
BDNF Overexpression in the Ventral Hippocampus Promotes Antidepressant- and Anxiolytic-Like Activity in Serotonin Transporter Knockout Rats

 , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
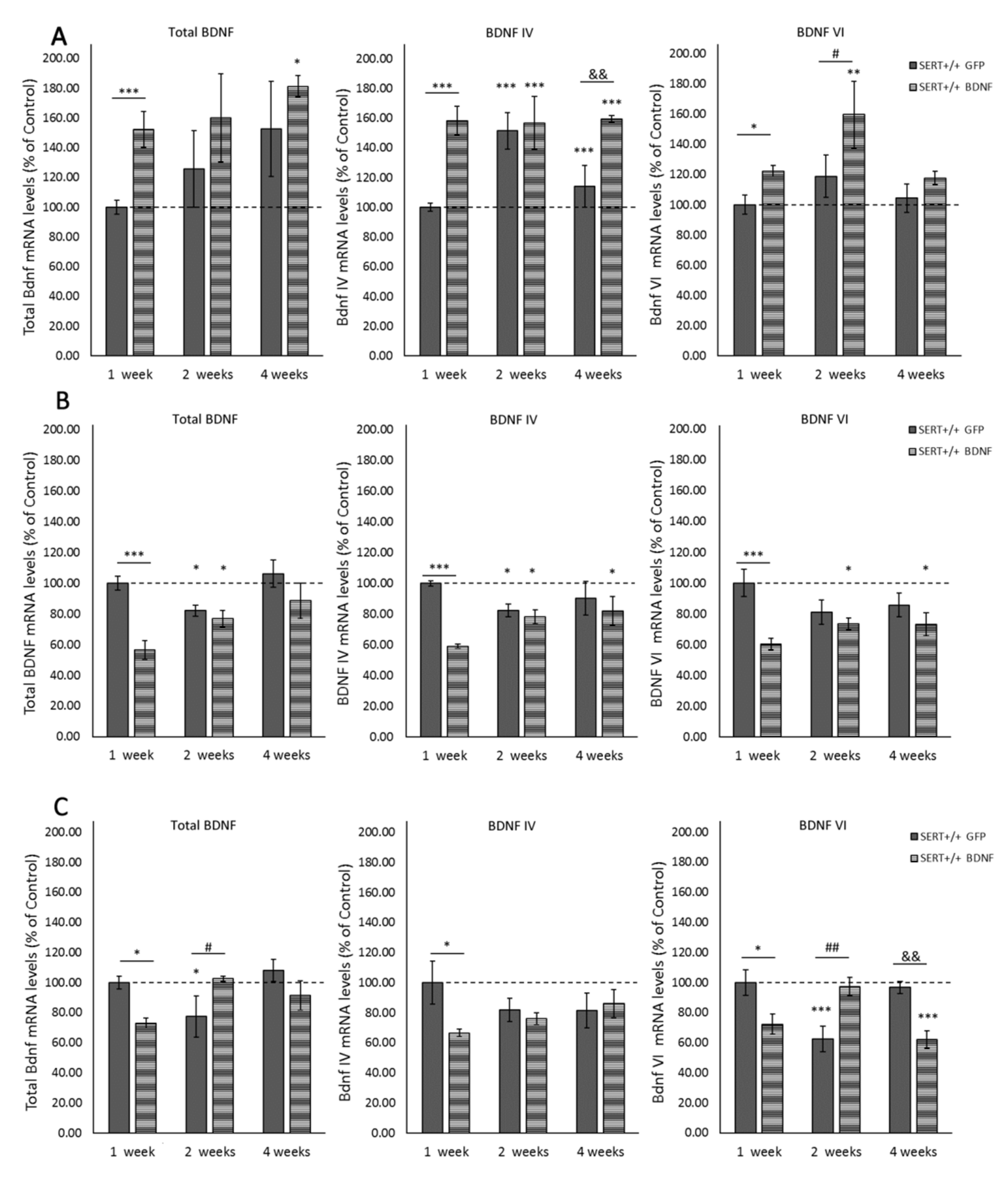
2.1. Lentivirus Transfection Leads to BDNF Overexpression in Ventral Hippocampus of SERT+/+ Rats Subsection
2.2. Local BDNF IV Lentivirus Infusion Leads to Overall BDNF Overexpression in the Ventral Hippocampus
2.3. BDNF Overexpression in the vHIP Leads to BDNF mRNA Downregulation in the Infralimbic Cortex
2.4. BDNF Overexpression in the vHIP Alters BDNF Expression in the Prelimbic Cortex
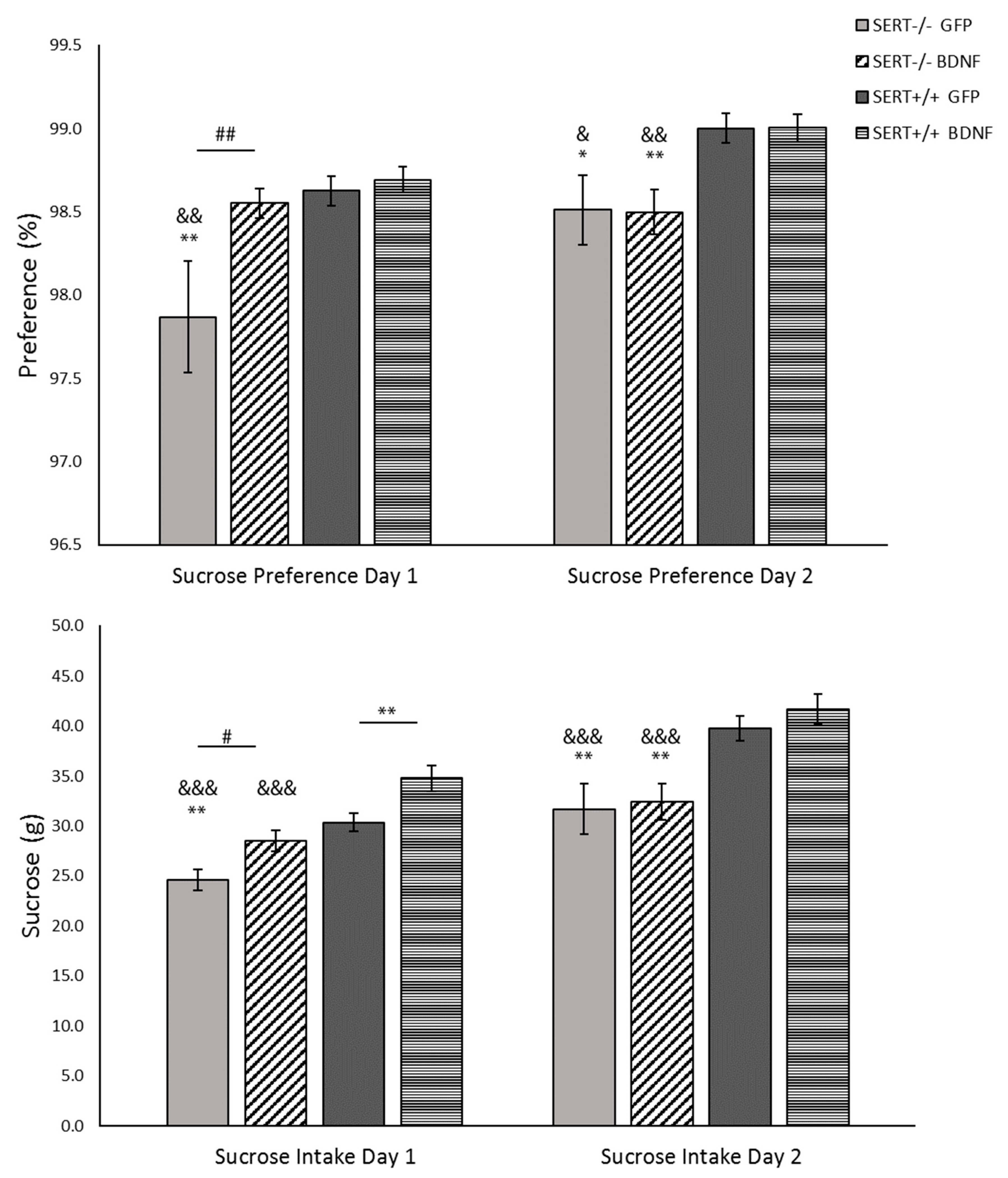
2.5. Sucrose Consumption Test
2.5.1. Sucrose Preference: BDNF Upregulation Meliorates Anhedonia-Like Behavior in SERT−/− Rats upon First Exposure to the Sucrose Solution
2.5.2. Sucrose Intake: BDNF Overexpression Exclusively Modulates Sucrose Intake on the First Day of the Sucrose Consumption Test
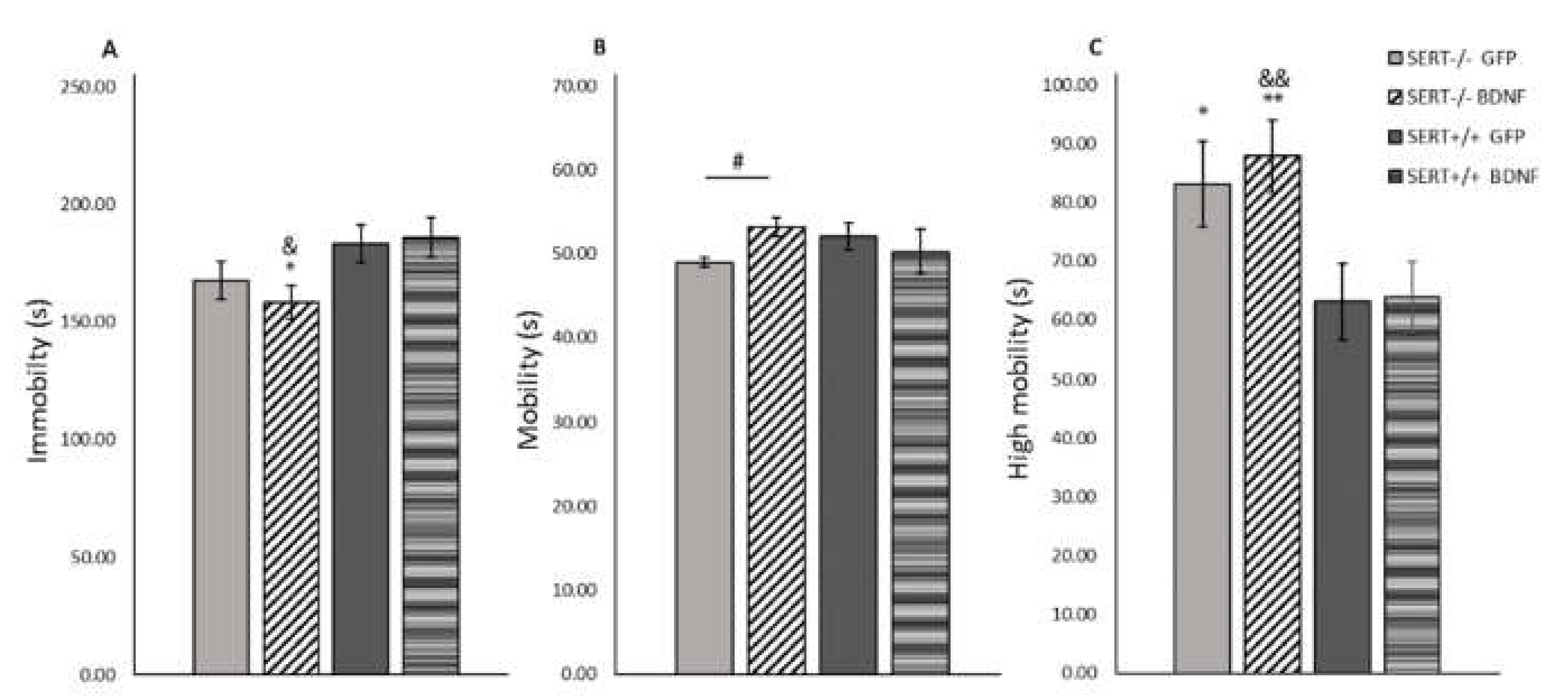
2.6. BDNF Overexpression Decreases Immobility in the Forced Swim Test in SERT−/− Rats
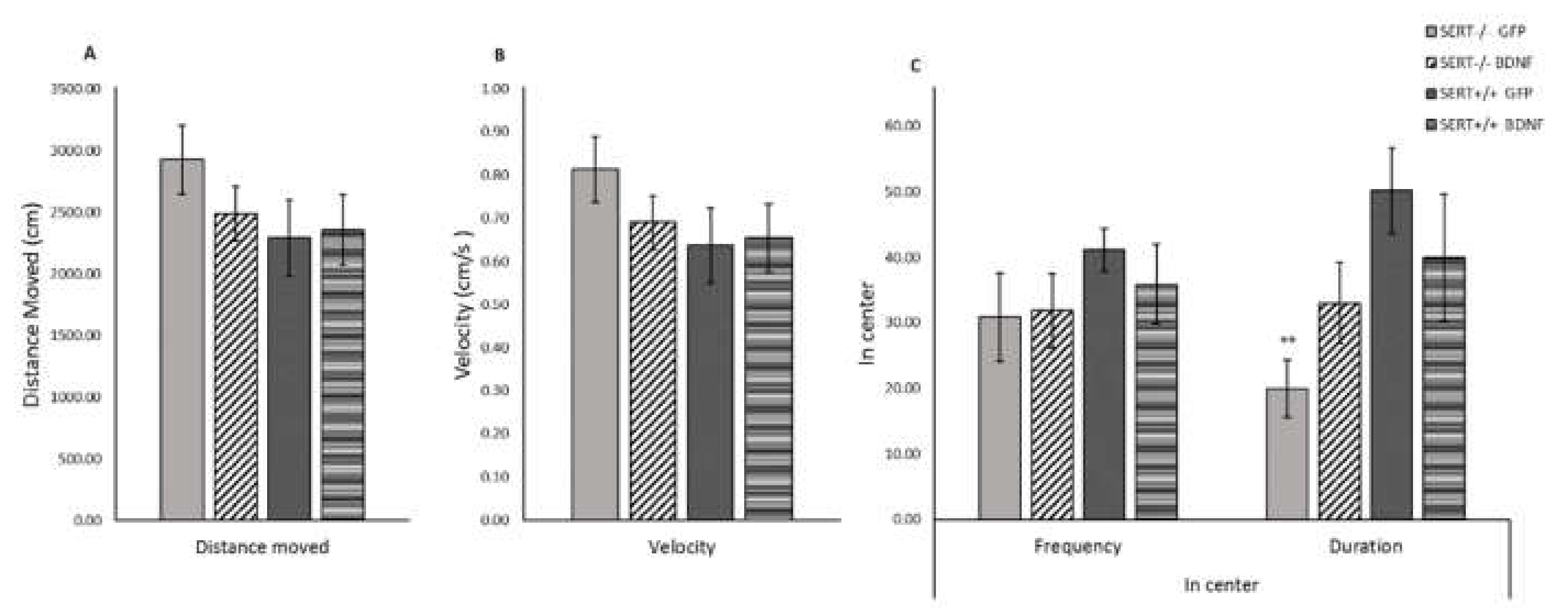
2.7. BDNF Upregulation Decreases Anxiety-Like Behavior in SERT−/− Rats in the Open Field Test
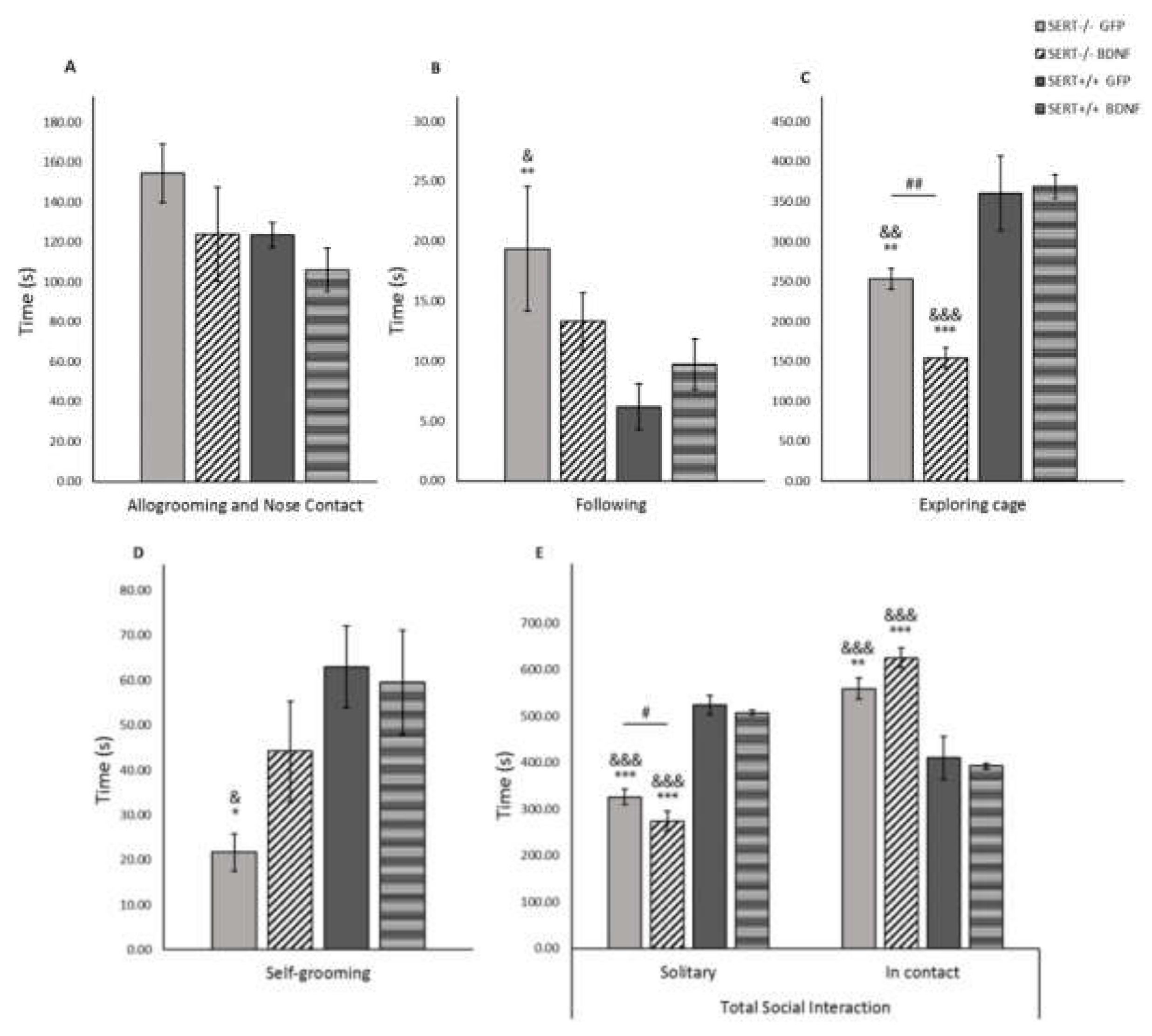
2.8. BDNF in the Social Interaction Test
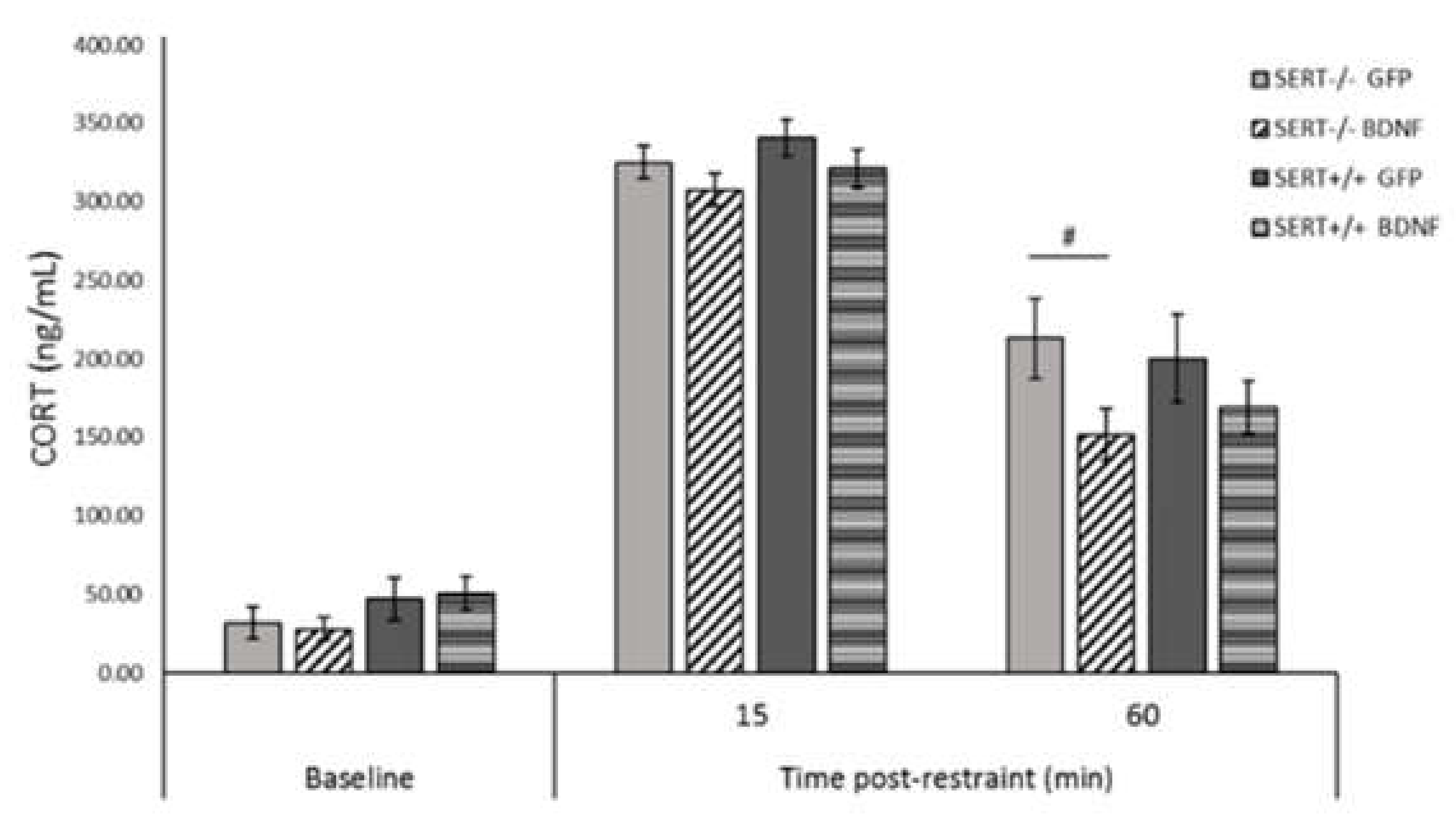
2.9. BDNF Overexpression Reduces CORT Levels in SERT−/− Rats in the HPA-Axis Reactivity Test
3. Discussion
4. Materials and Methods
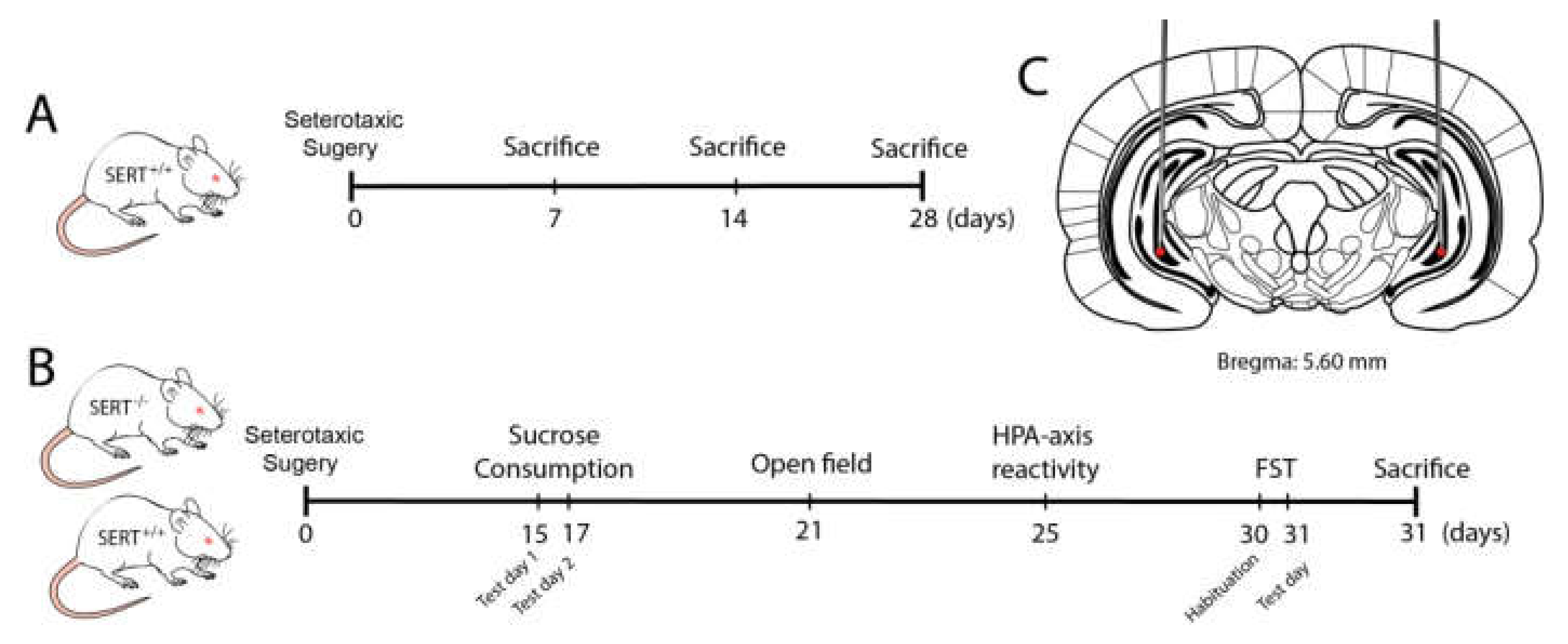
4.1. Animals
4.2. Stereotaxic Surgery
4.3. RNA Preparation and Gene Expression Analysis by Quantitative Real-Time PCR
4.4. Behavioral Tests
4.4.1. Sucrose Consumption Test
4.4.2. HPA-Axis Reactivity Test
4.4.3. Open Field Test
4.4.4. Social Interaction Test
4.4.5. Forced Swim Test
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mateos-Aparicio, P.; Rodríguez-Moreno, A. The Impact of Studying Brain Plasticity. Front. Cell. Neurosci. 2019, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Duman, R.S.; Malberg, J.; Nakagawa, S.; D’Sa, C. Neuronal plasticity and survival in mood disorders. Biol. Psychiatry 2000, 48, 732–739. [Google Scholar] [CrossRef]
- Coppen, A. The Biochemistry of Affective Disorders. Br. J. Psychiatry 1967, 113, 1237–1264. [Google Scholar] [CrossRef]
- Alenina, N.; Klempin, F. The role of serotonin in adult hippocampal neurogenesis. Behav. Brain Res. 2015, 277, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, M.; Gray, J.A.; Roth, B.L. The Expanded Biology of Serotonin. Annu. Rev. Med. 2009, 60, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Deltheil, T.; Guiard, B.; Cerdan, J.; David, D.; Tanaka, K.; Repérant, C.; Guilloux, J.-P.; Coudoré, F.; Hen, R.; Gardier, A. Behavioral and serotonergic consequences of decreasing or increasing hippocampus brain-derived neurotrophic factor protein levels in mice. Neuropharmacology 2008, 55, 1006–1014. [Google Scholar] [CrossRef]
- Duman, R.S.; Malberg, J.; Thome, J. Neural plasticity to stress and antidepressant treatment. Biol. Psychiatry 1999, 46, 1181–1191. [Google Scholar] [CrossRef]
- Edelmann, E.; Leßmann, V.; Brigadski, T. Pre- and postsynaptic twists in BDNF secretion and action in synaptic plasticity. Neuropharmacology 2014, 76, 610–627. [Google Scholar] [CrossRef]
- Mattson, M.P.; Maudsley, S.; Martin, B. BDNF and 5-HT: A dynamic duo in age-related neuronal plasticity and neurodegenerative disorders. Trends Neurosci. 2004, 27, 589–594. [Google Scholar] [CrossRef]
- Park, H.; Poo, M.-M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2012, 14, 7–23. [Google Scholar] [CrossRef]
- Adachi, N. New insight in expression, transport, and secretion of brain-derived neurotrophic factor: Implications in brain-related diseases. World J. Biol. Chem. 2014, 5, 409–428. [Google Scholar] [CrossRef]
- Sen, S.; Duman, R.; Sanacora, G. Serum Brain-Derived Neurotrophic Factor, Depression, and Antidepressant Medications: Meta-Analyses and Implications. Biol. Psychiatry 2008, 64, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Duclot, F.; Kabbaj, M. Epigenetic mechanisms underlying the role of brain-derived neurotrophic factor in depression and response to antidepressants. J. Exp. Biol. 2015, 218, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, R.S.; Monteggia, L.M. A Neurotrophic Model for Stress-Related Mood Disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef]
- Monteggia, L.M.; Barrot, M.; Powell, C.M.; Berton, O.; Galanis, V.; Gemelli, T.; Meuth, S.; Nagy, A.; Greene, R.W.; Nestler, E.J. Essential role of brain-derived neurotrophic factor in adult hippocampal function. Proc. Natl. Acad. Sci. USA 2004, 101, 10827–10832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, D.L.; Fox, M.A.; Timpano, K.R.; Moya, P.R.; Ren-Patterson, R.; Andrews, A.M.; Holmes, A.; Lesch, K.-P.; Wendland, J.R. How the serotonin story is being rewritten by new gene-based discoveries principally related to SLC6A4, the serotonin transporter gene, which functions to influence all cellular serotonin systems. Neuropharmacology 2008, 55, 932–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesch, K.-P.; Bengel, D.; Heils, A.; Sabol, S.Z.; Greenberg, B.D.; Petri, S.; Benjamin, J.; Müller, C.R.; Hamer, D.H.; Murphy, D.L. Association of Anxiety-Related Traits with a Polymorphism in the Serotonin Transporter Gene Regulatory Region. Science 1996, 274, 1527–1531. [Google Scholar] [CrossRef]
- Bleys, D.; Luyten, P.; Soenens, B.; Claes, S. Gene-environment interactions between stress and 5-HTTLPR in depression: A meta-analytic update. J. Affect. Disord. 2018, 226, 339–345. [Google Scholar] [CrossRef]
- Caspi, A.; Sugden, K.; Moffitt, T.E.; Taylor, A.; Craig, I.W.; Harrington, H.; McClay, J.; Mill, J.; Martin, J.; Braithwaite, A.; et al. Influence of Life Stress on Depression: Moderation by a Polymorphism in the 5-HTT Gene. Science 2003, 301, 386–389. [Google Scholar] [CrossRef]
- Fanelli, G.; Serretti, A. The influence of the serotonin transporter gene 5-HTTLPR polymorphism on suicidal behaviors: A meta-analysis. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2019, 88, 375–387. [Google Scholar] [CrossRef]
- Homberg, J.R.; Molteni, R.; Calabrese, F.; Riva, M.A. The serotonin–BDNF duo: Developmental implications for the vulnerability to psychopathology. Neurosci. Biobehav. Rev. 2014, 43, 35–47. [Google Scholar] [CrossRef]
- Kroeze, Y.; Zhou, H.; Homberg, J.R. The genetics of selective serotonin reuptake inhibitors. Pharmacol. Ther. 2012, 136, 375–400. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, F.; Ambrée, O.; Locatelli, C.; Lorenzi, C.; Poletti, S.; Colombo, C.; Arolt, V. The effect of childhood trauma on serum BDNF in bipolar depression is modulated by the serotonin promoter genotype. Neurosci. Lett. 2017, 656, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Bhang, S.; Ahn, J.-H.; Choi, S.-W. Brain-derived neurotrophic factor and serotonin transporter gene-linked promoter region genes alter serum levels of brain-derived neurotrophic factor in humans. J. Affect. Disord. 2011, 128, 299–304. [Google Scholar] [CrossRef]
- Molteni, R.; Cattaneo, A.; Calabrese, F.; Macchi, F.; Olivier, J.D.; Racagni, G.; Ellenbroek, B.A.; Gennarelli, M.; Riva, M.A. Reduced function of the serotonin transporter is associated with decreased expression of BDNF in rodents as well as in humans. Neurobiol. Dis. 2010, 37, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, F.; Van Der Doelen, R.H.A.; Guidotti, G.; Racagni, G.; Kozicz, T.; Homberg, J.R.; Riva, M.A. Exposure to early life stress regulates Bdnf expression in SERT mutant rats in an anatomically selective fashion. J. Neurochem. 2014, 132, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, F.; Guidotti, G.; Middelman, A.; Racagni, G.; Homberg, J.; Riva, M.A. Lack of Serotonin Transporter Alters BDNF Expression in the Rat Brain During Early Postnatal Development. Mol. Neurobiol. 2013, 48, 244–256. [Google Scholar] [CrossRef]
- Kalueff, A.; Olivier, J.; Nonkes, L.; Homberg, J. Conserved role for the serotonin transporter gene in rat and mouse neurobehavioral endophenotypes. Neurosci. Biobehav. Rev. 2010, 34, 373–386. [Google Scholar] [CrossRef]
- Olivier, J.; Van Der Hart, M.; Van Swelm, R.; Dederen, P.; Homberg, J.; Cremers, T.; Deen, P.; Cuppen, E.; Cools, A.; Ellenbroek, B. A study in male and female 5-HT transporter knockout rats: An animal model for anxiety and depression disorders. Neuroscience 2008, 152, 573–584. [Google Scholar] [CrossRef]
- O’Leary, O.F.; Cryan, J.F. A ventral view on antidepressant action: Roles for adult hippocampal neurogenesis along the dorsoventral axis. Trends Pharmacol. Sci. 2014, 35, 675–687. [Google Scholar] [CrossRef]
- Liu, W.; Ge, T.; Leng, Y.; Pan, Z.; Fan, J.; Yang, W.; Cui, R. The Role of Neural Plasticity in Depression: From Hippocampus to Prefrontal Cortex. Neural Plast. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbejjani, M.; Fuhrer, R.; Abrahamowicz, M.; Mazoyer, B.; Crivello, F.; Tzourio, C.; Dufouil, C. Depression, depressive symptoms, and rate of hippocampal atrophy in a longitudinal cohort of older men and women. Psychol. Med. 2015, 45, 1931–1944. [Google Scholar] [CrossRef]
- Elbejjani, M.; Fuhrer, R.; Abrahamowicz, M.; Mazoyer, B.; Crivello, F.; Tzourio, C.; Dufouil, C. Hippocampal Atrophy and Subsequent Depressive Symptoms in Older Men and Women: Results From a 10-Year Prospective Cohort. Am. J. Epidemiol. 2014, 180, 385–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, M.A.O.; Bezerra, L.S.; Carvalho, A.R.M.R.; Brainer-Lima, A.M. Global hippocampal atrophy in major depressive disorder: A meta-analysis of magnetic resonance imaging studies. Trends Psychiatry Psychother. 2018, 40, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Taylor, W.D.; McQuoid, D.R.; Payne, M.E.; Zannas, A.S.; MacFall, J.R.; Steffens, D.C. Hippocampus Atrophy and the Longitudinal Course of Late-life Depression. Am. J. Geriatr. Psychiatry 2014, 22, 1504–1512. [Google Scholar] [CrossRef] [Green Version]
- Castrén, E.; Rantamäki, T. The role of BDNF and its receptors in depression and antidepressant drug action: Reactivation of developmental plasticity. Dev. Neurobiol. 2010, 70, 289–297. [Google Scholar] [CrossRef]
- Castrén, E.; Võikar, V.; Rantamäki, T. Role of Neurotrophic Factors in Depression; Elsevier: Amsterdam, The Netherlands, 2007; Volume 7, pp. 18–21. [Google Scholar]
- Dwivedi, Y.; Rizavi, H.S.; Conley, R.R.; Roberts, R.C.; Tamminga, C.A.; Pandey, G.N. Altered Gene Expression of Brain-Derived Neurotrophic Factor and Receptor Tyrosine Kinase B in Postmortem Brain of Suicide Subjects. Arch. Gen. Psychiatry 2003, 60, 804–815. [Google Scholar] [CrossRef] [Green Version]
- Ray, M.T. Decreased BDNF, trkB-TK+ and GAD67 mRNA expression in the hippocampus of individuals with schizophrenia and mood disorders. J. Psychiatry Neurosci. 2011, 36, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, B.L.; Van Praag, H.; Gage, F.H. Adult brain neurogenesis and psychiatry: A novel theory of depression. Mol. Psychiatry 2000, 5, 262–269. [Google Scholar] [CrossRef]
- Quesseveur, G.; David, D.J.; Gaillard, M.C.; Pla, P.; Wu, M.V.; Nguyen, H.T.; Nicolas, V.; Auregan, G.; David, I.; Dranovsky, A.; et al. BDNF overexpression in mouse hippocampal astrocytes promotes local neurogenesis and elicits anxiolytic-like activities. Transl. Psychiatry 2013, 3, e253. [Google Scholar] [CrossRef]
- Rossi, C.; Angelucci, A.; Costantin, L.; Braschi, C.; Mazzantini, M.; Babbini, F.; Fabbri, M.E.; Tessarollo, L.; Maffei, L.; Berardi, N.; et al. Brain-derived neurotrophic factor (BDNF) is required for the enhancement of hippocampal neurogenesis following environmental enrichment. Eur. J. Neurosci. 2006, 24, 1850–1856. [Google Scholar] [CrossRef]
- Guidotti, G.; Calabrese, F.; Auletta, F.; Olivier, J.; Racagni, G.; Homberg, J.; Riva, M.A. Developmental Influence of the Serotonin Transporter on the Expression of Npas4 and GABAergic Markers: Modulation by Antidepressant Treatment. Neuropsychopharmacology 2011, 37, 746–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aid, T.; Kazantseva, A.; Piirsoo, M.; Palm, K.; Timmusk, T. Mouse and ratBDNF gene structure and expression revisited. J. Neurosci. Res. 2007, 85, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Sakata, K.; Duke, S. Lack of BDNF expression through promoter IV disturbs expression of monoamine genes in the frontal cortex and hippocampus. Neuroscience 2014, 260, 265–275. [Google Scholar] [CrossRef]
- Maynard, K.R.; Hill, J.L.; Calcaterra, N.E.; Palko, M.E.; Kardian, A.; Paredes, D.; Sukumar, M.; Adler, B.D.; Jimenez, D.V.; Schloesser, R.J.; et al. Functional Role of BDNF Production from Unique Promoters in Aggression and Serotonin Signaling. Neuropsychopharmacology 2016, 41, 1943–1955. [Google Scholar] [CrossRef] [PubMed]
- Palaniswamy, R.; Teglund, S.; Lauth, M.; Zaphiropoulos, P.G.; Shimokawa, T. Genetic variations regulate alternative splicing in the 5′ untranslated regions of the mouse glioma-associated oncogene 1, Gli1. BMC Mol. Biol. 2010. [Google Scholar] [CrossRef] [Green Version]
- Spielmann, M.; Mundlos, S. Looking beyond the genes: The role of non-coding variants in human disease. Hum. Mol. Genet. 2016, 25, R157–R165. [Google Scholar] [CrossRef]
- Maynard, K.R.; Hobbs, J.W.; Sukumar, M.; Kardian, A.S.; Jimenez, D.V.; Schloesser, R.J.; Martinowich, K. Bdnf mRNA splice variants differentially impact CA1 and CA3 dendrite complexity and spine morphology in the hippocampus. Brain Struct. Funct. 2017, 222, 3295–3307. [Google Scholar] [CrossRef]
- Verwer, R.W.H.; Meijer, R.J.; Van Uum, H.F.M.; Witter, M.P. Collateral projections from the rat hippocampal formation to the lateral and medial prefrontal cortex. Hippocampus 1997. [Google Scholar] [CrossRef]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nature 2008, 455, 894–902. [Google Scholar] [CrossRef]
- Schulz, P.E.; Arora, G. Depression. Contin. Lifelong Learn. Neurol. 2015, 21, 756–771. [Google Scholar] [CrossRef] [PubMed]
- Porsolt, R.D.; Le Pichon, M.; Jalfre, M. Depression: A new animal model sensitive to antidepressant treatments. Nature 1977, 266, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Menzaghi, F.; Heinrichs, S.C.; Merlo-Pich, E.; Tilders, F.J.; Koob, G.F. Involvement of hypothalamic corticotropin-releasing factor neurons in behavioral responses to novelty in rats. Neurosci. Lett. 1994, 168, 139–142. [Google Scholar] [CrossRef]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Schipper, P.; Nonkes, L.J.; Karel, P.; Kiliaan, A.J.; Homberg, J.R. Serotonin transporter genotype x construction stress interaction in rats. Behav. Brain Res. 2011, 223, 169–175. [Google Scholar] [CrossRef]
- File, S.E.; Hyde, J. Can social interaction be used to measure anxiety? Br. J. Pharmacol. 1978, 62, 19–24. [Google Scholar] [CrossRef] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; SAGE Publications, Inc.: Washington, DC, USA, 2013. [Google Scholar]
- Wenzel, A. Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition (DSM-5). In The SAGE Encyclopedia of Abnormal and Clinical Psychology; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2017. [Google Scholar]
- Baj, G.; Leone, E.; Chao, M.V.; Tongiorgi, E. Spatial segregation of BDNF transcripts enables BDNF to differentially shape distinct dendritic compartments. Proc. Natl. Acad. Sci. USA 2011, 108, 16813–16818. [Google Scholar] [CrossRef] [Green Version]
- Sakata, K.; Jin, L.; Jha, S. Lack of promoter IV-driven BDNF transcription results in depression-like behavior. Genes Brain Behav. 2010, 9, 712–721. [Google Scholar] [CrossRef]
- Matsuda, N.; Lu, H.; Fukata, Y.; Noritake, J.; Gao, H.; Mukherjee, S.; Nemoto, T.; Fukata, M.; Poo, M. Differential Activity-Dependent Secretion of Brain-Derived Neurotrophic Factor from Axon and Dendrite. J. Neurosci. 2009, 29, 14185–14198. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, A.; Topiwala, M.A.; Gordon, J.A. Synchronized Activity between the Ventral Hippocampus and the Medial Prefrontal Cortex during Anxiety. Neuron 2010, 65, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Vertes, R.P. Differential projections of the infralimbic and prelimbic cortex in the rat. Synapse 2003, 51, 32–58. [Google Scholar] [CrossRef]
- Vidal-Gonzalez, I.; Vidal-Gonzalez, B.; Rauch, S.L.; Quirk, G.J. Microstimulation reveals opposing influences of prelimbic and infralimbic cortex on the expression of conditioned fear. Learn. Mem. 2006, 13, 728–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, M.C.; Mielock, A.; Rao, U. The SAGE Encyclopedia of Abnormal and Clinical Psychology; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2017; NV-7. [Google Scholar] [CrossRef] [Green Version]
- Porsolt, R.D.; Anton, G.; Blavet, N.; Jalfre, M. Behavioural despair in rats: A new model sensitive to antidepressant treatments. Eur. J. Pharmacol. 1978, 47, 379–391. [Google Scholar] [CrossRef]
- Lira, A.; Zhou, M.; Castanon, N.; Ansorge, M.S.; Gordon, J.A.; Francis, J.H.; Bradley-Moore, M.; Lira, J.; Underwood, M.D.; Arango, V.; et al. Altered depression-related behaviors and functional changes in the dorsal raphe nucleus of serotonin transporter-deficient mice. Biol. Psychiatry 2003, 54, 960–971. [Google Scholar] [CrossRef]
- Anyan, J.; Amir, S. Too Depressed to Swim or Too Afraid to Stop? A Reinterpretation of the Forced Swim Test as a Measure of Anxiety-Like Behavior. Neuropsychopharmacology 2018, 43, 931–933. [Google Scholar] [CrossRef]
- Karpova, N.N. Role of BDNF epigenetics in activity-dependent neuronal plasticity. Neuropharmacology 2014, 76, 709–718. [Google Scholar] [CrossRef] [Green Version]
- Schipper, P.; Brivio, P.; De Leest, D.; Madder, L.; Asrar, B.; Rebuglio, F.; Verheij, M.M.M.; Kozicz, T.; Riva, M.A.; Calabrese, F.; et al. Impaired Fear Extinction Recall in Serotonin Transporter Knockout Rats Is Transiently Alleviated during Adolescence. Brain Sci. 2019, 9, 118. [Google Scholar] [CrossRef] [Green Version]
- Luoni, A.; Hulsken, S.; Cazzaniga, G.; Racagni, G.; Homberg, J.R.; Riva, M.A. Behavioural and neuroplastic properties of chronic lurasidone treatment in serotonin transporter knockout rats. Int. J. Neuropsychopharmacol. 2013, 16, 1319–1330. [Google Scholar] [CrossRef] [Green Version]
- Lydiard, R.B. The role of GABA in anxiety disorders. J. Clin. Psychiatry 2003, 64, 21–27. [Google Scholar]
- Millan, M.J. The neurobiology and control of anxious states. Prog. Neurobiol. 2003, 70, 83–244. [Google Scholar] [CrossRef]
- Homberg, J.R.; De Boer, S.F.; Raasø, H.S.; Olivier, J.D.A.; Verheul, M.; Ronken, E.; Cools, A.R.; Ellenbroek, B.A.; Schoffelmeer, A.N.M.; Vanderschuren, L.J.M.J.; et al. Adaptations in pre- and postsynaptic 5-HT1A receptor function and cocaine supersensitivity in serotonin transporter knockout rats. Psychopharmacology 2008, 200, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Homberg, J.R.; Olivier, J.; Smits, B.M.G.; Mul, J.D.; Mudde, J.; Verheul, M.; Nieuwenhuizen, O.F.M.; Cools, A.R.; Ronken, E.; Cremers, T.; et al. Characterization of the serotonin transporter knockout rat: A selective change in the functioning of the serotonergic system. Neuroscience 2007, 146, 1662–1676. [Google Scholar] [CrossRef]
- Olivier, J.; Cools, A.; Ellenbroek, B.; Cuppen, E.; Homberg, J. The serotonin transporter knock-out rat: A review. In Experimental Models in Serotonin Transporter Research; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Schipper, P.; Kiliaan, A.J.; Homberg, J.R. A mixed polyunsaturated fatty acid diet normalizes hippocampal neurogenesis and reduces anxiety in serotonin transporter knockout rats. Behav. Pharmacol. 2011, 22, 324–334. [Google Scholar] [CrossRef]
- Jolles, J.; Rompa-Barendregt, J.; Gispen, W. ACTH-induced excessive grooming in the rat: The influence of environmental and motivational factors. Horm. Behav. 1979, 12, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Gispen, W.; Isaacson, R. ACTH-induced excessive grooming in the rat. Pharmacol. Ther. 1981, 12, 209–246. [Google Scholar] [CrossRef] [Green Version]
- Kametani, H. Analysis of Age-related Changes in Stress-induced Grooming in the Rat. Ann. N. Y. Acad. Sci. 1988, 525, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Spruijt, B.M.; Van Hooff, J.A.; Gispen, W.H. Ethology and neurobiology of grooming behavior. Physiol. Rev. 1992, 72, 825–852. [Google Scholar] [CrossRef]
- De Kloet, E.R.; Joëls, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef]
- Chen, X.; Margolis, K.J.; Gershon, M.D.; Schwartz, G.J.; Sze, J.Y. Reduced Serotonin Reuptake Transporter (SERT) Function Causes Insulin Resistance and Hepatic Steatosis Independent of Food Intake. PLoS ONE 2012, 7, e32511. [Google Scholar] [CrossRef] [Green Version]
- Tjurmina, O.A.; Armando, I.; Saavedra, J.M.; Goldstein, D.S.; Murphy, D.L. Exaggerated Adrenomedullary Response to Immobilization in Mice with Targeted Disruption of the Serotonin Transporter Gene. Endocrinology 2002, 143, 4520–4526. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wichems, C.; Heils, A.; Van De Kar, L.D.; Lesch, K.P.; Murphy, D.L. Reduction of 5-hydroxytryptamine (5-HT)(1A)-mediated temperature and neuroendocrine responses and 5-HT(1A) binding sites in 5-HT transporter knockout mice. J. Pharmacol. Exp. Ther. 1999, 291, 999–1007. [Google Scholar]
- Jansen, F.; Heiming, R.S.; Lewejohann, L.; Touma, C.; Palme, R.; Schmitt, A.; Lesch, K.P.; Sachser, N. Modulation of behavioural profile and stress response by 5-HTT genotype and social experience in adulthood. Behav. Brain Res. 2010, 207, 21–29. [Google Scholar] [CrossRef]
- Van Der Doelen, R.H.A.; Deschamps, W.; D’Annibale, C.; Peeters, D.; Wevers, R.A.; Zelena, D.; Homberg, J.R.; Kozicz, T. Early life adversity and serotonin transporter gene variation interact at the level of the adrenal gland to affect the adult hypothalamo-pituitary-adrenal axis. Transl. Psychiatry 2014, 4, e409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belay, H.; Burton, C.L.; Lovic, V.; Meaney, M.J.; Sokolowski, M.; Fleming, A.S. Early adversity and serotonin transporter genotype interact with hippocampal glucocorticoid receptor mRNA expression, corticosterone, and behavior in adult male rats. Behav. Neurosci. 2011, 125, 150–160. [Google Scholar] [CrossRef] [Green Version]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of Depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, C.; Praça, C.; Ferreira, R.; Santos, T.; Ferreira, L.; Bernardino, L. Nanoparticle-mediated brain drug delivery: Overcoming blood-brain barrier to treat neurodegenerative diseases. J. Control. Release 2016, 235, 34–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, B.M.; Mudde, J.B.; van de Belt, J.; Verheul, M.; Olivier, J.; Homberg, J.; Guryev, V.; Cools, A.R.; Ellenbroek, B.A.; Plasterk, R.H.; et al. Generation of gene knockouts and mutant models in the laboratory rat by ENU-driven target-selected mutagenesis. Pharmacogenetics Genom. 2006, 16, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Elsevier Acad. Press: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Manfré, G.; Clemensson, E.K.H.; Kyriakou, E.I.; Clemensson, L.E.; Van Der Harst, J.E.; Homberg, J.R.; Nguyen, H.P. The BACHD Rat Model of Huntington Disease Shows Specific Deficits in a Test Battery of Motor Function. Front. Behav. Neurosci. 2017, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulle, F.; Pawluski, J.L.; Homberg, J.R.; Machiels, B.; Kroeze, Y.; Kumar, N.; Steinbusch, H.W.; Kenis, G.; Hove, D.L.V.D. Developmental fluoxetine exposure increases behavioral despair and alters epigenetic regulation of the hippocampal BDNF gene in adult female offspring. Horm. Behav. 2016, 80, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Hove, D.V.D.; Kenis, G.; Brass, A.; Opstelten, R.; Rutten, B.; Bruschettini, M.; Blanco, C.E.; Lesch, K.P.; Steinbusch, H.; Prickaerts, J. Vulnerability versus resilience to prenatal stress in male and female rats; Implications from gene expression profiles in the hippocampus and frontal cortex. Eur. Neuropsychopharmacol. 2013, 23, 1226–1246. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diniz, D.M.; Calabrese, F.; Brivio, P.; Riva, M.A.; Grandjean, J.; Homberg, J.R. BDNF Overexpression in the Ventral Hippocampus Promotes Antidepressant- and Anxiolytic-Like Activity in Serotonin Transporter Knockout Rats. Int. J. Mol. Sci. 2021, 22, 5040. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22095040
Diniz DM, Calabrese F, Brivio P, Riva MA, Grandjean J, Homberg JR. BDNF Overexpression in the Ventral Hippocampus Promotes Antidepressant- and Anxiolytic-Like Activity in Serotonin Transporter Knockout Rats. International Journal of Molecular Sciences. 2021; 22(9):5040. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22095040
Chicago/Turabian StyleDiniz, Danielle M., Francesca Calabrese, Paola Brivio, Marco A. Riva, Joanes Grandjean, and Judith R. Homberg. 2021. "BDNF Overexpression in the Ventral Hippocampus Promotes Antidepressant- and Anxiolytic-Like Activity in Serotonin Transporter Knockout Rats" International Journal of Molecular Sciences 22, no. 9: 5040. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22095040