
Insulin Resistance Is Not Sustained Following Denervation in Glycolytic Skeletal Muscle

 , ,
, ,
Abstract
:1. Introduction
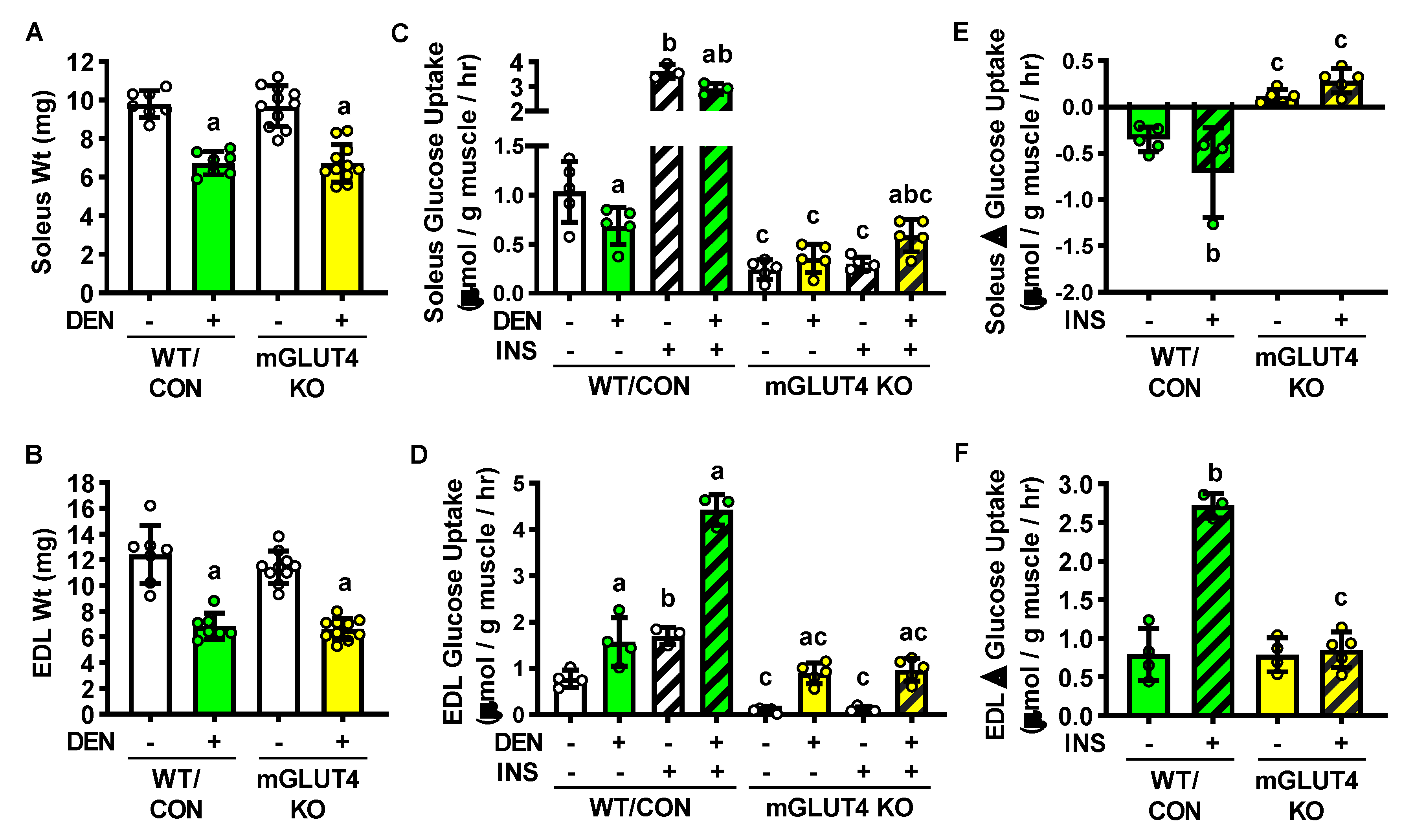
2. Results
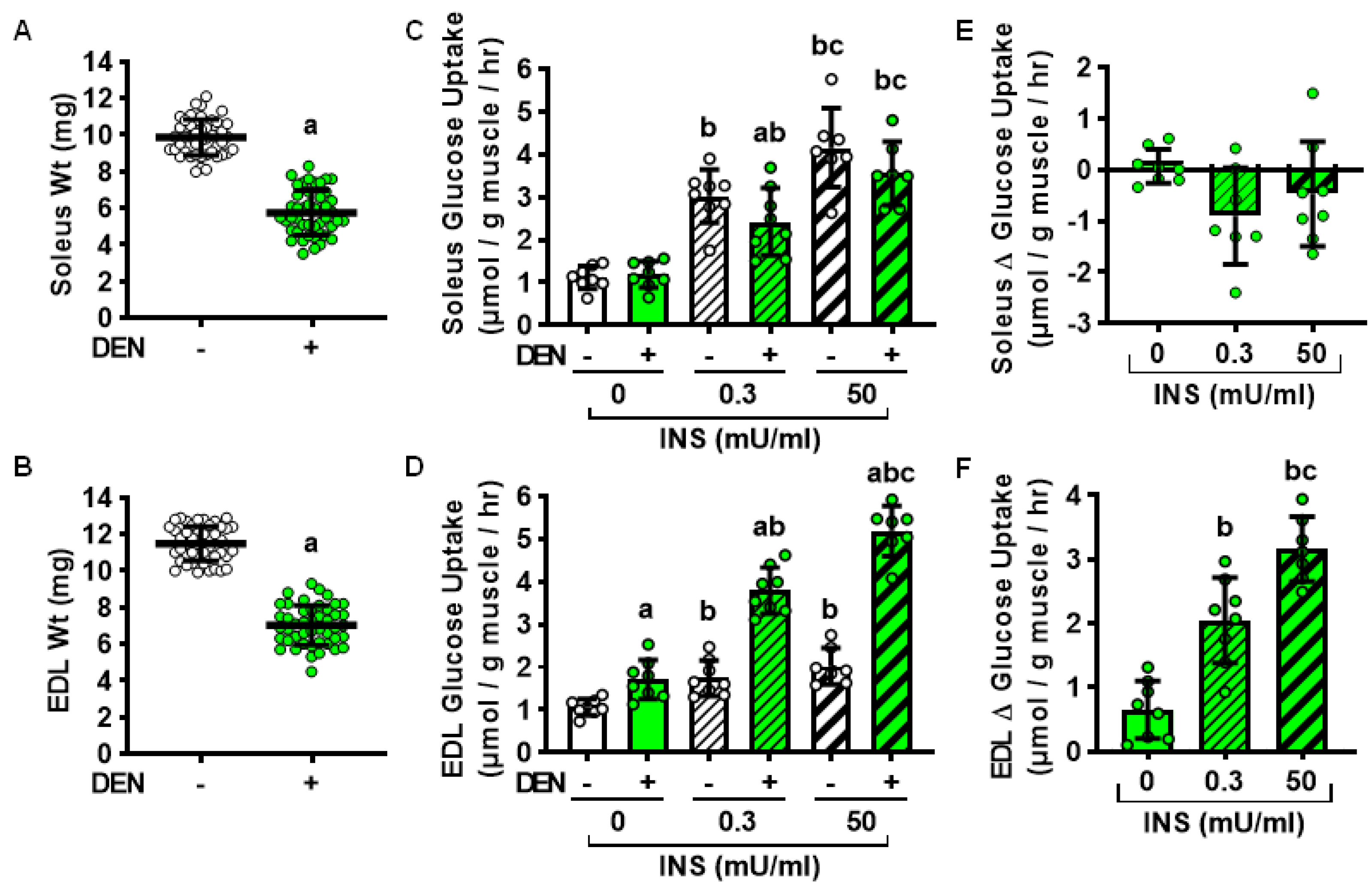
2.1. Effects of Long-Term Denervation on Skeletal Muscle Insulin Sensitivity
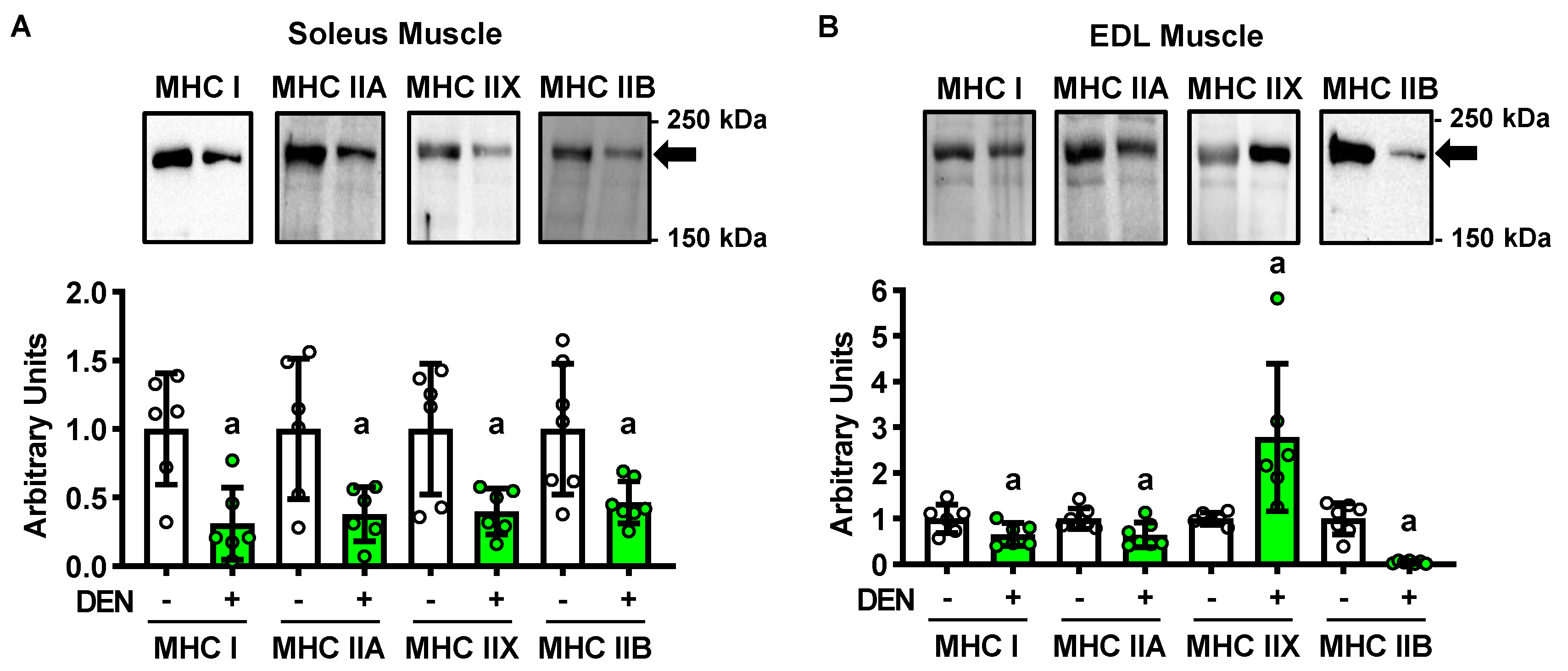
2.2. Long-Term Denervation and Muscle Fiber Type
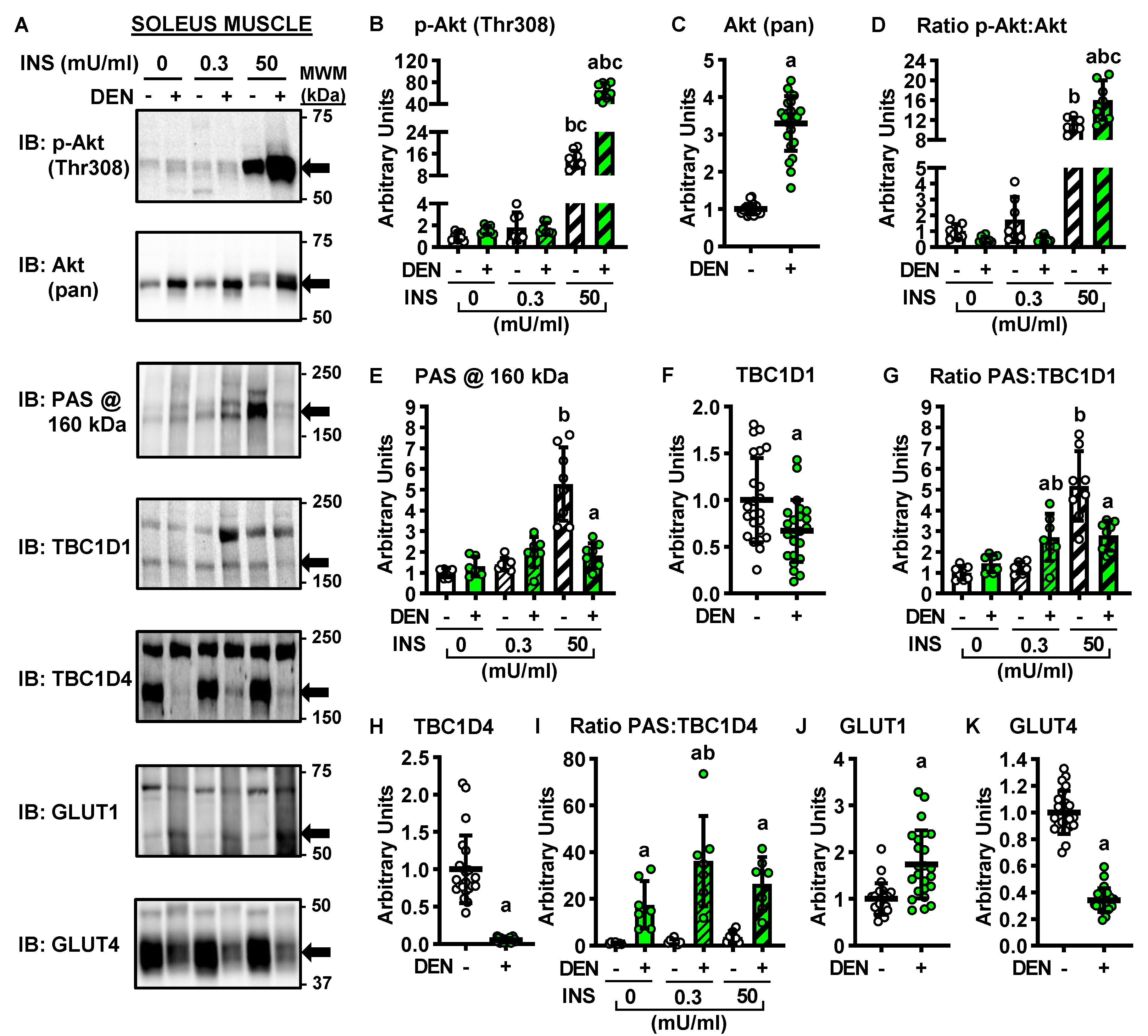
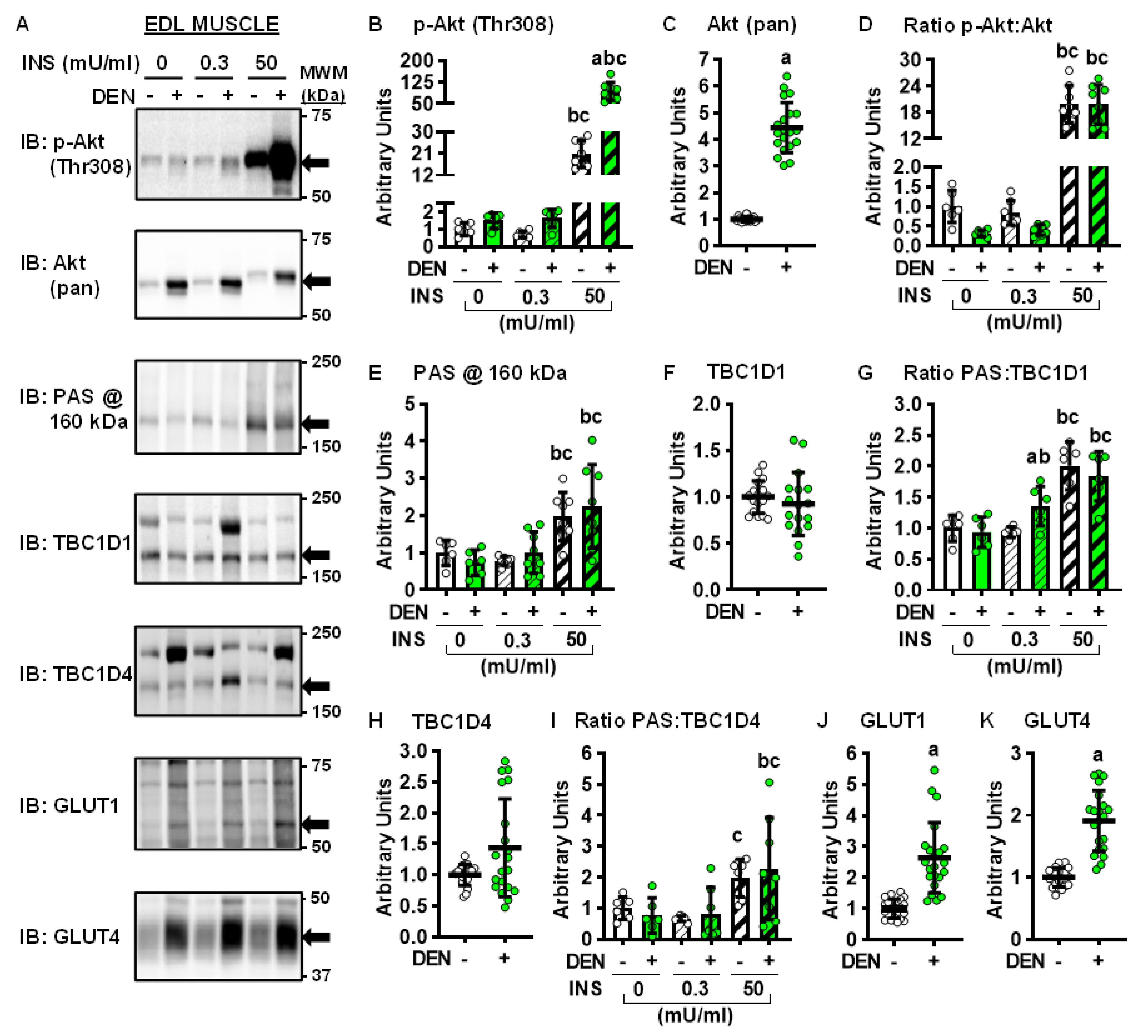
2.3. Long-Term Denervation and the Insulin-Signaling Cascade
2.4. Long-Term Denervation and Muscle Glucose Transporters
2.5. Long-term denervation and muscle-specific GLUT4 knockout mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Unilateral Denervation Surgery
4.3. Muscle [3H]-2-Deoxyglucose Uptake
4.4. Immunoblot Analyses
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DEN | Denervation |
| EDL | Extensor digitorum longus |
| GLUT | Glucose transporter |
| INS | Insulin |
| KRB | Krebs-Ringer Bicarbonate |
| mGLUT4 KO | Muscle-specific glucose transporter 4 knockout |
| MHC | Myosin heavy chain |
| PAS | Phospho-Akt substrate |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| WT/CON | Wild-type/control |
References
- Buse, M.G.; Buse, J. Glucose uptake and response to insulin of the isolated rat diaphragm: The effect of denervation. Diabetes 1959, 8, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Burant, C.F.; Lemmon, S.K.; Treutelaar, M.K.; Buse, M.G. Insulin resistance of denervated rat muscle: A model for impaired receptor-function coupling. Am. J. Physiol. 1984, 247, E657–E666. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.L.; Lawrence, J.C., Jr. Insulin action in denervated rat hemidiaphragms. Decreased hormonal stimulation of glycogen synthesis involves both glycogen synthase and glucose transport. J. Biol. Chem. 1984, 259, 2201–2207. [Google Scholar] [CrossRef]
- Turinsky, J. Dynamics of insulin resistance in denervated slow and fast muscles in vivo. Am. J. Physiol. 1987, 252, R531–R537. [Google Scholar] [CrossRef] [PubMed]
- Turinsky, J. Glucose and amino acid uptake by exercising muscles in vivo: Effect of insulin, fiber population, and denervation. Endocrinology 1987, 121, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Karl, I.E. Resistance of protein and glucose metabolism to insulin in denervated rat muscle. Biochem. J. 1988, 254, 667–675. [Google Scholar] [CrossRef] [Green Version]
- Sowell, M.O.; Dutton, S.L.; Buse, M.G. Selective in vitro reversal of the insulin resistance of glucose transport in denervated rat skeletal muscle. Am. J. Physiol. 1989, 257, E418–E425. [Google Scholar] [CrossRef]
- Henriksen, E.J.; Rodnick, K.J.; Mondon, C.E.; James, D.E.; Holloszy, J.O. Effect of denervation or unweighting on GLUT-4 protein in rat soleus muscle. J. Appl. Physiol. 1991, 70, 2322–2327. [Google Scholar] [CrossRef]
- Coderre, L.; Monfar, M.M.; Chen, K.S.; Heydrick, S.J.; Kurowski, T.G.; Ruderman, N.B.; Pilch, P.F. Alteration in the expression of GLUT-1 and GLUT-4 protein and messenger RNA levels in denervated rat muscles. Endocrinology 1992, 131, 1821–1825. [Google Scholar] [CrossRef]
- Megeney, L.A.; Michel, R.N.; Boudreau, C.S.; Fernando, P.K.; Prasad, M.; Tan, M.H.; Bonen, A. Regulation of muscle glucose transport and GLUT-4 by nerve-derived factors and activity-related processes. Am. J. Physiol. 1995, 269, R1148–R1153. [Google Scholar] [CrossRef] [PubMed]
- Megeney, L.A.; Neufer, P.D.; Dohm, G.L.; Tan, M.H.; Blewett, C.A.; Elder, G.C.; Bonen, A. Effects of muscle activity and fiber composition on glucose transport and GLUT-4. Am. J. Physiol. 1993, 264, E583–E593. [Google Scholar] [CrossRef]
- Handberg, A.; Megeney, L.A.; McCullagh, K.J.; Kayser, L.; Han, X.X.; Bonen, A. Reciprocal GLUT-1 and GLUT-4 expression and glucose transport in denervated muscles. Am. J. Physiol. 1996, 271, E50–E57. [Google Scholar] [CrossRef] [PubMed]
- Elmendorf, J.S.; Damrau-Abney, A.; Smith, T.R.; David, T.S.; Turinsky, J. Phosphatidylinositol 3-kinase and dynamics of insulin resistance in denervated slow and fast muscles in vivo. Am. J. Physiol. 1997, 272, E661–E670. [Google Scholar] [CrossRef]
- Turinsky, J.; Damrau-Abney, A.; Loegering, D.J. Blood flow and glucose uptake in denervated, insulin-resistant muscles. Am. J. Physiol. 1998, 274, R311–R317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkes, J.J.; Bonen, A. Reduced insulin-stimulated glucose transport in denervated muscle is associated with impaired Akt-alpha activation. Am. J. Physiol. Metab. 2000, 279, E912–E919. [Google Scholar] [CrossRef] [Green Version]
- Aksnes, A.K.; Hjeltnes, N.; Wahlstrom, E.O.; Katz, A.; Zierath, J.R.; Wallberg-Henriksson, H. Intact glucose transport in morphologically altered denervated skeletal muscle from quadriplegic patients. Am. J. Physiol. 1996, 271, E593–E600. [Google Scholar] [CrossRef] [PubMed]
- Pak, K.; Shin, M.J.; Hwang, S.-J.; Shin, J.-H.; Shin, H.K.; Kim, S.J.; Kim, I.J. Longitudinal Changes in Glucose Metabolism of Denervated Muscle after Complete Peripheral Nerve Injury. Mol. Imaging Biol. 2016, 18, 741–747. [Google Scholar] [CrossRef]
- Nunes, W.M.S.; Mello, M.A.R.D. Glucose metabolism in rats submitted to skeletal muscle denervation. Braz. Arch. Biol. Technol. 2005, 48, 541–548. [Google Scholar] [CrossRef]
- Callahan, Z.J.; Oxendine, M.; Wheatley, J.L.; Menke, C.; Cassell, E.A.; Bartos, A.; Geiger, P.C.; Schaeffer, P.J. Compensatory responses of the insulin signaling pathway restore muscle glucose uptake following long-term denervation. Physiol. Rep. 2015, 3. [Google Scholar] [CrossRef]
- Turinsky, J.; Damrau-Abney, A. Akt1 kinase and dynamics of insulin resistance in denervated muscles in vivo. Am. J. Physiol. 1998, 275, R1425–R1430. [Google Scholar] [CrossRef]
- Block, N.E.; Menick, D.R.; Robinson, K.A.; Buse, M.G. Effect of denervation on the expression of two glucose transporter isoforms in rat hindlimb muscle. J. Clin. Investig. 1991, 88, 1546–1552. [Google Scholar] [CrossRef]
- Jones, J.P.; Tapscott, E.B.; Olson, A.L.; Pessin, J.E.; Dohm, G.L. Regulation of glucose transporters GLUT-4 and GLUT-1 gene transcription in denervated skeletal muscle. J. Appl. Physiol. 1998, 84, 1661–1666. [Google Scholar] [CrossRef]
- Fogt, D.L.; Slentz, M.J.; Tischler, M.E.; Henriksen, E.J. GLUT-4 protein and citrate synthase activity in distally or proximally denervated rat soleus muscle. Am. J. Physiol. 1997, 272, R429–R432. [Google Scholar] [CrossRef]
- Zhou, M.; Vallega, G.; Kandror, K.V.; Pilch, P.F. Insulin-mediated translocation of GLUT-4-containing vesicles is preserved in denervated muscles. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E1019–E1026. [Google Scholar] [CrossRef] [Green Version]
- Cartee, G.D.; Arias, E.B.; Yu, C.S.; Pataky, M.W. Novel single skeletal muscle fiber analysis reveals a fiber type-selective effect of acute exercise on glucose uptake. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E818–E824. [Google Scholar] [CrossRef] [PubMed]
- Roseno, S.L.; Davis, P.R.; Bollinger, L.M.; Powell, J.J.; Witczak, C.A.; Brault, J.J. Short-term, high-fat diet accelerates disuse atrophy and protein degradation in a muscle-specific manner in mice. Nutr. Metab. 2015, 12, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloemberg, D.; Quadrilatero, J. Rapid determination of myosin heavy chain expression in rat, mouse, and human skeletal muscle using multicolor immunofluorescence analysis. PLoS ONE 2012, 7, e35273. [Google Scholar] [CrossRef] [PubMed]
- Cartee, G.D. Roles of TBC1D1 and TBC1D4 in insulin- and exercise-stimulated glucose transport of skeletal muscle. Diabetologia 2015, 58, 19–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumiya, Y.; Hopkins, T.; Morris, C.; Sato, K.; Zeng, L.; Viereck, J.; Hamilton, J.A.; Ouchi, N.; LeBrasseur, N.K.; Walsh, K. Fast/Glycolytic muscle fiber growth reduces fat mass and improves metabolic parameters in obese mice. Cell Metab. 2008, 7, 159–172. [Google Scholar] [CrossRef] [Green Version]
- Kramer, H.F.; Witczak, C.A.; Taylor, E.B.; Fujii, N.; Hirshman, M.F.; Goodyear, L.J. AS160 regulates insulin- and contraction-stimulated glucose uptake in mouse skeletal muscle. J. Biol. Chem. 2006, 281, 31478–31485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, D.; Toyoda, T.; Taylor, E.B.; Yu, H.; Fujii, N.; Hirshman, M.F.; Goodyear, L.J. TBC1D1 regulates insulin- and contraction-induced glucose transport in mouse skeletal muscle. Diabetes 2010, 59, 1358–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zisman, A.; Peroni, O.D.; Abel, E.D.; Michael, M.D.; Mauvais-Jarvis, F.; Lowell, B.B.; Wojtaszewski, J.F.; Hirshman, M.F.; Virkamaki, A.; Goodyear, L.J.; et al. Targeted disruption of the glucose transporter 4 selectively in muscle causes insulin resistance and glucose intolerance. Nat. Med. 2000, 6, 924–928. [Google Scholar] [CrossRef]
- Andrisse, S.; Patel, G.D.; Chen, J.E.; Webber, A.M.; Spears, L.D.; Koehler, R.M.; Robinson-Hill, R.M.; Ching, J.K.; Jeong, I.; Fisher, J.S. ATM and GLUT1-S490 phosphorylation regulate GLUT1 mediated transport in skeletal muscle. PLoS ONE 2013, 8, e66027. [Google Scholar] [CrossRef] [Green Version]
- McMillin, S.L.; Schmidt, D.L.; Kahn, B.B.; Witczak, C.A. GLUT4 Is Not Necessary for Overload-Induced Glucose Uptake or Hypertrophic Growth in Mouse Skeletal Muscle. Diabetes 2017, 66, 1491–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espelage, L.; Al-Hasani, H.; Chadt, A. RabGAPs in skeletal muscle function and exercise. J. Mol. Endocrinol 2020, 64, R1–R19. [Google Scholar] [CrossRef] [PubMed]
- Pigna, E.; Renzini, A.; Greco, E.; Simonazzi, E.; Fulle, S.; Mancinelli, R.; Moresi, V.; Adamo, S. HDAC4 preserves skeletal muscle structure following long-term denervation by mediating distinct cellular responses. Skelet. Muscle 2018, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Abel, E.D.; Kaulbach, H.C.; Tian, R.; Hopkins, J.C.; Duffy, J.; Doetschman, T.; Minnemann, T.; Boers, M.E.; Hadro, E.; Oberste-Berghaus, C.; et al. Cardiac hypertrophy with preserved contractile function after selective deletion of GLUT4 from the heart. J. Clin. Investig. 1999, 104, 1703–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinkley, J.M.; Ferey, J.L.; Brault, J.J.; Smith, C.S.; Gilliam, L.A.A.; Witczak, C.A. Constitutively active CaMKKalpha stimulates skeletal muscle glucose uptake in insulin resistant mice in vivo. Diabetes 2014, 63, 142–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferey, J.L.; Brault, J.J.; Smith, C.A.; Witczak, C.A. Constitutive Activation of CaMKKalpha Signaling is Sufficient but Not Necessary for mTORC1 Activation and Growth in Mouse Skeletal Muscle. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E686–E694. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Blocking | 1° Antibody | 1° Antibody RRID | 2° Antibody | ECL Reagent |
|---|---|---|---|---|---|
| Akt (11E7) | 5% BSA | 1:2000 in 5% BSA, cat#4685, lot#6, Cell Signaling Technology, Danvers, MA, USA | AB_2225340 | 1:2000 Rabbit-HRP | Western Lightning™ |
| p-Akt (Thr308) | 5% BSA | 1:2000 in 5% BSA, cat#9275, lot#20, Cell Signaling Technology, Danvers, MA, USA | AB_329828 | 1:2000 Rabbit-HRP | Western Lightning™ |
| p-Akt Substrate (PAS) | 5% BSA | 1:2000 in 5% BSA, cat#9611, lot#11, Cell Signaling Technology, Danvers, MA, USA | AB_330302 | 1:2000 Rabbit-HRP | Western Lightning™ |
| GLUT1 | 5% milk | 1:4000 in 5% BSA, cat#07-1401, lot#2956779, Millipore, St. Louis, MO, USA | AB_1587074 | 1:2000 Rabbit-HRP | Western Lightning™ |
| GLUT4 | 5% BSA | 1:2000 in 5% BSA, cat#07-1404, lot#2890837 and 2987632, Millipore, St. Louis, MO, USA | AB_1587080 | 1:2000 Rabbit-HRP | Western Lightning™ |
| MHC Type I | 5% BSA | 1:500 in 5% BSA, cat#BA-F8, lot# N/A,Developmental Studies Hybridoma Bank, University of Iowa, Iowa City, IA, USA | AB_10572253 | 1:5000 Mouse-HRP | EDL: Western Lightning™ Soleus: Super Signal™ |
| MHC Type IIA | 5% milk | 1:500 in 5% BSA, cat#SC-71-S, lot# N/A, Developmental Studies Hybridoma Bank, University of Iowa, Iowa City, IA, USA | AB_2147165 | 1:5000 Mouse-HRP | Western Lightning™ |
| MHC Type IIB | 5% BSA | 1:100 in 5% BSA, cat#BF-F3, lot# N/A, Developmental Studies Hybridoma Bank, University of Iowa, Iowa City, IA, USA | AB_2266724 | 1:5000 Mouse-HRP | EDL: Super Signal™ Soleus: Western Lightning™ |
| MHC Type IIX | 5% BSA | 1:500 in 5% BSA, cat#6H1, lot# N/A, Developmental Studies Hybridoma Bank, University of Iowa, Iowa City, IA, USA | AB_1157897 | 1:5000 Mouse-HRP | Western Lightning™ |
| TBC1D1 (V796) | 5% BSA | 1:2000 in 5% BSA, cat#4629, lot#1, Cell Signaling Technology, Danvers, MA, USA | AB_1904162 | 1:2000 Rabbit-HRP | Super Signal™ |
| TBC1D4 | 5% BSA | 1:1000 in 5% BSA, cat#07-741, lot#1962662, Millipore, St. Louis, MO, USA | AB_492639 | 1:2000 Rabbit-HRP | Western Lightning™ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McMillin, S.L.; Stanley, E.C.; Weyrauch, L.A.; Brault, J.J.; Kahn, B.B.; Witczak, C.A. Insulin Resistance Is Not Sustained Following Denervation in Glycolytic Skeletal Muscle. Int. J. Mol. Sci. 2021, 22, 4913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094913
McMillin SL, Stanley EC, Weyrauch LA, Brault JJ, Kahn BB, Witczak CA. Insulin Resistance Is Not Sustained Following Denervation in Glycolytic Skeletal Muscle. International Journal of Molecular Sciences. 2021; 22(9):4913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094913
Chicago/Turabian StyleMcMillin, Shawna L., Erin C. Stanley, Luke A. Weyrauch, Jeffrey J. Brault, Barbara B. Kahn, and Carol A. Witczak. 2021. "Insulin Resistance Is Not Sustained Following Denervation in Glycolytic Skeletal Muscle" International Journal of Molecular Sciences 22, no. 9: 4913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094913