
Thalidomide Alleviates Pulmonary Fibrosis Induced by Silica in Mice by Inhibiting ER Stress and the TLR4-NF-κB Pathway

 , and
, and 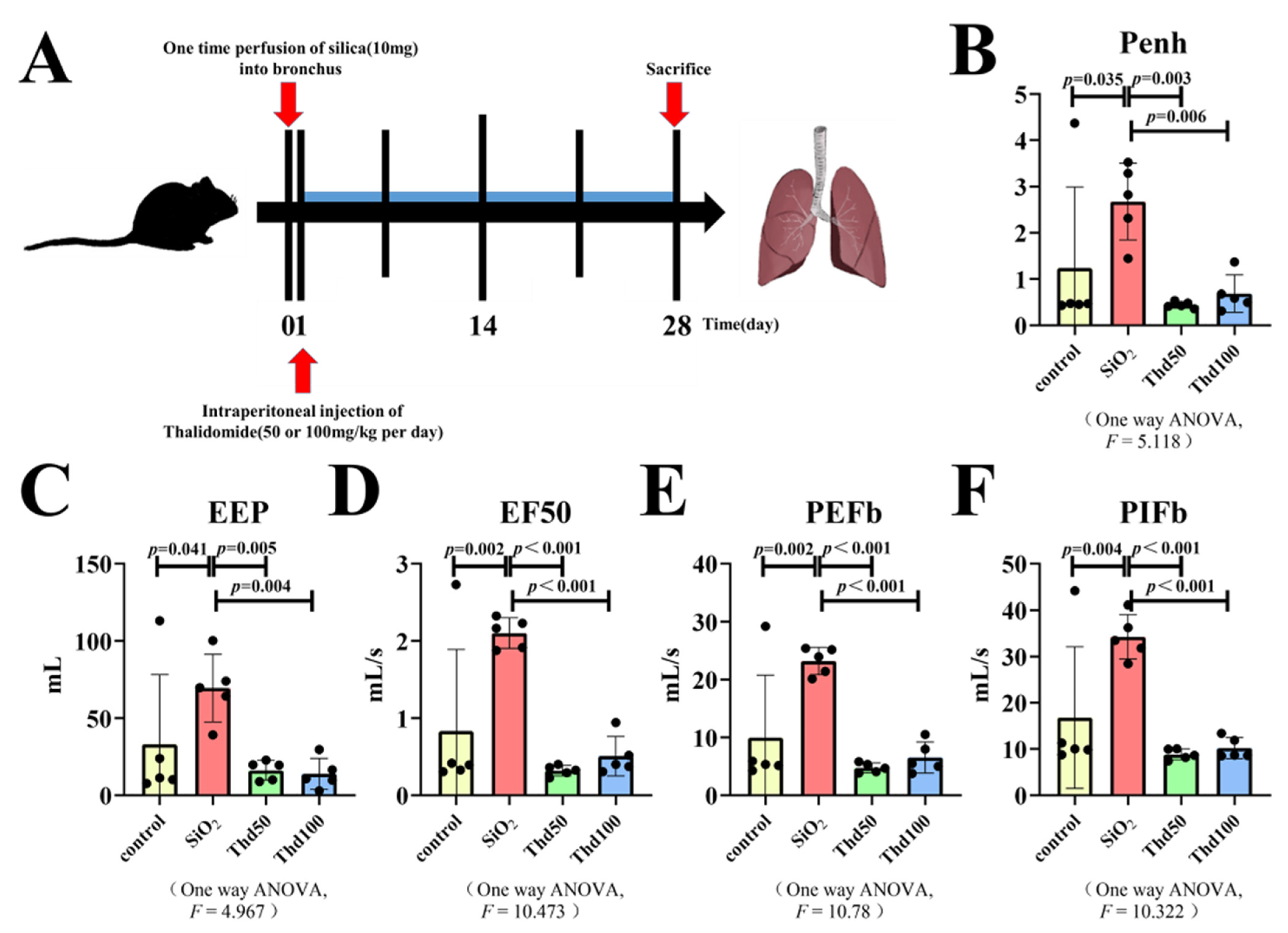{kind=link}
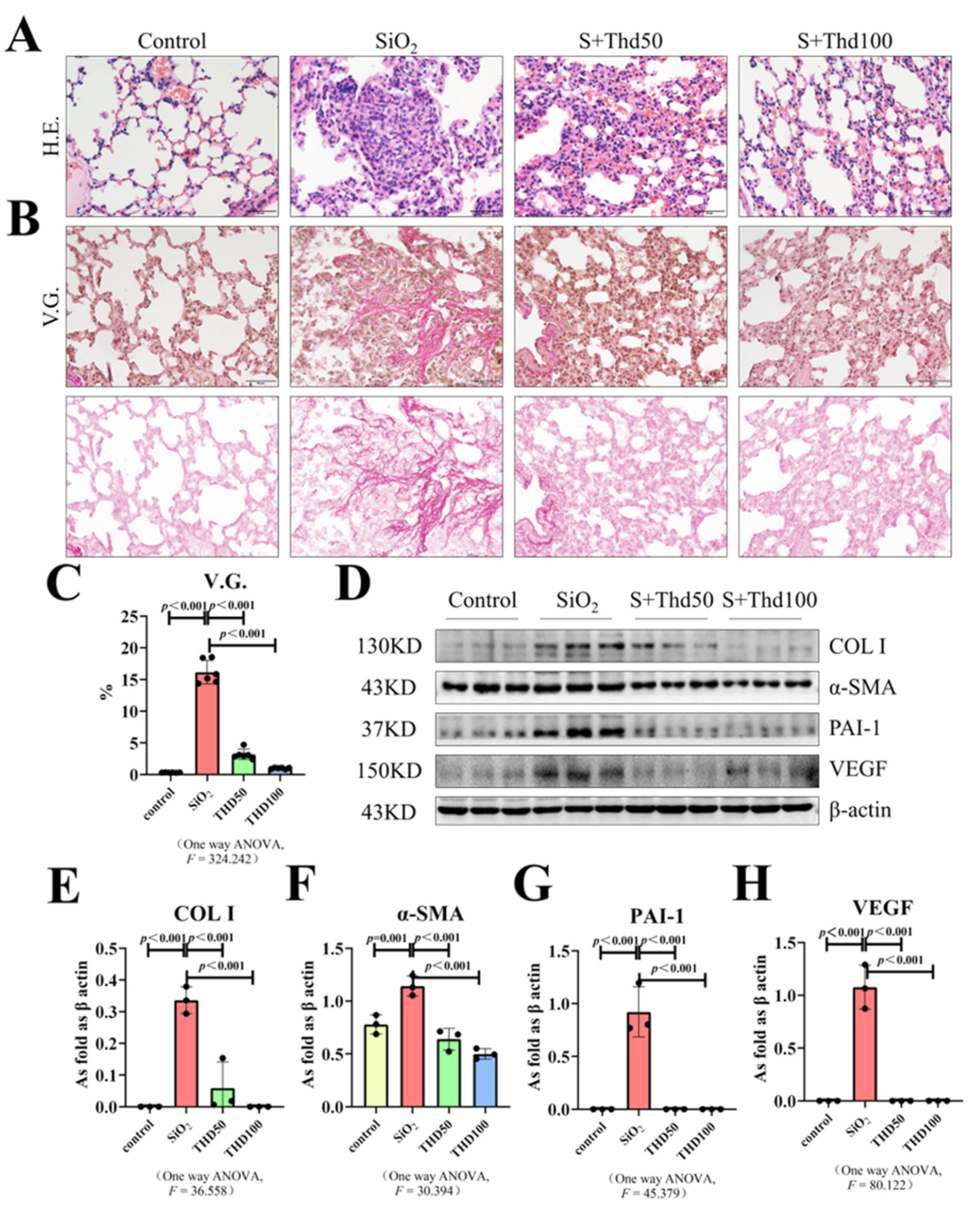{kind=link}
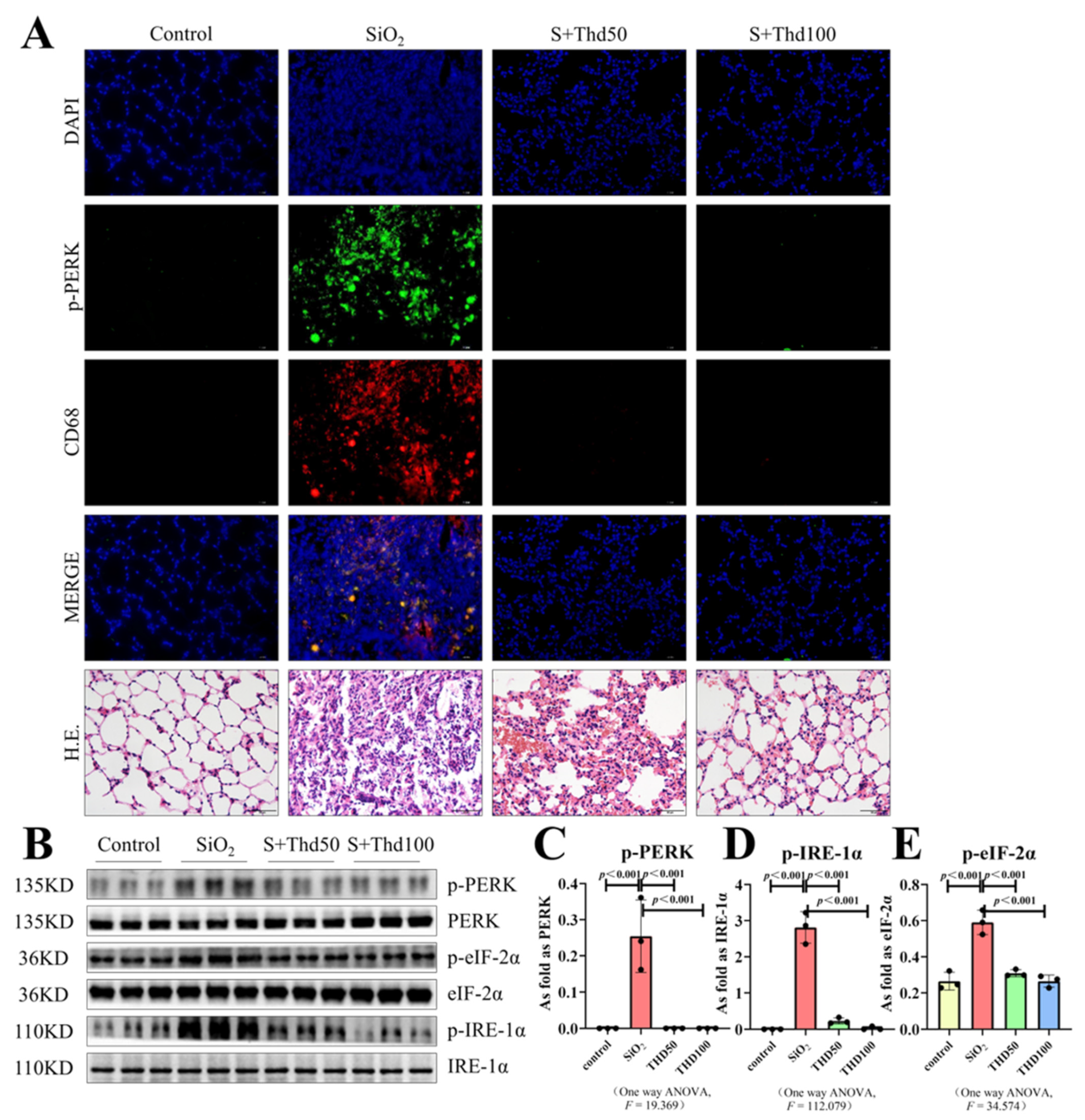{kind=link}
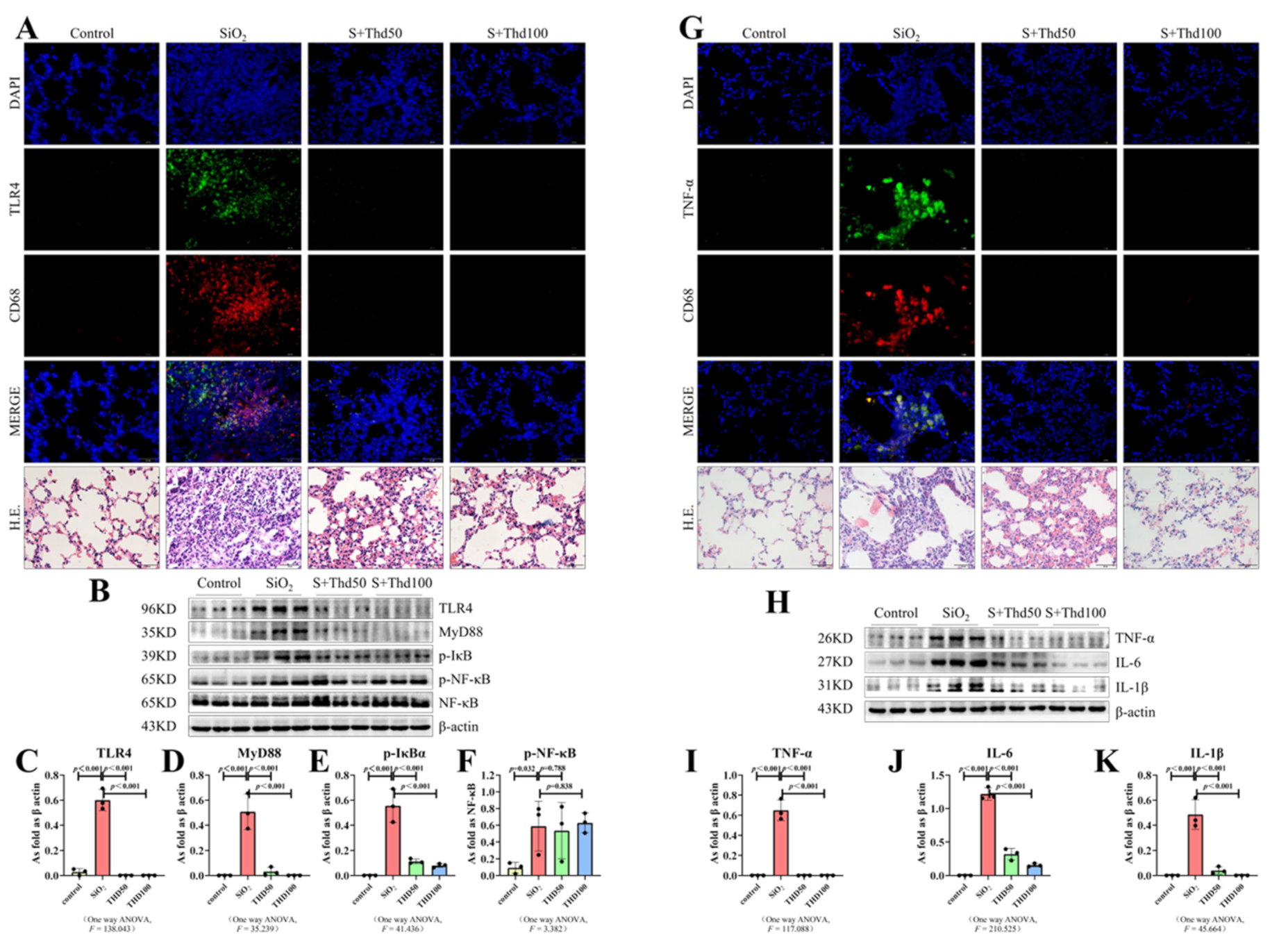{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Thalidomide Reduced Collagen Deposition in Silicotic Mice
2.2. Thalidomide Inhibited ER Stress in Silicotic Mice
2.3. Thalidomide Inhibited the TLR4-NF-κB Pathway and Inflammatory Response in Silicotic Mice
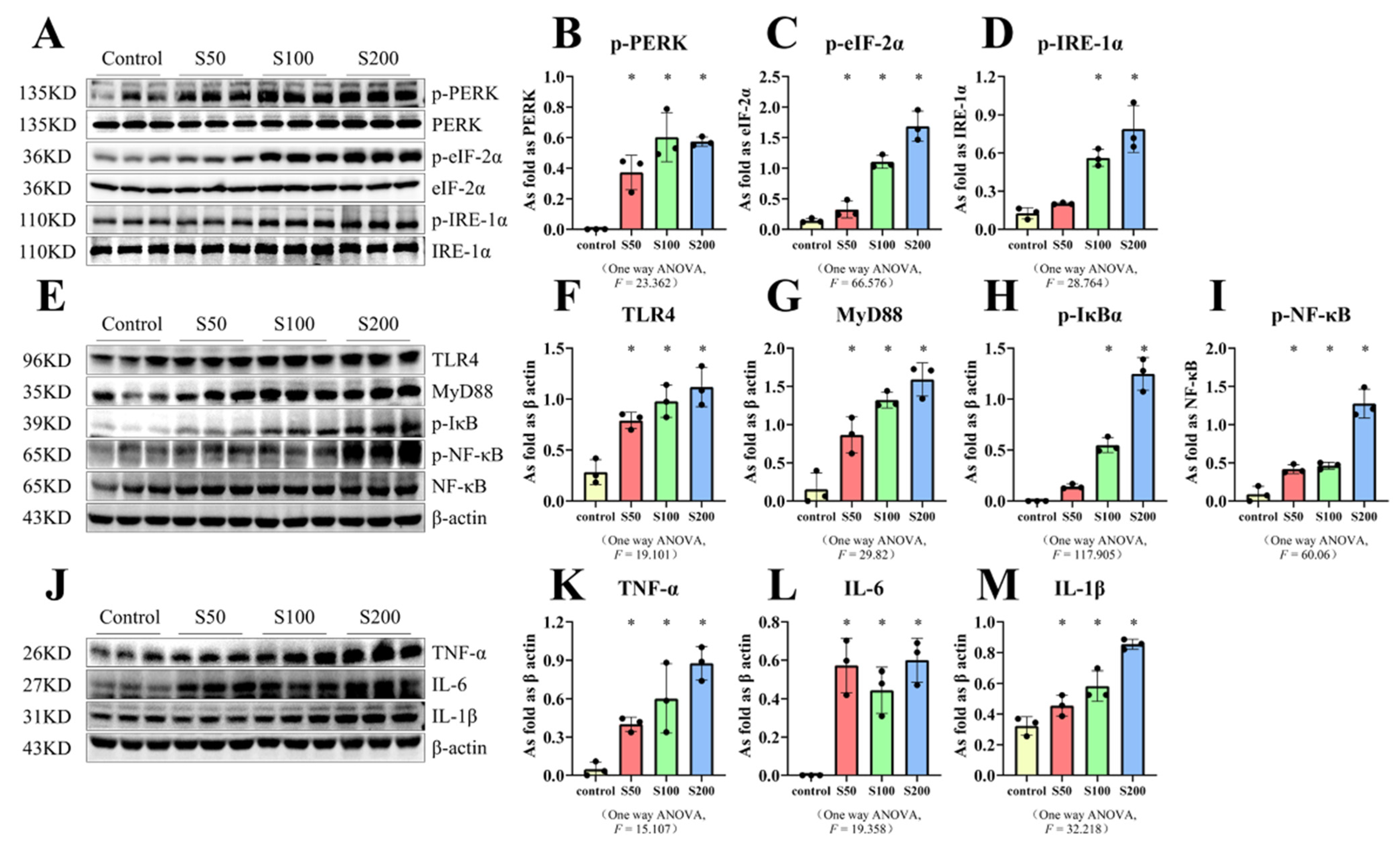
2.4. SiO2 Stimulated the Activation of ER Stress and TLR4-NF-κB as Well as Inflammation-Related Factors in MH-S Cells
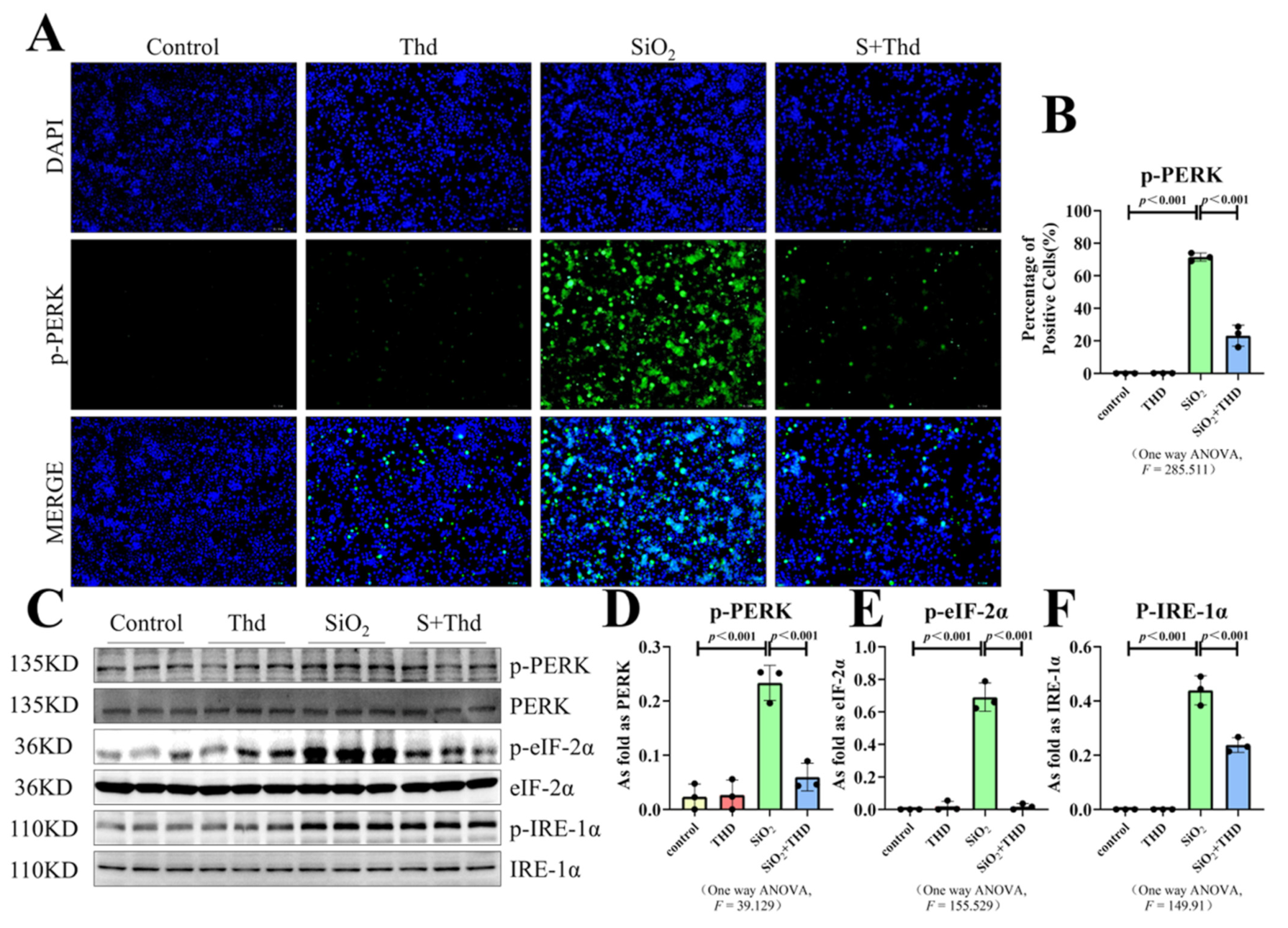
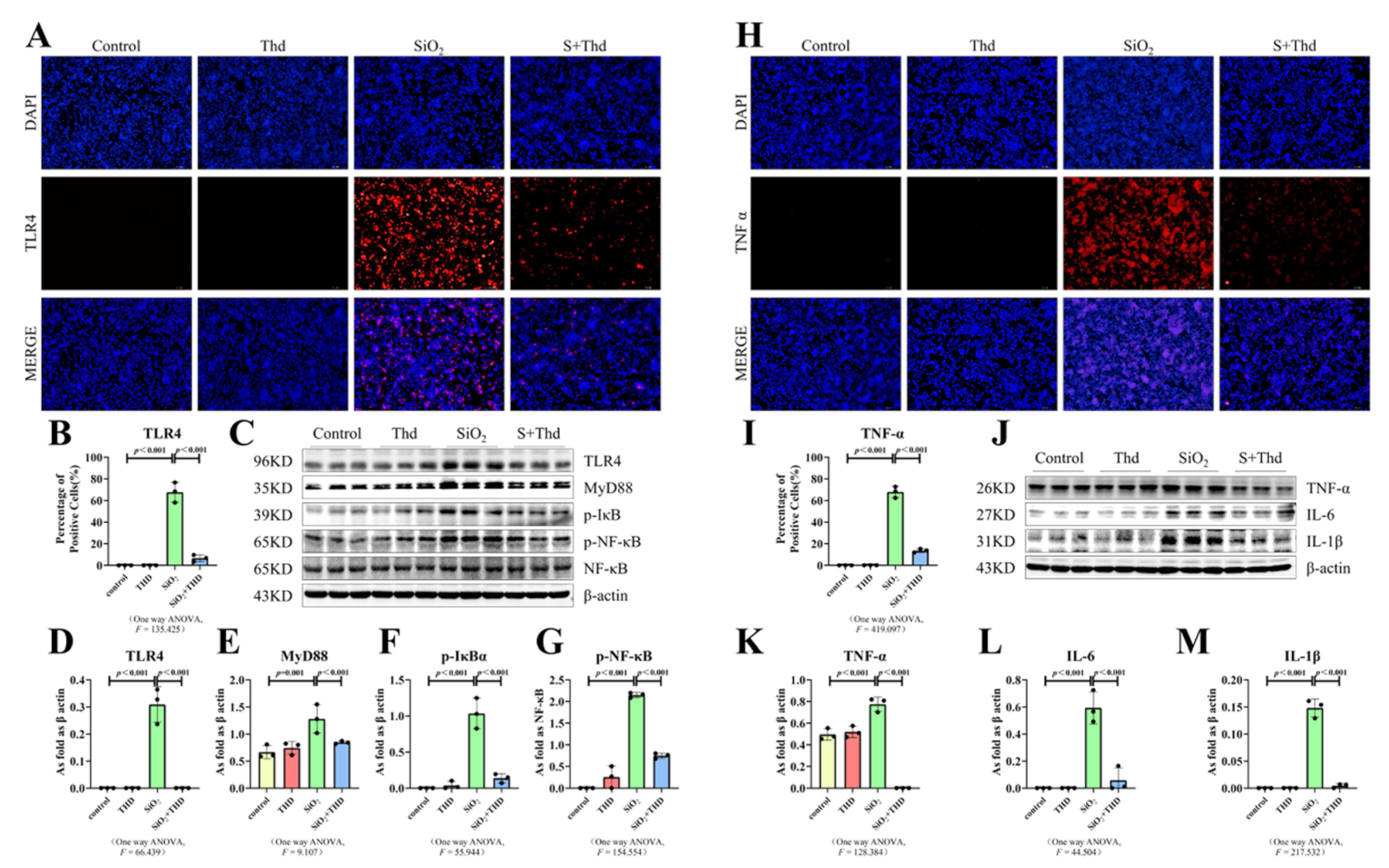
2.5. Thalidomide Inhibited ER Stress and the TLR4-NF-κB Pathway as Well as the Inflammatory Response in MH-S Cells Treated with SiO2
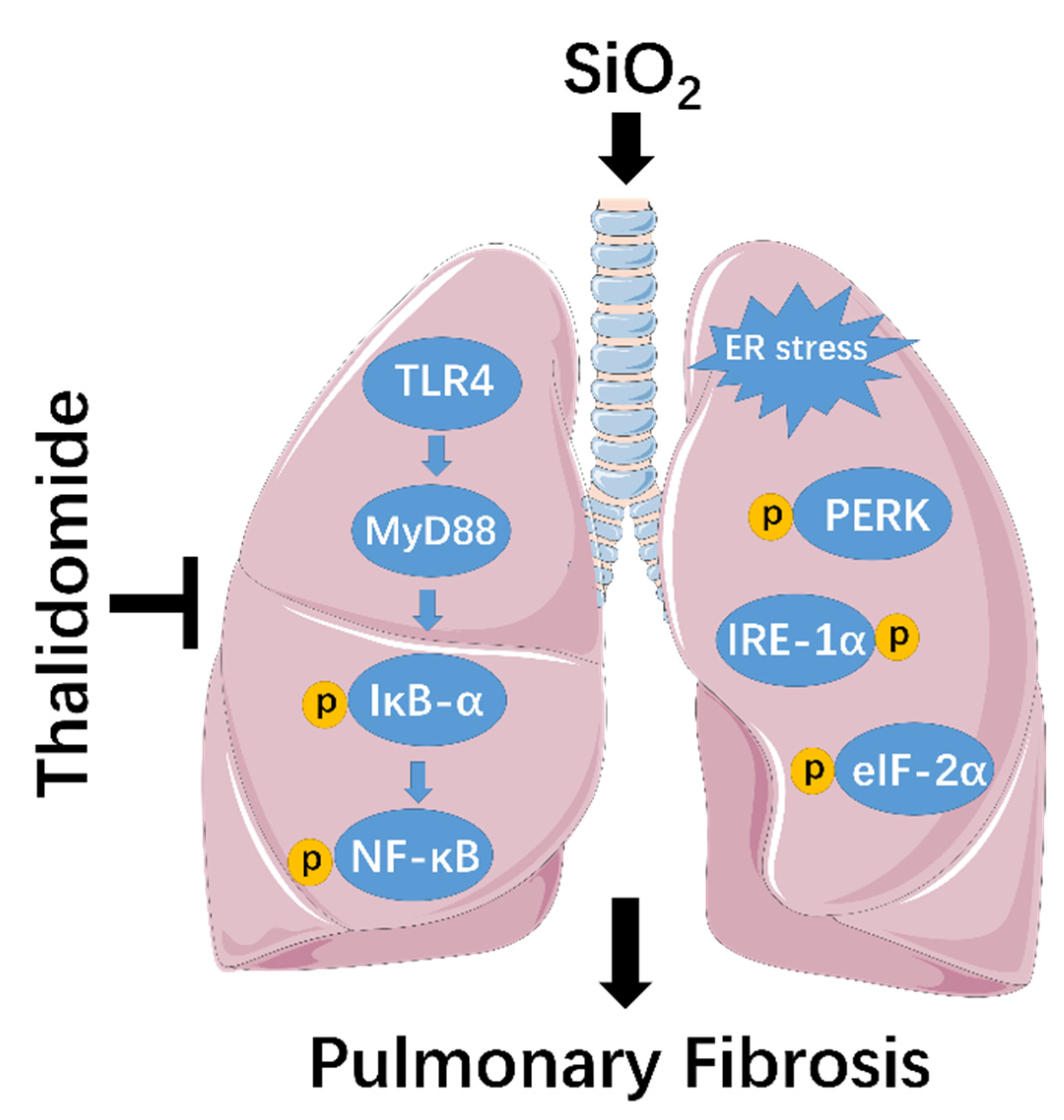
3. Discussion
4. Materials and Method
4.1. Establishment of the Mouse Model
4.2. Cell Culture
4.3. Hematoxylin–Eosin Staining
4.4. Van Gieson’s Staining
4.5. Western Blot
4.6. Immunofluorescence Staining (IF)
4.7. Lung Function Assessment
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoy, R.F.; Chambers, D.C. Silica-related diseases in the modern world. Allergy 2020, 75, 2805–2817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Yang, F.; Sun, Y.; Yuan, Y.; Cheng, H.; Wei, Z.; Li, S.; Cheng, T.; Brann, D.; Wang, R. A new antifibrotic target of Ac-SDKP: Inhibition of myofibroblast differentiation in rat lung with silicosis. PLoS ONE 2012, 7, e40301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shifeng, L.; Hong, X.; Xue, Y.; Siyu, N.; Qiaodan, Z.; Dingjie, X.; Lijuan, Z.; Zhongqiu, W.; Xuemin, G.; Wenchen, C.; et al. Ac-SDKP increases α-TAT 1 and promotes the apoptosis in lung fibroblasts and epithelial cells double-stimulated with TGF-β1 and silica. Toxicol. Appl. Pharmacol. 2019, 369, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, P.; Kropski, J.A.; Jones, M.G.; Lee, J.S.; Rossi, G.; Karampitsakos, T.; Maher, T.M.; Tzouvelekis, A.; Ryerson, C.J. Idiopathic pulmonary fibrosis: Disease mechanisms and drug development. Pharmacol. Ther. 2021, 222, 107798. [Google Scholar] [CrossRef]
- Song, M.-Y.; Wang, J.-X.; Sun, Y.-L.; Han, Z.-F.; Zhou, Y.-T.; Liu, Y.; Fan, T.-H.; Li, Z.-G.; Qi, X.-M.; Luo, Y.; et al. Tetrandrine alleviates silicosis by inhibiting canonical and non-canonical NLRP3 inflammasome activation in lung macrophages. Acta Pharmacol. Sin. 2022, 43, 1274–1284. [Google Scholar] [CrossRef]
- Bando, M.; Yamauchi, H.; Ogura, T.; Taniguchi, H.; Watanabe, K.; Azuma, A.; Homma, S.; Sugiyama, Y.; Japan Pirfenidone Clinical Study Group. Clinical Experience of the Long-term Use of Pirfenidone for Idiopathic Pulmonary Fibrosis. Intern. Med. 2016, 55, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Galli, J.A.; Pandya, A.; Vega-Olivo, M.; Dass, C.; Zhao, H.; Criner, G.J. Pirfenidone and nintedanib for pulmonary fibrosis in clinical practice: Tolerability and adverse drug reactions. Respirology 2017, 22, 1171–1178. [Google Scholar] [CrossRef] [Green Version]
- George, P.M.; Patterson, C.M.; Reed, A.K.; Thillai, M. Lung transplantation for idiopathic pulmonary fibrosis. Lancet Respir. Med. 2019, 7, 271–282. [Google Scholar] [CrossRef]
- Kenyon, B.M.; Browne, F.; D’Amato, R.J. Effects of thalidomide and related metabolites in a mouse corneal model of neovascularization. Exp. Eye Res. 1997, 64, 971–978. [Google Scholar] [CrossRef]
- Mellin, G.W.; Katzenstein, M. The saga of thalidomide. Neuropathy to embryopathy; with case reports of congenital anomalies. N. Engl. J. Med. 1962, 267, 1238–1244. [Google Scholar] [CrossRef]
- Petersen, C.E.; Wiedemann, H.R. Thalidomide and foetal abnormalities. Lancet 1963, 281, 174. [Google Scholar] [CrossRef]
- Franks, M.E.; Macpherson, G.R.; Figg, W.D. Thalidomide. Lancet 2004, 363, 1802–1811. [Google Scholar] [CrossRef] [Green Version]
- Rutter, T. Thalidomide ban to be lifted in the US. BMJ 1997, 315, 697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Li, X.; Li, M.; Chen, M.; Fan, Q.; Wei, W. Antiinflammation and Antioxidant Effects of Thalidomide on Pulmonary Fibrosis in Mice and Human Lung Fibroblasts. Inflammation 2017, 40, 1836–1846. [Google Scholar] [CrossRef]
- Bian, C.; Qin, W.-J.; Zhang, C.-Y.; Zou, G.-L.; Zhu, Y.-Z.; Chen, J.; Zhao, R.; Wang, Y.-Y.; Zhe, H. Thalidomide (THD) alleviates radiation induced lung fibrosis (RILF) via down-regulation of TGF-β/Smad3 signaling pathway in an Nrf2-dependent manner. Free Radic. Biol. Med. 2018, 129, 446–453. [Google Scholar] [CrossRef]
- Tanjore, H.; Lawson, W.E.; Blackwell, T.S. Endoplasmic reticulum stress as a pro-fibrotic stimulus. Biochim. Biophys. Acta 2013, 1832, 940–947. [Google Scholar] [CrossRef] [Green Version]
- Mao, N.; Fan, Y.; Liu, W.; Yang, H.; Yang, Y.; Li, Y.; Jin, F.; Li, T.; Yang, X.; Gao, X.; et al. Oxamate Attenuates Glycolysis and ER Stress in Silicotic Mice. Int. J. Mol. Sci. 2022, 23, 3013. [Google Scholar] [CrossRef]
- Li, T.; Yang, X.-Y.; Xu, D.-J.; Gao, Z.-Y.; Gao, Y.-B.; Jin, F.-Y.; Li, Y.-Q.; Liu, S.-P.; Li, S.-F.; Gao, X.-M.; et al. OC-STAMP Overexpression Drives Lung Alveolar Epithelial Cell Type II Senescence in Silicosis. Oxid. Med. Cell. Longev. 2021, 2021, 4158495. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, D.; Li, Q.; Yang, Y.; Xu, H.; Wei, Z.; Wang, R.; Zhang, W.; Liu, Y.; Geng, Y.; et al. N-acetyl-seryl-aspartyl-lysyl-proline (Ac-SDKP) attenuates silicotic fibrosis by suppressing apoptosis of alveolar type II epithelial cells via mediation of endoplasmic reticulum stress. Toxicol. Appl. Pharmacol. 2018, 350, 1–10. [Google Scholar] [CrossRef]
- Jin, F.; Geng, F.; Xu, D.; Li, Y.; Li, T.; Yang, X.; Liu, S.; Zhang, H.; Wei, Z.; Li, S.; et al. Ac-SDKP Attenuates Activation of Lung Macrophages and Bone Osteoclasts in Rats Exposed to Silica by Inhibition of TLR4 and RANKL Signaling Pathways. J. Inflamm. Res. 2021, 14, 1647–1660. [Google Scholar] [CrossRef]
- Noman, A.S.M.; Koide, N.; Hassan, F.; E.-Khuda, I.; Dagvadorj, J.; Tumurkhuu, G.; Islam, S.; Naiki, Y.; Yoshida, T.; Yokochi, T. Thalidomide inhibits lipopolysaccharide-induced tumor necrosis factor-α production via down-regulation of MyD88 expression. Innate Immun. 2009, 15, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, S.; Xue, A.; Shi, J.; Zheng, C.; Huang, Y. Thalidomide Inhibits Angiogenesis via Downregulation of VEGF and Angiopoietin-2 in Crohn’s Disease. Inflammation 2020, 44, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Burman, A.; Tanjore, H.; Blackwell, T.S. Endoplasmic reticulum stress in pulmonary fibrosis. Matrix Biol. 2018, 68–69, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Dancsok, A.R.; Gao, D.; Lee, A.F.; Steigen, S.E.; Blay, J.-Y.; Thomas, D.M.; Maki, R.G.; Nielsen, T.O.; Demicco, E.G. Tumor-associated macrophages and macrophage-related immune checkpoint expression in sarcomas. OncoImmunology 2020, 9, 1747340. [Google Scholar] [CrossRef] [Green Version]
- Hui, Z.; Dingjie, X.; Yuan, Y.; Zhongqiu, W.; Na, M.; Mingjian, B.; Yu, G.; Guangyuan, L.; Xu, H.; Shifeng, L.; et al. Silicosis decreases bone mineral density in rats. Toxicol. Appl. Pharmacol. 2018, 348, 117–122. [Google Scholar] [CrossRef]
- Liu, G.; Lu, Y.; Shi, L.; Ren, Y.; Kong, J.; Zhang, M.; Chen, M.; Liu, W. TLR4-MyD88 signaling pathway is responsible for acute lung inflammation induced by reclaimed water. J. Hazard. Mater. 2020, 396, 122586. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, W.; Wang, Y. STING is an essential regulator of heart inflammation and fibrosis in mice with pathological cardiac hypertrophy via endoplasmic reticulum (ER) stress. Biomed. Pharmacother. 2020, 125, 110022. [Google Scholar] [CrossRef]
- Delbrel, E.; Soumare, A.; Naguez, A.; Label, R.; Bernard, O.; Bruhat, A.; Fafournoux, P.; Tremblais, G.; Marchant, D.; Gille, T.; et al. HIF-1α triggers ER stress and CHOP-mediated apoptosis in alveolar epithelial cells, a key event in pulmonary fibrosis. Sci. Rep. 2018, 8, 17939. [Google Scholar] [CrossRef]
- Cybulsky, A.V. Endoplasmic reticulum stress, the unfolded protein response and autophagy in kidney diseases. Nat. Rev. Nephrol. 2017, 13, 681–696. [Google Scholar] [CrossRef]
- Qin, X.; Lin, X.; Liu, L.; Li, Y.; Li, X.; Deng, Z.; Chen, H.; Chen, H.; Niu, Z.; Li, Z.; et al. Macrophage-derived exosomes mediate silica-induced pulmonary fibrosis by activating fibroblast in an endoplasmic reticulum stress-dependent manner. J. Cell. Mol. Med. 2021, 25, 4466–4477. [Google Scholar] [CrossRef]
- Oakes, S.A.; Papa, F.R. The Role of Endoplasmic Reticulum Stress in Human Pathology. Annu. Rev. Pathol. Mech. Dis. 2015, 10, 173–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Chen, S.; Zhang, Y.; Xu, H.; Xu, D.; Wei, Z.; Gao, X.; Cai, W.; Mao, N.; Zhang, L.; et al. Matrix stiffness regulates α-TAT1-mediated acetylation of α-tubulin and promotes silica-induced epithelial–mesenchymal transition via DNA damage. J. Cell Sci. 2021, 134, jcs243394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-X.; Yuan, J.; Li, R.-S. Thalidomide Mitigates Apoptosis via Endoplasmic Reticulum Stress in Diabetic Nephropathy. Endocrine Metab. Immune Disord. Drug Targets 2021, 22, 1. [Google Scholar] [CrossRef]
- Puskás, L.G.; Fehér, L.Z.; Vizler, C.; Ayaydin, F.; Rásó, E.; Molnár, E.; Magyary, I.; Kanizsai, I.; Gyuris, M.; Madácsi, R.; et al. Polyunsaturated fatty acids synergize with lipid droplet binding thalidomide analogs to induce oxidative stress in cancer cells. Lipids Health Dis. 2010, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2020, 78, 1233–1261. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lu, B.; Fu, J.; Zhu, X.; Song, E.; Song, Y. Amorphous silica nanoparticles induce inflammation via activation of NLRP3 inflammasome and HMGB1/TLR4/MYD88/NF-kb signaling pathway in HUVEC cells. J. Hazard. Mater. 2021, 404, 124050. [Google Scholar] [CrossRef]
- Benmerzoug, S.; Rose, S.; Bounab, B.; Gosset, D.; Duneau, L.; Chenuet, P.; Mollet, L.; Le Bert, M.; Lambers, C.; Geleff, S.; et al. STING-dependent sensing of self-DNA drives silica-induced lung inflammation. Nat. Commun. 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Ishikawa, M.; Kanno, S.-I.; Takayanagi, M.; Takayanagi, Y.; Sasaki, K.-I. Thalidomide Promotes the Release of Tumor Necrosis Factor-ALPHA. (TNF-ALPHA.) and Lethality by Lipopolysaccharide in Mice. Biol. Pharm. Bull. 1998, 21, 638–640. [Google Scholar] [CrossRef] [Green Version]
- Maciel-Fiuza, M.F.; Costa, P.D.S.S.; Kowalski, T.W.; Schuler-Faccini, L.; Bonamigo, R.R.; Vetoratto, R.; Eidt, L.M.; de Moraes, P.C.; Silveira, M.I.D.S.; Camargo, L.M.A.; et al. Evaluation of Polymorphisms in Toll-Like Receptor Genes as Biomarkers of the Response to Treatment of Erythema Nodosum Leprosum. Front. Med. 2022, 8, 713143. [Google Scholar] [CrossRef]
- Han, J.; Ye, S.; Zou, C.; Chen, T.; Wang, J.; Li, J.; Jiang, L.; Xu, J.; Huang, W.; Wang, Y.; et al. Angiotensin II Causes Biphasic STAT3 Activation Through TLR4 to Initiate Cardiac Remodeling. Hypertension 2018, 72, 1301–1311. [Google Scholar] [CrossRef]
- Dange, R.B.; Agarwal, D.; Masson, G.S.; Vila, J.; Wilson, B.; Nair, A.; Francis, J. Central blockade of TLR4 improves cardiac function and attenuates myocardial inflammation in angiotensin II-induced hypertension. Cardiovasc. Res. 2014, 103, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Fazzi, P.; Manni, E.; Cristofani, R.; Cei, G.; Piazza, S.; Calabrese, R.; Antonelli, A.; Siciliano, G.; Barachini, P.; Carpi, A. Thalidomide for improving cutaneous and pulmonary sarcoidosis in patients resistant or with contraindications to corticosteroids. Biomed. Pharmacother. 2012, 66, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, Y.; Xu, H.; Wei, Z.; Yang, Y.; Jin, F.; Zhang, M.; Wang, C.; Song, W.; Huo, J.; et al. ACE2 Attenuates Epithelial-Mesenchymal Transition in MLE-12 Cells Induced by Silica. Drug Des. Dev. Ther. 2020, 14, 1547–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-A.; Jang, H.-J.; Hwang, D.H. Toll-like receptor 4-induced endoplasmic reticulum stress contributes to impairment of vasodilator action of insulin. Am. J. Physiol. Metab. 2015, 309, E767–E776. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Paul, J.; Wang, Y.; Gupta, M.; Vang, D.; Thompson, S.; Jha, R.; Nguyen, J.; Valverde, Y.; Lamarre, Y.; et al. Heme Causes Pain in Sickle Mice via Toll-Like Receptor 4-Mediated Reactive Oxygen Species- and Endoplasmic Reticulum Stress-Induced Glial Activation. Antioxidants Redox Signal. 2021, 34, 279–293. [Google Scholar] [CrossRef]
- Zheng, F.; Zhu, J.; Zhang, W.; Fu, Y.; Lin, Z. Thal protects against paraquat-induced lung injury through a microRNA-141/HDAC6/IκBα-NF-κB axis in rat and cell models. Basic Clin. Pharmacol. Toxicol. 2020, 128, 334–347. [Google Scholar] [CrossRef]
- Tabata, C.; Tabata, R.; Takahashi, Y.; Nakamura, K.; Nakano, T. Thalidomide prevents cigarette smoke extract-induced lung damage in mice. Int. Immunopharmacol. 2015, 25, 511–517. [Google Scholar] [CrossRef]
- Ito, T.; Ando, H.; Suzuki, T.; Ogura, T.; Hotta, K.; Imamura, Y.; Yamaguchi, Y.; Handa, H. Identification of a Primary Target of Thalidomide Teratogenicity. Science 2010, 327, 1345–1350. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Li, Y.; Zhang, Y.; Li, S.; Zhang, M.; Jin, F.; Wei, Z.; Yang, Y.; Gao, X.; Mao, N.; et al. N-Acetyl-Seryl-Asparyl-Lysyl-Proline regulates lung renin angiotensin system to inhibit epithelial–mesenchymal transition in silicotic mice. Toxicol. Appl. Pharmacol. 2020, 408, 115255. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Cai, W.; Jin, F.; Wang, X.; Liu, W.; Li, T.; Yang, X.; Liu, H.; Xu, H.; Yang, F. Thalidomide Alleviates Pulmonary Fibrosis Induced by Silica in Mice by Inhibiting ER Stress and the TLR4-NF-κB Pathway. Int. J. Mol. Sci. 2022, 23, 5656. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105656
Li Y, Cai W, Jin F, Wang X, Liu W, Li T, Yang X, Liu H, Xu H, Yang F. Thalidomide Alleviates Pulmonary Fibrosis Induced by Silica in Mice by Inhibiting ER Stress and the TLR4-NF-κB Pathway. International Journal of Molecular Sciences. 2022; 23(10):5656. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105656
Chicago/Turabian StyleLi, Yaqian, Wenchen Cai, Fuyu Jin, Xiaojing Wang, Wenjing Liu, Tian Li, Xinyu Yang, Heliang Liu, Hong Xu, and Fang Yang. 2022. "Thalidomide Alleviates Pulmonary Fibrosis Induced by Silica in Mice by Inhibiting ER Stress and the TLR4-NF-κB Pathway" International Journal of Molecular Sciences 23, no. 10: 5656. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105656