
Transcriptome and Physiological Analyses of a Navel Orange Mutant with Improved Drought Tolerance and Water Use Efficiency Caused by Increases of Cuticular Wax Accumulation and ROS Scavenging Capacity

 and
and
Abstract
:1. Introduction
2. Results
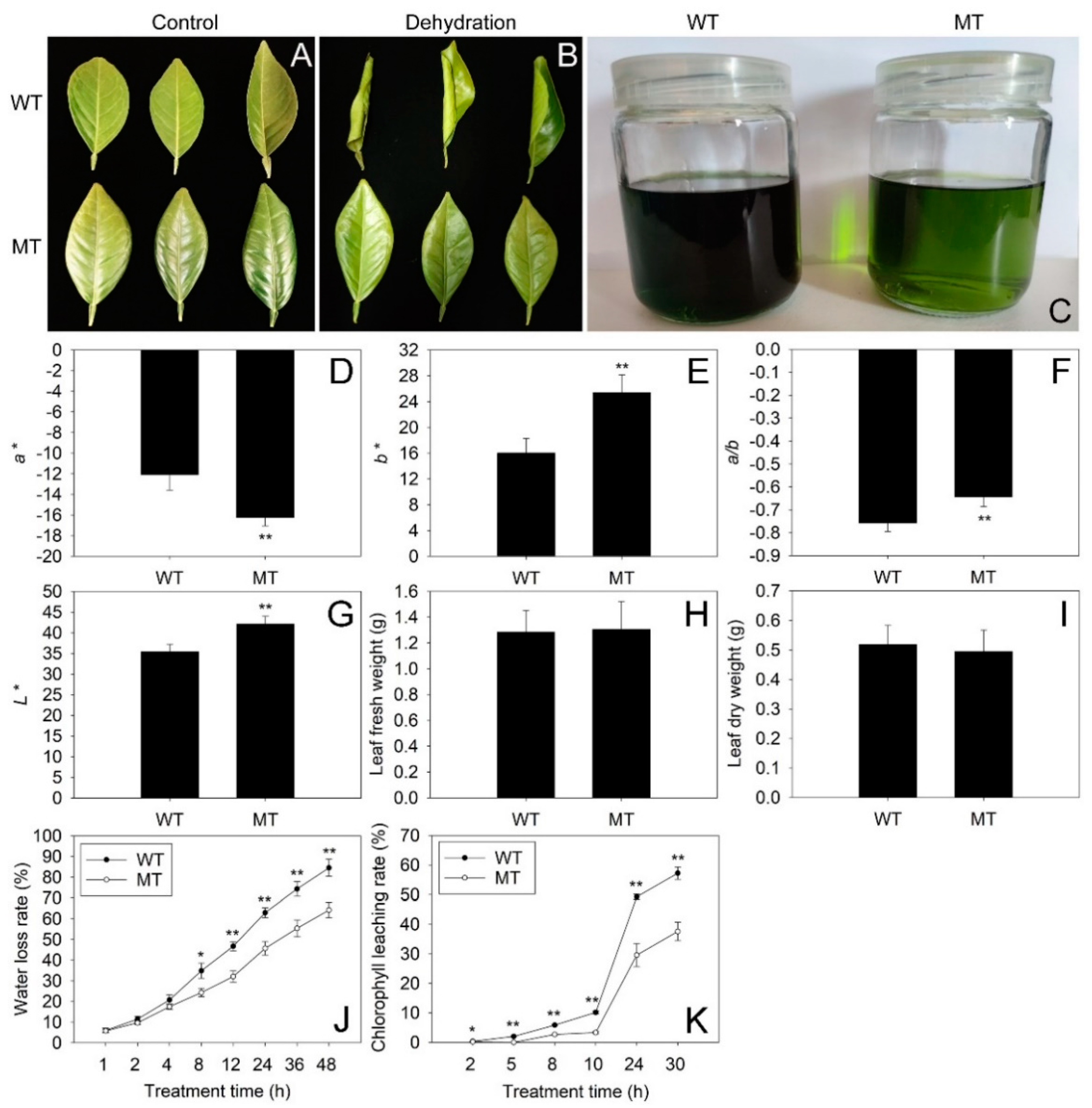
2.1. Comparison of Phenotype, Chromatic Aberration and Cuticular Permeability between WT and MT Leaves
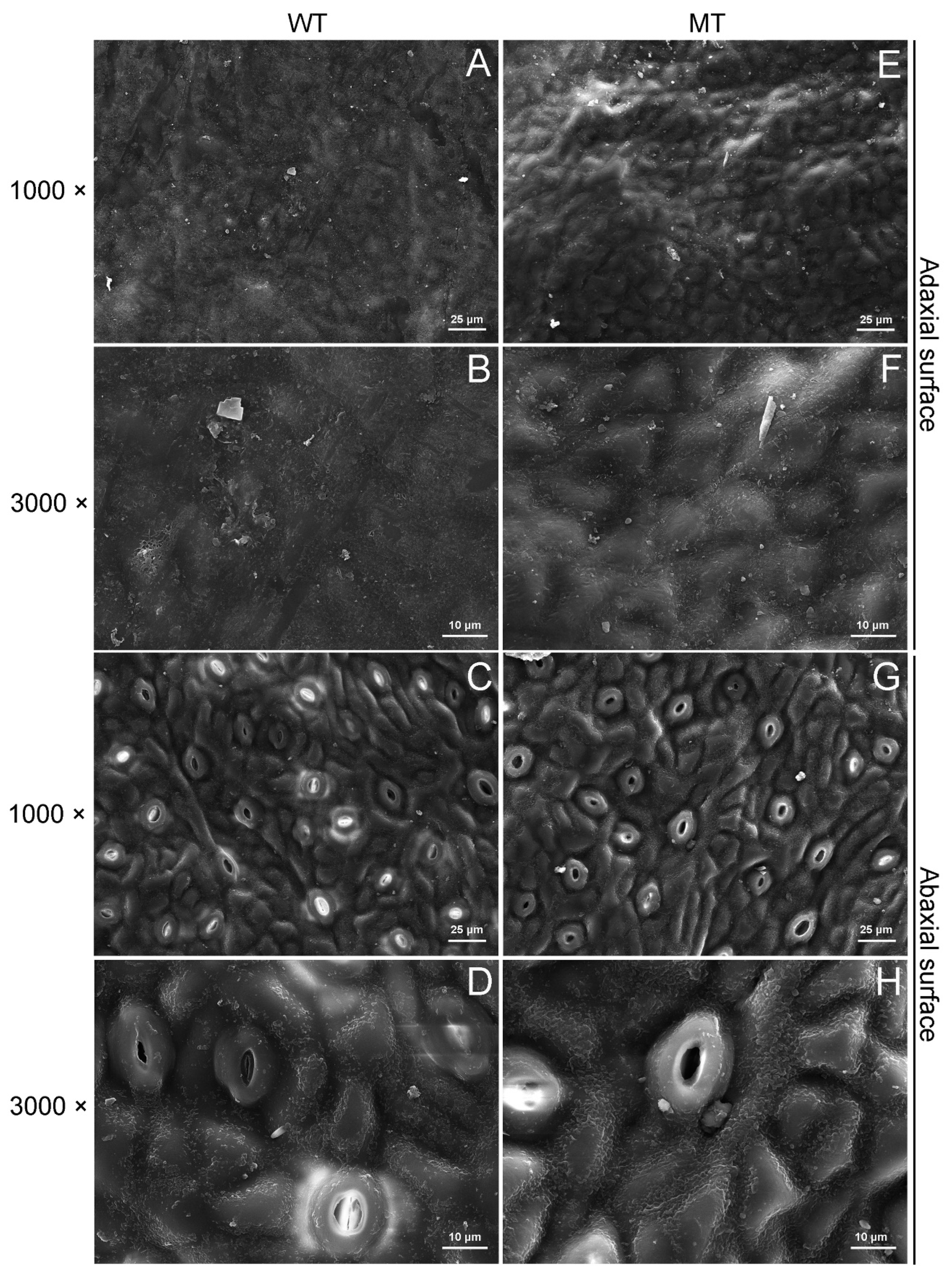
2.2. Comparison of Cuticular Wax Morphology and Chemical Composition between WT and MT Leaves
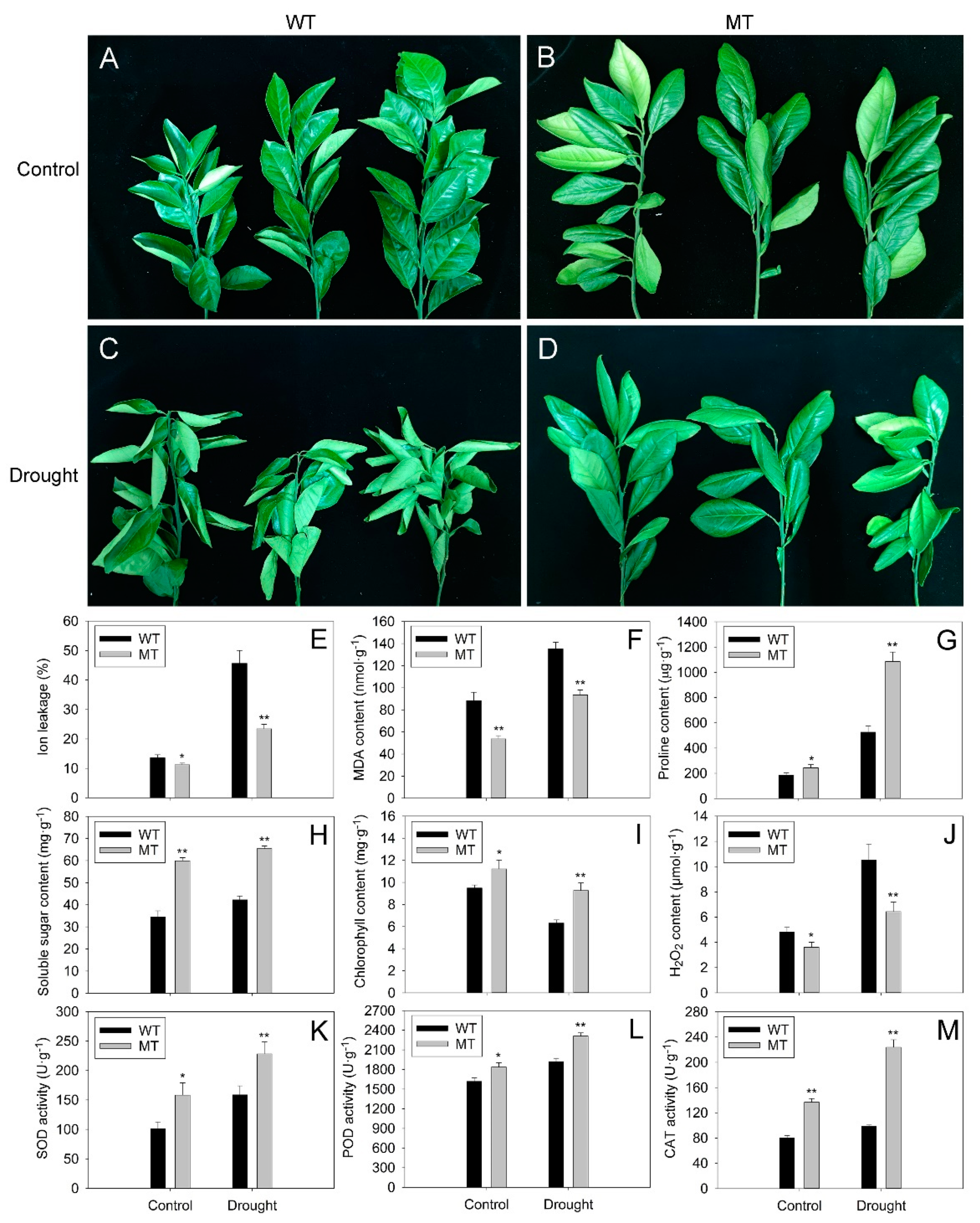
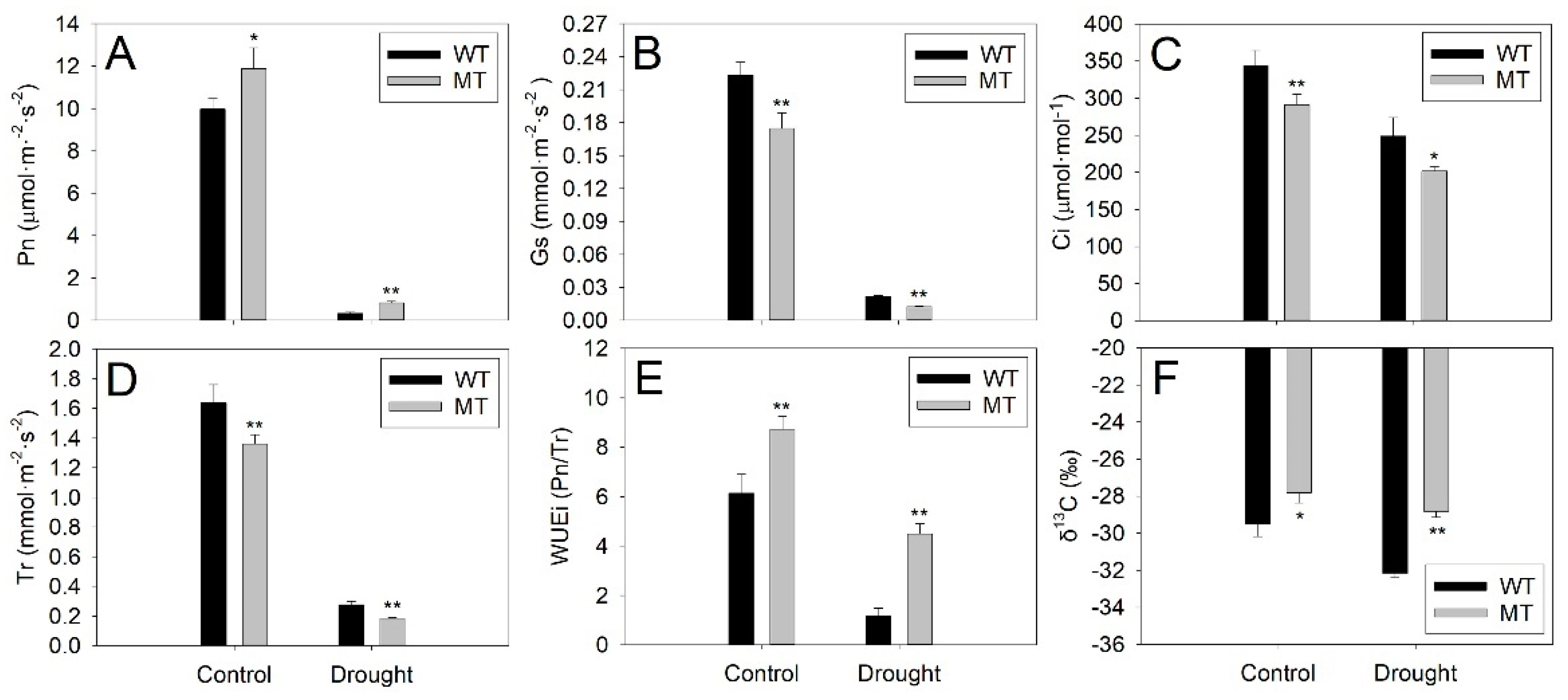
2.3. Comparison of Morphological and Physiological Responses of WT and MT Plants to Drought Stress
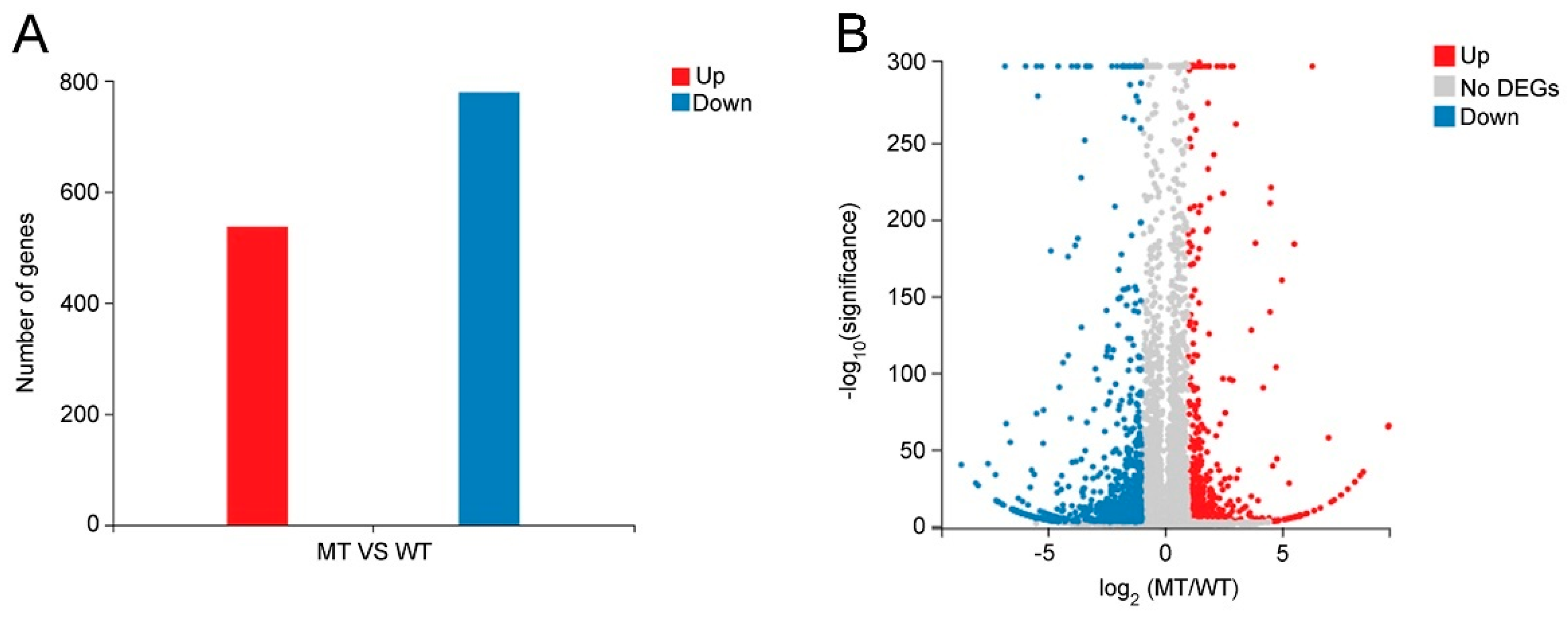
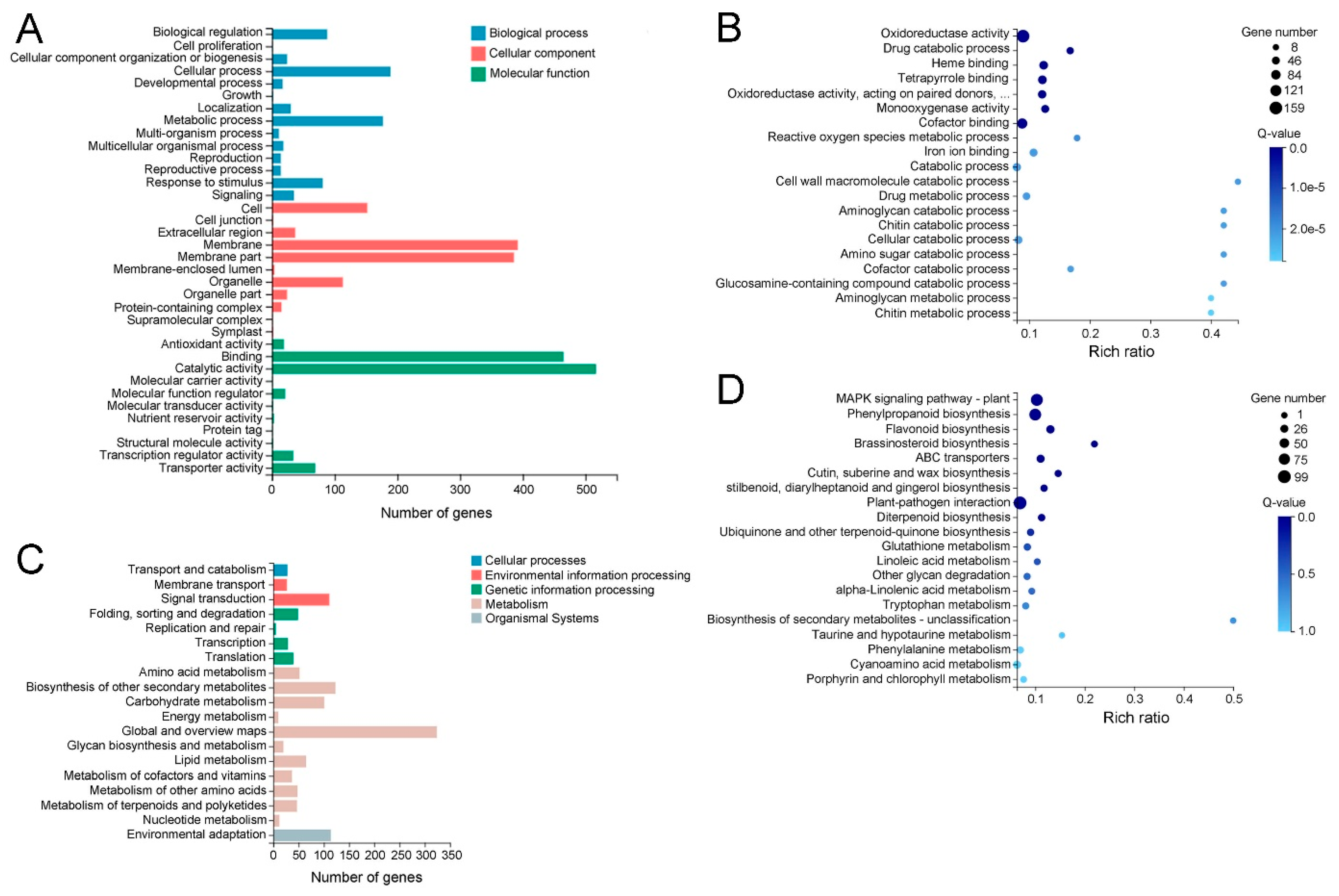
2.4. Functional Classification of Differentially Expressed Genes (DEGs) between WT and MT Leaves
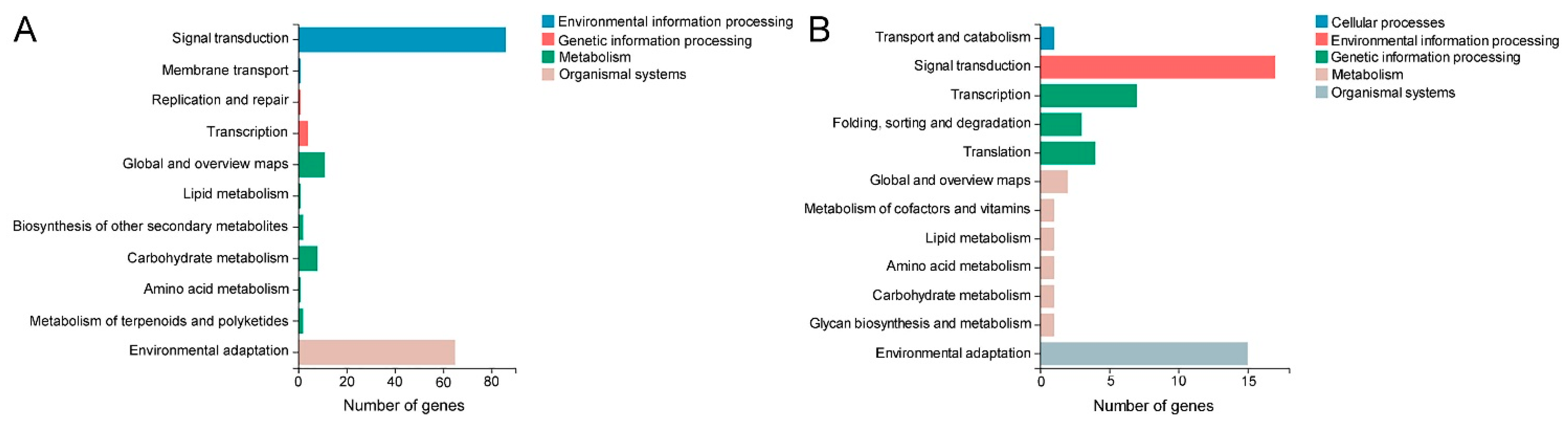
2.5. Identification of DEGs Involved in Cuticular Wax Biosynthesis and Transport
2.6. Identification of DEGs Involved in MAPK Cascade, Reactive Oxygen Species (ROS) Scavenging and Drought Response
2.7. Identification of DEGs Encoding Transcription Factor
2.8. Expression Analysis of Wax Biosynthesis, Transport and Drought Responsive DEGs by qRT-PCR
3. Discussion
3.1. The Decrease in Cuticular Permeability was Caused by the Increase in Aliphatic Wax Accumulation in MT Leaves
3.2. Increased Cuticular Wax Accumulation and Enhanced ROS Scavenging Capacity Contributes to the Improvement of Drought Tolerance and WUE in MT Plants
3.3. The Changes in Expression Levels of Wax Biosynthesis and Export Genes Contributed to the Alterations in Cuticular Wax Accumulation of MT Leaves
3.4. The DEGs Involved in the MAPK Signaling Pathway-Plant, ROS Scavenging and Other Enriched Pathways Might Contribute to Improve MT Tolerance to Drought Stress
3.5. The DEGs Encode Transcription Factor May Play an Important Role in Enhancing MT Tolerance to Drought Stress
4. Materials and Methods
4.1. Plant Materials
4.2. Analysis of Leaf Chromatic Aberration, Fresh and Dry Weight, Water Loss Rate and Chlorophyll Leaching Rate
4.3. SEM Analysis
4.4. Cuticular Wax Extraction and Analysis
4.5. Analysis of Physiological Indexes in WT and MT Leaves under Control and Drought Treatment
4.6. Total RNA Extraction and Transcriptome Sequencing
4.7. qRT-PCR Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Joubès, J. Arabidopsis cuticular waxes: Advances in synthesis, export and regulation. Prog. Lipid Res. 2013, 52, 110–129. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Suh, M.C. Advances in the understanding of cuticular waxes in Arabidopsis thaliana and crop species. Plant Cell Rep. 2015, 34, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Xu, W.; Yu, X.; Shen, H.; Li, H.; Cheng, D.; Liu, A.; Liu, J.; Liu, C.; Zhao, S.; et al. Cuticular wax accumulation is associated with drought tolerance in wheat near-isogenic lines. Front. Plant Sci. 2016, 7, 1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, P.J.; Lee, S.B.; Suh, M.C.; Park, M.J.; Go, Y.S.; Park, C.M. The MYB96 transcription factor regulates cuticular wax biosynthesis under drought conditions in Arabidopsis. Plant Cell 2011, 23, 1138–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.L.; Hu, X.; Zhang, Y.L.; Liu, Y.; Wang, G.L.; You, C.X.; Li, Y.Y.; Hao, Y.J. An apple long-chain acyl-CoA synthetase 2 gene enhances plant resistance to abiotic stress by regulating the accumulation of cuticular wax. Tree Physiol. 2020, 40, 1450–1465. [Google Scholar] [CrossRef]
- Zhou, L.; Ni, E.; Yang, J.; Zhou, H.; Liang, H.; Li, J.; Jiang, D.; Wang, Z.; Liu, Z.; Zhuang, C. Rice OsGL16 is involved in leaf cuticular wax accumulation and drought resistance. PLoS ONE 2013, 8, e65139. [Google Scholar] [CrossRef]
- Premachandra, G.S.; Hahn, D.T.; Axtell, J.D.; Joly, R.J. Epicuticular wax load and water-use efficiency in bloomless and sparse-bloom mutants of Sorghum bicolor L. Environ. Exp. Bot. 1994, 34, 293–301. [Google Scholar] [CrossRef]
- Cheng, C.; Hu, S.; Han, Y.; Xia, D.; Huang, B.L.; Wu, W.; Hussain, J.; Zhang, X.; Huang, B. Yellow nutsedge WRI4-like gene improves drought tolerance in Arabidopsis thaliana by promoting cuticular wax biosynthesis. BMC Plant Biol. 2020, 20, 498. [Google Scholar] [CrossRef]
- Kang, S.; Hao, X.; Du, T.; Tong, L.; Su, X.; Lu, H.; Li, X.; Huo, Z.; Li, S.; Ding, R. Improving agricultural water productivity to ensure food security in China under changing environment: From research to practice. Agric. Water Manag. 2017, 179, 5–17. [Google Scholar] [CrossRef]
- Rodríguez-Gamir, J.; Primo-Millo, E.; Forner, J.B.; Forner-Giner, M.A. Citrus rootstock responses to water. Sci. Hortic. 2010, 126, 95–102. [Google Scholar] [CrossRef]
- Miranda, M.T.; da Silva, S.F.; Moura, B.B.; Hayashi, A.H.; Machado, E.C.; Ribeiro, R.V. Hydraulic redistribution in Citrus rootstocks under drought. Theor. Exp. Plant Physiol. 2018, 30, 165–172. [Google Scholar] [CrossRef]
- Miranda, M.T.; da Silva, S.F.; Silveira, N.M.; Pereira, L.; Machado, E.C.; Ribeiro, R.V. Root osmotic adjustment and stomatal control of leaf gas exchange are dependent on citrus rootstocks under water deficit. J. Plant Growth Regul. 2021, 40, 11–19. [Google Scholar] [CrossRef]
- Sampaio, A.H.R.; Silva, R.O.; Brito, R.B.F.; Filho, W.S.S.; Gesteira, D.A.S.; Souza, L.D.; Filho, M.A.C. Sweet orange acclimatisation to water stress: A rootstock dependency. Sci. Hortic. 2021, 276, 109727. [Google Scholar] [CrossRef]
- Khan, F.S.; Gan, Z.M.; Li, E.Q.; Ren, M.K.; Hu, C.G.; Zhang, J.Z. Transcriptomic and physiological analysis reveals interplay between salicylic acid and drought stress in citrus tree floral initiation. Planta 2022, 255, 24. [Google Scholar] [CrossRef]
- Baker, E.A.; Procopiou, J.; Hunt, G.M. The cuticles of citrus species. composition of leaf and fruit waxes. J. Sci. Food Agric. 1975, 26, 1093–1101. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, Z.; Zhu, C.; Yue, J.; Yang, H.; Li, J.; Gao, J.; Xu, R.; Deng, X.; Cheng, Y. Variations of membrane fatty acids and epicuticular wax metabolism in response to oleocellosis in lemon fruit. Food Chem. 2021, 338, 127684. [Google Scholar] [CrossRef]
- Nordby, H.E.; Mcdonald, R.E. Variations in chilling injury and epicuticular wax composition of white grapefruit with canopy position and fruit development during the season. J. Agric. Food Chem. 1995, 43, 1828–1833. [Google Scholar] [CrossRef]
- Ding, S.; Zhang, J.; Wang, R.; Ou, S.; Shan, Y. Changes in cuticle compositions and crystal structure of ‘Bingtang’ sweet orange fruits (Citrus sinensis) during storage. Int. J. Food Prop. 2018, 21, 2411–2427. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.; Zhang, J.; Yang, L.; Wang, X.; Fu, F.; Wang, R.; Zhang, Q.; Shan, Y. Changes in cuticle components and morphology of ‘Satsuma’ Mandarin (Citrus unshiu) during ambient storage and their potential role on Penicillium digitatum infection. Molecules 2020, 25, 412. [Google Scholar] [CrossRef] [Green Version]
- Cajuste, J.F.; González-Candelas, L.; Veyrat, A.; García-Breijo, F.J.; Reig-Armiñana, J.; Lafuente, M.T. Epicuticular wax content and morphology as related to ethylene and storage performance of ‘Navelate’ orange fruit. Postharvest Biol. Technol. 2010, 55, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.C.; Yang, L.; Zheng, Q.; Wang, Y.C.; Wang, M.L.; Zhuang, X.; Wu, Q.; Liu, C.F.; Liu, S.B.; Liu, Y. Analysis of cuticular wax constituents and genes that contribute to the formation of ‘glossy Newhall’, a spontaneous bud mutant from the wild-type ‘Newhall’ navel orange. Plant Mol. Biol. 2015, 88, 573–590. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Han, J.; Liu, R.; Ding, Y.; Wang, J.; Sun, L.; Yang, X.; Zeng, Y.; Wen, W.; Xu, J.; et al. Integrated transcriptomic and metabolomic analyses of a wax deficient citrus mutant exhibiting jasmonic acid-mediated defense against fungal pathogens. Hortic. Res. 2018, 5, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, H.; Liu, H.; Zhang, J.; Lyu, Y.; Li, Z.; He, Y.; Zhang, X.; Deng, X.; Brotman, Y.; Fernie, A.R.; et al. Lipidomic and transcriptomic analysis reveals reallocation of carbon flux from cuticular wax into plastid membrane lipids in a glossy “Newhall” navel orange mutant. Hortic. Res. 2020, 7, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Hao, H.; Liu, R.; Ma, Q.; Xu, J.; Chen, F.; Cheng, Y.; Deng, X. Comparative analysis of surface wax in mature fruits between Satsuma mandarin (Citrus unshiu) and ‘Newhall’ navel orange (Citrus sinensis) from the perspective of crystal morphology, chemical composition and key gene expression. Food Chem. 2014, 153, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.C.; Ma, Q.L.; Yang, L.; Hu, W.; Guo, W.F.; Wang, M.L.; Zhou, R.; Liu, Y. Comparative analysis of the cuticular waxes and related gene expression between ‘Newhall’ and ‘Ganqi 3′ navel orange during long-term cold storage. Plant Physiol. Biochem. 2021, 167, 1049–1060. [Google Scholar] [CrossRef]
- Maréchaux, I.; Bartlett, M.K.; Sack, L.; Baraloto, C.; Engel, J.; Luo, Y.L.; Su, Z.L.; Cui, X.L.; Lan, Q.Y. Water loss prevention plays a greater role than ROS scavenging in dehydration tolerance of Kalanchoe tubiflora epiphyllous buds. Isr. J. Plant Sci. 2015, 62, 1–7. [Google Scholar] [CrossRef]
- Guo, W.F.; Wu, Q.; Yang, L.; Hu, W.; Liu, D.C.; Liu, Y. Ectopic Expression of CsKCS6 from navel orange promotes the production of very-long-chain fatty acids (VLCFAs) and increases the abiotic stress tolerance of Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 564656. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, J.; Liu, R.; Liu, H.; Yang, H.; Zhu, Z.; Xu, R.; Wang, P.; Deng, X.; Xue, S.; et al. CsMYB96 confers water loss resistance in citrus fruit by simultaneous regulation of water transport and wax biosynthesis. J. Exp. Bot. 2022, 73, 953–966. [Google Scholar] [CrossRef]
- Yuan, G.; Chun, C.; Peng, L.; Huang, Z.; Huang, T.; Yang, H.; Fu, X.; Ling, L.; Cao, L.A. Study on the difference of ‘Newhall’ navel orange and its sport ‘Longhuihong’ navel orange. J. Fruit Sci. 2017, 34, 1117–1124. [Google Scholar] [CrossRef]
- Riederer, M.; Schreiber, L. Protecting against water loss: Analysis of the barrier properties of plant cuticles. J. Exp. Bot. 2001, 52, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Jetter, R.; Riederer, M. Localization of the transpiration barrier in the epi- and intracuticular waxes of eight plant species: Water transport resistances are associated with fatty acyl rather than alicyclic components. Plant Physiol. 2016, 170, 921–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchholz, A. Characterization of the diffusion of non-electrolytes across plant cuticles: Properties of the lipophilic pathway. J. Exp. Bot. 2006, 57, 2501–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, M.S.; Jiang, H.; Cao, Y.; Wang, Y.X.; You, C.X.; Li, Y.Y.; Hao, Y.J. MdCER2 conferred to wax accumulation and increased drought tolerance in plants. Plant Physiol. Biochem. 2020, 149, 277–285. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, X.; Wang, M.; Bi, Q.; Cui, Y.; Wang, L. Transcriptome and physiological analyses provide insights into the leaf epicuticular wax accumulation mechanism in yellowhorn. Hortic. Res. 2021, 8, 134. [Google Scholar] [CrossRef]
- Yamada, M.; Morishita, H.; Urano, K.; Shiozaki, N.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Yoshiba, Y. Effects of free proline accumulation in petunias under drought stress. J. Exp. Bot. 2005, 56, 1975–1981. [Google Scholar] [CrossRef] [Green Version]
- Bolouri-Moghaddam, M.R.; Roy, K.L.; Xiang, L.; Rolland, F.; Van den Ende, W. Sugar signaling and antioxidant network connections in plant cells. FEBS J. 2010, 277, 2022–2037. [Google Scholar] [CrossRef]
- Janero, D.R. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radic. Biol. Med. 1990, 9, 515–540. [Google Scholar] [CrossRef]
- Bajji, M.; Kinet, J.M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Mathobo, R.; Marais, D.; Steyn, J. The effect of drought stress on yield, leaf gaseous exchange and chlorophyll fluorescence of dry beans (Phaseolus vulgaris L.). Agric. Water Manag. 2017, 180, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, E.; Ekinci, M.; Turan, M. Impact of biochar in mitigating the negative effect of drought stress on cabbage seedlings. J. Soil Sci. Plant Nut. 2021, 21, 2297–2309. [Google Scholar] [CrossRef]
- Yuan, M.; Zhao, Y.Q.; Zhang, Z.W.; Chen, Y.E.; Ding, C.B.; Yuan, S. Light regulates transcription of chlorophyll biosynthetic genes during chloroplast biogenesis. Crit. Rev. Plant Sci. 2017, 36, 35–54. [Google Scholar] [CrossRef]
- Schuster, A.C.; Burghardt, M.; Riederer, M. The ecophysiology of leaf cuticular transpiration: Are cuticular water permeabilities adapted to ecological conditions? J. Exp. Bot. 2017, 68, 5271–5279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bögelein, R.; Hassdenteufel, M.; Thomas, F.M.; Werner, W. Comparison of leaf gas exchange and stable isotope signature of water-soluble compounds along canopy gradients of co-occurring Douglas-fir and European beech. Plant Cell Environ. 2012, 35, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Gago, J.; Douthe, C.; Florezsarasa, I.; Escalona, J.M.; Galmes, J.; Fernie, A.R.; Flexas, J.; Medrano, H. Opportunities for improving leaf water use efficiency under climate change conditions. Plant Sci. 2014, 226, 108–119. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Ubierna, N.; Winter, K.; Holtum, J.A.M.; Marshall, J.D.; Farquhar, G.D. Environmental and physiological determinants of carbon isotope is crimination in terrestrial plants. New Phytol. 2013, 200, 950–965. [Google Scholar] [CrossRef]
- Burow, G.B.; Franks, C.D.; Xin, Z. Genetic and physiological analysis of an irradiated bloomless mutant (epicuticular wax mutant) of Sorghum. Crop Sci. 2008, 48, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.H.; Li, K.T. Leaf color as a morpho-physiological index for screening heat tolerance and improved water use efficiency in rabbiteye blueberry (Vaccinium virgatum Aiton). Sci. Hortic. 2021, 278, 109864. [Google Scholar] [CrossRef]
- Reicosky, D.A.; Hanover, J.W. Physiological effects of surface waxes: I. Light reflectance for glaucous and nonglaucous Picea pungens. Plant Physiol. 1978, 62, 101–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamshiri, M.H.; Usha, K.; Singh, B. Carbon-Isotope discrimination, water use efficiency, net photosynthesis and transpiration in mycorrhizal kinnow (Citrus nobilis × C. deliciosa) plants. Acta Hortic. 2008, 773, 261–265. [Google Scholar] [CrossRef]
- Rowland, O.; Lee, R.; Franke, R.; Schreiber, L.; Kunst, L. The CER3 wax biosynthetic gene from Arabidopsis thaliana is allelic to WAX2/YRE/FLP1. FEBS Lett. 2007, 581, 3538–3544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, A.; Domergue, F.; Pascal, S.; Jetter, R.; Renne, C.; Faure, J.D.; Haslam, R.P.; Napier, J.A.; Lessire, R.; Joubès, J. Reconstitution of plant alkane biosynthesis in yeast demonstrates that Arabidopsis ECERIFERUM1 and ECERIFERUM3 are core components of a very-long-chain alkane synthesis complex. Plant Cell 2012, 24, 3106–3118. [Google Scholar] [CrossRef] [Green Version]
- Panikashvili, D.; Savaldi-Goldstein, S.; Mandel, T.; Yifhar, T.; Franke, R.B.; Hofer, R.; Schreiber, L.; Chory, J.; Aharoni, A. The Arabidopsis DESPERADO/AtWBC11 transporter is required for cutin and wax secretion. Plant Physiol. 2007, 145, 1345–1360. [Google Scholar] [CrossRef] [Green Version]
- Bird, D.; Beisson, F.; Brigham, A.; Shin, J.; Greer, S.; Jetter, R.; Kunst, L.; Wu, X.; Yephremov, A.; Samuels, L. Characterization of Arabidopsis ABCG11/WBC11, an ATP binding cassette (ABC) transporter that is required for cuticular lipid secretion. Plant J. 2007, 52, 485–498. [Google Scholar] [CrossRef]
- Buda, G.J.; Barnes, W.J.; Fich, E.A.; Park, S.; Yeats, T.H.; Zhao, L.; Domozych, D.S.; Rose, J.K.C. An ATP binding cassette transporter is required for cuticular wax deposition and desiccation tolerance in the moss Physcomitrella patens. Plant Cell 2013, 25, 4000–4013. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, D.; Liu, S.; Ma, X.; Dietrich, C.R.; Hu, H.C.; Zhang, G.; Liu, Z.; Zheng, J.; Wang, G.; et al. The Maize glossy13 gene, cloned via BSR-seq and seq-walking encodes a putative ABC transporter required for the normal accumulation of epicuticular waxes. PLoS ONE 2013, 8, e82333. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.N.T.; Lee, S.B.; Suh, M.C.; An, G.; Jung, K.H. OsABCG9 is an important ABC transporter of cuticular wax deposition in rice. Front. Plant Sci. 2018, 9, 960. [Google Scholar] [CrossRef]
- Piironen, V.; Lindsay, D.G.; Miettinen, T.A.; Toivo, J.; Lampi, A.M. Plant sterols: Biosynthesis, biological function and their importance to human nutrition. J. Sci. Food Agric. 2000, 80, 939–966. [Google Scholar] [CrossRef]
- Rowland, O.; Zheng, H.; Hepworth, S.R.; Lam, P.; Jetter, R.; Kunst, L. CER4 encodes an alcohol-forming fatty acyl-coenzyme A reductase involved in cuticular wax production in Arabidopsis. Plant Physiol. 2006, 14, 866–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, O.; Domergue, F. Plant fatty acyl reductases: Enzymes generating fatty alcohols for protective layers with potential for industrial applications. Plant Sci. 2012, 194, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Wang, Y.; Wu, H.Q.; Xu, J.; Li, T.T.; Hegebarth, D.; Jetter, R.; Chen, L.; Wang, Z.H. Three TaFAR genes function in the biosynthesis of primary alcohols and the response to abiotic stresses in Triticum aestivum. Sci. Rep. 2016, 6, 25008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; Arkel, G.V.; Pereira, A. The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Y.; Broeckling, C.D.; Sumner, L.W.; Wang, Z.Y. Heterologous expression of two Medicago truncatula putative ERF transcription factor genes, WXP1 and WXP2, in Arabidopsis led to increased leaf wax accumulation and improved drought tolerance, but differential response in freezing tolerance. Plant Mol. Biol. 2007, 64, 265–278. [Google Scholar] [CrossRef]
- Javelle, M.; Vernoud, V.; Depege-Fargeix, N.; Arnould, C.; Oursel, D.; Domergue, F.; Sarda, X.; Rogowsky, P.M. Over-expression of the epidermis-specific HD-ZIP IV transcription factor OCL1 in maize identifies target genes involved in lipid metabolism and cuticle biosynthesis. Plant Physiol. 2010, 154, 273–286. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Li, S.; He, S.; Wassmann, F.; Yu, C.; Qin, G.; Schreiber, L.; Qu, L.J.; Gu, H. CFL1, a WW domain protein, regulates cuticle development by modulating the function of HDG1, a class IV homeodomain transcription factor, in rice and Arabidopsis. Plant Cell 2011, 23, 3392–3411. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Bai, X.; Sun, X.; Zhu, D.; Liu, B.; Ji, W.; Cai, H.; Cao, L.; Wu, J.; Hu, M.; et al. Expression of wild soybean WRKY20 in Arabidopsis enhances drought tolerance and regulates ABA signaling. J. Exp. Bot. 2013, 64, 2155–2169. [Google Scholar] [CrossRef] [Green Version]
- Oshima, Y.; Mitsuda, N. The MIXTA-like Transcription factor MYB16 is a major regulator of cuticle formation in vegetative organs. Plant Signal. Behav. 2013, 8, e26826. [Google Scholar] [CrossRef]
- Go, Y.S.; Kim, H.; Kim, H.J.; Suh, M.C. Arabidopsis cuticular wax biosynthesis is negatively regulated by the DEWAX gene encoding an AP2/ERF-Type transcription factor. Plant Cell 2014, 26, 1666–1680. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.L.; Zhang, C.L.; Wang, G.L.; Wang, Y.X.; Qi, C.H.; Zhao, Q.; You, C.X.; Li, Y.Y.; Hao, Y.J. The R2R3 MYB transcription factor MdMYB30 modulates plant resistance against pathogens by regulating cuticular wax biosynthesis. BMC Plant Biol. 2019, 19, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castorina, G.; Domergue, F.; Chiara, M.; Zilio, M.; Persico, M.; Ricciardi, V.; Horner, D.S.; Consonni, G. Drought-responsive ZmFDL1/MYB94 regulates cuticle biosynthesis and cuticle-dependent leaf permeability. Plant Physiol. 2020, 184, 266–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xue, W.; Li, X.; Li, J.; Wu, J.; Xie, L.; Kawabata, S.; Li, Y.; Zhang, Y. EgMIXTA1, a MYB-type transcription factor, promotes cuticular wax formation in Eustoma grandiflorum Leaves. Front. Plant Sci. 2020, 11, 524947. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Mao, L.; Wei, X.; Xia, M.; Xu, C. MYB41, MYB107, and MYC2 promote ABA-mediated primary fatty alcohol accumulation via activation of AchnFAR in wound suberization in kiwifruit. Hortic. Res. 2020, 7, 86. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Xie, Q.; Yang, Q.; Sun, P.; Gao, S.; Li, H.; Zhang, J.; Wang, T.; Ye, Z.; Yang, C. Woolly, interacting with MYB transcription factor SlMYB31, regulates cuticular wax biosynthesis by modulating SlCER6 expression in tomato. Plant J. 2020, 103, 323–337. [Google Scholar] [CrossRef]
- Yang, S.U.; Kim, H.; Kim, R.J.; Kim, J.; Suh, M.C. AP2/DREB transcription factor RAP2.4 activates cuticular wax biosynthesis in Arabidopsis leaves under drought. Front. Plant Sci. 2020, 11, 895. [Google Scholar] [CrossRef]
- Zhang, C.L.; Wang, Y.X.; Hu, X.; Zhang, Y.L.; Wang, G.L.; You, C.X.; Li, Y.Y.; Hao, Y.J. An apple AP2/EREBP-type transcription factor, MdWRI4, enhances plant resistance to abiotic stress by increasing cuticular wax load. Environ. Exp. Bot. 2020, 180, 104206. [Google Scholar] [CrossRef]
- Mahmood, T.; Khalid, S.; Abdullah, M.; Ahmed, Z.; Shah, M.K.N.; Ghafoor, A.; Du, X. Insights into drought stress signaling in plants and the molecular genetic basis of cotton drought tolerance. Cells 2020, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhang, S. Mitogen-activated protein kinase cascades in signaling plant growth and development. Trends Plant Sci. 2015, 20, 56–64. [Google Scholar] [CrossRef]
- Virk, N.; Li, D.; Tian, L.; Huang, L.; Hong, Y.; Li, X.; Zhang, Y.; Liu, B.; Zhang, H.; Song, F. Arabidopsis Raf-like mitogen-activated protein kinase kinase kinase gene Raf43 is required for tolerance to multiple abiotic stresses. PLoS ONE 2015, 10, e0133975. [Google Scholar] [CrossRef]
- Kim, J.M.; Woo, D.H.; Kim, S.H.; Lee, S.Y.; Park, H.Y.; Seok, H.Y.; Chung, W.; Moon, Y.H. Arabidopsis MKKK20 is involved in osmotic stress response via regulation of MPK6 activity. Plant Cell Rep. 2012, 31, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cai, H.; Liu, P.; Wang, C.; Gao, H.; Wu, C.; Yan, K.; Zhang, S.; Huang, J.; Zheng, C. Arabidopsis MAPKKK18 positively regulates drought stress resistance via downstream MAPKK3. Biochem. Biophys. Res. Commun. 2017, 484, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, T.; Irie, K.; Hirayama, T.; Hayashida, N.; Yamaguchi-Shinozaki, K.; Matsumoto, K.; Shinozaki, K. A gene encoding a mitogen-activated protein kinase kinase kinase is induced simultaneously with genes for a mitogen-activated protein kinase and an S6 ribosomal protein kinase by touch, cold, and water stress in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1996, 93, 765–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, J.; Li, X.; Hicks, L.M.; Xiong, L. A Raf-like MAPKKK gene DSM1 mediates drought resistance through reactive oxygen species scavenging in rice. Plant Physiol. 2010, 152, 876–890. [Google Scholar] [CrossRef] [Green Version]
- Negi, N.P.; Shrivastava, D.C.; Sharma, V.; Sarin, N.B. Overexpression of CuZnSOD from Arachis hypogaea alleviates salinity and drought stress in tobacco. Plant Cell Rep. 2015, 34, 1109–1126. [Google Scholar] [CrossRef]
- Kim, Y.H.; Hong, J.K.; Kim, H.S.; Kwak, S.S. Overexpression of the sweetpotato peroxidase gene swpa4 enhances tolerance to methyl viologen-mediated oxidative stress and dehydration in Arabidopsis thaliana. J. Plant Biochem. Biotechnol. 2021, 30, 215–220. [Google Scholar] [CrossRef]
- Dossa, K.; Wei, X.; Li, D.; Fonceka, D.; Zhang, Y.; Wang, L.; Yu, J.; Boshou, L.; Diouf, D.; Cissé, N.; et al. Insight into the AP2/ERF transcription factor superfamily in sesame and expression profiling of DREB subfamily under drought stress. BMC Plant Biol. 2016, 16, 171. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, Y.; Wu, M.; Li, L.; Li, C.; Han, Z.; Yuan, J.; Chen, C.; Song, W.; Wang, C. Genome-wide identification of AP2/ERF transcription factors in cauliflower and expression profiling of the ERF family under salt and drought stresses. Front. Plant Sci. 2017, 8, 946. [Google Scholar] [CrossRef]
- Gao, Y.; Han, D.; Jia, W.; Ma, X.; Yang, Y.; Xu, Z. Molecular characterization and systematic analysis of NtAP2/ERF in tobacco and functional determination of NtRAV-4 under drought stress. Plant Physiol. Biochem. 2020, 156, 420–435. [Google Scholar] [CrossRef]
- Dong, C.; Xi, Y.; Chen, X.; Cheng, Z.M. Genome-wide identification of AP2/EREBP in Fragaria vesca and expression pattern analysis of the FvDREB subfamily under drought stress. BMC Plant Biol. 2021, 21, 295. [Google Scholar] [CrossRef]
- He, H.; Dong, Q.; Shao, Y.; Jiang, H.; Zhu, S.; Cheng, B.; Xiang, Y. Genome-wide survey and characterization of the WRKY gene family in Populus trichocarpa. Plant Cell Rep. 2012, 31, 1199–1217. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, J.; Wang, L.; Wang, S. Genome-wide investigation of WRKY transcription factors involved in terminal drought stress response in common bean. Front. Plant Sci. 2017, 8, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, N.; He, M.; Li, L.; Cui, S.; Hou, M.; Wang, L.; Mu, G.; Liu, L.; Yang, X.; Mondal, T.K. Identification and expression analysis of WRKY gene family under drought stress in peanut (Arachis hypogaea L.). PLoS ONE 2020, 15, e0231396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrmova, M.; Hussain, S.S. Plant transcription factors involved in drought and associated stresses. Int. J. Mol. Sci. 2021, 22, 5662. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.J.; Park, J.B.; Cho, Y.J.; Jung, C.; Seo, H.S.; Park, S.K.; Nahm, B.H.; Song, J.T. Overexpression of the ethylene-responsive factor gene BrERF4 from Brassica rapaincreases tolerance to salt and drought in Arabidopsis plants. Mol. Cells 2010, 30, 271–277. [Google Scholar] [CrossRef]
- Zhai, Y.; Shao, S.; Sha, W.; Zhao, Y.; Zhang, J.; Ren, W.; Zhang, C. Overexpression of soybean GmERF9 enhances the tolerance to drought and cold in the transgenic tobacco. Plant Cell Tiss. Organ Cult. 2017, 128, 607–618. [Google Scholar] [CrossRef]
- Butt, H.I.; Yang, Z.; Chen, E.; Zhao, G.; Gong, Q.; Yang, Z.; Zhang, X.; Li, F.; Zhang, H. Functional characterization of cotton GaMYB62L, a novel R2R3 TF in transgenic Arabidopsis. PLoS ONE 2017, 12, e0170578. [Google Scholar] [CrossRef]
- Nguyen, X.C.; Kim, S.H.; Hussain, S.; An, J.; Yoo, Y.; Han, H.J.; Yoo, J.S.; Lim, C.O.; Yun, D.J.; Chung, W.S. A positive transcription factor in osmotic stress tolerance, ZAT10, is regulated by MAP kinases in Arabidopsis. J. Plant Biol. 2016, 59, 55–61. [Google Scholar] [CrossRef]
- Hong, Y.; Zhang, H.; Huang, L.; Li, D.; Song, F. Overexpression of a stress-responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice. Front. Plant Sci. 2016, 7, 4. [Google Scholar] [CrossRef]
- Lin, J.; Dang, F.; Chen, Y.; Guan, D.; He, S. CaWRKY27 negatively regulates salt and osmotic stress responses in pepper. Plant Physiol. Biochem. 2019, 145, 43–51. [Google Scholar] [CrossRef]
- Xu, X.; Wang, C.; Ma, X.; Pan, Y.; Ying, Q.; Song, H.; Wang, H. Overexpression of DnWRKY29 in tobacco impaired plants tolerance to salt and drought stresses. Russ. J. Plant Physiol. 2015, 62, 262–269. [Google Scholar] [CrossRef]
- Huang, X.S.; Liu, J.H.; Chen, X.J. Overexpression of PtrABF gene, a bZIP transcription factor isolated from Poncirus trifoliata, enhances dehydration and drought tolerance in tobacco via scavenging ROS and modulating expression of stress-responsive genes. BMC Plant Biol. 2010, 10, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verlinden, M.S.; Fichot, R.; Broeckx, L.S.; Vanholme, B.; Boerjan, W.; Ceulemans, R. Carbon isotope compositions (δ13C) of leaf, wood and holocellulose differ among genotypes of poplar and between previous land uses in a short-rotation biomass plantation. Plant Cell Environ. 2015, 38, 144–156. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | log2 (MT/WT) | Function Definition |
|---|---|---|---|
| CsCER3-LIKE | Cs4g02580 | 2.8143 | Very-long-chain aldehyde decarbonylase |
| CsFAR1-LIKE | Cs8g15290 | −2.3009 | Fatty acyl-CoA reductase 1 |
| CsFAR3-LIKE | Cs5g15350 | −1.3315 | Fatty acyl-CoA reductase 3 |
| CsFAR4-LIKE | Cs5g15345 | −2.0485 | Fatty acyl-CoA reductase 3 |
| CsNSDHL-LIKE | BGI_novel_G000534 | −4.7498 | Sterol-4alpha-carboxylate 3-dehydrogenase |
| CsChDI-LIKE | orange1.1t04155 | −1.1913 | Cholestenol Delta-isomerase |
| CsABCG11-LIKE | orange1.1t01993 | 1.4780 | ATP-binding cassette, subfamily G, member 11 |
| CsABCG21-LIKE | Cs7g12620 | 1.2926 | ATP-binding cassette, subfamily G, member 21 |
| CsMEKK1-LIKE | Cs5g22020 | 1.2434 | MAP kinase kinase kinase |
| CsSOD1-LIKE | orange1.1t05755 | 2.2530 | Superoxide dismutase 1, Cu-Zn family |
| CsPRX5-LIKE | orange1.1t01747 | 1.8422 | Peroxidase 5 |
| CsPRX10-LIKE | Cs5g34200 | 2.8727 | Peroxidase 10 |
| CsPRX24-LIKE | Cs9g05140 | 1.1134 | Peroxidase 24 |
| CsPRX25-LIKE | Cs4g17860 | 1.4794 | Peroxidase 25 |
| CsERF4-LIKE | Cs1g07950 | 1.0323 | Ethylene-responsive transcription factor 4 |
| CsERF9-LIKE | Cs2g23660 | 1.5729 | Ethylene-responsive transcription factor 9 |
| CsMYB62-LIKE | Cs7g26930 | 1.3835 | Transcription factor MYB62 |
| CsZAT10-LIKE1 | Cs3g15900 | 1.1106 | Zinc finger protein ZAT10 |
| CsZAT10-LIKE2 | Cs8g04280 | 1.0356 | Zinc finger protein ZAT10 |
| CsNAC22-LIKE | Cs5g29650 | 2.0431 | NAC domain-containing protein 22 |
| CsWRKY27-LIKE | Cs9g19070 | −1.5064 | WRKY transcription factor 27 |
| CsWRKY29-LIKE | Cs5g03010 | −4.1799 | WRKY transcription factor 29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, B.; Wan, S.; Ma, Q.; Yang, L.; Hu, W.; Kuang, L.; Xie, J.; Liu, D.; Liu, Y. Transcriptome and Physiological Analyses of a Navel Orange Mutant with Improved Drought Tolerance and Water Use Efficiency Caused by Increases of Cuticular Wax Accumulation and ROS Scavenging Capacity. Int. J. Mol. Sci. 2022, 23, 5660. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105660
Liang B, Wan S, Ma Q, Yang L, Hu W, Kuang L, Xie J, Liu D, Liu Y. Transcriptome and Physiological Analyses of a Navel Orange Mutant with Improved Drought Tolerance and Water Use Efficiency Caused by Increases of Cuticular Wax Accumulation and ROS Scavenging Capacity. International Journal of Molecular Sciences. 2022; 23(10):5660. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105660
Chicago/Turabian StyleLiang, Beibei, Shiguo Wan, Qingling Ma, Li Yang, Wei Hu, Liuqing Kuang, Jingheng Xie, Dechun Liu, and Yong Liu. 2022. "Transcriptome and Physiological Analyses of a Navel Orange Mutant with Improved Drought Tolerance and Water Use Efficiency Caused by Increases of Cuticular Wax Accumulation and ROS Scavenging Capacity" International Journal of Molecular Sciences 23, no. 10: 5660. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105660