
Pulsed Electromagnetic Field (PEMF) Treatment Reduces Lipopolysaccharide-Induced Septic Shock in Mice

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
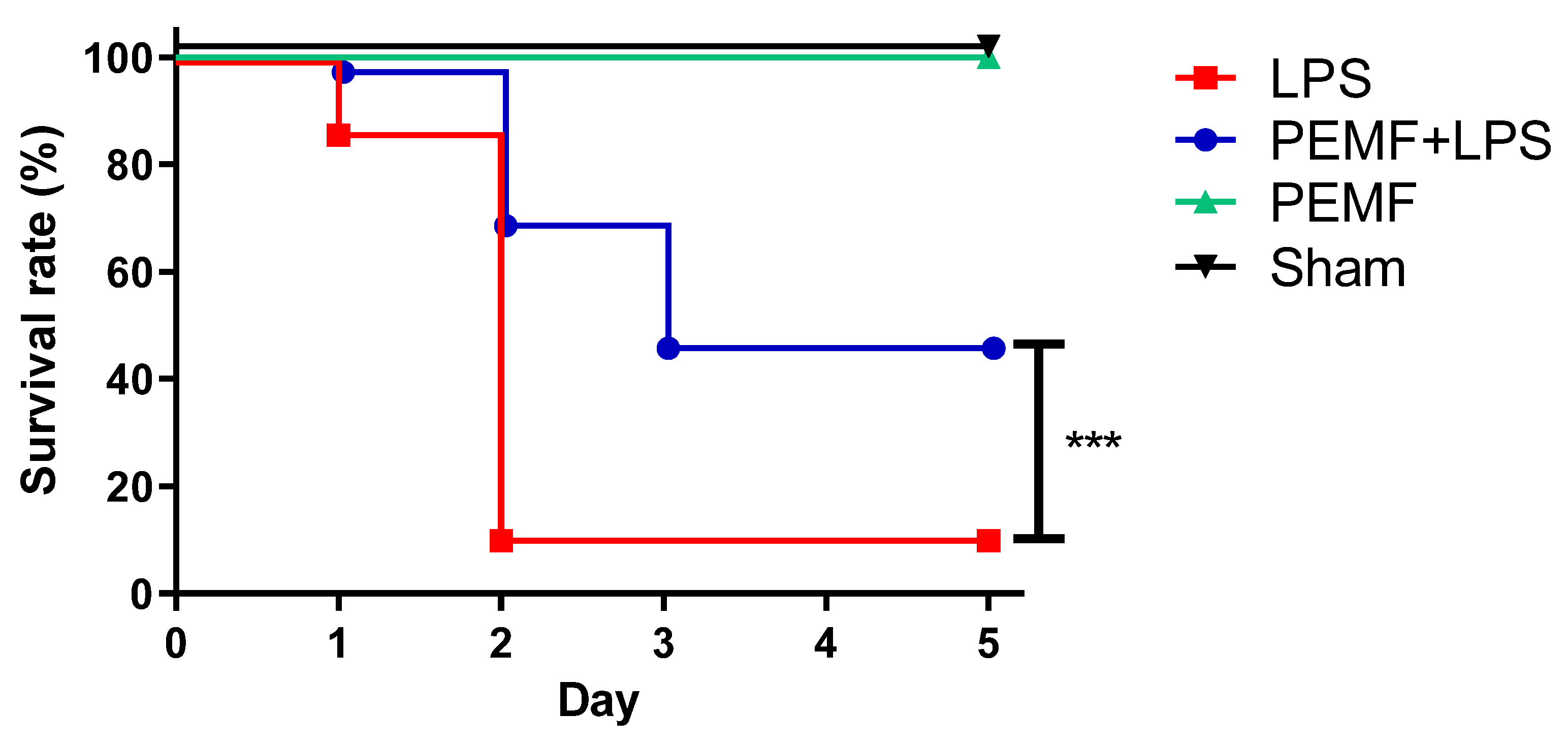
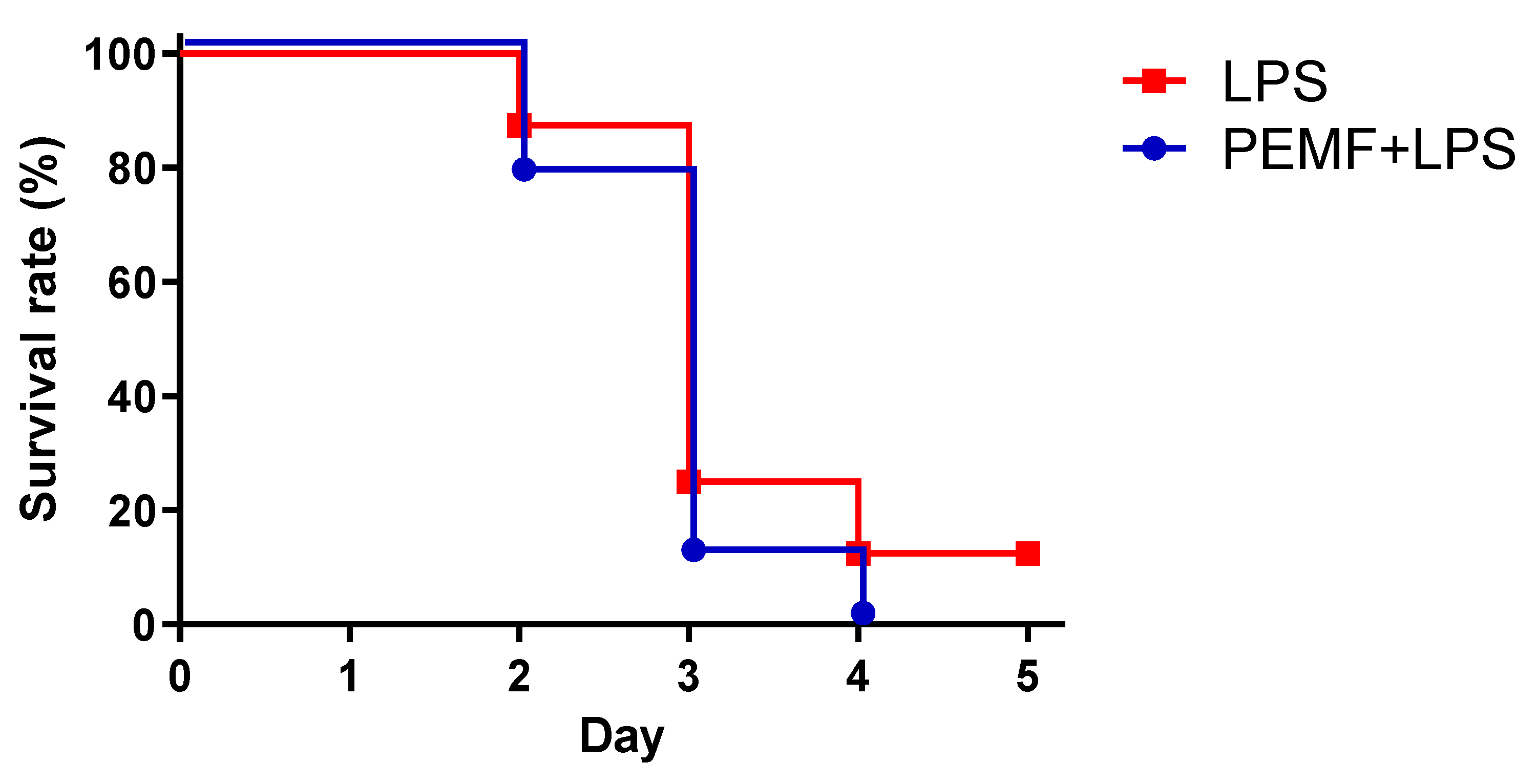
2.1. PEMF Treatment Decreased Septic Shock-Induced Mortality in Mice
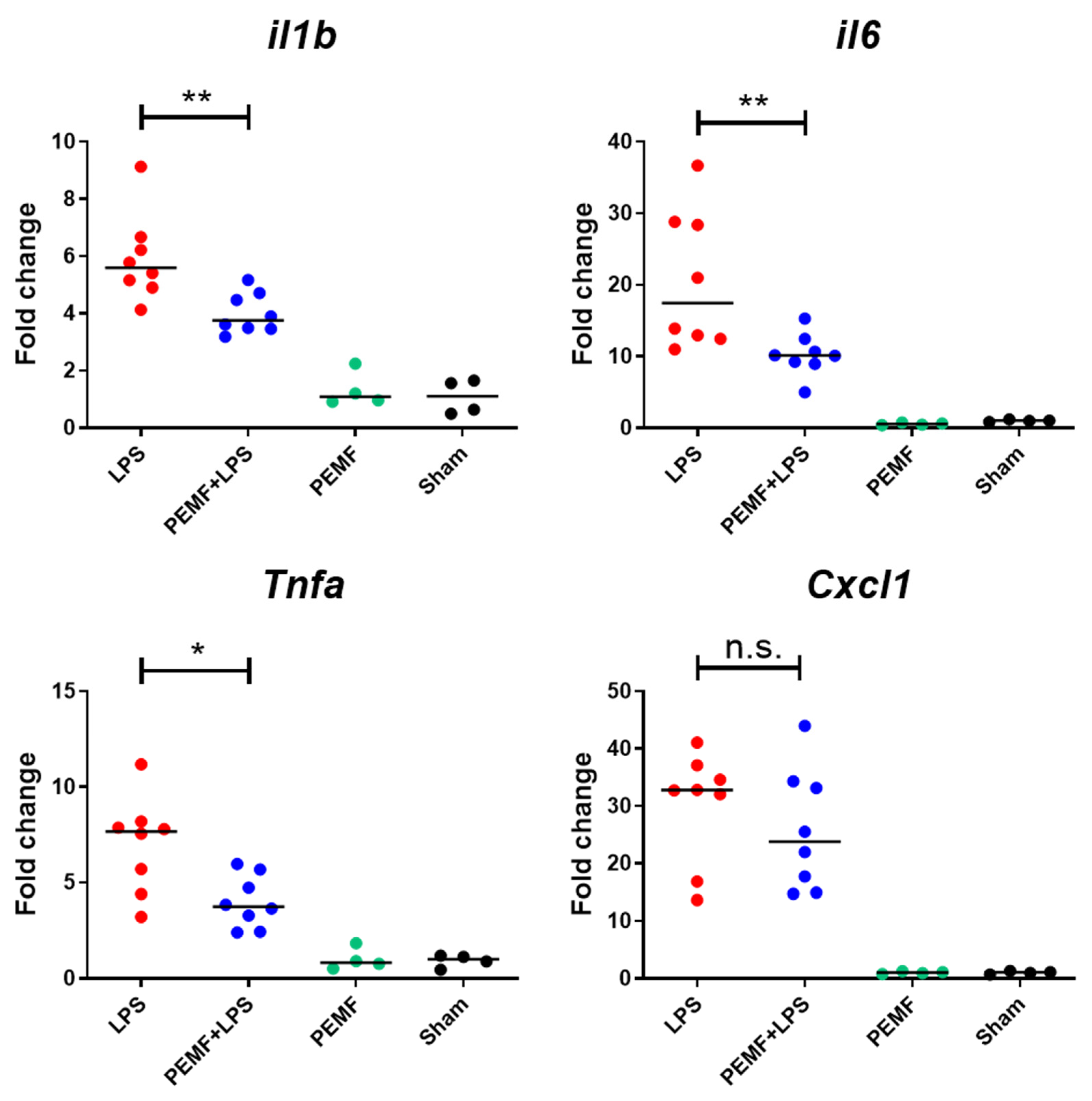
2.2. PEMF Treatment Attenuated Expression of Inflammatory Cytokines in Liver of Septic Shock-Induced Mice
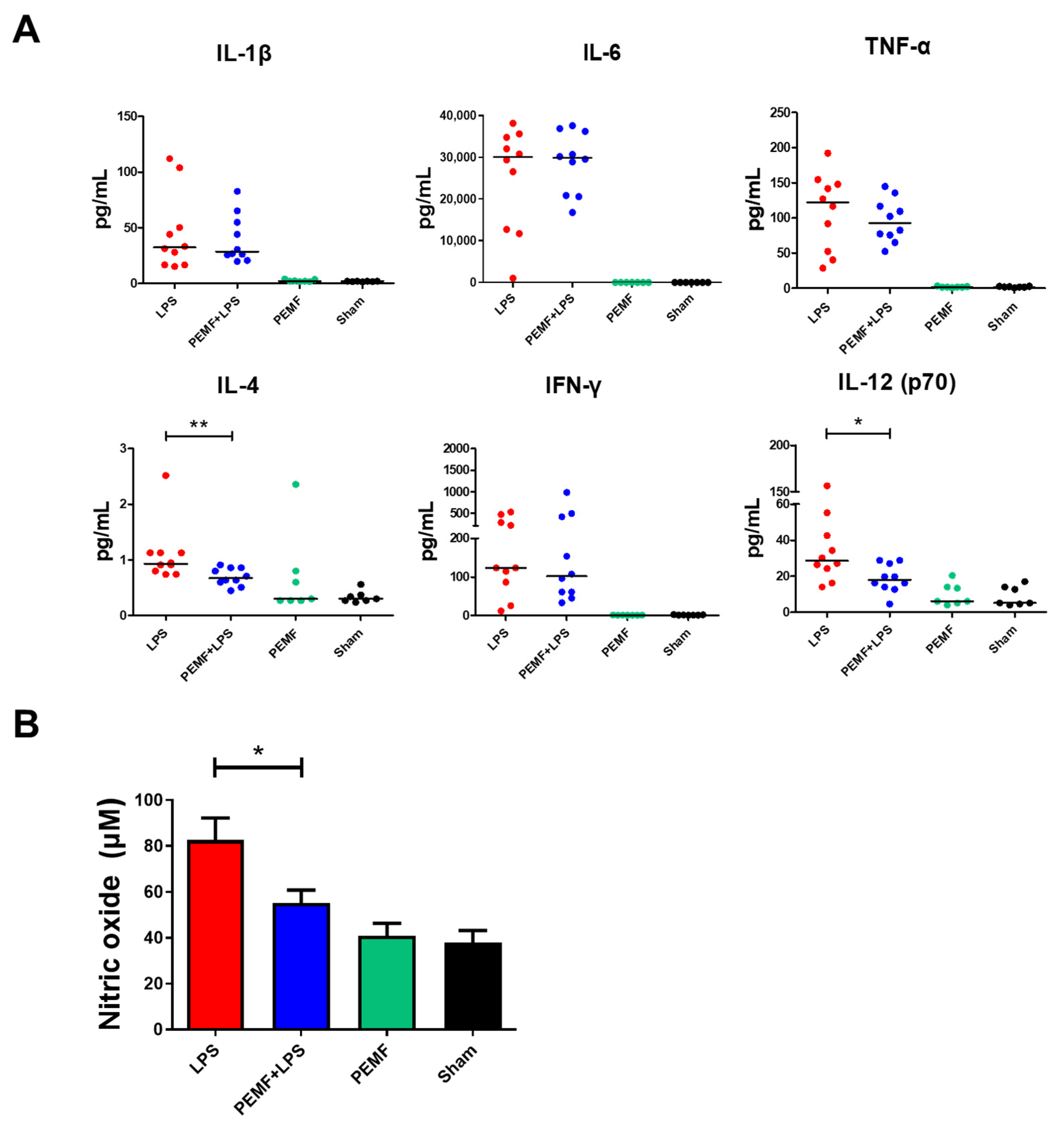
2.3. PEMF Attenuated Serum IL-12 and Nitric Oxide Levels in Septic Shock-Induced Mice
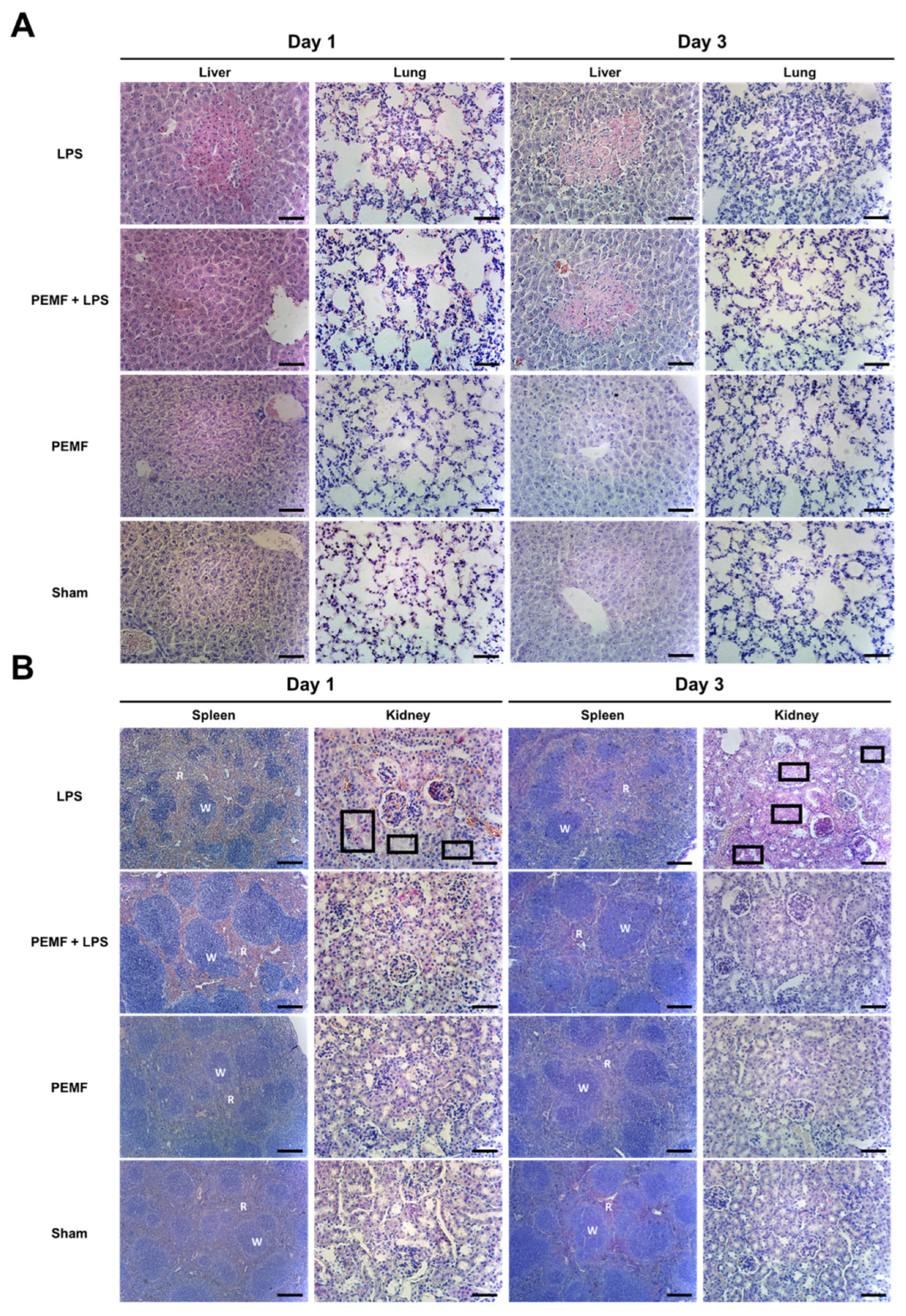
2.4. PEMF Treatment Attenuated Multiple Organ Damage in Septic Shock-Induced Mice
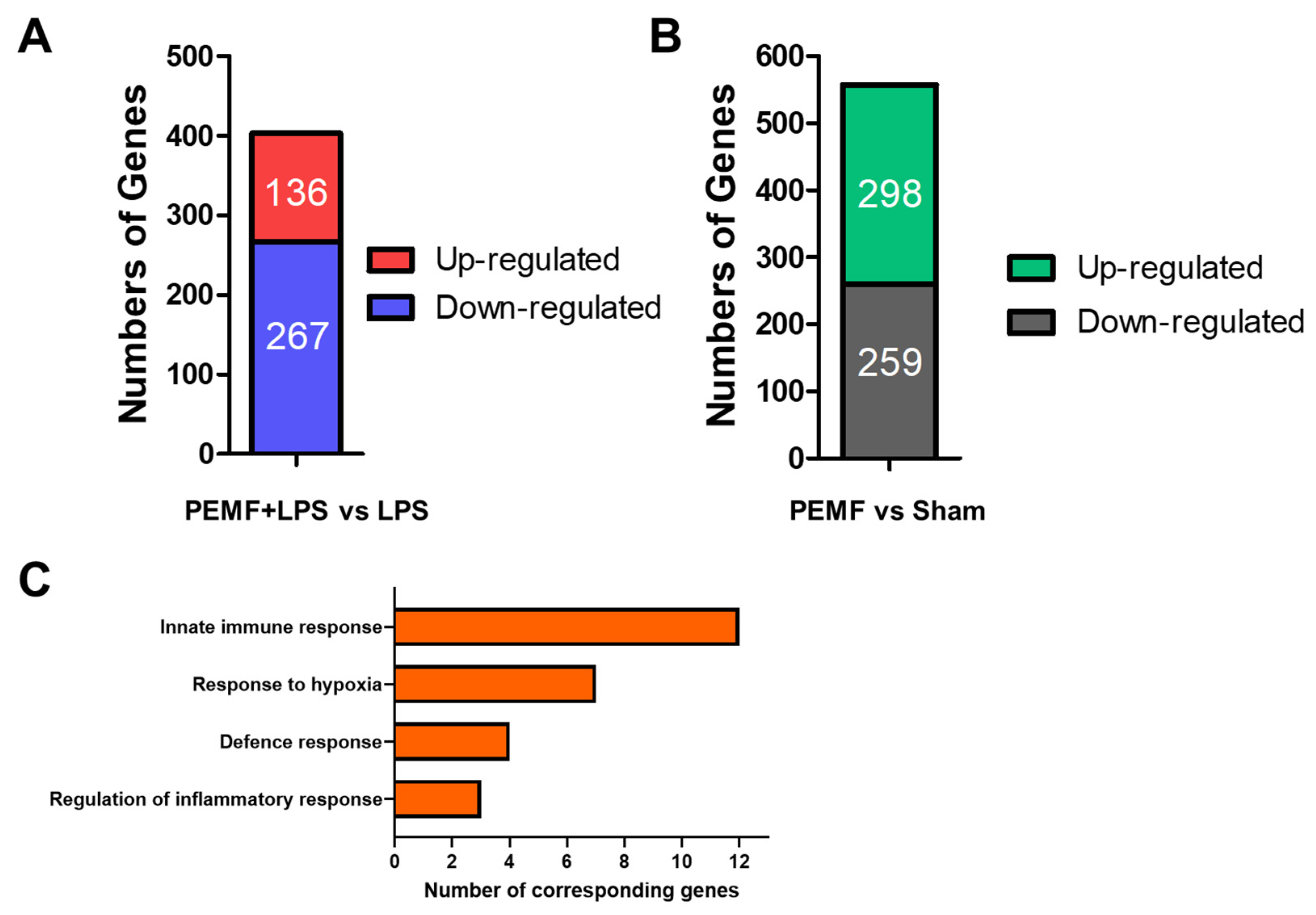
2.5. PEMF Treatment Induces Multiple Gene Expression Changes in Mice
2.6. PEMF Treatment on B Cell Deficient Mice Showed No Protective Effect in Septic Shock-Induced Mice
3. Discussion
4. Materials and Methods
4.1. Animals
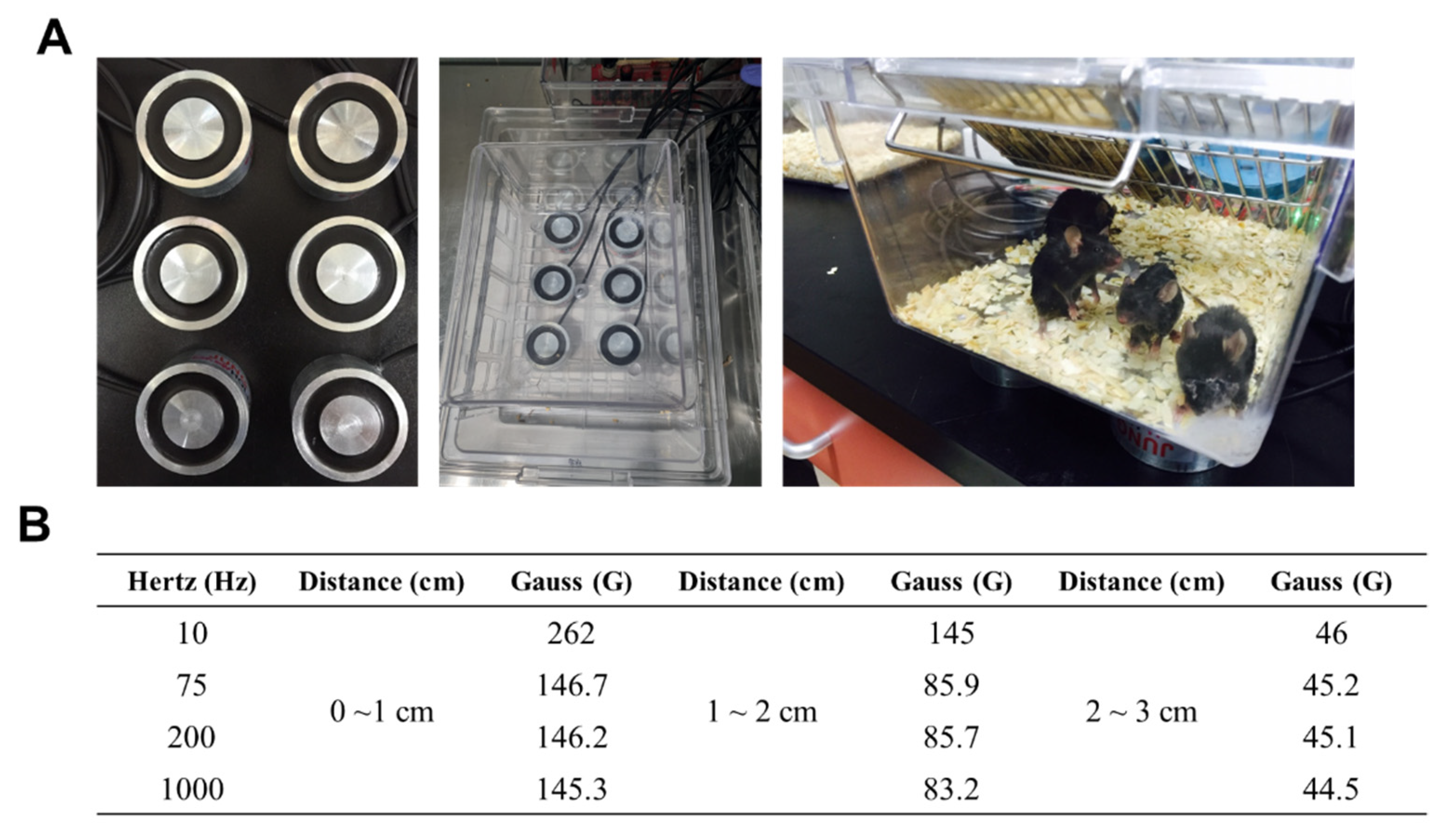
4.2. LPS-Induced Septic Shock Experiment and Pulsed-Electromagnetic Field (PEMF) Treatment
4.3. Histology
4.4. Quantitative Reverse-Transcriptase Polymerase Chain Reaction (qRT-PCR)
4.5. Determination of Serum Nitric Oxide and Serum Cytokines
4.6. Next-Generation Sequencing and Gene Ontology (GO) Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Font, M.D.; Thyagarajan, B.; Khanna, A.K. Sepsis and Septic Shock-Basics of diagnosis, pathophysiology and clinical decision making. Med. Clin. N. Am. 2020, 104, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.S.; Coopersmith, C.M.; McDunn, J.E.; Ferguson, T.A. The sepsis seesaw: Tilting toward immunosuppression. Nat. Med. 2009, 15, 496–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potschke, C.; Kessler, W.; Maier, S.; Heidecke, C.D.; Broker, B.M. Experimental sepsis impairs humoral memory in mice. PLoS ONE 2013, 8, e81752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosmann, M.; Ward, P.A. The inflammatory response in sepsis. Trends Immunol. 2013, 34, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulte, W.; Bernhagen, J.; Bucala, R. Cytokines in sepsis: Potent immunoregulators and potential therapeutic targets--an updated view. Mediat. Inflamm. 2013, 2013, 165974. [Google Scholar] [CrossRef] [PubMed]
- Chousterman, B.G.; Swirski, F.K.; Weber, G.F. Cytokine storm and sepsis disease pathogenesis. Semin. Immunopathol. 2017, 39, 517–528. [Google Scholar] [CrossRef]
- Lambden, S. Bench to bedside review: Therapeutic modulation of nitric oxide in sepsis-an update. Intensiv. Care Med. Exp. 2019, 7, 64. [Google Scholar] [CrossRef] [Green Version]
- Gotur, D.B. Sepsis in a Panorama: What the Cardiovascular Physician Should Know. Methodist Debakey Cardiovasc. J. 2018, 14, 89–100. [Google Scholar] [CrossRef]
- Gotts, J.E.; Matthay, M.A. Sepsis: Pathophysiology and clinical management. BMJ 2016, 353, i1585. [Google Scholar] [CrossRef] [Green Version]
- Lelubre, C.; Vincent, J.L. Mechanisms and treatment of organ failure in sepsis. Nat. Rev. Nephrol. 2018, 14, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Mayr, F.B.; Yende, S.; Angus, D.C. Epidemiology of severe sepsis. Virulence 2014, 5, 4–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, L.; van Meegern, A.; Doemming, S.; Schuerholz, T. Antimicrobial Peptides in Human Sepsis. Front. Immunol. 2015, 6, 404. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Roberts, D.; Wood, K.E.; Light, B.; Parrillo, J.E.; Sharma, S.; Suppes, R.; Feinstein, D.; Zanotti, S.; Taiberg, L.; et al. Duration of hypotension before initiation of effective antimicrobial therapy is the critical determinant of survival in human septic shock. Crit. Care Med. 2006, 34, 1589–1596. [Google Scholar] [CrossRef]
- Epstein, L.; Dantes, R.; Magill, S.; Fiore, A. Varying Estimates of Sepsis Mortality Using Death Certificates and Administrative Codes--United States, 1999–2014. MMWR. Morb. Mortal. Wkly. Rep. 2016, 65, 342–345. [Google Scholar] [CrossRef] [Green Version]
- Napolitano, L.M. Sepsis 2018: Definitions and Guideline Changes. Surg. Infect. 2018, 19, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Perman, S.M.; Goyal, M.; Gaieski, D.F. Initial emergency department diagnosis and management of adult patients with severe sepsis and septic shock. Scand. J. Trauma Resusc. Emerg. Med. 2012, 20, 41. [Google Scholar] [CrossRef] [Green Version]
- Gavelli, F.; Castello, L.M.; Avanzi, G.C. Management of sepsis and septic shock in the emergency department. Intern. Emerg. Med. 2021, 16, 1649–1661. [Google Scholar] [CrossRef]
- Perner, A.; Gordon, A.C.; De Backer, D.; Dimopoulos, G.; Russell, J.A.; Lipman, J.; Jensen, J.U.; Myburgh, J.; Singer, M.; Bellomo, R.; et al. Sepsis: Frontiers in diagnosis, resuscitation and antibiotic therapy. Intensiv. Care Med. 2016, 42, 1958–1969. [Google Scholar] [CrossRef]
- Shahrami, B.; Sharif, M.; Sefidani Forough, A.; Najmeddin, F.; Arabzadeh, A.A.; Mojtahedzadeh, M. Antibiotic therapy in sepsis: No next time for a second chance! J. Clin. Pharm. Ther. 2021, 46, 872–876. [Google Scholar] [CrossRef]
- Asner, S.A.; Desgranges, F.; Schrijver, I.T.; Calandra, T. Impact of the timeliness of antibiotic therapy on the outcome of patients with sepsis and septic shock. J. Infect. 2021, 82, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Waldorff, E.I.; Zhang, N.; Ryaby, J.T. Pulsed electromagnetic field applications: A corporate perspective. J. Orthop. Translat. 2017, 9, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Pieber, K.; Schuhfried, O.; Fialka-Moser, V. Pulsed electromagnetic fields (PEMF)--results in evidence based medicine. Wien. Med. Wochenschr. 2007, 157, 34–36. [Google Scholar] [CrossRef] [PubMed]
- Novickij, V.; Grainys, A.; Lastauskiene, E.; Kananaviciute, R.; Pamedytyte, D.; Kalediene, L.; Novickij, J.; Miklavcic, D. Pulsed Electromagnetic Field Assisted in vitro Electroporation: A Pilot Study. Sci. Rep. 2016, 6, 33537. [Google Scholar] [CrossRef] [Green Version]
- Kubat, N.J.; Moffett, J.; Fray, L.M. Effect of pulsed electromagnetic field treatment on programmed resolution of inflammation pathway markers in human cells in culture. J. Inflamm. Res. 2015, 8, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Gessi, S.; Merighi, S.; Bencivenni, S.; Battistello, E.; Vincenzi, F.; Setti, S.; Cadossi, M.; Borea, P.A.; Cadossi, R.; Varani, K. Pulsed electromagnetic field and relief of hypoxia-induced neuronal cell death: The signaling pathway. J. Cell. Physiol. 2019, 234, 15089–15097. [Google Scholar] [CrossRef]
- Vadala, M.; Morales-Medina, J.C.; Vallelunga, A.; Palmieri, B.; Laurino, C.; Iannitti, T. Mechanisms and therapeutic effectiveness of pulsed electromagnetic field therapy in oncology. Cancer Med. 2016, 5, 3128–3139. [Google Scholar] [CrossRef]
- Markov, M.S. Expanding use of pulsed electromagnetic field therapies. Electromagn. Biol. Med. 2007, 26, 257–274. [Google Scholar] [CrossRef]
- Martiny, K.; Lunde, M.; Bech, P. Transcranial low voltage pulsed electromagnetic fields in patients with treatment-resistant depression. Biol. Psychiatry 2010, 68, 163–169. [Google Scholar] [CrossRef]
- Strauch, B.; Herman, C.; Dabb, R.; Ignarro, L.J.; Pilla, A.A. Evidence-based use of pulsed electromagnetic field therapy in clinical plastic surgery. Aesthetic Surg. J. 2009, 29, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahim, A.; Hassanein, H.R.; Dahaba, M. Effect of pulsed electromagnetic field on healing of mandibular fracture: A preliminary clinical study. J. Oral Maxillofac. Surg. 2011, 69, 1708–1717. [Google Scholar] [CrossRef] [PubMed]
- Stocchero, M.; Gobbato, L.; De Biagi, M.; Bressan, E.; Sivolella, S. Pulsed electromagnetic fields for postoperative pain: A randomized controlled clinical trial in patients undergoing mandibular third molar extraction. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2015, 119, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, F.; Botarelli, E.; Mele, G.; Polo, L.; Zoncu, D.; Renati, P.; Sgarlata, C.; Rollone, M.; Ricevuti, G.; Maurizi, N.; et al. Effectiveness of an Innovative Pulsed Electromagnetic Fields Stimulation in Healing of Untreatable Skin Ulcers in the Frail Elderly: Two Case Reports. Case Rep. Dermatol. Med. 2015, 2015, 576580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadossi, R.; Massari, L.; Racine-Avila, J.; Aaron, R.K. Pulsed Electromagnetic Field Stimulation of Bone Healing and Joint Preservation: Cellular Mechanisms of Skeletal Response. J. Am. Acad. Orthop. Surg. Glob. Res. Rev. 2020, 4, e1900155. [Google Scholar] [CrossRef]
- Ross, C.L.; Ang, D.C.; Almeida-Porada, G. Targeting Mesenchymal Stromal Cells/Pericytes (MSCs) With Pulsed Electromagnetic Field (PEMF) Has the Potential to Treat Rheumatoid Arthritis. Front. Immunol. 2019, 10, 266. [Google Scholar] [CrossRef]
- Yang, X.; Guo, H.; Ye, W.; Yang, L.; He, C. Pulsed Electromagnetic Field Attenuates Osteoarthritis Progression in a Murine Destabilization-Induced Model through Inhibition of TNF-α and IL-6 Signaling. Cartilage 2021, 13, 1665s–1675s. [Google Scholar] [CrossRef]
- Selvam, R.; Ganesan, K.; Narayana Raju, K.V.; Gangadharan, A.C.; Manohar, B.M.; Puvanakrishnan, R. Low frequency and low intensity pulsed electromagnetic field exerts its antiinflammatory effect through restoration of plasma membrane calcium ATPase activity. Life Sci. 2007, 80, 2403–2410. [Google Scholar] [CrossRef]
- Kingsley, S.M.; Bhat, B.V. Differential Paradigms in Animal Models of Sepsis. Curr. Infect. Dis. Rep. 2016, 18, 26. [Google Scholar] [CrossRef]
- Fink, M.P. Animal models of sepsis. Virulence 2014, 5, 143–153. [Google Scholar] [CrossRef]
- Zanotti-Cavazzoni, S.L.; Goldfarb, R.D. Animal models of sepsis. Crit. Care Clin. 2009, 25, 703–719. [Google Scholar] [CrossRef]
- Lewis, A.J.; Seymour, C.W.; Rosengart, M.R. Current Murine Models of Sepsis. Surg. Infect. (Larchmt) 2016, 17, 385–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, M.; Scott, M.J.; Loughran, P.; Gibson, G.; Sodhi, C.; Watkins, S.; Hackam, D.; Billiar, T.R. Lipopolysaccharide clearance, bacterial clearance, and systemic inflammatory responses are regulated by cell type-specific functions of TLR4 during sepsis. J. Immunol. 2013, 190, 5152–5160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickson, K.; Lehmann, C. Inflammatory Response to Different Toxins in Experimental Sepsis Models. Int. J. Mol. Sci. 2019, 20, 4341. [Google Scholar] [CrossRef] [Green Version]
- Reyes, M.; Filbin, M.R.; Bhattacharyya, R.P.; Billman, K.; Eisenhaure, T.; Hung, D.T.; Levy, B.D.; Baron, R.M.; Blainey, P.C.; Goldberg, M.B.; et al. An immune-cell signature of bacterial sepsis. Nat. Med. 2020, 26, 333–340. [Google Scholar] [CrossRef]
- Arfaras-Melainis, A.; Polyzogopoulou, E.; Triposkiadis, F.; Xanthopoulos, A.; Ikonomidis, I.; Mebazaa, A.; Parissis, J. Heart failure and sepsis: Practical recommendations for the optimal management. Hear. Fail. Rev. 2020, 25, 183–194. [Google Scholar] [CrossRef]
- Vanderhaeghen, T.; Vandewalle, J.; Libert, C. Hypoxia-inducible factors in metabolic reprogramming during sepsis. FEBS J. 2020, 287, 1478–1495. [Google Scholar] [CrossRef] [Green Version]
- Cecconi, M.; Evans, L.; Levy, M.; Rhodes, A. Sepsis and septic shock. Lancet 2018, 392, 75–87. [Google Scholar] [CrossRef]
- Chimenz, R.; Chirico, V.; Basile, P.; Carcione, A.; Conti, G.; Monardo, P.; Lacquaniti, A. HMGB-1 and TGFβ-1 highlight immuno-inflammatory and fibrotic processes before proteinuria onset in pediatric patients with Alport syndrome. J. Nephrol. 2021, 34, 1915–1924. [Google Scholar] [CrossRef]
- Chimenz, R.; Chirico, V.; Cuppari, C.; Ceravolo, G.; Concolino, D.; Monardo, P.; Lacquaniti, A. Fabry disease and kidney involvement: Starting from childhood to understand the future. Pediatr. Nephrol. 2022, 37, 95–103. [Google Scholar] [CrossRef]
- Cavaillon, J.M. Exotoxins and endotoxins: Inducers of inflammatory cytokines. Toxicon 2018, 149, 45–53. [Google Scholar] [CrossRef]
- Zhao, H.Q.; Li, W.M.; Lu, Z.Q.; Sheng, Z.Y.; Yao, Y.M. The growing spectrum of anti-inflammatory interleukins and their potential roles in the development of sepsis. J. Interf. Cytokine Res. 2015, 35, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V. Toll-like receptors in sepsis-associated cytokine storm and their endogenous negative regulators as future immunomodulatory targets. Int. Immunopharmacol. 2020, 89, 107087. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Ochoa, I.; Gomez-Ochoa, P.; Gomez-Casal, F.; Cativiela, E.; Larrad-Mur, L. Pulsed electromagnetic fields decrease proinflammatory cytokine secretion (IL-1beta and TNF-alpha) on human fibroblast-like cell culture. Rheumatol. Int. 2011, 31, 1283–1289. [Google Scholar] [CrossRef] [PubMed]
- de Girolamo, L.; Stanco, D.; Galliera, E.; Vigano, M.; Colombini, A.; Setti, S.; Vianello, E.; Corsi Romanelli, M.M.; Sansone, V. Low frequency pulsed electromagnetic field affects proliferation, tissue-specific gene expression, and cytokines release of human tendon cells. Cell Biochem. Biophys. 2013, 66, 697–708. [Google Scholar] [CrossRef]
- Cronkite, D.A.; Strutt, T.M. The Regulation of Inflammation by Innate and Adaptive Lymphocytes. J. Immunol. Res. 2018, 2018, 1467538. [Google Scholar] [CrossRef]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH Oxidase Activation and Bacterial Resistance. Front. Cell Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef]
- Patruno, A.; Amerio, P.; Pesce, M.; Vianale, G.; Di Luzio, S.; Tulli, A.; Franceschelli, S.; Grilli, A.; Muraro, R.; Reale, M. Extremely low frequency electromagnetic fields modulate expression of inducible nitric oxide synthase, endothelial nitric oxide synthase and cyclooxygenase-2 in the human keratinocyte cell line HaCat: Potential therapeutic effects in wound healing. Br. J. Dermatol. 2010, 162, 258–266. [Google Scholar] [CrossRef]
- Garrean, S.; Gao, X.P.; Brovkovych, V.; Shimizu, J.; Zhao, Y.Y.; Vogel, S.M.; Malik, A.B. Caveolin-1 regulates NF-kappaB activation and lung inflammatory response to sepsis induced by lipopolysaccharide. J. Immunol. 2006, 177, 4853–4860. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Fan, G.; Liu, Y.; Liu, L.; Zhang, T.; Liu, P.; Tu, Q.; Zhang, X.; Luo, S.; Yao, L.; et al. The transcription factor KLF14 regulates macrophage glycolysis and immune function by inhibiting HK2 in sepsis. Cell Mol. Immunol. 2022, 19, 504–515. [Google Scholar] [CrossRef]
- Zhao, D.M.; Ding, R.Y.; Mao, Y.R.; Wang, L.; Zhang, Z.D.; Ma, X.C. Heparin Rescues Sepsis-Associated Acute Lung Injury and Lethality Through the Suppression of Inflammatory Responses. Inflammation 2012, 35, 1825–1832. [Google Scholar] [CrossRef]
- Wu, Y.; Lu, J.; Antony, S.; Juhasz, A.; Liu, H.; Jiang, G.; Meitzler, J.L.; Hollingshead, M.; Haines, D.C.; Butcher, D.; et al. Activation of TLR4 is required for the synergistic induction of dual oxidase 2 and dual oxidase A2 by IFN-γ and lipopolysaccharide in human pancreatic cancer cell lines. J. Immunol. 2013, 190, 1859–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipinski, S.; Till, A.; Sina, C.; Arlt, A.; Grasberger, H.; Schreiber, S.; Rosenstiel, P. DUOX2-derived reactive oxygen species are effectors of NOD2-mediated antibacterial responses. J. Cell Sci. 2009, 122, 3522–3530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfieri, A.; Malito, E.; Orru, R.; Fraaije, M.W.; Mattevi, A. Revealing the moonlighting role of NADP in the structure of a flavin-containing monooxygenase. Proc. Natl. Acad. Sci. USA 2008, 105, 6572–6577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Chaluvadi, M.R.; Reddy, R.; Motika, M.S.; Richardson, T.A.; Cashman, J.R.; Morgan, E.T. Hepatic flavin-containing monooxygenase gene regulation in different mouse inflammation models. Drug Metab. Dispos. 2009, 37, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Kelly-Scumpia, K.M.; Scumpia, P.O.; Weinstein, J.S.; Delano, M.J.; Cuenca, A.G.; Nacionales, D.C.; Wynn, J.L.; Lee, P.Y.; Kumagai, Y.; Efron, P.A.; et al. B cells enhance early innate immune responses during bacterial sepsis. J. Exp. Med. 2011, 208, 1673–1682. [Google Scholar] [CrossRef]
- Aziz, M.; Holodick, N.E.; Rothstein, T.L.; Wang, P. B-1a Cells Protect Mice from Sepsis: Critical Role of CREB. J. Immunol. 2017, 199, 750–760. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Gene Name | Description | Log2FC | p Value |
|---|---|---|---|---|
| Down-regulated | Cav1 | Caveolin 1, caveolae protein | −2.21 | 0.0225 |
| Hk2 | Hexokinase 2 | −1.23 | 0.0456 | |
| Cxcl5 | Chemokine (C-X-C motif) ligand 5 | −1.48 | 0.00085 | |
| Duox2 | Dual oxidase 2 | −4.84 | 0.0046 | |
| Mpo | Myeloperoxidase | −4.53 | 0.04695 | |
| Car9 | Carbonic anhydrase 9 | −4.37 | 0.02705 | |
| Cd24a | CD24a antigen | −1.25 | 5.00 × 10−5 | |
| Plat | Plasminogen activator, tissue | −2.42 | 0.03025 | |
| Ltf | Lactotransferrin | −1.96 | 0.0029 | |
| Selp | Selectin, platelet | −0.921 | 0.02645 | |
| Timp2 | Tissue inhibitor of metalloproteinase 2 | −1.13 | 0.0452 | |
| Up-regulated | Itga2 | Integrin alpha 2 | 0.617 | 0.0154 |
| Fmo3 | Flavin containing monooxygenase 3 | 1.93 | 5.00 × 10−5 | |
| Adc | Arginine decarboxylase | 2 | 0.04095 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.-G.; Park, C.; Hwang, S.; Hong, J.-E.; Jo, M.; Eom, M.; Lee, Y.; Rhee, K.-J. Pulsed Electromagnetic Field (PEMF) Treatment Reduces Lipopolysaccharide-Induced Septic Shock in Mice. Int. J. Mol. Sci. 2022, 23, 5661. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105661
Lee C-G, Park C, Hwang S, Hong J-E, Jo M, Eom M, Lee Y, Rhee K-J. Pulsed Electromagnetic Field (PEMF) Treatment Reduces Lipopolysaccharide-Induced Septic Shock in Mice. International Journal of Molecular Sciences. 2022; 23(10):5661. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105661
Chicago/Turabian StyleLee, Chang-Gun, Chanoh Park, Soonjae Hwang, Ju-Eun Hong, Minjeong Jo, Minseob Eom, Yongheum Lee, and Ki-Jong Rhee. 2022. "Pulsed Electromagnetic Field (PEMF) Treatment Reduces Lipopolysaccharide-Induced Septic Shock in Mice" International Journal of Molecular Sciences 23, no. 10: 5661. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105661