
Penicillin-Binding Proteins, β-Lactamases, and β-Lactamase Inhibitors in β-Lactam-Producing Actinobacteria: Self-Resistance Mechanisms

Abstract
:1. Introduction
Resistance to Antibiotics and Its Relation to Antibiotic-Producing Bacteria
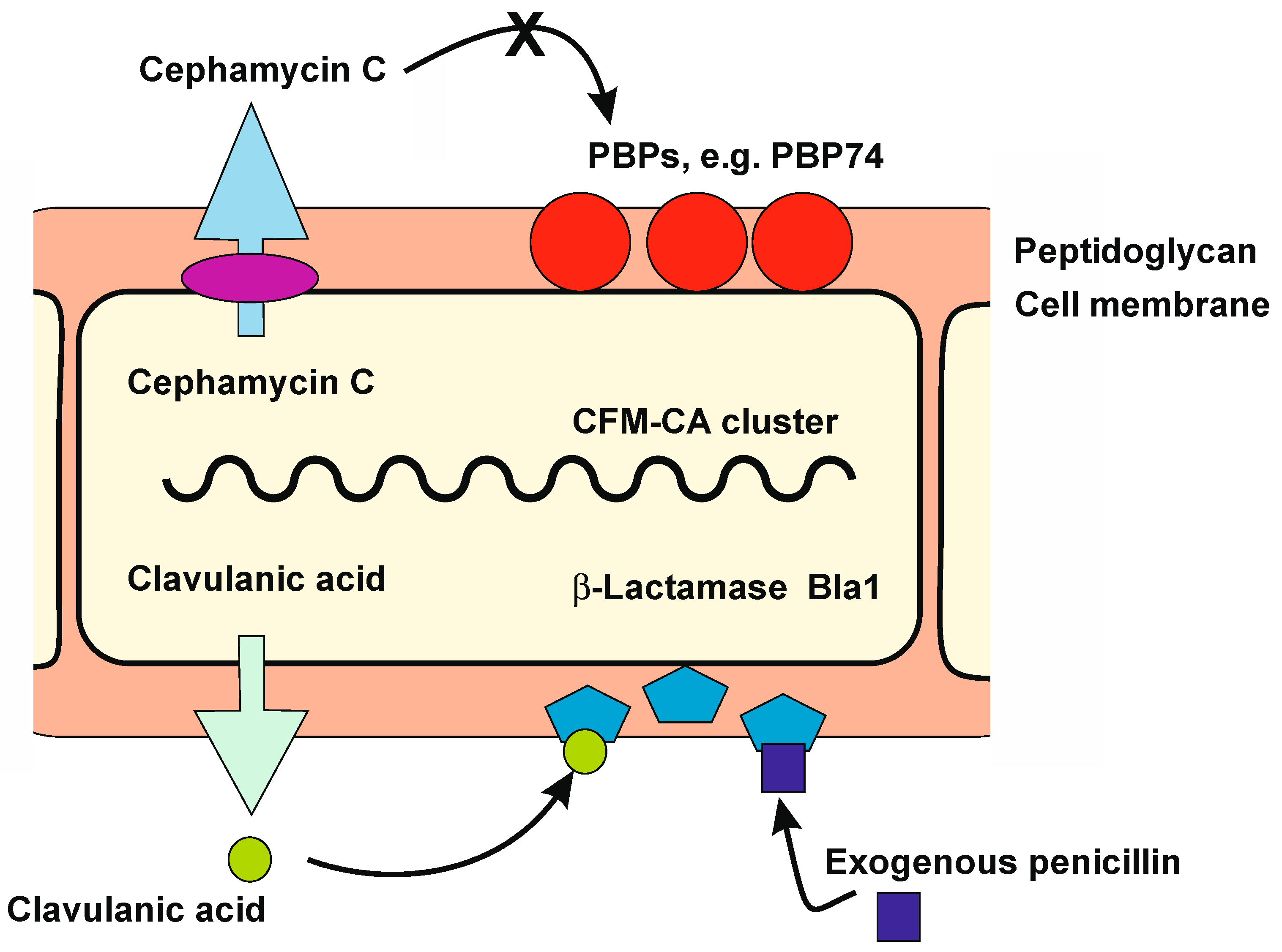
2. Overview of Resistance Mechanisms to Antibiotics in Antibiotic-Producing Actinobacteria
3. Peptidoglycan Biosynthesis and Penicillin-Binding Proteins
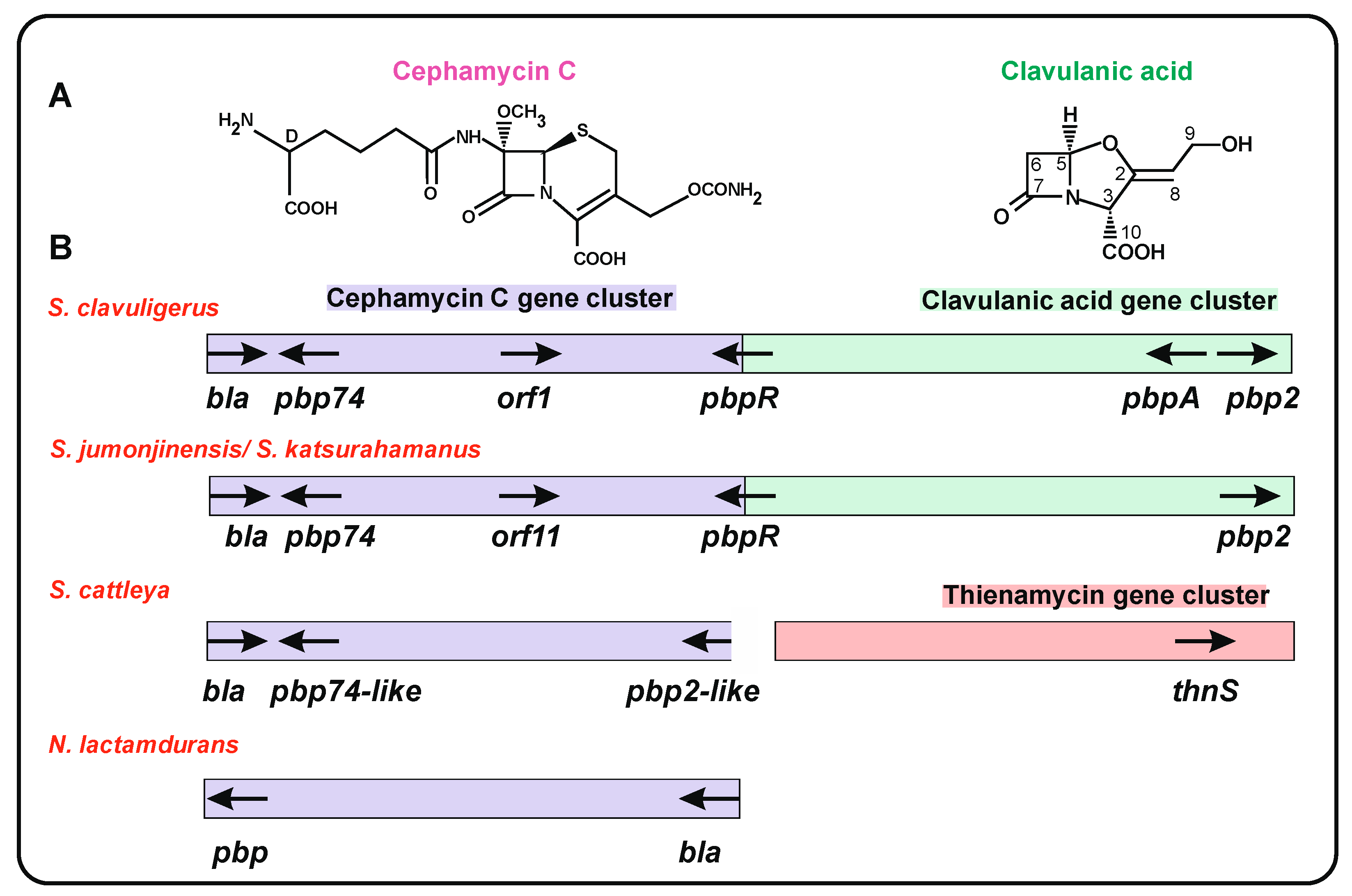
4. Clusters of Genes for β-Lactam Antibiotics in Actinobacteria
5. Genes Encoding PBP Proteins in S. clavuligerus and Other Actinobacteria β-Lactam Gene Clusters

{kind=link}
{kind=link}
{kind=link}
| Protein | Amino Acid Number | Characteristics | Motifs | Accession Number |
|---|---|---|---|---|
| PBP-74 | 693 | Located at the CFM cluster. Amino acids 1–300 are proline rich. Amino acids 300–693 contain carboxypeptidase motifs. An α-helix trans-membrane motif (aa 286–309) separates the proline-rich N-terminal half and the carboxypeptidase moiety. | The PBP-conserved motifs * STAK, SGN (instead of SDN), and KTG are present in the second half of the protein [24]. | WP_003952489 |
| PBP-R | 551 | Located between the clavulanic acid and the cephamycin C cluster. The N-terminal end contains a hydrophobic stretch of 30–60 amino acids, which may act as a membrane anchor. | The C-terminal domain has the penicillin-binding region; it contains the * STFK, SCN (instead of SDN), and KTG motifs of BPBs and β-lactamases [48]. | WP_003952508 |
| PBP-A | 494 | Located at the end of the AC cluster. Contains a pfam0095 transpeptidase domain and an ATP/GTP-binding motif. | Contains the * STFK, STN, and KTG motifs, but lacks the EPELN motif [49]. | WP_003952525 |
| PBP-2 | 717 | Located at the end of the AC cluster. Domain pbp2_mrdA. | Contains SIFK and FTG (instead of KTG) [49]. | WP_003952526 |
| Bla1 | 332 | Located at the CFM cluster.Class A β-lactamase. | Contain the active center 70 STFK and the motifs 130 SDG (instead of SDN), 166 EPELN, and 234 KTG of class A β-lactamases [51]. | WP_003952487 |
| BlaB1 | 338 | Located at the CFM cluster [42]. Class B β-lactamase [this work]. | Motif 117 HGHFD121. | WP_003952502 |
5.1. The PBP-74 Protein
5.2. The PBP-R Protein
5.3. PBP-A and PBP-2
6. β-Lactamases
6.1. Analysis and Role of β-Lactamases Located in the S. clavuligerus β-Lactam Supercluster
Class A β-Lactamases of S. clavuligerus
6.2. Class A β-Lactamases Associated with β-Lactam Clusters in Other Actinobacteria
6.3. Class B β-Lactamases
6.4. Class C β-Lactamases
7. β-Lactamase Inhibitors and β-Lactamase Inhibitory Proteins
7.1. β-Lactamases Inhibitors
7.2. The BLIP Protein of S. clavuligerus and Other Actinobacteria
8. Distinct Arrangement of the Clavulanic Acid Cluster in Actinobacteria
8.1. Is a PBP Gene in the β-Lactam Gene Cluster Required for Self Resistance?

8.2. Distribution of β-Lactamase and Blip Genes in the Cephamycin and Clavulanic Acid GC
8.3. Are Some Genes in the Clavams or Cephamycin Clusters Required for Biosynthesis of Clavulanic Acid
9. Conclusions and Future Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Waksman, S.A.; Reilly, H.C.; Schatz, A. Strain specificity and production of antibiotic substances. V. Strain resistance of bacteria to antibiotic substances, especially streptomycin. Proc. Natl. Acad. Sci. USA 1945, 31, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Demain, A.L. Antibiotics: Natural products essential to human health. Med. Res. Rev. 2009, 29, 821–842. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Wells, V.; Piddock, L.J.V. Addressing antimicrobial resistance in the UK and Europe. Lancet Infect. Dis. 2017, 17, 1230–1231. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.A.G.; Worthington, T. The ‘antibiotic apocalypse’—Scaremongering or scientific reporting? Trends Microbiol. 2017, 25, 167–169. [Google Scholar] [CrossRef] [Green Version]
- Wright, G.D. The antibiotic resistome: The nexus of chemical and genetic diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef]
- D’Costa, V.M.; Griffiths, E.; Wright, G.D. Expanding the soil antibiotic resistome: Exploring environmental diversity. Curr. Opin. Microbiol. 2007, 10, 481–489. [Google Scholar] [CrossRef]
- Azargun, R.; Gholizadeh, P.; Sadeghi, V.; Hosainzadegan, H.; Tarhriz, V.; Memar, M.Y.; Pormohammad, A.; Eyvazi, S. Molecular mechanisms associated with quinolone resistance in Enterobacteriaceae: Review and update. Trans. R. Soc. Trop. Med. Hyg. 2020, 114, 770–781. [Google Scholar] [CrossRef]
- Gomis-Font, M.A.; Cabot, G.; Sánchez-Diener, I.; Fraile-Ribot, P.A.; Juan, C.; Moya, B.; Zamorano, L.; Oliver, A. In vitro dynamics and mechanisms of resistance development to imipenem and imipenem/relebactam in Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2020, 75, 2508–2515. [Google Scholar] [CrossRef]
- Benveniste, R.; Davies, J. Mechanisms of antibiotic resistance in bacteria. Annu. Rev. Biochem. 1973, 42, 471–506. [Google Scholar] [CrossRef]
- Barlow, M.; Hall, B.G. Phylogenetic analysis shows that the OXA beta-lactamase genes have been on plasmids for millions of years. J. Mol. Evol. 2002, 55, 314–321. [Google Scholar] [CrossRef]
- Wencewicz, T.A. Crossroads of Antibiotic Resistance and Biosynthesis. J. Mol. Biol. 2019, 431, 3370–3399. [Google Scholar] [CrossRef]
- Jiang, X.; Ellabaan, M.M.H.; Charusanti, P.; Munck, C.; Blin, K.; Tong, Y.; Weber, T.; Sommer, M.O.A.; Lee, S.Y. Dissemination of antibiotic resistance genes from antibiotic producers to pathogens. Nat. Commun. 2017, 8, 15784. [Google Scholar] [CrossRef] [Green Version]
- Cundliffe, E. How antibiotic-producing organisms avoid suicide. Annu. Rev. Microbiol. 1989, 43, 207–233. [Google Scholar] [CrossRef]
- Cundliffe, E.; Demain, A.L. Avoidance of suicide in antibiotic-producing microbes. J. Ind. Microbiol. Biotechnol. 2010, 37, 643–672. [Google Scholar] [CrossRef]
- Hopwood, D.A. How do antibiotic-producing bacteria ensure their self-resistance before antibiotic biosynthesis incapacitates them? Mol. Microbiol. 2007, 63, 937–940. [Google Scholar] [CrossRef]
- Ogawara, H. Penicillin-binding proteins in Actinobacteria. J. Antibiot. 2015, 68, 223–245. [Google Scholar] [CrossRef] [Green Version]
- Ogawara, H. Comparison of Antibiotic Resistance Mechanisms in Antibiotic-Producing and Pathogenic Bacteria. Molecules 2019, 24, 3430. [Google Scholar] [CrossRef] [Green Version]
- Frére, J.M. β-Lactamases and bacterial resistance to antibiotics. Mol. Microbiol. 1995, 16, 385–395. [Google Scholar] [CrossRef]
- King, D.T.; Sobhanifar, S.; Strynadka, N.C. One ring to rule them all: Current trends in combating bacterial resistance to the β-lactams. Protein Sci. 2016, 25, 787–803. [Google Scholar] [CrossRef] [Green Version]
- Piddock, L.J. Multidrug-resistance efflux pumps—Not just for resistance. Nat. Rev. Microbiol. 2006, 4, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Severi, E.; Thomas, G.H. Antibiotic export: Transporters involved in the final step of natural product production. Microbiology 2019, 165, 805–818. [Google Scholar] [CrossRef]
- Coque, J.J.; Liras, P.; Martín, J.F. Genes for a beta-lactamase, a penicillin-binding protein and a transmembrane protein are clustered with the cephamycin biosynthetic genes in Nocardia lactamdurans. EMBO J. 1993, 12, 631–639. [Google Scholar] [CrossRef]
- Pérez-Llarena, F.J.; Rodríguez-García, A.; Enguita, F.J.; Martín, J.F.; Liras, P. The pcd gene encoding piperideine-6-carboxylate dehydrogenase involved in biosynthesis of alpha-aminoadipic acid is located in the cephamycin cluster of Streptomyces clavuligerus. J. Bacteriol. 1998, 180, 4753–4756. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Burgo, Y.; Álvarez-Álvarez, R.; Pérez-Redondo, R.; Liras, P. Heterologous expression of Streptomyces clavuligerus ATCC 27064 cephamycin C gene cluster. J. Biotechnol. 2014, 186, 21–29. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.A.; Wright, G.D. The antibiotic resistance “mobilome”: Searching for the link between environment and clinic. Front. Microbiol. 2013, 4, 138. [Google Scholar] [CrossRef] [Green Version]
- Forsberg, K.J.; Reyes, A.; Wang, B.; Selleck, E.M.; Sommer, M.O.; Dantas, G. The shared antibiotic resistome of soil bacteria and human pathogens. Science 2012, 337, 1107–1111. [Google Scholar] [CrossRef] [Green Version]
- Durão, P.; Balbontín, R.; Gordo, I. Evolutionary Mechanisms Shaping the Maintenance of Antibiotic Resistance. Trends Microbiol. 2018, 26, 677–691. [Google Scholar] [CrossRef] [Green Version]
- Rioseras, B.; Yagüe, P.; López-García, M.T.; González-Quiñonez, N.; Binda, E.; Marinelli, F.; Manteca, A. Characterization of SCO4439, a D-alanyl-D-alanine carboxypeptidase involved in spore cell wall maturation, resistance, and germination in Streptomyces coelicolor. Sci. Rep. 2016, 6, 21659. [Google Scholar] [CrossRef]
- Jacobs, C.; Huang, L.J.; Bartowsky, E.; Normark, S.; Park, J.T. Bacterial cell wall recycling provides cytosolic muropeptides as effectors for β-lactamase induction. EMBO J. 1994, 13, 4684–4694. [Google Scholar] [CrossRef] [PubMed]
- Van der Aart, L.T.; Spijksm, G.K.; Harms, A.; Vollmer, W.; Hankemeier, T.; van Wezel, G.P. High-Resolution Analysis of the Peptidoglycan Composition in Streptomyces coelicolor. J. Bacteriol. 2018, 200, e00290-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haiser, H.J.; Yousef, M.R.; Elliot, M.A. Cell wall hydrolases affect germination, vegetative growth, and sporulation in Streptomyces coelicolor. J. Bacteriol. 2009, 191, 6501–6512. [Google Scholar] [CrossRef] [Green Version]
- Hugonnet, J.E.; Haddache, N.; Veckerle, C.; Dubost, L.; Marie, A.; Shikura, N.; Mainardi, J.L.; Rice, L.B.; Arthur, M. Peptidoglycan cross-linking in glycopeptide-resistant Actinomycetales. Antimicrob. Agents Chemother. 2014, 58, 1749–1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stapley, E.O.; Jackson, M.; Hernández, S.; Zimmerman, S.B.; Currie, S.A.; Mochales, S.; Mata, J.M.; Woodruff, H.B.; Hendlin, D. Cephamycins, a new family of beta-lactam antibiotics I. Production by actinomycetes, including Streptomyces lactamdurans sp. Antimicrob. Agents Chemother. 1972, 2, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Aharonowitz, Y.; Cohen, G.; Martín, J.F. Penicillin and cephalosporin biosynthetic genes: Structure, organization, regulation, and evolution. Annu. Rev. Microbiol. 1992, 46, 461–495. [Google Scholar] [CrossRef]
- Liras, P. Biosynthesis and Molecular Genetics of Cephamycins. Ant. Van Leeuwenhoek 1999, 75, 109–124. [Google Scholar] [CrossRef]
- Brown, A.G.; Butterworth, D.; Cole, M.; Hanscomb, G.; Hood, J.D.; Reading, C.; Rolinson, G.N. Naturally-occurring beta-lactamase inhibitors with antibacterial activity. J. Antibiot. 1976, 29, 668–669. [Google Scholar] [CrossRef]
- Reading, C.; Cole, M. Clavulanic acid: A beta-lactamase-inhibiting beta-lactam from Streptomyces clavuligerus. Antimicrob. Agents Chemother. 1977, 11, 852–857. [Google Scholar] [CrossRef] [Green Version]
- Drawz, S.M.; Bonomo, R.A. Three decades of beta-lactamase inhibitors. Clin. Microbiol. Rev. 2010, 23, 160–201. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.M.; Hodgson, J.E. The biosynthetic genes for clavulanic acid and cephamycin production occur as a ‘super-cluster’ in three Streptomyces. FEMS Microbiol. Lett. 1993, 110, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Llarena, F.J.; Liras, P.; Rodríguez-García, A.; Martín, J.F. A regulatory gene (ccaR) required for cephamycin and clavulanic acid production in Streptomyces clavuligerus: Amplification results in overproduction of both beta-lactam compounds. J. Bacteriol. 1997, 179, 2053–2059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, D.C.; Jensen, S.E. Investigation of the Streptomyces clavuligerus cephamycin C gene cluster and its regulation by the CcaR protein. J. Bacteriol. 1998, 180, 4068–4079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamarta, I.; López-García, M.T.; Kurt, A.; Nárdiz, N.; Alvarez-Álvarez, R.; Pérez-Redondo, R.; Martín, J.F.; Liras, P. Characterization of DNA-binding sequences for CcaR in the cephamycin-clavulanic acid supercluster of Streptomyces clavuligerus. Mol. Microbiol. 2011, 81, 968–981. [Google Scholar] [CrossRef]
- Álvarez-Álvarez, R.; Martínez-Burgo, Y.; Pérez-Redondo, R.; Braña, A.F.; Martín, J.F.; Liras, P. Expression of the endogenous and heterologous clavulanic acid cluster in Streptomyces flavogriseus: Why a silent cluster is sleeping. Appl. Microb. Biotechnol. 2013, 97, 9451–9463. [Google Scholar] [CrossRef]
- Ogawara, H. Antibiotic resistance in pathogenic and producing bacteria, with special reference to β-lactam antibiotics. Microbiol. Rev. 1981, 45, 591–619. [Google Scholar] [CrossRef]
- Medema, M.H.; Trefzer, A.; Kovalchuk, A.; van den Berg, M.; Müller, U.; Heijne, W.; Wu, L.; Alam, M.T.; Ronning, C.M.; Nierman, W.C.; et al. The sequence of a 1.8-mb bacterial linear plasmid reveals a rich evolutionary reservoir of secondary metabolic pathways. Genome Biol. Evol. 2010, 2, 212–224. [Google Scholar] [CrossRef]
- Paradkar, A.S.; Aidoo, K.A.; Wong, A.; Jensen, S.E. Molecular analysis of a beta-lactam resistance gene encoded within the cephamycin gene cluster of Streptomyces clavuligerus. J. Bacteriol. 1996, 178, 6266–6274. [Google Scholar] [CrossRef] [Green Version]
- Mellado, E.; Lorenzana, L.M.; Rodríguez-Sáiz, M.; Díez, B.; Liras, P.; Barredo, J.L. The clavulanic acid biosynthetic cluster of Streptomyces clavuligerus: Genetic organization of the region upstream of the car gene. Microbiology 2002, 148, 1427–1438. [Google Scholar] [CrossRef] [Green Version]
- Jensen, S.E.; Paradkar, A.S.; Mosher, R.H.; Anders, C.; Beatty, P.H.; Brumlik, M.J.; Griffin, A.; Barton, B. Five additional genes are involved in clavulanic acid biosynthesis in Streptomyces clavuligerus. Antimicrob. Agents Chemother. 2004, 48, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Llarena, F.; Martín, J.F.; Galleni, M.; Coque, J.J.; Fuente, J.L.; Frère, J.M.; Liras, P. The bla gene of the cephamycin cluster of Streptomyces clavuligerus encodes a class A beta-lactamase of low enzymatic activity. J. Bacteriol. 1997, 179, 6035–6040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, K.; Hung, T.V.; Liou, K.; Lee, H.C.; Shin, C.-H.; Sohng, J.K. Characterization of pbpA and pbp2 encoding penicillin-binding proteins located on the downstream of clavulanic acid gene cluster in Streptomyces clavuligerus. Biotechnol. Lett. 2006, 28, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Ogawara, H. Self-resistance in Streptomyces, with Special Reference to β-Lactam Antibiotics. Molecules 2016, 10, 605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, H.; Martin, M.T.; Waley, S.G. Beta-lactamases as fully efficient enzymes. Determination of all the rate constants in the acyl-enzyme mechanism. Biochem. J. 1990, 266, 853–861. [Google Scholar] [PubMed]
- Strynadka, N.C.; Adachi, H.; Jensen, S.E.; Johns, K.; Sielecki, A.; Betzel, C.; Sutoh, K.; James, M.N. Molecular structure of the acyl-enzyme intermediate in beta-lactam hydrolysis at 1.7 A resolution. Nature 1992, 359, 700–705. [Google Scholar] [CrossRef]
- Adachi, H.; Ohta, T.; Matsuzawa, H. Site-directed mutants, at position 166, of RTEM-1 beta-lactamase that form a stable acyl-enzyme intermediate with penicillin. J. Biol. Chem. 1991, 266, 3186–3191. [Google Scholar] [CrossRef]
- Guillaume, G.; Vanhove, M.; Lamotte-Brasseur, J.; Ledent, P.; Jamin, M.; Joris, B.; Frère, J.M. Site-directed mutagenesis of glutamate 166 in two beta-lactamases: Kinetic and molecular modelling studies. J. Biol. Chem. 1997, 272, 5438–5444. [Google Scholar] [CrossRef] [Green Version]
- Valegård, K.; Iqbal, A.; Kershaw, N.J.; Ivison, D.; Généreux, C.; Dubus, A.; Blikstad, C.; Demetriades, M.; Hopkinson, R.J.; Lloyd, A.J.; et al. Structural and mechanistic studies of the orf12 gene product from the clavulanic acid biosynthesis pathway. Acta Crystallogr. Sect. D Biol. Crystallogr. 2013, 69, 1567–1579. [Google Scholar] [CrossRef]
- Srivastava, S.K.; King, K.S.; AbuSara, N.F.; Malayny, C.J.; Piercey, B.M.; Wilson, J.A.; Tahlan, K. In vivo functional analysis of a class A β-lactamase-related protein essential for clavulanic acid biosynthesis in Streptomyces clavuligerus. PLoS ONE 2019, 14, e0215960. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; de la Fuente, J.L.; Leitão, A.L.; Liras, P.; Martín, J.F. Effect of amplification or targeted disruption of the beta-lactamase gene of Nocardia lactamdurans on cephamycin biosynthesis. Appl. Microbiol. Biotechnol. 1996, 45, 621–628. [Google Scholar] [CrossRef]
- Barbe, V.; Bouzon, M.; Mangenot, S.; Badet, B.; Poulain, J.; Segurens, B.; Vallenet, D.; Marlière, P.; Weissenbach, J. Complete genome sequence of Streptomyces cattleya NRRL 8057, a producer of antibiotics and fluorometabolites. J. Bacteriol. 2011, 193, 5055–5056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diene, S.M.; Pinault, L.; Baron, S.A.; Azza, S.; Armstrong, N.; Hadjadj, L.; Chabrière, E.; Rolain, J.M.; Pontarotti, P.; Raoult, D. A metallo-beta-lactamase enzyme for internal detoxification of the antibiotic thienamycin. Sci. Rep. 2021, 11, 10062. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.; Belasco, J.G.; Charnas, R.L.; Khosla, S.; Knowles, J.R. β-Lactamase inactivation by mechanism-based reagents. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1980, 289, 309–319. [Google Scholar]
- Bush, K. β-Lactamase inhibitors from laboratory to clinic. Clin. Microbiol. Rev. 1988, 1, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.E.; Paradkar, A.S. Biosynthesis and molecular genetics of clavulanic acid. Ant. Van Leeuwenhoek 1999, 75, 125–133. [Google Scholar] [CrossRef]
- Liras, P.; Rodríguez-García, A. Clavulanic acid, a β-lactamase inhibitor: Biosynthesis and molecular genetics. Appl. Microbiol. Biotechnol. 1999, 54, 467–475. [Google Scholar] [CrossRef]
- Liras, P.; Martín, J.F. Streptomyces clavuligerus: The Omics Era. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab072. [Google Scholar] [CrossRef]
- Spratt, B.G.; Jobanputra, V.; Zimmermann, W. Binding of thienamycin and clavulanic acid to the penicillin-binding proteins of Escherichia coli K-12. Antimicrob. Agents Chemother. 1977, 12, 406–409. [Google Scholar] [CrossRef] [Green Version]
- Strynadka, N.C.; Jensen, S.E.; Johns, K.; Blanchard, H.; Page, M.; Matagne, A.; Frère, J.M.; James, M.N. Structural and kinetic characterization of a beta-lactamase-inhibitor protein. Nature 1994, 368, 657–660. [Google Scholar] [CrossRef]
- Doran, J.L.; Leskiw, B.K.; Aippersbach, S.; Jensen, S.E. Isolation and characterization of a beta-lactamase-inhibitory protein from Streptomyces clavuligerus and cloning and analysis of the corresponding gene. J. Bacteriol. 1990, 172, 4909–4918. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Palzkill, T. Dissecting the protein-protein interface between beta-lactamase inhibitory protein and class A beta-lactamases. J. Biol. Chem. 2004, 279, 42860–42866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Huang, W.; Chow, D.-C.; Palzkill, T. Fine mapping of the sequence requirements for binding of beta-lactamase inhibitory protein (BLIP) to TEM-1 beta-lactamase using a genetic screen for BLIP function. J. Mol. Biol. 2009, 389, 401–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gretes, M.; Lim, D.C.; de Castro, L.; Jensen, S.E.; Kang, S.G.; Lee, K.J.; Strynadka, N.C. Insights into positive and negative requirements for protein-protein interactions by crystallographic analysis of the beta-lactamase inhibitory proteins BLIP, BLIP-I, and BLP-II. J. Mol. Biol. 2009, 389, 289–305. [Google Scholar] [CrossRef] [PubMed]
- Park, H.U.; Lee, K.J. Cloning and heterologous expression of the gene for BLIP-II, a β-lactamase-inhibitory protein from Streptomyces exfoliatus SMF19. Microbiology 1998, 144, 2161–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, D.; Park, H.U.; De Castro, L.; Kang, S.G.; Lee, H.S.; Jensen, S.; Lee, K.J.; Strynadka, N.C. Crystal structure and kinetic analysis of beta-lactamase inhibitor protein-II in complex with TEM-1 beta-lactamase. Nat. Struct. Biol. 2001, 8, 848–852. [Google Scholar] [CrossRef]
- AbuSara, N.F.; Piercey, B.M.; Moore, M.A.; Shaikh, A.A.; Nothias, L.F.; Srivastava, S.K.; Cruz-Morales, P.; Dorrestein, P.C.; Barona-Gómez, F.; Tahlan, K. Comparative Genomics and Metabolomics Analyses of Clavulanic Acid-Producing Streptomyces Species Provides Insight Into Specialized Metabolism. Front. Microbiol. 2019, 10, 2550. [Google Scholar] [CrossRef] [PubMed]
- Tahlan, K.; Park, H.U.; Jensen, S.E. Three unlinked gene clusters are involved in clavam metabolite biosynthesis in Streptomyces clavuligerus. Can. J. Microbiol. 2004, 50, 803–810. [Google Scholar] [CrossRef]
- Tahlan, K.; Park, H.U.; Wong, A.; Beatty, P.H.; Jensen, S.E. Two sets of paralogous genes encode the enzymes involved in the early stages of clavulanic acid and clavam metabolite biosynthesis in Streptomyces clavuligerus. Antimicrob. Agents Chemother. 2004, 48, 930–939. [Google Scholar] [CrossRef] [Green Version]
- Challis, G.L.; Hopwood, D.A. Synergy and contingency as driving forces for the evolution of multiple secondary metabolite production by Streptomyces species. Proc. Nat. Acad. Sci. USA 2003, 100, 14555–14561. [Google Scholar] [CrossRef] [Green Version]
- Egan, L.A.; Busby, R.W.; Iwata-Reuyl, D.; Townsend, C.A. Probable role of clavaminic acid as the terminal intermediate in the common pathway to clavulanic acid and the antipodal clavam metabolites. J. Am. Chem. Soc. 1997, 119, 2348–2355. [Google Scholar] [CrossRef]
- Jensen, S.E. Biosynthesis of clavam metabolites. J. Ind. Microbiol. Biotechnol. 2012, 39, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Romero, J.; Liras, P.; Martín, J.F. Dissociation of cephamycin and clavulanic acid biosynthesis in Streptomyces clavuligerus. Appl. Microbiol. Biotechnol. 1984, 20, 318–325. [Google Scholar] [CrossRef]
- Pérez-Redondo, R.; Rodríguez-García, A.; Martín, J.F.; Liras, P. The claR gene of Streptomyces clavuligerus, encoding a LysR-type regulatory protein controlling clavulanic acid biosynthesis, is linked to the clavulanate-9-aldehyde reductase (car) gene. Gene 1998, 211, 311–321. [Google Scholar] [CrossRef]
- Martín, J.F.; Casqueiro, J.; Liras, P. Secretion systems for secondary metabolites: How producer cells send out messages of intercellular communication. Curr. Opin. Microbiol. 2005, 8, 282–293. [Google Scholar] [CrossRef] [PubMed]


| Strain | Amino Acid Number 1 | Identity/ Similarity 2 (%) | Location | Accession Number |
|---|---|---|---|---|
| A. Proteins with identity/similarity to S. clavuligerus PBP-74 | ||||
| S. clavuligerus | 693 | 100 | CFM cluster | WP_003952489 |
| S. jumonjinensis | 437 | 67/77 | CFM cluster | MQT02641 |
| S. katsurahamanus | 862 | 71/81 | CFM cluster | MQS35996 |
| S. sulfonofaciens | 843 | 62 | CFM cluster | WP_229924744 |
| S. megasporus | 388 | 68 | CFM cluster | WP_031505760 |
| S. cattleya | 419 | 63/74 | CFM cluster | CCB78378 |
| B. Proteins with identity/similarity to S. clavuligerus PBP-R | ||||
| Streptomyces clavuligerus | 551 | 100 to PBP-R | CFM-CA 3 | WP_003952508 |
| Streptomyces jumonjinensis | 550 | 76/87 | CFM-CA 3 | MQT02628 |
| Streptomyces katsurahamanus | 550 | 76/87 | CFM-CA 3 | MQS35981 |
| Streptomyces fulvorobeus | 555 | 66/77 | CA cluster | WP_173313670 |
| Streptomyces albiflavescens | 548 | 71/83 | CA cluster | WP_189188591 |
| Streptomyces sp. M41 | 548 | 72/83 | CA cluster | WP_081218566 |
| Streptomyces sp. SID14446 | 548 | 71/82 | CA cluster | WP_164372891 |
| Streptomyces sp. B93 | 549 | 71/83 | CA cluster | WP_210923884 |
| Streptomyces sp. SID2888 | 547 | 67/79 | CA cluster | WP_161240914 |
| Streptomyces sp. NRRL 24051 | 553 | 61/74 | CA cluster | WP_014152699 |
| Streptomyces sp. S-325 | 553 | 61/74 | CA cluster | WP_014152699 |
| Streptomyces sp. SM10 | 556 | 61/74 | CA cluster | WP_103513573 |
| Streptomyces sp. PMAC2608 | 553 | 61/74 | CA cluster | WP_014152699 |
| Streptomyces flavovirens | 553 | 60/74 | CA cluster | WP_030636769 |
| Streptomyces flavogriseus | 551 | 61/74 | CA cluster | MBD28344891 |
| Streptomyces finlayi | 550 | 67/79 | CA cluster | WP_185300304 |
| Streptomyces sulfonofaciens | 549 | 70/80 | CFM cluster | WP_189933512 |
| C. Proteins with identity/similarity to S. clavuligerus PBP-A | ||||
| Streptomyces clavuligerus | 494 | 100 | CA cluster | WP_003952525 |
| Streptomyces sp. B93 | 494 | 78/89 | CA cluster | WP_210923906 |
| D. Proteins with identity/similarity to S. clavuligerus PBP-2 | ||||
| Streptomyces clavuligerus | 764 | 100 | CA cluster | WP_003952526 |
| Streptomyces jumonjinensis | 721 | 80 | CA cluster | MQ535963 |
| Streptomyces katsurahamanus | 721 | 80 | CA cluster | MQT02611 |
| Streptomyces cattleya | 695 | 66/78 | CFM cluster | CCB78364 |
| Strain | Amino Acid Number | Identity/ Similarity 1 (%) | Location | Accession Number |
|---|---|---|---|---|
| A. Genes encoding class A β-lactamases | ||||
| Streptomyces clavuligerus | 332 | 100 | CFM cluster | WP_003952487 |
| Streptomyces jumonjinensis | 332 | 73 | CFM cluster | WP_153524219 |
| Streptomyces katsurahamanus | 332 | 76 | CFM cluster | WP_153482554 |
| Streptomyces megasporus | 312 | 63/73 | CFM cluster | WP_031505759 |
| Streptomyces fulvorobeus | 332 | 42/54 | CFM cluster | WP_179764187 |
| Nocardia lactamdurans | 302 | 46/59 | CFM cluster | Z13971 |
| Streptomyces cattleya | 310 | 60/74 | CFM cluster | CCB78379.1 |
| B. Genes encoding class B MBL fold Metallo-hydrolases | ||||
| Streptomyces clavuligerus BlaB1 | 338 | 100 | CFM cluster | WP_003952502 |
| Streptomyces SID2888 | 331 | 73/81 | CA cluster | WP_161240912 |
| Streptomyces albiflavescens | 332 | 71/92 | CA cluster | WP_189188593 |
| Streptomyces M41 | 332 | 70/78 | CA cluster | WP_081218564 |
| Streptomyces SID1446 | 332 | 70/78 | CA cluster | WP_164372889 |
| Streptomyces fulvorobeus | 342 | 52/65 | CA cluster | WP_173313669 |
| Streptomyces sp. SM10 | 334 | 52/65 | CA cluster | WP_103513567 |
| Streptomyces sp. PMAC2608 | 335 | 52/66 | CA cluster | WP_014152693 |
| Saccharomonospora viridis | 315 | 39/52 | CA cluster | WP_015787604 |
| Streptomyces flavovirens | 335 | 53/67 | CA cluster | WP_030636771 |
| Streptomyces sp. S-325 | 335 | 53/67 | CA cluster | WP_030636771 |
| Streptomyces sp. NRRL-24051 | 335 | 53/66 | CA cluster | WP_030124616 |
| Streptomyces sulfonofaciens | 338 | 41/52 | CFM cluster | WP_189933518 |
| Streptomyces flavogriseus ATCC 33331 | 354 | 58/67 | THN cluster | ADW01616 |
| Streptomyces cattleya ThnS | 329 | 43/68 | THN cluster | CCB71864 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin, J.F.; Alvarez-Alvarez, R.; Liras, P. Penicillin-Binding Proteins, β-Lactamases, and β-Lactamase Inhibitors in β-Lactam-Producing Actinobacteria: Self-Resistance Mechanisms. Int. J. Mol. Sci. 2022, 23, 5662. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105662
Martin JF, Alvarez-Alvarez R, Liras P. Penicillin-Binding Proteins, β-Lactamases, and β-Lactamase Inhibitors in β-Lactam-Producing Actinobacteria: Self-Resistance Mechanisms. International Journal of Molecular Sciences. 2022; 23(10):5662. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105662
Chicago/Turabian StyleMartin, Juan F., Ruben Alvarez-Alvarez, and Paloma Liras. 2022. "Penicillin-Binding Proteins, β-Lactamases, and β-Lactamase Inhibitors in β-Lactam-Producing Actinobacteria: Self-Resistance Mechanisms" International Journal of Molecular Sciences 23, no. 10: 5662. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105662