
Strategies to Tackle Antimicrobial Resistance: The Example of Escherichia coli and Pseudomonas aeruginosa

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Epidemiology and Pathogenesis of Pseudomonas aeruginosa and Escherichia coli
2.1. Pseudomonas aeruginosa
2.2. Escherichia coli
3. State of the Art of Vaccines and mAbs to Tackle Pseudomonas aeruginosa and Escherichia coli
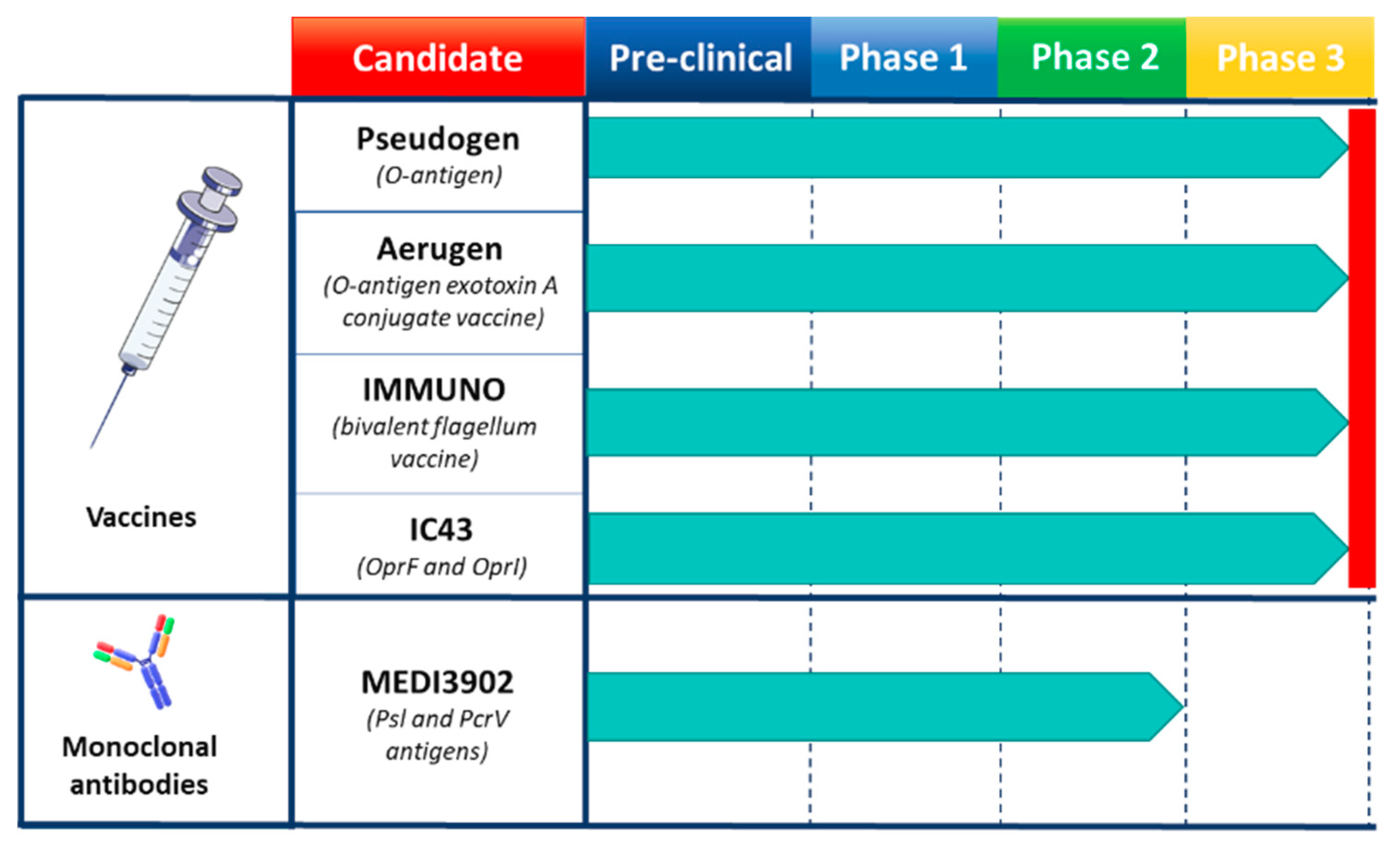
3.1. Pseudomonas aeuroginosa Vaccines and mAbs
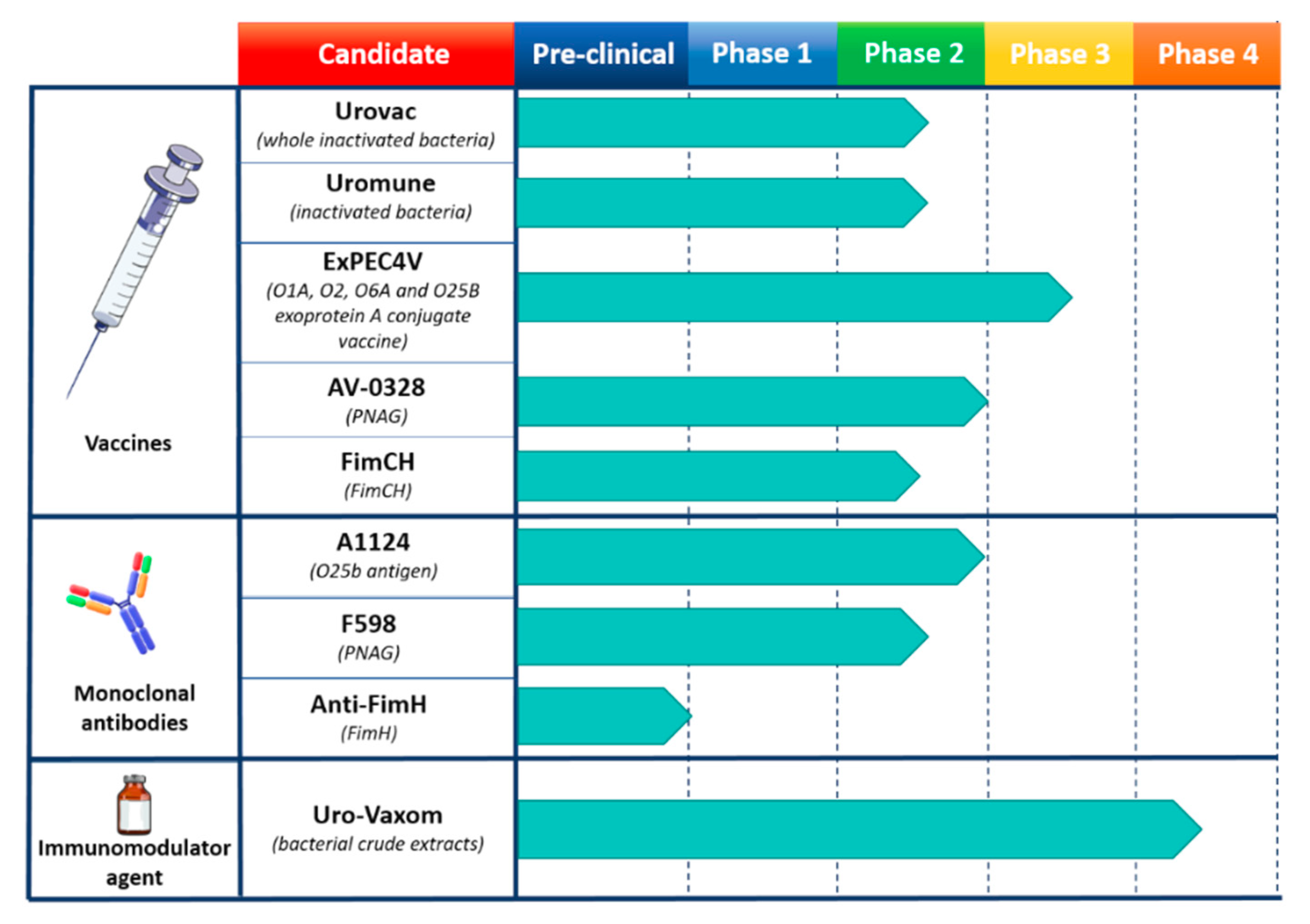
3.2. Escherichia coli Vaccines and mAbs
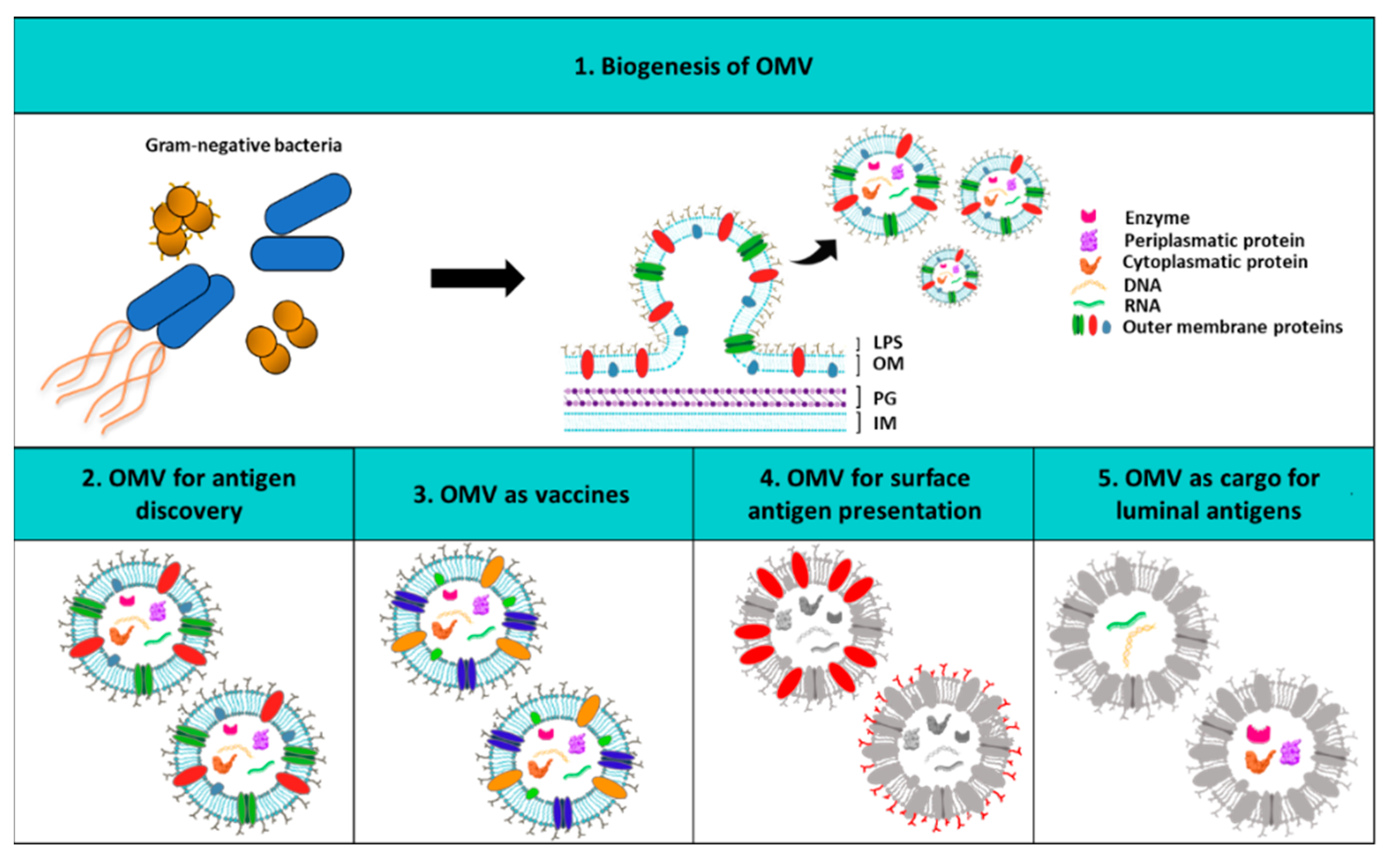
4. Outer Membrane Vesicles as Vaccine Platform
5. Gram-Negative AMR Future Prospective
Funding
Acknowledgments
Conflicts of Interest
References
- Christensen, K.; Doblhammer, G.; Rau, R.; Vaupel, J.W. Ageing populations: The challenges ahead. Lancet 2009, 374, 1196–1208. [Google Scholar] [CrossRef] [Green Version]
- Rappuoli, R. Twenty-first century vaccines. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2756–2758. [Google Scholar] [CrossRef] [Green Version]
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Delany, I.; Rappuoli, R.; Seib, K.L. Vaccines, Reverse Vaccinology, and Bacterial Pathogenesis. Cold Spring Harb. Perspect. Med. 2013, 3, a012476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosini, R.; Nicchi, S.; Pizza, M.; Rappuoli, R. Vaccines Against Antimicrobial Resistance. Front. Immunol. 2020, 11, 1048. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Alekshun, M.N.; Levy, S.B. Molecular Mechanisms of Antibacterial Multidrug Resistance. Cell 2007, 128, 1037–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedijanto, C.; Olesen, S.W.; Grad, Y.H.; Lipsitch, M. Estimating the proportion of bystander selection for antibiotic resistance among potentially pathogenic bacterial flora. Proc. Natl. Acad. Sci. USA 2018, 115, E11988–E11995. [Google Scholar] [CrossRef] [Green Version]
- Blake, K.L.; O’Neill, A.J. Transposon library screening for identification of genetic loci participating in intrinsic susceptibility and acquired resistance to antistaphylococcal agents. J. Antimicrob. Chemother. 2012, 68, 12–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, K.S.; Dallman, T.J.; Field, N.; Childs, T.; Mitchell, H.; Day, M.; Weill, F.-X.; Lefèvre, S.; Tourdjman, M.; Hughes, G.; et al. Horizontal antimicrobial resistance transfer drives epidemics of multiple Shigella species. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. The antibiotic resistome: The nexus of chemical and genetic diversity. Nat. Rev. Genet. 2007, 5, 175–186. [Google Scholar] [CrossRef]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Genet. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Motta, S.; Aldana, M. Adaptive resistance to antibiotics in bacteria: A systems biology perspective. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Sehar, S.; Manefield, M. The roles of extracellular DNA in the structural integrity of extracellular polymeric substance and bacterial biofilm development. Environ. Microbiol. Rep. 2013, 5, 778–786. [Google Scholar] [CrossRef]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Wu, Y.; Vulić, M.; Keren, I.; Lewis, K. Role of Oxidative Stress in Persister Tolerance. Antimicrob. Agents Chemother. 2012, 56, 4922–4926. [Google Scholar] [CrossRef] [Green Version]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, Targets, and Triggers: An Overview of Toxin-Antitoxin Biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef] [Green Version]
- Dörr, T.; Lewis, K.; Vulić, M. SOS Response Induces Persistence to Fluoroquinolones in Escherichia coli. PLoS Genet. 2009, 5, e1000760. [Google Scholar] [CrossRef] [Green Version]
- Bumann, D.; Fanous, J.; Li, J.; Goormaghtigh, F. Antibiotic chemotherapy against heterogeneous pathogen populations in complex host tissues. F1000Research 2019, 8, 1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Miyoshi-Akiyama, T.; Tada, T.; Ohmagari, N.; Hung, N.V.; Tharavichitkul, P.; Pokhrel, B.M.; Gniadkowski, M.; Shimojima, M.; Kirikae, T. Emergence and Spread of Epidemic Multidrug-Resistant Pseudomonas aeruginosa. Genome Biol. Evol. 2017, 9, 3238–3245. [Google Scholar] [CrossRef] [PubMed]
- Shah, C.; Baral, R.; Bartaula, B.; Shrestha, L.B. Virulence factors of uropathogenic Escherichia coli (UPEC) and correlation with antimicrobial resistance. BMC Microbiol. 2019, 19, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhen, X.; Lundborg, C.S.; Sun, X.; Hu, X.; Dong, H. Economic burden of antibiotic resistance in ESKAPE organisms: A systematic review. Antimicrob. Resist. Infect. Control. 2019, 8, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Nayak, R.P. Dry antibiotic pipeline: Regulatory bottlenecks and regulatory reforms. J. Pharmacol. Pharmacother. 2014, 5, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.F.; Drown, B.S.; Riley, A.P.; Garcia, A.; Shirai, T.; Svec, R.L.; Hergenrother, P.J. Predictive compound accumulation rules yield a broad-spectrum antibiotic. Nat. Cell Biol. 2017, 545, 299–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappuoli, R.; Pizza, M.; Del Giudice, G.; De Gregorio, E. Vaccines, new opportunities for a new society. Proc. Natl. Acad. Sci. USA 2014, 111, 12288–12293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreano, E.; Seubert, A.; Rappuoli, R. Human monoclonal antibodies for discovery, therapy, and vaccine acceleration. Curr. Opin. Immunol. 2019, 59, 130–134. [Google Scholar] [CrossRef]
- Balhuizen, M.D.; Veldhuizen, E.J.A.; Haagsman, H.P. Outer Membrane Vesicle Induction and Isolation for Vaccine Development. Front. Microbiol. 2021, 12, 629090. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Liu, Q. Engineered Bacterial Outer Membrane Vesicles as Multifunctional Delivery Platforms. Front. Mater. 2020, 7, 7. [Google Scholar] [CrossRef]
- Li, M.; Zhou, H.; Yang, C.; Wu, Y.; Zhou, X.; Liu, H.; Wang, Y. Bacterial outer membrane vesicles as a platform for biomedical applications: An update. J. Control. Release 2020, 323, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; MacLennan, C.A. Outer membrane vesicle vaccines. Semin. Immunol. 2020, 50, 101433. [Google Scholar] [CrossRef]
- Bartolini, E.; Ianni, E.; Frigimelica, E.; Petracca, R.; Galli, G.; Scorza, F.B.; Norais, N.; Laera, D.; Giusti, F.; Pierleoni, A.; et al. Recombinant outer membrane vesicles carrying Chlamydia muridarum HtrA induce antibodies that neutralize chlamydial infection in vitro. J. Extracell. Vesicles 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Grandi, A.; Tomasi, M.; Zanella, I.; Ganfini, L.; Caproni, E.; Fantappiè, L.; Irene, C.; Frattini, L.; Isaac, S.J.; König, E.; et al. Synergistic Protective Activity of Tumor-Specific Epitopes Engineered in Bacterial Outer Membrane Vesicles. Front. Oncol. 2017, 7, 253. [Google Scholar] [CrossRef] [Green Version]
- Watkins, H.C.; Rappazzo, C.G.; Higgins, J.S.; Sun, X.; Brock, N.; Chau, A.; Misra, A.; Cannizzo, J.P.; King, M.R.; Maines, T.R.; et al. Safe Recombinant Outer Membrane Vesicles that Display M2e Elicit Heterologous Influenza Protection. Mol. Ther. 2017, 25, 989–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Pol, L.; Stork, M.; Van Der Ley, P. Outer membrane vesicles as platform vaccine technology. Biotechnol. J. 2015, 10, 1689–1706. [Google Scholar] [CrossRef]
- Gujrati, V.B.; Jon, S. Bioengineered bacterial outer membrane vesicles: What is their potential in cancer therapy? Nanomedicine 2014, 9, 933–935. [Google Scholar] [CrossRef]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef] [Green Version]
- Valentini, M.; Gonzalez, D.; Mavridou, D.A.; Filloux, A. Lifestyle transitions and adaptive pathogenesis of Pseudomonas aeruginosa. Curr. Opin. Microbiol. 2018, 41, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Botelho, J.; Grosso, F.; Peixe, L. Antibiotic resistance in Pseudomonas aeruginosa—Mechanisms, epidemiology and evolution. Drug Resist. Updat. 2019, 44, 100640. [Google Scholar] [CrossRef] [PubMed]
- Gellatly, S.L.; Hancock, R.E.W. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winstanley, C.; O’Brien, S.; Brockhurst, M.A. Pseudomonas aeruginosa Evolutionary Adaptation and Diversification in Cystic Fibrosis Chronic Lung Infections. Trends Microbiol. 2016, 24, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Kuchma, S.L.; O’Toole, G.A. Keeping Their Options Open: Acute versus Persistent Infections. J. Bacteriol. 2006, 188, 1211–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, A.; Mulet, X.; López-Causapé, C.; Juan, C. The increasing threat of Pseudomonas aeruginosa high-risk clones. Drug Resist. Updat. 2015, 21-22, 41–59. [Google Scholar] [CrossRef]
- Maciá, M.D.; Blanquer, D.; Togores, B.; Sauleda, J.; Pérez, J.L.; Oliver, A. Hypermutation Is a Key Factor in Development of Multiple-Antimicrobial Resistance in Pseudomonas aeruginosa Strains Causing Chronic Lung Infections. Antimicrob. Agents Chemother. 2005, 49, 3382–3386. [Google Scholar] [CrossRef] [Green Version]
- Peña, C.; Cabot, G.; Gómez-Zorrilla, S.; Zamorano, L.; Ocampo-Sosa, A.; Murillas, J.; Almirante, B.; Pomar, V.; Aguilar, M.; Granados, A.; et al. Influence of Virulence Genotype and Resistance Profile in the Mortality of Pseudomonas aeruginosa Bloodstream Infections. Clin. Infect. Dis. 2015, 60, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Nesta, B.; Pizza, M. Vaccines against Escherichia coli. In Escherichia coli, a Versatile Pathogen; Frankel, G., Ron, E.Z., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 213–242. [Google Scholar]
- Mazzariol, A.; Bazaj, A.; Cornaglia, G. Multi-drug-resistant Gram-negative bacteria causing urinary tract infections: A review. J. Chemother. 2017, 29, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Foxman, B.; Brown, P. Epidemiology of urinary tract infections. Infect. Dis. Clin. North Am. 2003, 17, 227–241. [Google Scholar] [CrossRef]
- A Russo, T. Medical and economic impact of extraintestinal infections due to Escherichia coli: Focus on an increasingly important endemic problem. Microbes Infect. 2003, 5, 449–456. [Google Scholar] [CrossRef]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global Extraintestinal Pathogenic Escherichia coli (ExPEC) Lineages. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti-infective Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Stamm, W.E.; Norrby, S.R. Urinary Tract Infections: Disease Panorama and Challenges. J. Infect. Dis. 2001, 183, S1–S4. [Google Scholar] [CrossRef]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef]
- Mann, R.; Mediati, D.G.; Duggin, I.G.; Harry, E.J.; Bottomley, A.L. Metabolic Adaptations of Uropathogenic E. coli in the Urinary Tract. Front. Cell. Infect. Microbiol. 2017, 7, 241. [Google Scholar] [CrossRef] [PubMed]
- Chenoweth, C.E.; Gould, C.V.; Saint, S. Diagnosis, Management, and Prevention of Catheter-Associated Urinary Tract Infections. Infect. Dis. Clin. North Am. 2014, 28, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Lo, E.; Nicolle, L.E.; Coffin, S.E.; Gould, C.; Maragakis, L.L.; Meddings, J.; Pegues, D.A.; Pettis, A.M.; Saint, S.; Yokoe, D.S. Strategies to prevent catheter-associated urinary tract infections in acute care hospitals: 2014 update. Infect. Control Hosp. Epidemiol. 2014, 35, S32–S47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fihn, S.D. Acute Uncomplicated Urinary Tract Infection in Women. N. Engl. J. Med. 2003, 349, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Duell, B.L.; Carey, A.J.; Dando, S.J.; Schembri, M.A.; Ulett, G.C. Human Bladder Uroepithelial Cells Synergize with Monocytes to Promote IL-10 Synthesis and Other Cytokine Responses to Uropathogenic Escherichia coli. PLoS ONE 2013, 8, e78013. [Google Scholar] [CrossRef] [Green Version]
- Magruder, M.; Sholi, A.N.; Gong, C.; Zhang, L.; Edusei, E.; Huang, J.; Albakry, S.; Satlin, M.J.; Westblade, L.F.; Crawford, C.; et al. Gut uropathogen abundance is a risk factor for development of bacteriuria and urinary tract infection. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Bartoletti, R.; Cai, T.; Wagenlehner, F.M.; Naber, K.; Johansen, T.E.B. Treatment of Urinary Tract Infections and Antibiotic Stewardship. Eur. Urol. Suppl. 2016, 15, 81–87. [Google Scholar] [CrossRef]
- Doran, K.S.; Fulde, M.; Gratz, N.; Kim, B.J.; Nau, R.; Prasadarao, N.V.; Schubert-Unkmeir, A.; Tuomanen, E.I.; Valentin-Weigand, P. Host–pathogen interactions in bacterial meningitis. Acta Neuropathol. 2016, 131, 185–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zlatkov, N.; Uhlin, B.E. Absence of Global Stress Regulation in Escherichia coli Promotes Pathoadaptation and Novel c-di-GMP-dependent Metabolic Capability. Sci. Rep. 2019, 9, 2600. [Google Scholar] [CrossRef] [Green Version]
- Wijetunge, D.S.S.; Gongati, S.; Debroy, C.; Kim, K.S.; Couraud, P.-O.; A Romero, I.; Weksler, B.B.; Kariyawasam, S. Characterizing the pathotype of neonatal meningitis causing Escherichia coli (NMEC). BMC Microbiol. 2015, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Dufendach, K.R.; Wellons, J.C.; Kuba, M.G.; Nickols, H.H.; Gómez-Duarte, O.G.; Wynn, J.L. Lethal neonatal meningoencephalitis caused by multi-drug resistant, highly virulent Escherichia coli. Infect. Dis. 2016, 48, 461–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bronzwaer, S.L.; Cars, O.; Buchholz, U.; Mölstad, S.; Goettsch, W.G.; Veldhuijzen, I.K.; Kool, J.L.; Sprenger, M.J.; Degener, J.E.; System, P.I.T.E.A.R.S. The Relationship between Antimicrobial Use and Antimicrobial Resistance in Europe. Emerg. Infect. Dis. 2002, 8, 278–282. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 2: Management strategies and new agents. Pharm. Ther. 2015, 40, 344–352. [Google Scholar] [PubMed]
- A Goff, D.; Kullar, R.; Goldstein, E.J.C.; Gilchrist, M.; Nathwani, D.; Cheng, A.C.; A Cairns, K.; Escandón-Vargas, K.; Villegas, M.V.; Brink, A.; et al. A global call from five countries to collaborate in antibiotic stewardship: United we succeed, divided we might fail. Lancet Infect. Dis. 2017, 17, e56–e63. [Google Scholar] [CrossRef] [Green Version]
- Fleming-Dutra, K.E.; Hersh, A.L.; Shapiro, D.J.; Bartoces, M.; Enns, E.A.; File, T.M.; Finkelstein, J.A.; Gerber, J.S.; Hyun, D.Y.; Linder, J.A.; et al. Prevalence of Inappropriate Antibiotic Prescriptions Among US Ambulatory Care Visits, 2010–2011. JAMA 2016, 315, 1864–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landers, T.F.; Cohen, B.; Wittum, T.E.; Larson, E.L. A Review of Antibiotic Use in Food Animals: Perspective, Policy, and Potential. Public Heal. Rep. 2012, 127, 4–22. [Google Scholar] [CrossRef] [Green Version]
- Jansen, K.U.; Knirsch, C.; Anderson, A.S. The role of vaccines in preventing bacterial antimicrobial resistance. Nat. Med. 2018, 24, 10–19. [Google Scholar] [CrossRef]
- Tümmler, B. Emerging therapies against infections with Pseudomonas aeruginosa. F1000Research 2019, 8, 1371. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.A.; Read, A.F. Why does drug resistance readily evolve but vaccine resistance does not? Proc. R. Soc. B Boil. Sci. 2017, 284, 20162562. [Google Scholar] [CrossRef]
- Lipsitch, M.; Siber, G.R. How Can Vaccines Contribute to Solving the Antimicrobial Resistance Problem? mBio 2016, 7, e00428-16. [Google Scholar] [CrossRef] [Green Version]
- Kwong, J.C.; Maaten, S.; Upshur, R.E.G.; Patrick, D.M.; Marra, F. The Effect of Universal Influenza Immunization on Antibiotic Prescriptions: An Ecological Study. Clin. Infect. Dis. 2009, 49, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Pecetta, S.; Pizza, M.; Sala, C.; Andreano, E.; Pileri, P.; Troisi, M.; Pantano, E.; Manganaro, N.; Rappuoli, R. Antibodies, epicenter of SARS-CoV-2 immunology. Cell Death Differ. 2021, 28, 821–824. [Google Scholar] [CrossRef] [PubMed]
- McConnell, M.J. Where are we with monoclonal antibodies for multidrug-resistant infections? Drug Discov. Today 2019, 24, 1132–1138. [Google Scholar] [CrossRef]
- Alexander, J.W.; Fisher, M.W.; Macmillan, B.G. Immunological Control of Pseudomonas Infection in Burn Patients: A Clinical Evaluation. Arch. Surg. 1971, 102, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Young, L.S.; Meyer, R.D.; Armstrong, D. Pseudomonas aeruginosa Vaccine in Cancer Patients. Ann. Intern. Med. 1973, 79, 518–527. [Google Scholar] [CrossRef]
- Haghbin, M.; Armstrong, D.; Murphy, M.L. Controlled prospective trial of Pseudomonas aeruginosa vaccine in children with acute leukemia. Cancer 1973, 32, 761–766. [Google Scholar] [CrossRef]
- Pennington, J.E.; Reynolds, H.Y.; Wood, R.E.; Robinson, R.A.; Levine, A.S. Use of a Pseudomonas aeruginosa vaccine in patients with acute leukemia and cystic fibrosis. Am. J. Med. 1975, 58, 629–636. [Google Scholar] [CrossRef]
- Priebe, G.P.; Goldberg, J.B. Vaccines for Pseudomonas aeruginosa: A long and winding road. Expert Rev. Vaccines 2014, 13, 507–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatano, K.; Pier, G.B. Complex Serology and Immune Response of Mice to Variant High-Molecular-Weight O Polysaccharides Isolated from Pseudomonas aeruginosa Serogroup O2 Strains. Infect. Immun. 1998, 66, 3719–3726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansen, H.K.; Gøtzsche, P.C. Vaccines for preventing infection with Pseudomonas aeruginosa in cystic fibrosis. Cochrane Database Syst. Rev. 2013, 6, CD001399. [Google Scholar] [CrossRef] [Green Version]
- Cryz, S.J.; Lang, A.; Rüdeberg, A.; Wedgwood, J.; Que, J.U.; Fürer, E.; Schaad, U. Immunization of cystic fibrosis patients with a Pseudomonas aeruginosa O-polysaccharide-toxin A conjugate vaccine. Behring Inst. Mitteilungen 1997, 98, 345–349. [Google Scholar]
- Döring, G.; Meisner, C.; Stern, M.; Flagella Vaccine Trial Study Group. A double-blind randomized placebo-controlled phase III study of a Pseudomonas aeruginosa flagella vaccine in cystic fibrosis patients. Proc. Natl. Acad. Sci. USA 2007, 104, 11020–11025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Döring, G.; Pier, G.B. Vaccines and immunotherapy against Pseudomonas aeruginosa. Vaccine 2008, 26, 1011–1024. [Google Scholar] [CrossRef]
- Campfield, B.; Chen, K.; Kolls, J.K. Vaccine approaches for multidrug resistant Gram negative infections. Curr. Opin. Immunol. 2014, 28, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Larbig, M.; Mansouri, E.; Freihorst, J.; Tümmler, B.; Köhler, G.; Domdey, H.; Knapp, B.; Hungerer, K.D.; Hundt, E.; Gabelsberger, J.; et al. Safety and immunogenicity of an intranasal Pseudomonas aeruginosa hybrid outer membrane protein F-I vaccine in human volunteers. Vaccine 2001, 19, 2291–2297. [Google Scholar] [CrossRef]
- Rello, J.; Krenn, C.-G.; Locker, G.; Pilger, E.; Madl, C.; Balica, L.; Dugernier, T.; Laterre, P.-F.; Spapen, H.; Depuydt, P.; et al. A randomized placebo-controlled phase II study of a Pseudomonas vaccine in ventilated ICU patients. Crit. Care 2017, 21, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westritschnig, K.; Hochreiter, R.; Wallner, G.; Firbas, C.; Schwameis, M.; Jilma, B. A randomized, placebo-controlled phase I study assessing the safety and immunogenicity of a Pseudomonas aeruginosa hybrid outer membrane protein OprF/I vaccine (IC43) in healthy volunteers. Hum. Vaccines Immunother. 2013, 10, 170–183. [Google Scholar] [CrossRef] [Green Version]
- Burrows, L.L. The Therapeutic Pipeline for Pseudomonas aeruginosa Infections. ACS Infect. Dis. 2018, 4, 1041–1047. [Google Scholar] [CrossRef]
- Merakou, C.; Schaefers, M.M.; Priebe, G.P. Progress Toward the Elusive Pseudomonas aeruginosa Vaccine. Surg. Infect. 2018, 19, 757–768. [Google Scholar] [CrossRef]
- Ali, S.O.; Yu, X.Q.; Robbie, G.J.; Wu, Y.; Shoemaker, K.; Yu, L.; DiGiandomenico, A.; Keller, A.E.; Anude, C.; Hernandez-Illas, M.; et al. Phase 1 study of MEDI3902, an investigational anti–Pseudomonas aeruginosa PcrV and Psl bispecific human monoclonal antibody, in healthy adults. Clin. Microbiol. Infect. 2019, 25, 629.e1–629.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chastre, J.; François, B.; Bourgeois, M.; Komnos, A.; Ferrer, R.; Rahav, G.; De Schryver, N.; Lepape, A.; Koksal, I.; Luyt, C.-E.; et al. 635. Efficacy, Pharmacokinetics (PK), and Safety Profile of MEDI3902, an Anti-Pseudomonas aeruginosa Bispecific Human Monoclonal Antibody in Mechanically Ventilated Intensive Care Unit Patients; Results of the Phase 2 EVADE Study Conducted by the Public-Private COMBACTE-MAGNET Consortium in the Innovative Medicines Initiative (IMI) Program. Open Forum Infect. Dis. 2020, 7, S377–S378. [Google Scholar] [CrossRef]
- Cryz, S.J.; Cross, A.S.; Sadoff, J.C.; Wegmann, A.; Que, J.U.; Fürer, E. Safety and Immunogenicity of Escherichia coli 018 O-Specific Polysaccharide (O-PS)-Toxin A and O-PS-Cholera Toxin Conjugate Vaccines in Humans. J. Infect. Dis. 1991, 163, 1040–1045. [Google Scholar] [CrossRef]
- Poolman, J.T.; Wacker, M. Extraintestinal Pathogenic Escherichia coli, a Common Human Pathogen: Challenges for Vaccine Development and Progress in the Field. J. Infect. Dis. 2016, 213, 6–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loubet, P.; Ranfaing, J.; Dinh, A.; Dunyach-Remy, C.; Bernard, L.; Bruyère, F.; Lavigne, J.-P.; Sotto, A. Alternative Therapeutic Options to Antibiotics for the Treatment of Urinary Tract Infections. Front. Microbiol. 2020, 11, 1509. [Google Scholar] [CrossRef] [PubMed]
- Tammen, H. Immunobiotherapy with Uro-Vaxom in Recurrent Urinary Tract Infection. BJU Int. 1990, 65, 6–9. [Google Scholar] [CrossRef]
- Bauer, H.W.; Alloussi, S.; Egger, G.; Blümlein, H.-M.; Cozma, G.; Schulman, C.C. A Long-Term, Multicenter, Double-Blind Study of an Escherichia coli Extract (OM-89) in Female Patients with Recurrent Urinary Tract Infections. Eur. Urol. 2005, 47, 542–548. [Google Scholar] [CrossRef]
- Magasi, P.; Pánovics, J.; Illés, A.; Nagy, M. Uro-Vaxom^® and the Management of Recurrent Urinary Tract Infection in Adults: A Randomized Multicenter Double-Blind Trial. Eur. Urol. 1994, 26, 137–140. [Google Scholar] [CrossRef]
- Schulman, C.; Corbusier, A.; Michiels, H.; Taenzer, H. Oral Immunotherapy of Recurrent Urinary Tract Infections: A Double-Blind Placebo-Controlled Multicenter Study. J. Urol. 1993, 150, 917–921. [Google Scholar] [CrossRef]
- Brodie, A.; El-Taji, O.; Jour, I.; Foley, C.; Hanbury, D. A Retrospective Study of Immunotherapy Treatment with Uro-Vaxom (OM-89®) for Prophylaxis of Recurrent Urinary Tract Infections. Curr. Urol. 2020, 14, 130–134. [Google Scholar] [CrossRef]
- Wagenlehner, F.M.; Ballarini, S.; Pilatz, A.; Weidner, W.; Lehr, L.; Naber, K.G. A Randomized, Double-Blind, Parallel-Group, Multicenter Clinical Study of Escherichia coli-Lyophilized Lysate for the Prophylaxis of Recurrent Uncomplicated Urinary Tract Infections. Urol. Int. 2015, 95, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.J.; Uehling, D.T. Vaccine development for the prevention of urinary tract infections. Curr. Infect. Dis. Rep. 2002, 4, 509–513. [Google Scholar] [CrossRef]
- Yang, B.; Foley, S. Urinary tract infection vaccines—The ‘burning’issue. BJU Int. 2019, 123, 743–744. [Google Scholar] [CrossRef] [Green Version]
- Aziminia, N.; Hadjipavlou, M.; Philippou, Y.; Pandian, S.S.; Malde, S.; Hammadeh, M.Y. Vaccines for the prevention of recurrent urinary tract infections: A systematic review. BJU Int. 2019, 123, 753–768. [Google Scholar] [CrossRef] [Green Version]
- Sevilla, C.R.; Lanza, E.G.; Manzanera, J.L.; Martín, J.A.R.; Sanz, M.Á.B. Active immunoprophyilaxis with uromune® decreases the recurrence of urinary tract infections at three and six months after treatment without relevant secondary effects. BMC Infect. Dis. 2019, 19, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Nickel, J.C. Could sublingual vaccination be a viable option for the prevention of recurrent urinary tract infection in Canada? A systematic review of the current literature and plans for the future. Can. Urol. Assoc. J. 2020, 14, 281–287. [Google Scholar] [CrossRef]
- Huttner, A.; Gambillara, V. The development and early clinical testing of the ExPEC4V conjugate vaccine against uropathogenic Escherichia coli. Clin. Microbiol. Infect. 2018, 24, 1046–1050. [Google Scholar] [CrossRef]
- Huttner, A.; Hatz, C.; Dobbelsteen, G.V.D.; Abbanat, D.; Hornacek, A.; Frölich, R.; Dreyer, A.M.; Martin, P.; Davies, T.; Fae, K.; et al. Safety, immunogenicity, and preliminary clinical efficacy of a vaccine against extraintestinal pathogenic Escherichia coli in women with a history of recurrent urinary tract infection: A randomised, single-blind, placebo-controlled phase 1b trial. Lancet Infect. Dis. 2017, 17, 528–537. [Google Scholar] [CrossRef]
- Inoue, M.; Ogawa, T.; Tamura, H.; Hagiwara, Y.; Saito, Y.; Abbanat, D.; Dobbelsteen, G.V.D.; Hermans, P.; Thoelen, S.; Poolman, J.; et al. Safety, tolerability and immunogenicity of the ExPEC4V (JNJ-63871860) vaccine for prevention of invasive extraintestinal pathogenic Escherichia coli disease: A phase 1, randomized, double-blind, placebo-controlled study in healthy Japanese participants. Hum. Vaccines Immunother. 2018, 14, 2150–2157. [Google Scholar] [CrossRef] [Green Version]
- Frenck, R.W.; Ervin, J.; Chu, L.; Abbanat, D.; Spiessens, B.; Go, O.; Haazen, W.; Dobbelsteen, G.V.D.; Poolman, J.; Thoelen, S.; et al. Safety and immunogenicity of a vaccine for extra-intestinal pathogenic Escherichia coli (ESTELLA): A phase 2 randomised controlled trial. Lancet Infect. Dis. 2019, 19, 631–640. [Google Scholar] [CrossRef]
- Eldridge, G.R.; Hughey, H.; Rosenberger, L.; Martin, S.M.; Shapiro, A.M.; D’Antonio, E.; Krejci, K.G.; Shore, N.; Peterson, J.; Lukes, A.S.; et al. Safety and immunogenicity of an adjuvanted Escherichia coli adhesin vaccine in healthy women with and without histories of recurrent urinary tract infections: Results from a first-in-human phase 1 study. Hum. Vaccines Immunother. 2021, 17, 1262–1270. [Google Scholar] [CrossRef] [PubMed]
- Motley, M.P.; Fries, B.C. A New Take on an Old Remedy: Generating Antibodies against Multidrug-Resistant Gram-Negative Bacteria in a Postantibiotic World. mSphere 2017, 2, e00397-17. [Google Scholar] [CrossRef] [Green Version]
- Guachalla, L.M.; Hartl, K.; Varga, C.; Stulik, L.; Mirkina, I.; Malafa, S.; Nagy, E.; Nagy, G.; Szijártó, V. Multiple Modes of Action of a Monoclonal Antibody against Multidrug-Resistant Escherichia coli Sequence Type 131-H30. Antimicrob. Agents Chemother. 2017, 61, e01428-17. [Google Scholar] [CrossRef] [Green Version]
- Guachalla, L.M.; Ramoni, K.; Varga, C.; Mutti, M.; Ghazawi, A.; Pál, T.; Nagy, E.; Sonnevend, Á.; Nagy, G.; Szijártó, V. Retained Activity of an O25b-Specific Monoclonal Antibody against an Mcr-1-Producing Escherichia coli Sequence Type 131 Strain. Antimicrob. Agents Chemother. 2018, 62, e00046-18. [Google Scholar] [CrossRef] [Green Version]
- Henriques, P.; Iacono, L.D.; Gimeno, A.; Biolchi, A.; Romano, M.R.; Arda, A.; Bernardes, G.J.L.; Jimenez-Barbero, J.; Berti, F.; Rappuoli, R.; et al. Structure of a protective epitope reveals the importance of acetylation of Neisseria meningitidisserogroup A capsular polysaccharide. Proc. Natl. Acad. Sci. USA 2020, 117, 29795–29802. [Google Scholar] [CrossRef]
- Kisiela, D.I.; Avagyan, H.; Friend, D.; Jalan, A.; Gupta, S.; Interlandi, G.; Liu, Y.; Tchesnokova, V.; Rodriguez, V.B.; Sumida, J.P.; et al. Inhibition and Reversal of Microbial Attachment by an Antibody with Parasteric Activity against the FimH Adhesin of Uropathogenic E. coli. PLoS Pathog. 2015, 11, e1004857. [Google Scholar] [CrossRef] [Green Version]
- Cywes-Bentley, C.; Skurnik, D.; Zaidi, T.; Roux, D.; DeOliveira, R.B.; Garrett, W.S.; Lu, X.; O’Malley, J.; Kinzel, K.; Rey, A.; et al. Antibody to a conserved antigenic target is protective against diverse prokaryotic and eukaryotic pathogens. Proc. Natl. Acad. Sci. USA 2013, 110, E2209–E2218. [Google Scholar] [CrossRef] [Green Version]
- Skurnik, D.; Cywes-Bentley, C.; Pier, G.B. The exceptionally broad-based potential of active and passive vaccination targeting the conserved microbial surface polysaccharide PNAG. Expert Rev. Vaccines 2016, 15, 1041–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, D.; Pier, G.B.; Skurnik, D. Magic bullets for the 21st century: The reemergence of immunotherapy for multi- and pan-resistant microbes. J. Antimicrob. Chemother. 2012, 67, 2785–2787. [Google Scholar] [CrossRef] [Green Version]
- Soliman, C.; Walduck, A.K.; Yuriev, E.; Richards, J.S.; Cywes-Bentley, C.; Pier, G.B.; Ramsland, P.A. Structural basis for antibody targeting of the broadly expressed microbial polysaccharide poly-N-acetylglucosamine. J. Biol. Chem. 2018, 293, 5079–5089. [Google Scholar] [CrossRef] [Green Version]
- Bjune, G.; Iby, E.H.O.; Nnesby, J.G.O.; Arnesen, O.; Fredriksen, J.; Lindbak, A.-K.; Kleby, H.N.O.; Rosenqvist, E.; Solberg, L.; Closs, O.; et al. Effect of outer membrane vesicle vaccine against group B meningococcal disease in Norway. Lancet 1991, 338, 1093–1096. [Google Scholar] [CrossRef]
- Oster, P.; Lennon, D.; O’Hallahan, J.; Mulholland, K.; Reid, S.; Martin, D. MeNZB?: A safe and highly immunogenic tailor-made vaccine against the New Zealand serogroup B disease epidemic strain. Vaccine 2005, 23, 2191–2196. [Google Scholar] [CrossRef]
- Sierra, G.V.; Campa, H.C.; Varcacel, N.M.; Garcia, I.L.; Izquierdo, P.L.; Sotolongo, P.F.; Casanueva, G.V.; O Rico, C.; Rodriguez, C.R.; Terry, M.H. Vaccine against group B Neisseria meningitidis: Protection trial and mass vaccination results in Cuba. NIPH Ann. 1991, 14, 195–207. [Google Scholar]
- Holst, J.; Martin, D.; Arnold, R.; Huergo, C.C.; Oster, P.; O’Hallahan, J.; Rosenqvist, E. Properties and clinical performance of vaccines containing outer membrane vesicles from Neisseria meningitidis. Vaccine 2009, 27, B3–B12. [Google Scholar] [CrossRef]
- Serruto, D.; Bottomley, M.J.; Ram, S.; Giuliani, M.M.; Rappuoli, R. The new multicomponent vaccine against meningococcal serogroup B, 4CMenB: Immunological, functional and structural characterization of the antigens. Vaccine 2012, 30, B87–B97. [Google Scholar] [CrossRef] [Green Version]
- Bennett-Guerrero, E.; McIntosh, T.J.; Barclay, G.R.; Snyder, D.S.; Gibbs, R.J.; Mythen, M.G.; Poxton, I.R. Preparation and Preclinical Evaluation of a Novel Liposomal Complete-Core Lipopolysaccharide Vaccine. Infect. Immun. 2000, 68, 6202–6208. [Google Scholar] [CrossRef]
- Zariri, A.; Van Der Ley, P. Biosynthetically engineered lipopolysaccharide as vaccine adjuvant. Expert Rev. Vaccines 2015, 14, 861–876. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, F.; Zou, J.; Wu, W.; Jing, H.; Gou, Q.; Li, H.; Gu, J.; Zou, Q.; Zhang, J. Immunization with Pseudomonas aeruginosa outer membrane vesicles stimulates protective immunity in mice. Vaccine 2018, 36, 1047–1054. [Google Scholar] [CrossRef]
- Valguarnera, E.; Feldman, M.F. Glycoengineered Outer Membrane Vesicles as a Platform for Vaccine Development. RNA Interference 2017, 597, 285–310. [Google Scholar] [CrossRef]
- Wurpel, D.J.; Moriel, D.G.; Totsika, M.; Easton, D.M.; Schembri, M.A. Comparative analysis of the uropathogenic Escherichia coli surface proteome by tandem mass-spectrometry of artificially induced outer membrane vesicles. J. Proteom. 2015, 115, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Scorza, F.B.; Doro, F.; Rodríguez-Ortega, M.J.; Stella, M.; Liberatori, S.; Taddei, A.R.; Serino, L.; Moriel, D.G.; Nesta, B.; Fontana, M.R.; et al. Proteomics Characterization of Outer Membrane Vesicles from the Extraintestinal Pathogenic Escherichia coli ΔtolR IHE3034 Mutant. Mol. Cell. Proteom. 2008, 7, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Zanella, I.; König, E.; Tomasi, M.; Gagliardi, A.; Frattini, L.; Fantappiè, L.; Irene, C.; Zerbini, F.; Caproni, E.; Isaac, S.J.; et al. Proteome-minimized outer membrane vesicles from Escherichia coli as a generalized vaccine platform. J. Extracell. Vesicles 2021, 10, e12066. [Google Scholar] [CrossRef]
- Wojdyla, J.A.; Cutts, E.; Kaminska, R.; Papadakos, G.; Hopper, J.T.S.; Stansfeld, P.J.; Staunton, D.; Robinson, C.V.; Kleanthous, C. Structure and Function of the Escherichia coli Tol-Pal Stator Protein TolR. J. Biol. Chem. 2015, 290, 26675–26687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwechheimer, C.; Rodriguez, D.L.; Kuehn, M.J. NlpI-mediated modulation of outer membrane vesicle production through peptidoglycan dynamics in Escherichia coli. Microbiol. 2015, 4, 375–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojima, Y.; Sawabe, T.; Konami, K.; Azuma, M. Construction of hypervesiculation Escherichia coli strains and application for secretory protein production. Biotechnol. Bioeng. 2020, 117, 701–709. [Google Scholar] [CrossRef]
- Rueter, C.; Bielaszewska, M. Secretion and Delivery of Intestinal Pathogenic Escherichia coli Virulence Factors via Outer Membrane Vesicles. Front. Cell. Infect. Microbiol. 2020, 10, 91. [Google Scholar] [CrossRef]
- Qing, G.; Gong, N.; Chen, X.; Chen, J.; Zhang, H.; Wang, Y.; Wang, R.; Zhang, S.; Zhang, Z.; Zhao, X.; et al. Natural and engineered bacterial outer membrane vesicles. Biophys. Rep. 2019, 5, 184–198. [Google Scholar] [CrossRef] [Green Version]
- Klimentová, J.; Stulík, J. Methods of isolation and purification of outer membrane vesicles from gram-negative bacteria. Microbiol. Res. 2015, 170, 1–9. [Google Scholar] [CrossRef]
- Bauman, S.J.; Kuehn, M.J. Purification of outer membrane vesicles from Pseudomonas aeruginosa and their activation of an IL-8 response. Microbes Infect. 2006, 8, 2400–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, A.T. Outer Membrane Vesicles (OMVs) of Gram-negative Bacteria: A Perspective Update. Front. Microbiol. 2017, 8, 1053. [Google Scholar] [CrossRef]
- Wurpel, D.J.; Totsika, M.; Allsopp, L.P.; Webb, R.I.; Moriel, D.G.; Schembri, M.A. Comparative proteomics of uropathogenic Escherichia coli during growth in human urine identify UCA-like (UCL) fimbriae as an adherence factor involved in biofilm formation and binding to uroepithelial cells. J. Proteom. 2016, 131, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Park, A.J.; Murphy, K.; Surette, M.D.; Bandoro, C.; Krieger, J.R.; Taylor, P.; Khursigara, C.M. Tracking the Dynamic Relationship between Cellular Systems and Extracellular Subproteomes in Pseudomonas aeruginosa Biofilms. J. Proteome Res. 2015, 14, 4524–4537. [Google Scholar] [CrossRef]
- Donnarumma, D.; Faleri, A.; Costantino, P.; Rappuoli, R.; Norais, N. The role of structural proteomics in vaccine development: Recent advances and future prospects. Expert Rev. Proteom. 2016, 13, 55–68. [Google Scholar] [CrossRef]
- Lee, E.-Y.; Bang, J.Y.; Park, G.W.; Choi, D.-S.; Kang, J.S.; Kim, H.-J.; Park, K.-S.; Lee, J.-O.; Kim, Y.-K.; Kwon, K.-H.; et al. Global proteomic profiling of native outer membrane vesicles derived from Escherichia coli. Proteomics 2007, 7, 3143–3153. [Google Scholar] [CrossRef]
- Gasperini, G.; Biagini, M.; Arato, V.; Gianfaldoni, C.; Vadi, A.; Norais, N.; Bensi, G.; Delany, I.; Pizza, M.; Aricò, B.; et al. Outer Membrane Vesicles (OMV)-based and Proteomics-driven Antigen Selection Identifies Novel Factors Contributing to Bordetella pertussis Adhesion to Epithelial Cells. Mol. Cell. Proteom. 2018, 17, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P.; Poirel, L. Epidemiology and Diagnostics of Carbapenem Resistance in Gram-negative Bacteria. Clin. Infect. Dis. 2019, 69, S521–S528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akram, W.M.; Menezes, G.A.; Abbas, N.; Ahmad, W.; Ahmed, A.M. Treatment of Multi-Drug Resistant Gram-Negative Bacterial Pathogenic Infections. J. Pure Appl. Microbiol. 2020, 14, 1639–1647. [Google Scholar] [CrossRef]
- Silhavy, T.J.; Kahne, D.; Walker, S. The Bacterial Cell Envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonelli, G.; Cappelli, L.; Cinelli, P.; Cuffaro, R.; Manca, B.; Nicchi, S.; Tondi, S.; Vezzani, G.; Viviani, V.; Delany, I.; et al. Strategies to Tackle Antimicrobial Resistance: The Example of Escherichia coli and Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 4943. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094943
Antonelli G, Cappelli L, Cinelli P, Cuffaro R, Manca B, Nicchi S, Tondi S, Vezzani G, Viviani V, Delany I, et al. Strategies to Tackle Antimicrobial Resistance: The Example of Escherichia coli and Pseudomonas aeruginosa. International Journal of Molecular Sciences. 2021; 22(9):4943. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094943
Chicago/Turabian StyleAntonelli, Giada, Luigia Cappelli, Paolo Cinelli, Rossella Cuffaro, Benedetta Manca, Sonia Nicchi, Serena Tondi, Giacomo Vezzani, Viola Viviani, Isabel Delany, and et al. 2021. "Strategies to Tackle Antimicrobial Resistance: The Example of Escherichia coli and Pseudomonas aeruginosa" International Journal of Molecular Sciences 22, no. 9: 4943. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094943