman, J. Platelets and the immune continuum. Nat. Rev. Immunol. 2011, 11, 264–274. [Google Scholar] [CrossRef]
- Tunjungputri, R.N.; van der Ven, A.J.; Riksen, N.; Rongen, G.; Tacke, S.; van den Berg, T.N.A.D.; Fijnheer, R.; Gomes, M.E.; Dinarello, C.A.; van de Veerdonk, F.L.; et al. Differential effects of platelets and platelet inhibition by ticagrelor on TLR2- and TLR4-mediated inflammatory responses. Thromb. Haemost. 2015, 113, 1035–1045. [Google Scholar] [PubMed]
- Gros, A.; Ollivier, V.; Ho-Tin-Noé, B. Platelets in inflammation: Regulation of leukocyte activities and vascular repair. Front. Immunol. 2015, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kral, J.B.; Schrottmaier, W.C.; Salzmann, M.; Assinger, A. Platelet Interaction with Innate Immune Cells. Transfus. Med. Hemotherapy 2016, 43, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Herter, J.M.; Rossaint, J.; Zarbock, A. Platelets in inflammation and immunity. J. Thromb. Haemost. 2014, 12, 1764–1775. [Google Scholar] [CrossRef] [PubMed]
- Morrell, C.N.; Aggrey, A.A.; Chapman, L.M.; Modjeski, K.L. Review Article Emerging roles for platelets as immune and in fl ammatory cells. Blood 2014, 123, 2759–2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, K.C.; Jun, A.J.; Pawar, P.; Jadhav, S.; Napier, S.; McCarty, O.J.T.; Konstantopoulos, K. Preferential binding of platelets to monocytes over neutrophils under flow. Biochem. Biophys. Res. Commun. 2005, 329, 345–355. [Google Scholar] [CrossRef]
- Joseph, J.E.; Harrison, P.; Mackie, I.J.; Isenberg, D.A.; Machin, S.J. Increased circulating platelet-leucocyte complexes and platelet activation in patients with antiphospholipid syndrome, systemic lupus erythematosus and rheumatoid arthritis. Br. J. Haematol. 2001, 115, 451–459. [Google Scholar] [CrossRef]
- Li, N.; Hu, H.; Lindqvist, M.; Wikström-Jonsson, E.; Goodall, A.H.; Hjemdahl, P. Platelet-leukocyte cross talk in whole blood. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2702–2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamora, C.; Riudavets, M.; Anguera, G.; Alserawan, L.; Sullivan, I.; Barba, A.; Serra, J.; Ortiz, M.A.; Gallardo, P.; Perea, L.; et al. Circulating leukocyte–platelet complexes as a predictive biomarker for the development of immune-related adverse events in advanced nonsmall cell lung cancer patients receiving anti-PD-(L)1 blocking agents. Cancer Immunol. Immunother. 2021, 1–14. [Google Scholar]
- Zarbock, A.; Müller, H.; Kuwano, Y.; Ley, K. PSGL-1-dependent myeloid leukocyte activation. J. Leukoc. Biol. 2009, 86, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Kuligowski, M.P.; Kitching, A.R.; Hickey, M.J. Leukocyte Recruitment to the Inflamed Glomerulus: A Critical Role for Platelet-Derived P-Selectin in the Absence of Rolling. J. Immunol. 2006, 176, 6991–6999. [Google Scholar] [CrossRef] [Green Version]
- Da Costa Martins, P.A.; van Gils, J.M.; Mol, A.; Hordijk, P.L.; Zwaginga, J.J. Platelet binding to monocytes increases the adhesive properties of monocytes by up-regulating the expression and functionality of beta1 and beta2 integrins. J. Leukoc. Biol. 2006, 79, 499–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyrich, A.S.; Elstad, M.R.; McEver, R.P.; McIntyre, T.M.; Moore, K.L.; Morrissey, J.H.; Prescott, S.M.; Zimmerman, G.A. Activated platelets signal chemokine synthesis by human monocytes. J. Clin. Investig. 1996, 97, 1525–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, D.; Dohle, F.; Terstesse, M.; Bangen, P.; August, C.; Pauels, H.-G.; Heidenreich, S. Down-Regulation of Monocyte Apoptosis by Phagocytosis of Platelets: Involvement of a Caspase-9, Caspase-3, and Heat Shock Protein 70-Dependent Pathway. J. Immunol. 2002, 168, 6152–6158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passacquale, G.; Vamadevan, P.; Pereira, L.; Hamid, C.; Corrigall, V.; Ferro, A. Monocyte-platelet interaction induces a pro-inflammatory phenotype in circulating monocytes. PLoS ONE 2011, 6, e25595. [Google Scholar] [CrossRef] [Green Version]
- Ammon, C.; Kreutz, M.; Rehli, M.; Krause, S.W.; Andreesen, R. Platelets induce monocyte differentiation in serum-free coculture Abstract: Terminal maturation of blood mono- cytes (MO) in vitro and in vivo into macrophages connections that may be important for MO differen- and/or with platelets. Ammon C, Kreutz M, Rehli M, Krause SW, Andreesen R. Platelets induce monocyte differentiation in serum-free coculture. J. Leukoc. Biol. 1998, 63, 469–476. [Google Scholar] [PubMed]
- Gudbrandsdottir, S.; Hasselbalch, H.C.; Nielsen, C.H. Activated platelets enhance IL-10 secretion and reduce TNF-α secretion by monocytes. J. Immunol. 2013, 191, 4059–4067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartzkopff, F.; Petersen, F.; Grimm, T.A.; Brandt, E. CXC chemokine ligand 4 (CXCL4) down-regulates CC chemokine receptor expression on human monocytes. Innate Immun. 2012, 18, 124–139. [Google Scholar] [CrossRef]
- Linke, B.; Schreiber, Y.; Picard-Willems, B.; Slattery, P.; Nüsing, R.M.; Harder, S.; Geisslinger, G.; Scholich, K. Activated Platelets Induce an Anti-Inflammatory Response of Monocytes/Macrophages through Cross-Regulation of PGE2 and Cytokines. Mediators Inflamm. 2017, 2017, 1463216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bournazos, S.; Rennie, J.; Hart, S.P.; Dransfield, I. Choice of anticoagulant critically affects measurement of circulating platelet-leukocyte complexes. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Takeda, Y.; Marumo, M.; Nara, H.; Feng, Z.-G.; Asao, H.; Wakabayashi, I. Selective induction of anti-inflammatory monocyte-platelet aggregates in a model of pulsatile blood flow at low shear rates. Platelets 2016, 27, 583–592. [Google Scholar] [CrossRef]
- Sümegi, A.; Antal-Szalmás, P.; Aleksza, M.; Kovács, I.; Sipka, S.; Zeher, M.; Kiss, E.; Szegedi, G. Glucocorticosteroid therapy decreases CD14-expression and CD14-mediated LPS-binding and activation of monocytes in patients suffering from systemic lupus erythematosus. Clin. Immunol. 2005, 117, 271–279. [Google Scholar] [CrossRef]
- Mukherjee, R.; Kanti Barman, P.; Kumar Thatoi, P.; Tripathy, R.; Kumar Das, B.; Ravindran, B. Non-Classical monocytes display inflammatory features: Validation in Sepsis and Systemic Lupus Erythematous. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuentes Q., E.; Fuentes Q., F.; Andrés, V.; Pello, O.M.; de Mora, J.F.; Palomo G., I. Role of platelets as mediators that link inflammation and thrombosis in atherosclerosis. Platelets 2012, 24, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Alcocer Varela, J.; Laffon, A.; Alarcon Segovia, D. Defective monocyte production of, and T lymphocyte response to, interleukin-1 in the peripheral blood of patients with systemic lupus erythematosus. Clin. Exp. Immunol. 1984, 55, 125–132. [Google Scholar]
- Yuan, W.; Dimartino, S.J.; Redecha, P.B.; Ivashkiv, L.B.; Salmon, J.E. Systemic lupus erythematosus monocytes are less responsive to interleukin-10 in the presence of immune complexes. Arthritis Rheum. 2011, 63, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linker-Israeli, M.; Deans, R.J.; Wallace, D.J.; Prehn, J.; Ozeri-Chen, T.; Klinenberg, J.R. Elevated levels of endogenous IL-6 in systemic lupus erythematosus. J. Immunol. 2015, 147, 117–123. [Google Scholar]
- Herrmann, M.; Voll, R.E.; Zoller, O.M.; Hagenhofer, M.; Ponner, B.B.; Kalden, J.R. Impaired phagocytosis of apoptotic cell material by monocyte-derived macrophages from patients with systemic lupus erythematosus. Arthritis Rheum. 1998, 41, 1241–1250. [Google Scholar] [CrossRef]
- Munoz, L.E.; Gaipl, U.S.; Franz, S.; Sheriff, A.; Voll, R.E.; Kalden, J.R.; Herrmann, M. SLE - A disease of clearance deficiency? Rheumatology 2005, 44, 1101–1107. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.Y.; Chien, P.; Indik, Z.K.; Schreiber, A.D. Human platelet FcγRIIA and phagocytes in immune-complex clearance. Mol. Immunol. 2011, 48, 691–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linge, P.; Fortin, P.R.; Lood, C.; Bengtsson, A.A.; Boilard, E. The non-haemostatic role of platelets in systemic lupus erythematosus. Nat. Rev. Rheumatol. 2018, 14, 195–213. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.I.; Chen, Z.; Xu, H.; Li, C.Q.; Dong, J.F.; McIntire, L.V.; Ballantyne, C.M.; Zhang, L.; Furman, M.I.; Berndt, M.C.; et al. Platelet glycoprotein Ibα is a counterreceptor for the leukocyte integrin Mac-1 (CD11b/CD18). J. Exp. Med. 2000, 192, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Lindmark, E.; Tenno, T.; Siegbahn, A. Role of platelet P-selectin and CD40 ligand in the induction of monocytic tissue factor expression. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2322–2328. [Google Scholar] [CrossRef] [Green Version]
- Silverstein, R.L.; Asch, A.S.; Nachman, R.L. Glycoprotein IV mediates thrombospondin-dependent platelet-monocyte and platelet-U937 cell adhesion. J. Clin. Investig. 1989, 84, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Von Brühl, M.L.; Barocke, V.; Cullen, P.; Mayer, K.; Okrojek, R.; Steinhart, A.; Ahmad, Z.; Kremmer, E.; Nieswandt, B.; et al. EMMPRIN (CD147/basigin) mediates platelet-monocyte interactions in vivo and augments monocyte recruitment to the vascular wall. J. Thromb. Haemost. 2011, 9, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Rong, M.; Wang, C.; Wu, Z.; Zeng, W.; Zheng, Z.; Han, Q.; Jia, J.; Li, X.; Zhu, P. Platelets induce a proinflammatory phenotype in monocytes via the CD147 pathway in rheumatoid arthritis. Arthritis Res. Ther. 2014, 16, 478. [Google Scholar] [CrossRef] [Green Version]
- Van Gils, J.M.; Zwaginga, J.J.; Hordijk, P.L. Molecular and functional interactions among monocytes, platelets, and endothelial cells and their relevance for cardiovascular diseases. J. Leukoc. Biol. 2008, 85, 195–204. [Google Scholar] [CrossRef]
- Larsen, E.; Celi, A.; Gilbert, G.E.; Furie, B.C.; Erban, J.K.; Bonfanti, R.; Wagner, D.D.; Furie, B. PADGEM protein: A receptor that mediates the interaction of activated platelets with neutrophils and monocytes. Cell 1989, 59, 305–312. [Google Scholar] [CrossRef]
- Michelson, A.D.; Barnard, M.R.; Krueger, L.A.; Valeri, C.R.; Furman, M.I. Circulating monocyte-platelet aggregates are a more sensitive marker of in vivo platelet activation than platelet surface P-selectin: Studies in baboons, human coronary intervention, and human acute myocardial infarction. Circulation 2001, 104, 1533–1537. [Google Scholar] [CrossRef] [Green Version]
- Christersson, C.; Johnell, M.; Siegbahn, A. Tissue factor and IL8 production by P-selectin-dependent platelet-monocyte aggregates in whole blood involves phosphorylation of Lyn and is inhibited by IL10. J. Thromb. Haemost. 2008, 6, 986–994. [Google Scholar] [CrossRef]
- Wong, C.H.Y.; Jenne, C.N.; Petri, B.; Chrobok, N.L.; Kubes, P. Nucleation of platelets with blood-borne pathogens on Kupffer cells precedes other innate immunity and contributes to bacterial clearance. Nat. Immunol. 2013, 14, 785–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarma, J.; Laan, C.A.; Alam, S.; Jha, A.; Fox, K.A.A.; Dransfield, I. Increased platelet binding to circulating monocytes in acute coronary syndromes. Circulation 2002, 105, 2166–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, S.A.; Sommerfield, A.J.; Sarma, J.; Twomey, P.J.; Newby, D.E.; Frier, B.M.; Fox, K.A.A. Increased CD40 ligand and platelet – monocyte aggregates in patients with type 1 diabetes mellitus. Atherosclerosis 2004, 176, 321–325. [Google Scholar] [CrossRef]
- Pamuk, G.E.; Nurı Pamuk, Ö.; Örüm, H.; Arican, Ö.; Turgut, B.; Demır, M. Elevated platelet-monocyte complexes ın patıents wıth psorıatıc arthrıtıs. Platelets 2009, 20, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, J.; Hamada, E.; Shodai, T.; Kamoshida, G.; Kudo, S.; Itoh, S.; Koike, J.; Nagata, K.; Irimura, T.; Tsuji, T. Cytokine secretion from human monocytes potentiated by P-selectin-mediated cell adhesion. Int. Arch. Allergy Immunol. 2013, 160, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Da Costa Martins, P.; Van Den Berk, N.; Ulfman, L.H.; Koenderman, L.; Hordijk, P.L.; Zwaginga, J.J. Platelet-Monocyte Complexes Support Monocyte Adhesion to Endothelium by Enhancing Secondary Tethering and Cluster Formation. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawley, K.L.; Cruz, A.R.; Benjamin, S.J.; La Vake, C.J.; Cervantes, J.L.; LeDoyt, M.; Ramirez, L.G.; Mandich, D.; Fiel-Gan, M.; Caimano, M.J.; et al. IFNγ enhances CD64-potentiated phagocytosis of Treponema pallidum opsonized with human syphilitic serum by human macrophages. Front. Immunol. 2017, 8, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Andonegui, G.; Kerfoot, S.M.; McNagny, K.; Ebbert, K.V.J.; Patel, K.D.; Kubes, P. Platelets express functional Toll-like receptor-4. Blood 2005, 106, 2417–2423. [Google Scholar] [CrossRef]
- Semple, J.W.; Aslam, R.; Kim, M.; Speck, E.R.; Freedman, J. Brief report Platelet-bound lipopolysaccharide enhances Fc receptor–mediated phagocytosis of IgG-opsonized platelets. Blood. 2007, 109, 4803–4806. [Google Scholar] [CrossRef] [Green Version]
- Ståhl, A.L.; Svensson, M.; Mörgelin, M.; Svanborg, C.; Tarr, P.I.; Mooney, J.C.; Watkins, S.L.; Johnson, R.; Karpman, D. Lipopolysaccharide from enterohemorrhagic Escherichia coli binds to platelets through TLR4 and CD62 and is detected on circulating platelets in patients with hemolytic uremic syndrome. Blood 2006, 108, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Han, J.; Welch, E.J.; Ye, R.D.; Voyno-Yasenetskaya, T.A.; Malik, A.B.; Du, X.; Li, Z. Lipopolysaccharide stimulates platelet secretion and potentiates platelet aggregation via TLR4/MyD88 and the cGMP-dependent protein kinase pathway. J. Immunol. 2009, 182, 7997–8004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skrzeczyńska-Moncznik, J.; Bzowska, M.; Loseke, S.; Grage-Griebenow, E.; Zembala, M.; Pryjma, J. Peripheral blood CD14high CD16+ monocytes are main producers of IL-10. Scand. J. Immunol. 2008, 67, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Hirose, S.; Lin, Q.; Ohtsuji, M.; Nishimura, H.; Verbeek, J.S. Monocyte subsets involved in the development of systemic lupus erythematosus and rheumatoid arthritis. Int. Immunol. 2019, 31, 687–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsik, C.; Mayr, F.; Cardona, F.; Schaller, G.; Wagner, O.F.; Jilma, B. Endotoxin down-modulates P-selectin glycoprotein ligand-1 (PSGL-1, CD162) on neutrophils in humans. J. Clin. Immunol. 2004, 24, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Duan, Z.; Li, D.; Li, D.; Wang, Z.; Ren, L.; Shen, T.; Shao, Y. Higher levels of circulating monocyte-platelet aggregates are correlated with viremia and increased sCD163 levels in HIV-1 infection. Cell. Mol. Immunol. 2015, 12, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Davenpeck, K.L.; Brummet, M.E.; Hudson, S.A.; Mayer, R.J.; Bochner, B.S. Activation of Human Leukocytes Reduces Surface P-Selectin Glycoprotein Ligand-1 (PSGL-1, CD162) and Adhesion to P-Selectin In Vitro. J. Immunol. 2000, 165, 2764–2772. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lee, P.Y.; Reeves, W.H. Monocyte and macrophage abnormalities in systemic lupus erythematosus. Arch. Immunol. Ther. Exp. (Warsz). 2010, 58, 355–364. [Google Scholar] [CrossRef]
- Le Blanc, J.; Lordkipanidzé, M. Platelet Function in Aging. Front. Cardiovasc. Med. 2019, 6, 1–8. [Google Scholar] [CrossRef]
- Katsiari, C.G.; Liossis, S.N.C.; Souliotis, V.L.; Dimopoulos, A.M.; Manoussakis, M.N.; Sfikakis, P.P. Aberrant expression of the costimulatory molecule CD40 ligand on monocytes from patients with systemic lupus erythematosus. Clin. Immunol. 2002, 103, 54–62. [Google Scholar] [CrossRef]
- Boilard, E.; Blanco, P.; Nigrovic, P.A. Platelets: Active players in the pathogenesis of arthritis and SLE. Nat. Rev. Rheumatol. 2012, 8, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Zamora, C.; Toniolo, E.; Diaz-Torné, C.; Cantó, E.; Magallares, B.; Ortiz, M.A.; Perea, L.; Corominas, H.; Vidal, S. Association of Platelet Binding to Lymphocytes with B Cell Abnormalities and Clinical Manifestations in Systemic Lupus Erythematosus. Mediators Inflamm. 2019, 2019, 1–12. [Google Scholar] [CrossRef]
- Morel, Y.; Truneh, A.; Sweet, R.W.; Olive, D.; Costello, R.T. The TNF Superfamily Members LIGHT and CD154 (CD40 Ligand) Costimulate Induction of Dendritic Cell Maturation and Elicit Specific CTL Activity. J. Immunol. 2001, 167, 2479–2486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Ji, Q.; Hjemdahl, P. Platelet-lymphocyte conjugation differs between lymphocyte subpopulations. J. Thromb. Haemost. 2006, 4, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Llorente, L.; Richaud-Patin, Y.; Wijdenes, J.; Alcocer-Varela, J.; Maillot, M.C.; Durand-Gasselin, I.; Fourrier, B.M.; Galanaud, P.; Emilie, D. Spontaneous production of interleukin-10 by B lymphocytes and monocytes in systemic lupus erythematosus. Eur. Cytokine Netw. 1993, 4, 421–427. [Google Scholar] [PubMed]
- Llorente, L.; Richaud-Patin, Y.; García-Padilla, C.; Claret, E.; Jakez-Ocampo, J.; Cardiel, M.H.; Alcocer-Varela, J.; Grangeot-Keros, L.; Alarcón-Segovia, D.; Wijdenes, J.; et al. Clinical and biologic effects of anti-interleukin-10 monoclonal antibody administration in systemic Lupus erythematosus. Arthritis Rheum. 2000, 43, 1790–1800. [Google Scholar] [CrossRef]
- Uchida, M.; Ooka, S.; Goto, Y.; Suzuki, K.; Fujimoto, H.; Ishimori, K.; Matsushita, H.; Takakuwa, Y.; Kawahata, K. Anti-IL-10 antibody in systemic lupus erythematosus. Open Access Rheumatol. Res. Rev. 2019, 11, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Go, N.F.; Castle, B.E.; Barrett, R.; Kastelein, R.; Dang, W.; Mosmann, T.R.; Moore, K.W.; Howard, M. Interleukin 10, a novel B cell stimulatory factor: Unresponsiveness of X chromosome-linked immunodeficiency B cells. J. Exp. Med. 1990, 172, 1625–1631. [Google Scholar] [CrossRef] [Green Version]
- Levy, Y.; Brouet, J.C. Interleukin-10 prevents spontaneous death of germinal center B cells by induction of the bcl-2 protein. J. Clin. Investig. 1994, 93, 424–428. [Google Scholar] [CrossRef]
- Houssiau, F.A.; Vanden Berghe, M.; Devogelaer, J.P.; Lefebvre, C.; Lambert, M.; Renauld, J.C. Serum interleukin 10 titers in systemic lupus erythematosus reflect disease activity. Lupus 1995, 4, 393–395. [Google Scholar] [CrossRef]
- Conti, F.; Ceccarelli, F.; Perricone, C.; Massaro, L.; Marocchi, E.; Miranda, F.; Spinelli, F.R.; Truglia, S.; Alessandri, C.; Valesini, G. Systemic Lupus Erythematosus with and without Anti-dsDNA Antibodies: Analysis from a Large Monocentric Cohort. Mediators Inflamm. 2015, 2015, 328078. [Google Scholar] [PubMed]
- Satoh, M.; Hamamoto, T.; Shibata, T.; Katoh, M.; Ashihara, M.; Takahashi, M.; Ishitani, K.; Natsume, I.; Abe, Y.; Yoshida, S.; et al. An Evaluation on the 1982 Revised Criteria for the Classification of Systemic Lupus Erythematosus. Jpn. J. Clin. Immunol. 1987, 10, 186–193. [Google Scholar] [CrossRef]
- Guzmán, J.; Cardiel, M.H.; Arce-Salinas, A.; Sánchez-Guerrero, J.; Alarcón-Segovia, D. Measurement of disease activity in systemic lupus erythematosus. Prospective validation of 3 clinical indices. J. Rheumatol. 1992, 19, 1551–1558. [Google Scholar] [PubMed]
- Ziegler-Heitbrock, L.; Hofer, T.P.J. Toward a refined definition of monocyte subsets. Front. Immunol. 2013, 4, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Zamora, C.; Cantó, E.; Nieto, J.C.; Angels Ortiz, M.; Juarez, C.; Vidal, S. Functional consequences of CD36 downregulation by TLR signals. Cytokine 2012, 60, 257–265. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| General Conditions | SLE Patients | Healthy Donors |
|---|---|---|
| Age in years, mean ± SD | 50.4 ± 15.29 | 52.2 ± 17.7 |
| Gender, % (n) women | 93.75 (45) | 90 (18) |
| Years of development, median (IQR) | 9 (3–17) | |
| SLEDAI, mean (range) | 2.89 (0–10) | |
| Laboratory parameters | ||
| C3 (mg/dL, median (IQR)) | 75.1 (63.03–100.8) | |
| Decreased C3 or C4, % (n) | 66.7 (32) | |
| Positive anti-dsDNA, % (n) | 41.7 (20) | |
| Positive ANA, % (n) | 91.7 (44) | |
| Positive anti-Sm, % (n) | 23 (11) | |
| Positive anti-C1q, % (n) | 43 (20) | |
| Positive anti-SSB-La, % (n) | 16 (7) | |
| Positive anti-SSA-Ro, % (n) | 44 (20) | |
| Positive anti-U1-RNP, % (n) | 30 (14) | |
| Positive antiphospholipid antibody, % (n) | 27 (13) | |
| Leukopenia and/or lymphopenia, % (n) | 37.5 (18) | |
| Clinical features, % (n) | ||
| Renal involvement | 41.7 (20) | |
| Hematuria | 27.1 (13) | |
| Albumin/creatinine > 2.5 | 14.6 (7) | |
| Cutaneous involvement | 18.8 (9) | |
| Arthritis | 14.6 (7) | |
| Treatment, % (n) | ||
| None | 6 (3) | |
| Mycophenolate | 17 (8) | |
| Prednisone | 37 (18) | |
| Hydroxychloroquine | 79 (38) | |
| Azathioprine | 8 (4) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariscal, A.; Zamora, C.; Magallares, B.; Salman-Monte, T.C.; Ortiz, M.À.; Díaz-Torné, C.; Castellví, I.; Corominas, H.; Vidal, S. Phenotypic and Functional Consequences of PLT Binding to Monocytes and Its Association with Clinical Features in SLE. Int. J. Mol. Sci. 2021, 22, 4719. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094719
Mariscal A, Zamora C, Magallares B, Salman-Monte TC, Ortiz MÀ, Díaz-Torné C, Castellví I, Corominas H, Vidal S. Phenotypic and Functional Consequences of PLT Binding to Monocytes and Its Association with Clinical Features in SLE. International Journal of Molecular Sciences. 2021; 22(9):4719. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094719
Chicago/Turabian StyleMariscal, Anaís, Carlos Zamora, Berta Magallares, Tarek Carlos Salman-Monte, Mª Àngels Ortiz, Cesar Díaz-Torné, Iván Castellví, Héctor Corominas, and Silvia Vidal. 2021. "Phenotypic and Functional Consequences of PLT Binding to Monocytes and Its Association with Clinical Features in SLE" International Journal of Molecular Sciences 22, no. 9: 4719. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094719