
Cobalt Regulates Activation of Camk2α in Neurons by Influencing Fructose 1,6-Bisphosphatase 2 Quaternary Structure and Subcellular Localization


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
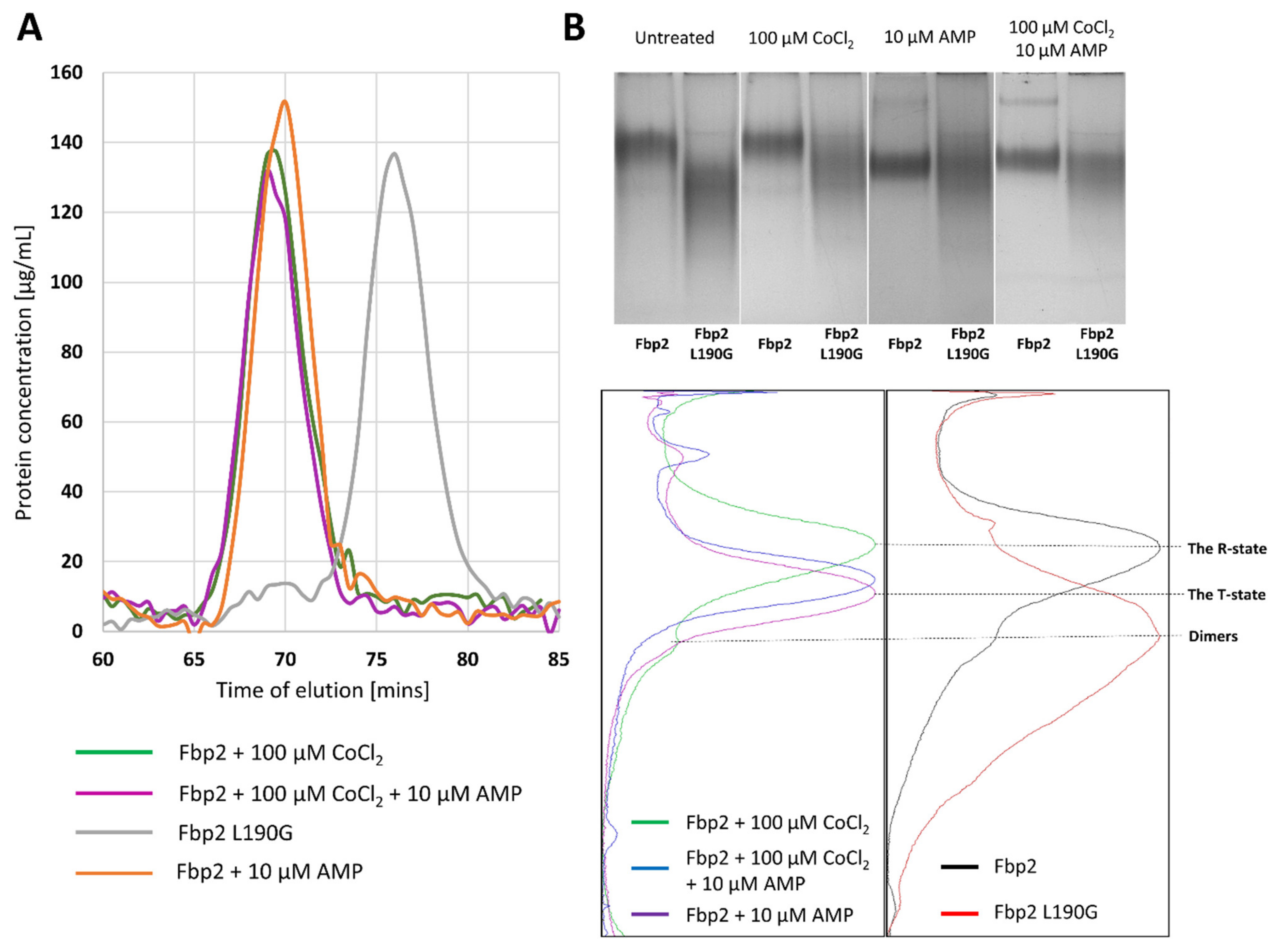
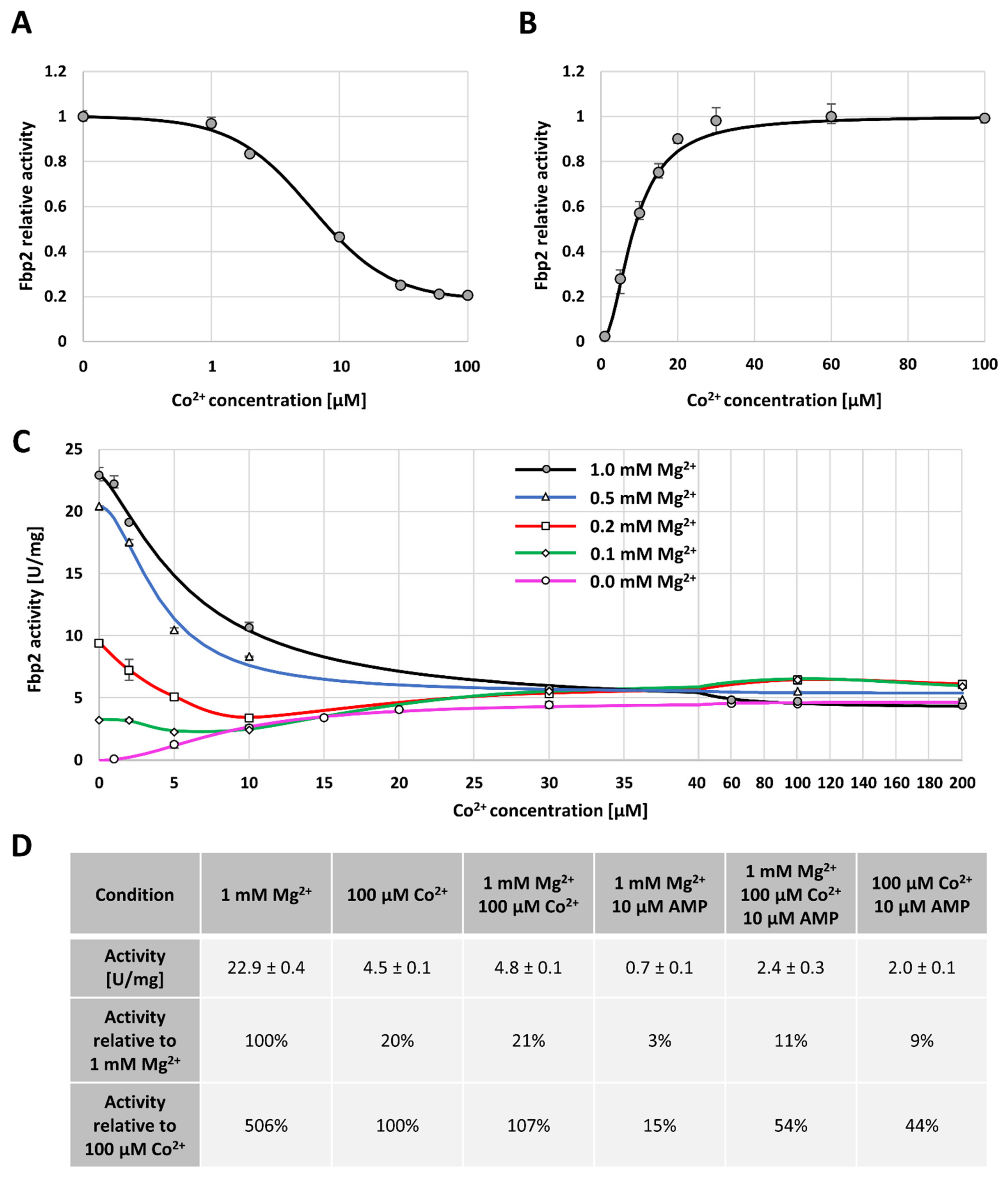
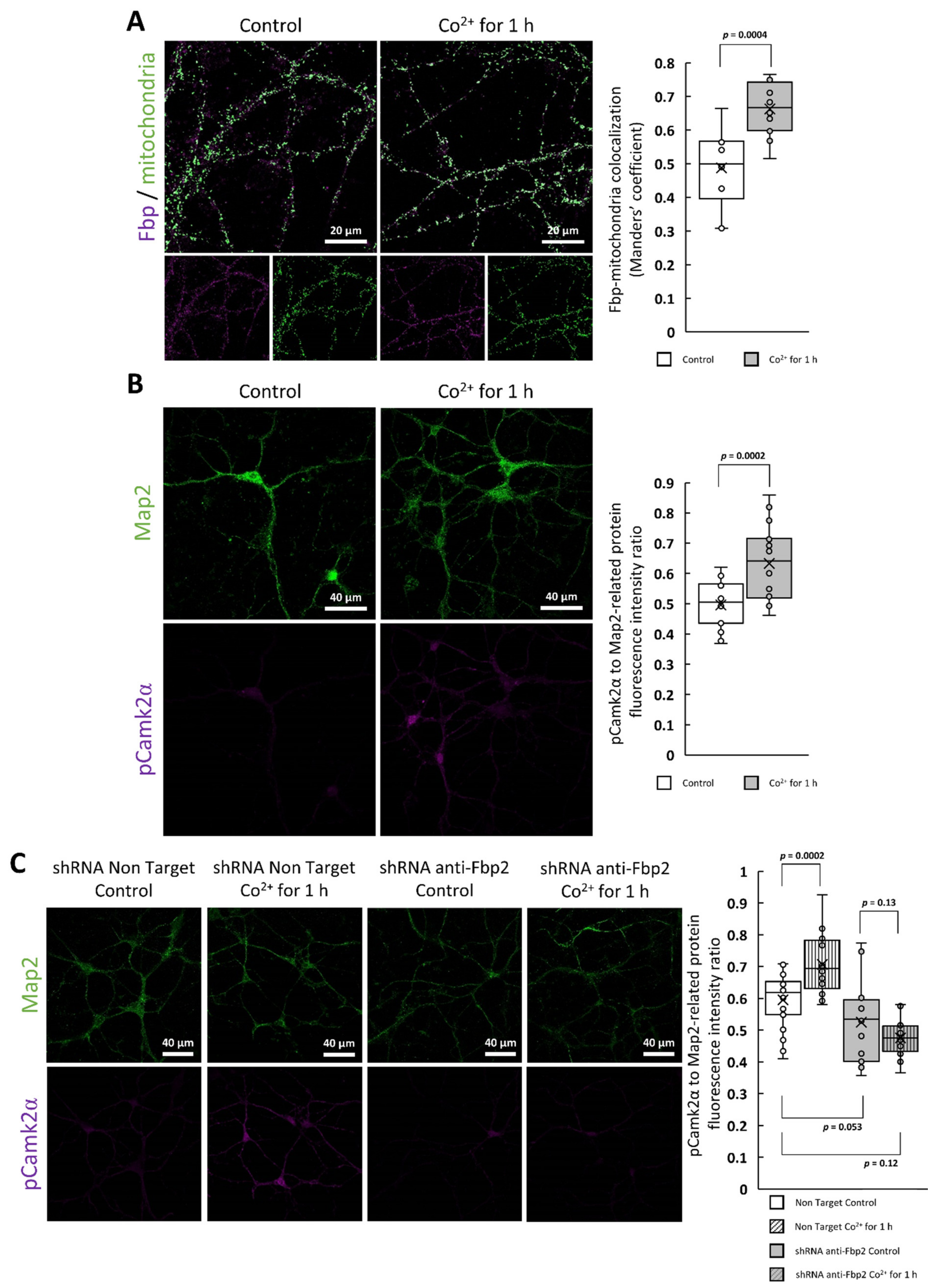
2.1. The Effect of Cobalt Ions on Fpb2 Kinetics, Structure and Subcellular Localization
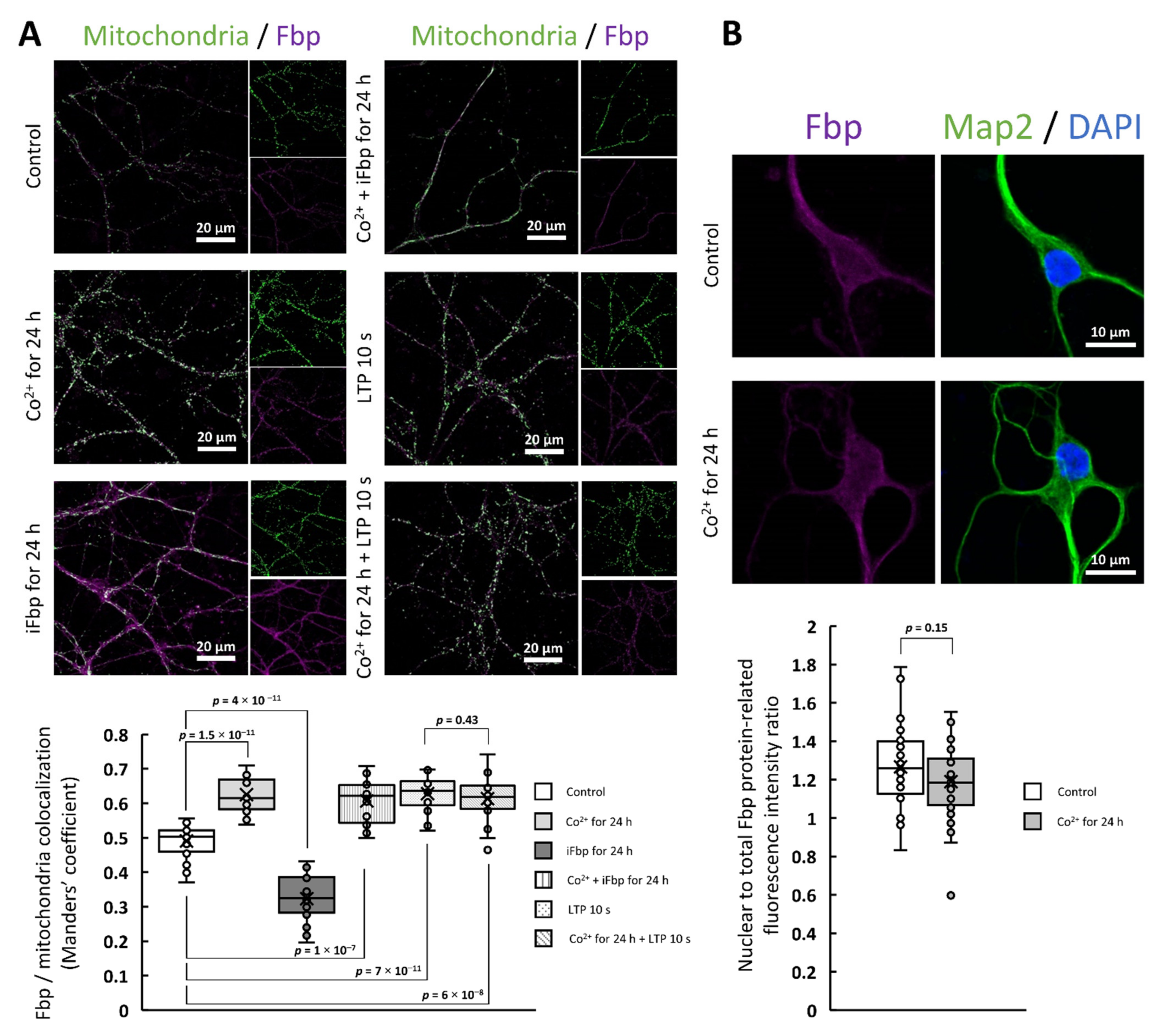
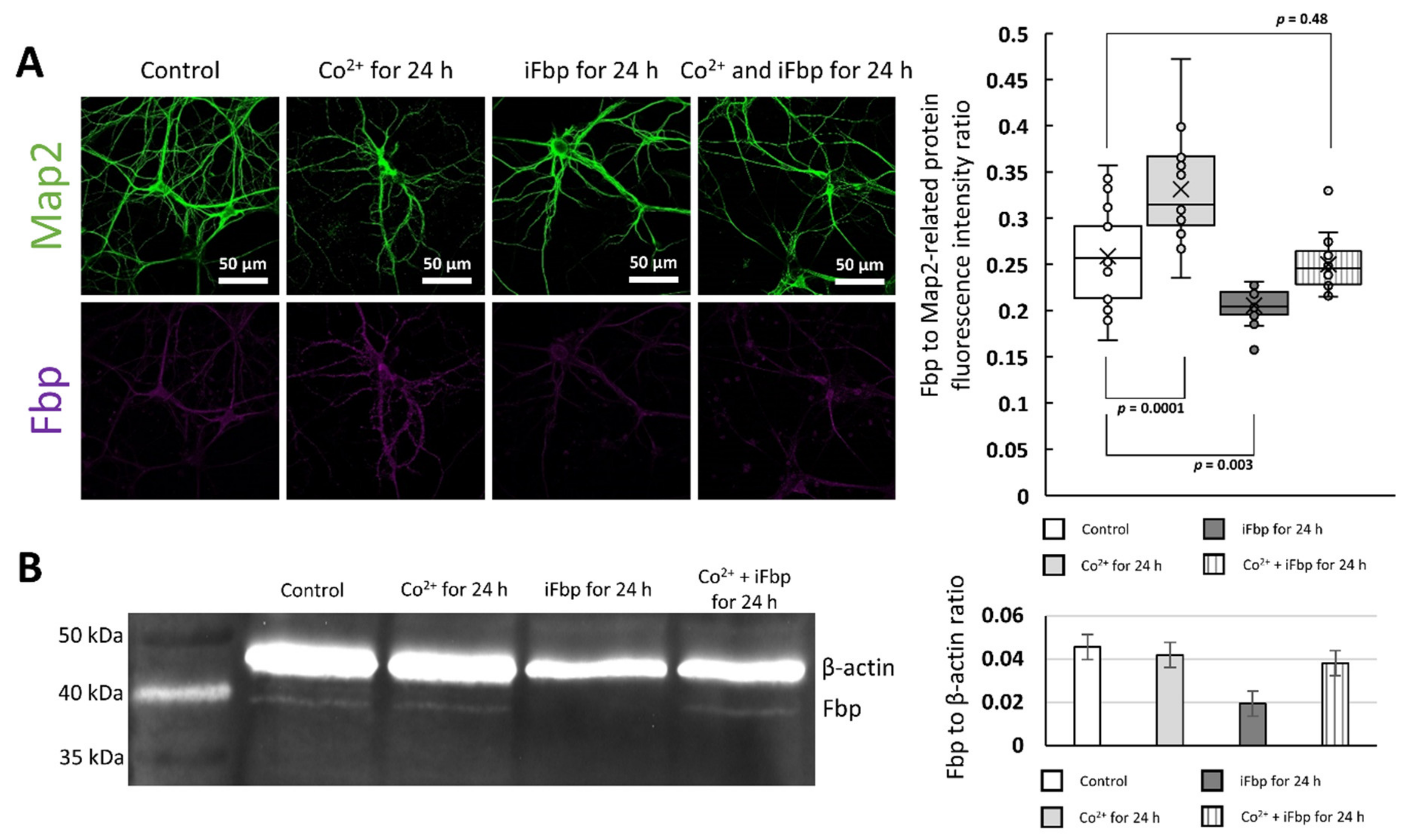
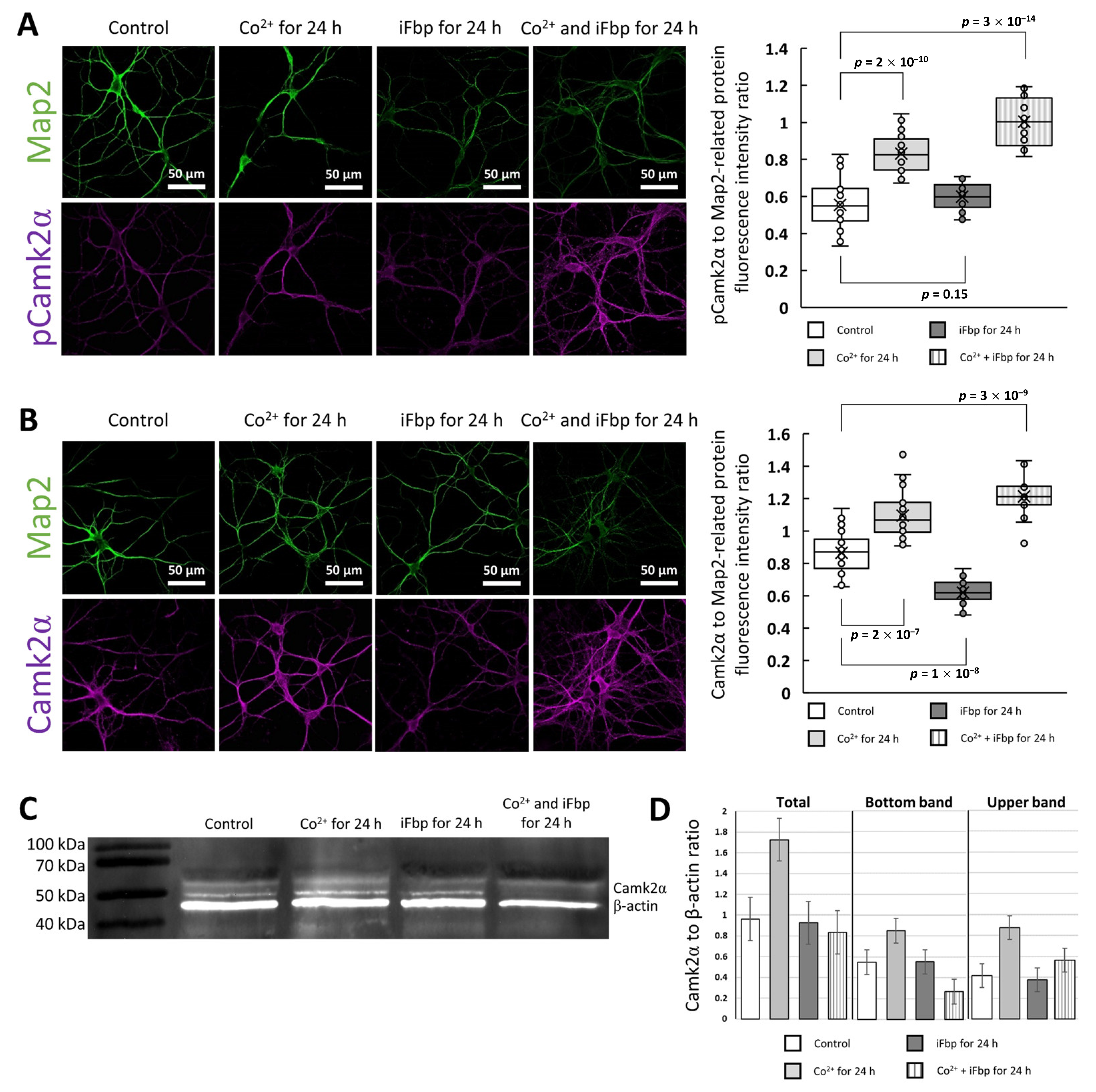
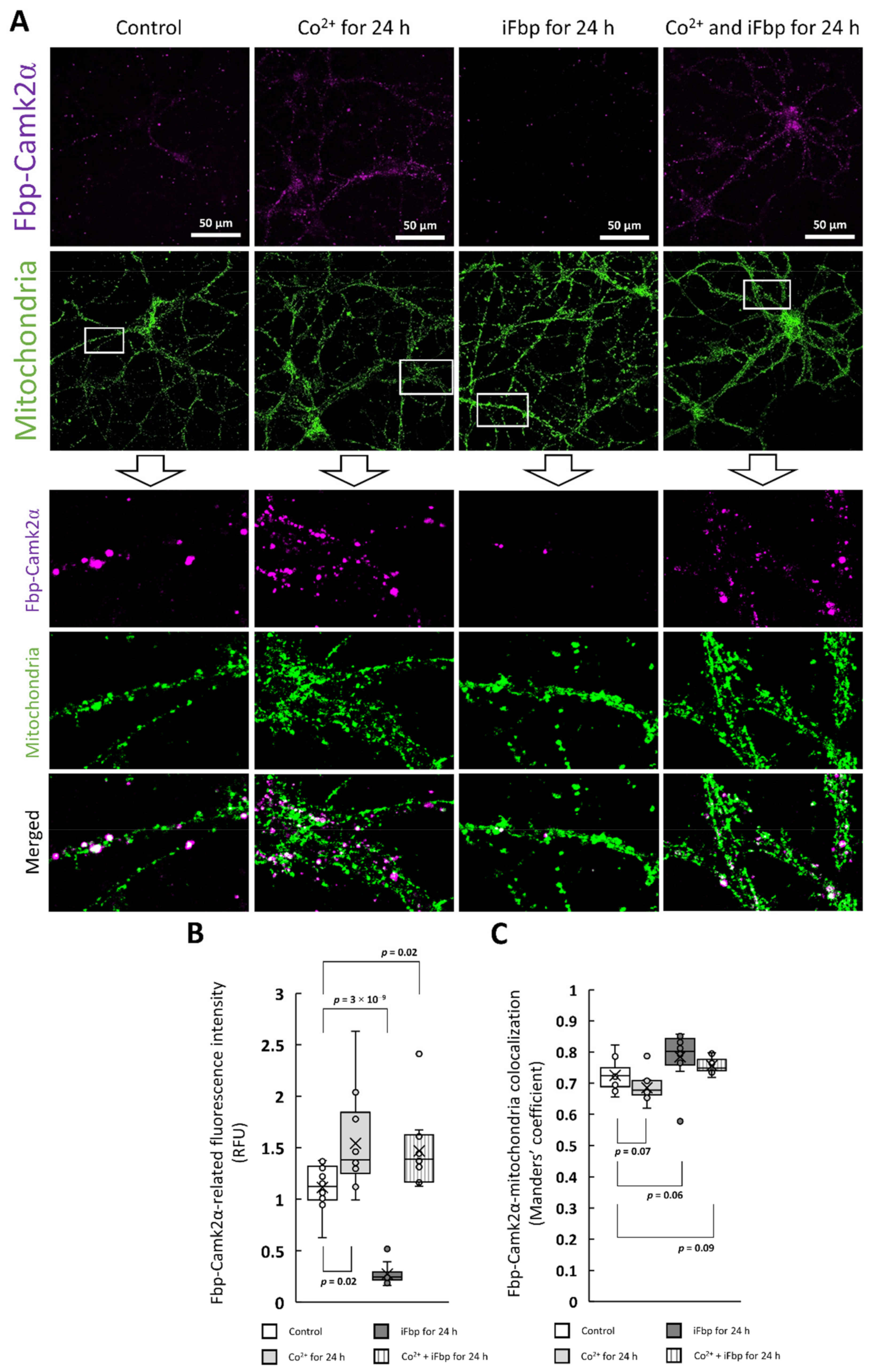
2.2. The Cobalt-Induced Fbp2 Mitochondrial Localization Enhances the Camk2α Autoactivation in Hippocampal Neurons
3. Materials and Methods
3.1. Protein Expression and Purification
3.2. Size-Exclusion Chromatography (SEC)
3.3. Native-PAGE
3.4. Enzymatic Activity Assay
3.5. Cell Culture, LTP Induction, and Fbp2 Expression Silencing
3.6. Chemicals
3.7. Immunofluorescence
3.8. In Situ Detection of Proteins Interaction
3.9. Western Blot
3.10. Confocal Microscopy and Fluorescence Analysis
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gizak, A.; Duda, P.; Wisniewski, J.; Rakus, D. Fructose-1,6-bisphosphatase: From a glucose metabolism enzyme to multifaceted regulator of a cell fate. Adv. Biol. Regul. 2019, 72, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Duda, P.; Wójtowicz, T.; Janczara, J.; Krowarsch, D.; Czyrek, A.; Gizak, A.; Rakus, D. Fructose 1,6-Bisphosphatase 2 Plays a Crucial Role in the Induction and Maintenance of Long-Term Potentiation. Cells 2020, 9, 1375. [Google Scholar] [CrossRef] [PubMed]
- Huangyang, P.; Li, F.; Lee, P.; Nissim, I.; Weljie, A.M.; Mancuso, A.; Li, B.; Keith, B.; Yoon, S.S.; Celeste Simon, M. Fructose-1,6-Bisphosphatase 2 Inhibits Sarcoma Progression by Restraining Mitochondrial Biogenesis. Cell Metab. 2020, 31, 174–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duda, P.; Janczara, J.; McCubrey, J.A.; Gizak, A.; Rakus, D. The Reverse Warburg Effect is Associated with Fbp2-Dependent Hif1α Regulation in Cancer Cells Stimulated by Fibroblasts. Cells 2020, 14, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Qiu, B.; Lee, D.S.M.; Walton, Z.E.; Ochocki, J.D.; Mathew, L.K.; Mancuso, A.; Gade, T.P.F.; Keith, B.; Nissim, I.; et al. Fructose-1,6-bisphosphatase opposes renal carcinoma progression. Nature 2014, 513, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Hajka, D.; Duda, P.; Wójcicka, O.; Drulis-Fajdasz, D.; Rakus, D.; Gizak, A. Expression of FBP2, a newly discovered constituent of memory formation mechanisms, is regulated by astrocyte–neuron crosstalk. Int. J. Mol. Sci. 2020, 21, 6903. [Google Scholar] [CrossRef]
- Gizak, A.; Pirog, M.; Rakus, D. Muscle FBPase binds to cardiomyocyte mitochondria under glycogen synthase kinase-3 inhibition or elevation of cellular Ca 2+ level. FEBS Lett. 2012, 586, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Wiśniewski, J.; Piróg, M.; Hołubowicz, R.; Dobryszycki, P.; McCubrey, J.A.; Rakus, D.; Gizak, A. Dimeric and tetrameric forms of muscle fructose-1,6-bisphosphatase play different roles in the cell. Oncotarget 2017, 8, 115420–115433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barciszewski, J.; Wisniewski, J.; Kolodziejczyk, R.; Jaskolski, M.; Rakus, D.; Dzugaj, A. T-to-R switch of muscle fructose-1,6-bisphosphatase involves fundamental changes of secondary and quaternary structure. Acta. Crystallogr. Sect. D Struct. Biol. 2016, 72, 536–550. [Google Scholar] [CrossRef] [Green Version]
- Rakus, D.; Tillmann, H.; Wysocki, R.; Ulaszewski, S.; Eschrich, K.; Dzugaj, A. Different sensitivities of mutants and chimeric forms of human muscle and liver fructose-1,6-bisphosphatases towards AMP. Biological. Chem. 2003, 384, 51–58. [Google Scholar] [CrossRef]
- Pirog, M.; Gizak, A.; Rakus, D. Changes in quaternary structure of muscle fructose-1,6-bisphosphatase regulate affinity of the enzyme to mitochondria. Int. J. Biochem. Cell Biol. 2014, 48, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Gizak, A.; Zarzycki, M.; Rakus, D. Nuclear targeting of FBPase in HL-1 cells is controlled by beta-1 adrenergic receptor-activated Gs protein signaling cascade. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 871–877. [Google Scholar] [CrossRef] [Green Version]
- Gizak, A.; Majkowski, M.; Dus, D.; Dzugaj, A. Calcium inhibits muscle FBPase and affects its intracellular localization in cardiomyocytes. FEBS Lett. 2004, 576, 445–448. [Google Scholar] [CrossRef] [Green Version]
- Tejwani, G.A.; Pedrosa, F.O.; Pontremoli, S.; Horecker, B.L. Dual role of Zn2+ as inhibitor and activator of fructose 1,6 bisphosphatase of rat liver. Proc. Natl. Acad. Sci. USA 1976, 73, 2692–2695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi-Iñiguez, T.; García-Hernandez, E.; Arreguín-Espinosa, R.; Flores, M.E. Role of Vitamin B12 on methylmalonyl-CoA mutase activity. J. Zhejiang Univ. Sci. B 2012, 13, 423–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Froese, D.S.; Fowler, B.; Baumgartner, M.R. Vitamin B12, folate, and the methionine remethylation cycle—Biochemistry, pathways, and regulation. J. Inherit. Metab. Dis. 2019, 42, 673–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderón-Ospina, C.A.; Nava-Mesa, M.O. B Vitamins in the nervous system: Current knowledge of the biochemical modes of action and synergies of thiamine, pyridoxine, and cobalamin. CNS Neurosci. Ther. 2020, 26, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Catalani, S.; Rizzetti, M.C.; Padovani, A.; Apostoli, P. Neurotoxicity of cobalt. Hum. Exp. Toxicol. 2012, 31, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Ensinger, H.A.; Pauly, H.E.; Pfleiderer, G.; Stiefel, T. The role of Zn(II) in calf intestinal alkaline phosphatase studied by the influence of chelating agents and chemical modification of histidine residues. Biochim. Biophys. Acta. 1978, 527, 432–441. [Google Scholar] [CrossRef]
- Mao, X.; Wong, A.A.; Crawford, R.W. Cobalt toxicity—An emerging clinical problem in patients with metal-on-metal hip prostheses? Med. J. Aust. 2011, 20, 649–651. [Google Scholar] [CrossRef] [Green Version]
- Schirrmacher, U.O.E. Case of Cobalt Poisoning. Br. Med. J. 1967, 1, 544–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Licht, A.; Oliver, M.; Rachmilewitz, E.A. Optic atrophy following treatment with cobalt chloride in a patient with pancytopenia and hypercellular marrow. Isr. J. Med. Sci. 1972, 8, 61–66. [Google Scholar]
- Dow, R.S.; Fernández-Guardiola, A.; Manni, E. The production of cobalt experimental epilepsy in the rat. Electroencephalogr. Clin. Neurophysiol. 1962, 14, 399–407. [Google Scholar] [CrossRef]
- Mutani, R. Cobalt Experimental Hippocampal Epilepsy in the Cat. Epilepsia 1967, 8, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Grimm, R.J.; Frazee, J.G.; Kawasaki, T.; Savić, M. Cobalt epilepsy in the squirrel monkey. Electroencephalogr. Clin. Neurophysiol. 1970, 29, 525–528. [Google Scholar] [CrossRef]
- Chang, J.H.; Yang, X.F.; Zempel, J.M.; Rothman, S.M. The unilateral cobalt wire model of neocortical epilepsy: A method of producing subacute focal seizures in rodents. Epilepsy Res. 2004, 61, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Craig, C.R.; Colasanti, B.K. GABA receptors, lipids, and gangliosides in cobalt epileptic focus. Adv. Neurol. 1986, 44, 379–391. [Google Scholar] [PubMed]
- He, J.; Hsiang, H.L.; Wu, C.; Mylvagnanam, S.; Carlen, P.L.; Zhang, L. Cellular mechanisms of cobalt-induced hippocampal epileptiform discharges. Epilepsia 2009, 50, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.; Gum, R.J.; Daly, M.; Fry, E.H.; Hutchins, C.; Abad-Zapatero, C.; von Geldern, T.W. Benzoxazole benzenesulfonamides as allosteric inhibitors of fructose-1,6-bisphosphatase. Bioorg. Med. Chem. Lett. 2006, 16, 1807–1810. [Google Scholar] [CrossRef]
- Von Geldern, T.W.; Lai, C.; Gum, R.J.; Daly, M.; Sun, C.; Fry, E.H.; Abad-Zapatero, C. Benzoxazole benzenesulfonamides are novel allosteric inhibitors of fructose-1,6-bisphosphatase with a distinct binding mode. Bioorg. Med. Chem. Lett. 2006, 16, 1811–1815. [Google Scholar] [CrossRef]
- Pedrosa, F.O.; Pontremoli, S.; Horecker, B.L. Binding of Zn2+ to rat liver fructose-1,6-bisphosphatase and its effect on the catalytic properties. Proc. Natl. Acad. Sci. USA 1977, 74, 2742–2745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choe, J.Y.; Fromm, H.J.; Honzatko, R.B. Crystal structures of fructose 1,6-bisphosphatase: Mechanism of catalysis and allosteric inhibition revealed in product complexes. Biochemistry 2000, 39, 8565–8574. [Google Scholar] [CrossRef] [PubMed]
- Tejwani, G.A. Regulation of fructose-bisphosphatase activity. Adv. Enzymol. Relat. Areas Mol Biol. 1983, 54, 121–194. [Google Scholar] [CrossRef]
- Rulisek, L.; Vondrasek, J. Coordination geometries of selected transition metal ion (Co2+, Ni2+, Cu2+, Zn2+, Cd2+, and Hg2+) in metalloproteins. J. Inorg. Biochem. 1998, 71, 115–127. [Google Scholar] [CrossRef]
- Sellin, S.; Mannervik, B. Metal dissociation constants for glyoxalase I reconstituted with Zn2+, Co2+, Mn2+, and Mg2+. J. Biol. Chem. 1984, 259, 11426–11429. [Google Scholar] [CrossRef]
- Schneider, G.; Zeppezauer, M. Kinetics and mechanisms of the recombination of Zn2+, Co2+, and Ni2+ with the metal-depleted catalytic site of horse liver alcohol dehydrogenase. J. Inorg. Bioch. 1983, 18, 59–69. [Google Scholar] [CrossRef]
- Hanada, T. Ionotropic glutamate receptors in epilepsy: A review focusing on ampa and nmda receptors. Biomolecules 2020, 18, 464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lua, W.; Isozaki, K.; Roche, K.W.; Nicoll, R.A. Synaptic targeting of AMPA receptors is regulated by a CaMKII site in the first intracellular loop of GluA1. Proc. Natl. Acad. Sci. USA 2010, 107, 22266–22271. [Google Scholar] [CrossRef] [Green Version]
- Barria, A.; Muller, D.; Derkach, V.; Griffith, L.C.; Soderling, T.R. Regulatory phosphorylation of AMPA-type glutamate receptors by CaM-KII during long-term potentiation. Science 1997, 276, 2042–2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristensen, A.S.; Jenkins, M.A.; Banke, T.G.; Schousboe, A.; Makino, Y.; Johnson, R.C.; Huganir, R.; Traynelis, S.F. Mechanism of Ca2+/calmodulin-dependent kinase II regulation of AMPA receptor gating. Nat. Neurosci. 2011, 14, 727–735. [Google Scholar] [CrossRef]
- Carruthers, N.J.; Parker, G.C.; Gratsch, T.; Caruso, J.A.; Stemmer, P.M. Protein mobility shifts contribute to gel electrophoresis liquid chromatography analysis. J. Biomol. Tech. 2015, 26, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, J.B.; Zaegel, V.; Coultrap, S.J.; Miller, A.P.; Bayer, K.U.; Reichow, S.L. The CaMKII holoenzyme structure in activation-competent conformations. Nat. Commun. 2017, 8, 15742. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, M.; Stratton, M.M.; Going, C.C.; McSpadden, E.D.; Huang, Y.; Susa, A.C.; Elleman, A.; Cao, Y.M.; Pappireddi, N.; Burkhardt, P.; et al. Molecular mechanism of activation-triggered subunit exchange in Ca2+/calmodulin-dependent protein kinase II. Elife 2016, 5, e13405. [Google Scholar] [CrossRef]
- Skalecki, K.; Mularczyk, W.; Dzugaj, A. Kinetic properties of D-fructose-1,6-bisphosphate 1-phosphohydrolase isolated from human muscle. Biochem. J. 1995, 310, 1029–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mozrzymas, J.; Szczęsny, T.; Rakus, D. The effect of glycogen phosphorolysis on basal glutaminergic transmission. Biochem. Biophys Res. Commun. 2011, 404, 652–655. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Man, H.; Ju, W.; Trimble, W.S.; MacDonald, J.F.; Wang, Y.T. Activation of synaptic NMDA receptors induces membrane insertion of new AMPA receptors and LTP in cultured hippocampal neurons. Neuron 2001, 29, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.-Q.; Wang, S.-H.; Yao, W.; Wang, J.-J.; Ji, F.; Yan, J.-Z.; Ren, S.-Q.; Chen, Z.; Liu, S.-Y.; Lu, W. Role of glycine receptors in glycine-induced LTD in hippocampal CA1 pyramidal neurons. Neuropsychopharmacology 2011, 36, 1948–1958. [Google Scholar] [CrossRef] [Green Version]
- Mamczur, P.; Borsuk, B.; Paszko, J.; Sas, Z.; Mozrzymas, J.; Wiśniewski, J.R.; Gizak, A.; Rakus, D. Astrocyte-neuron crosstalk regulates the expression and subcellular localization of carbohydrate metabolism enzymes. Glia 2015, 63, 328–340. [Google Scholar] [CrossRef]
- Gizak, A.; Dzugaj, A. FBPase is in the nuclei of cardiomyocytes. FEBS Lett. 2003, 539, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Bolte, S.; Cordelières, F.P. A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef] [PubMed]
- Dunn, K.W.; Kamocka, M.M.; McDonald, J.H. A practical guide to evaluating colocalization in biological microscopy. Am. J. Physiol. Cell Physiol. 2011, 300, C723–C742. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duda, P.; Budziak, B.; Rakus, D. Cobalt Regulates Activation of Camk2α in Neurons by Influencing Fructose 1,6-Bisphosphatase 2 Quaternary Structure and Subcellular Localization. Int. J. Mol. Sci. 2021, 22, 4800. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094800
Duda P, Budziak B, Rakus D. Cobalt Regulates Activation of Camk2α in Neurons by Influencing Fructose 1,6-Bisphosphatase 2 Quaternary Structure and Subcellular Localization. International Journal of Molecular Sciences. 2021; 22(9):4800. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094800
Chicago/Turabian StyleDuda, Przemysław, Bartosz Budziak, and Dariusz Rakus. 2021. "Cobalt Regulates Activation of Camk2α in Neurons by Influencing Fructose 1,6-Bisphosphatase 2 Quaternary Structure and Subcellular Localization" International Journal of Molecular Sciences 22, no. 9: 4800. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094800