
Binding Properties of RNA Quadruplex of SARS-CoV-2 to Berberine Compared to Telomeric DNA Quadruplex

 ,
,
Abstract
:1. Introduction
2. Results
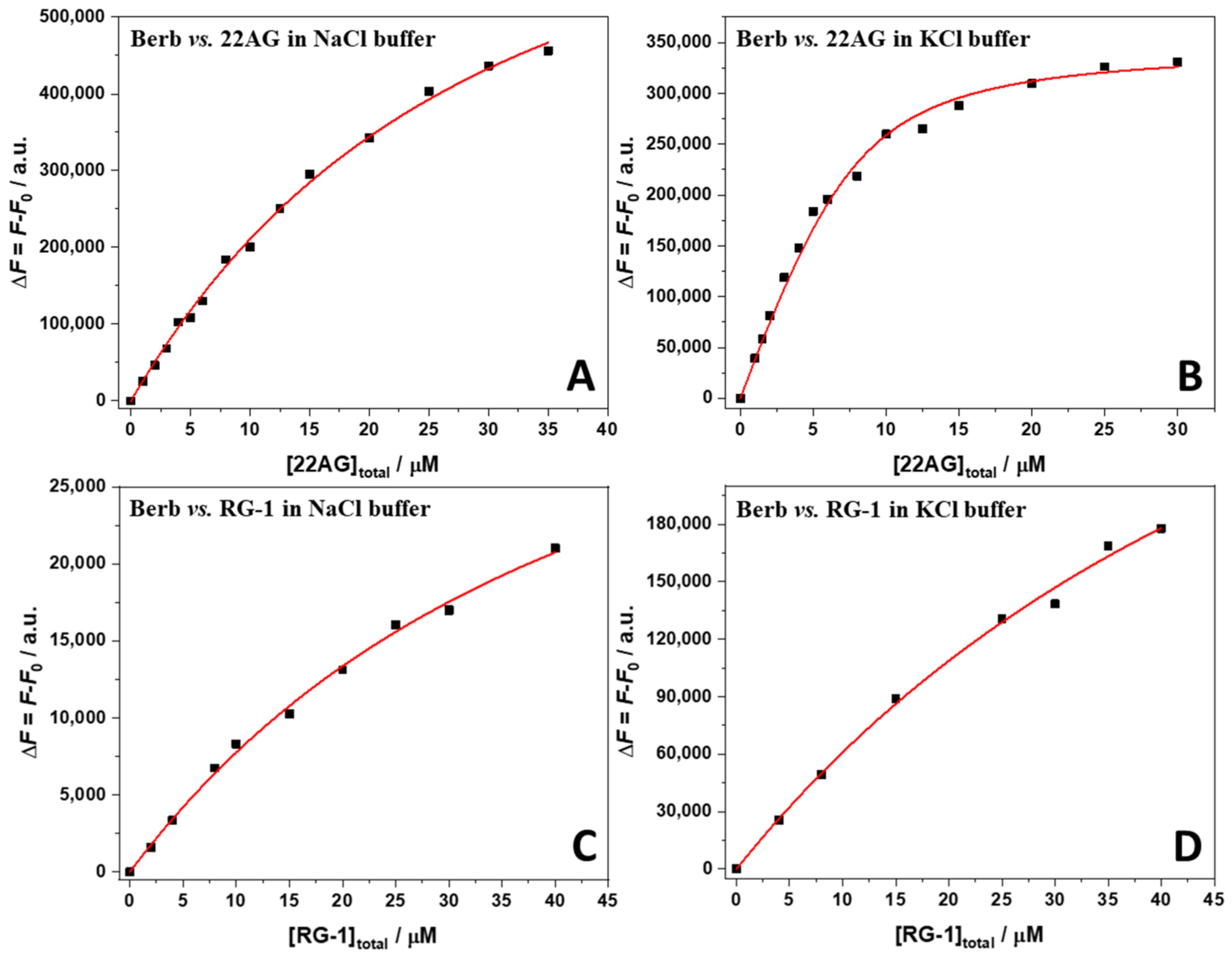
2.1. Binding of Berberine to 22AG and RG-1 at Different Solution Conditions
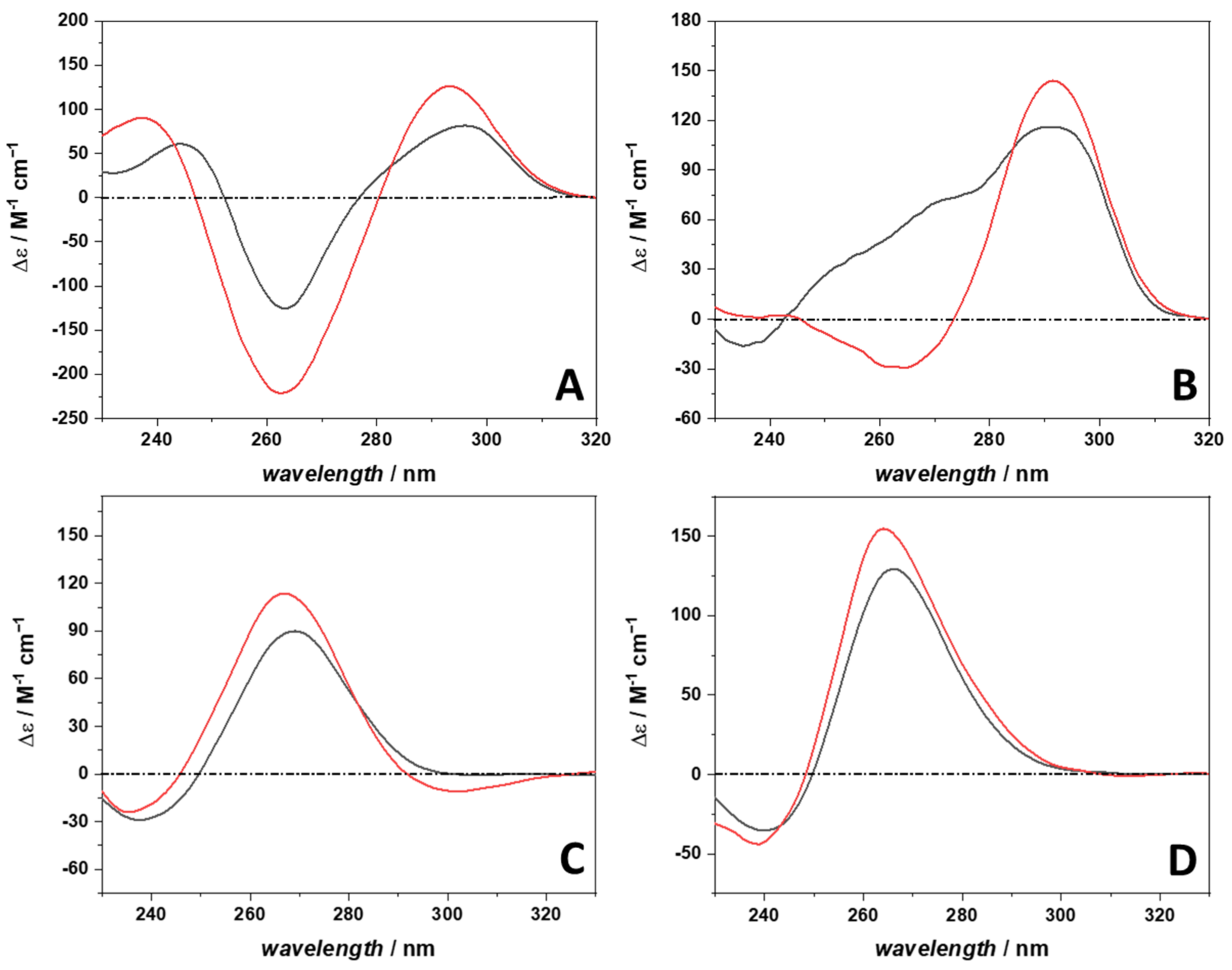
2.2. The Conformational Behavior of 22AG and RG-1 in Complex with Berberine
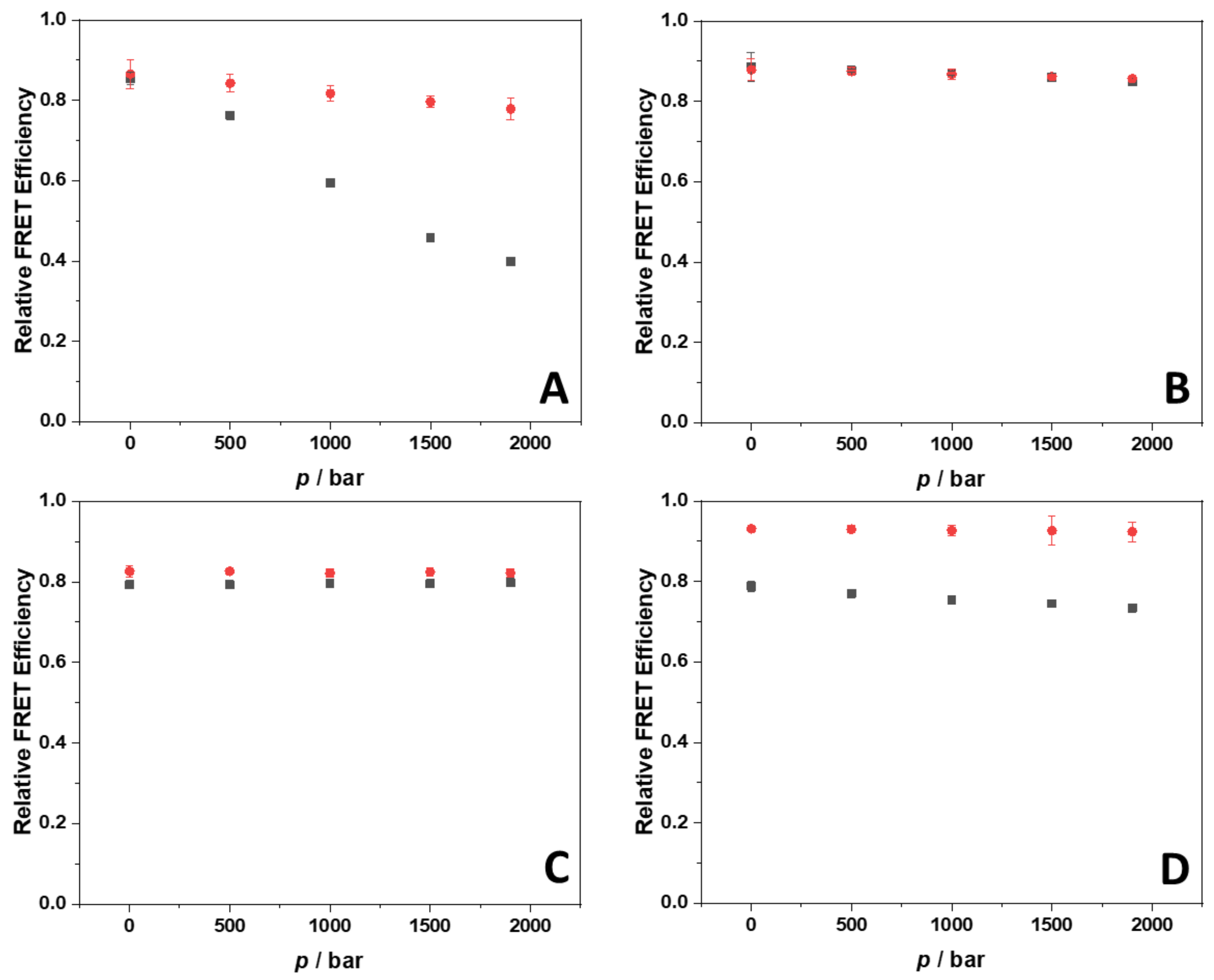
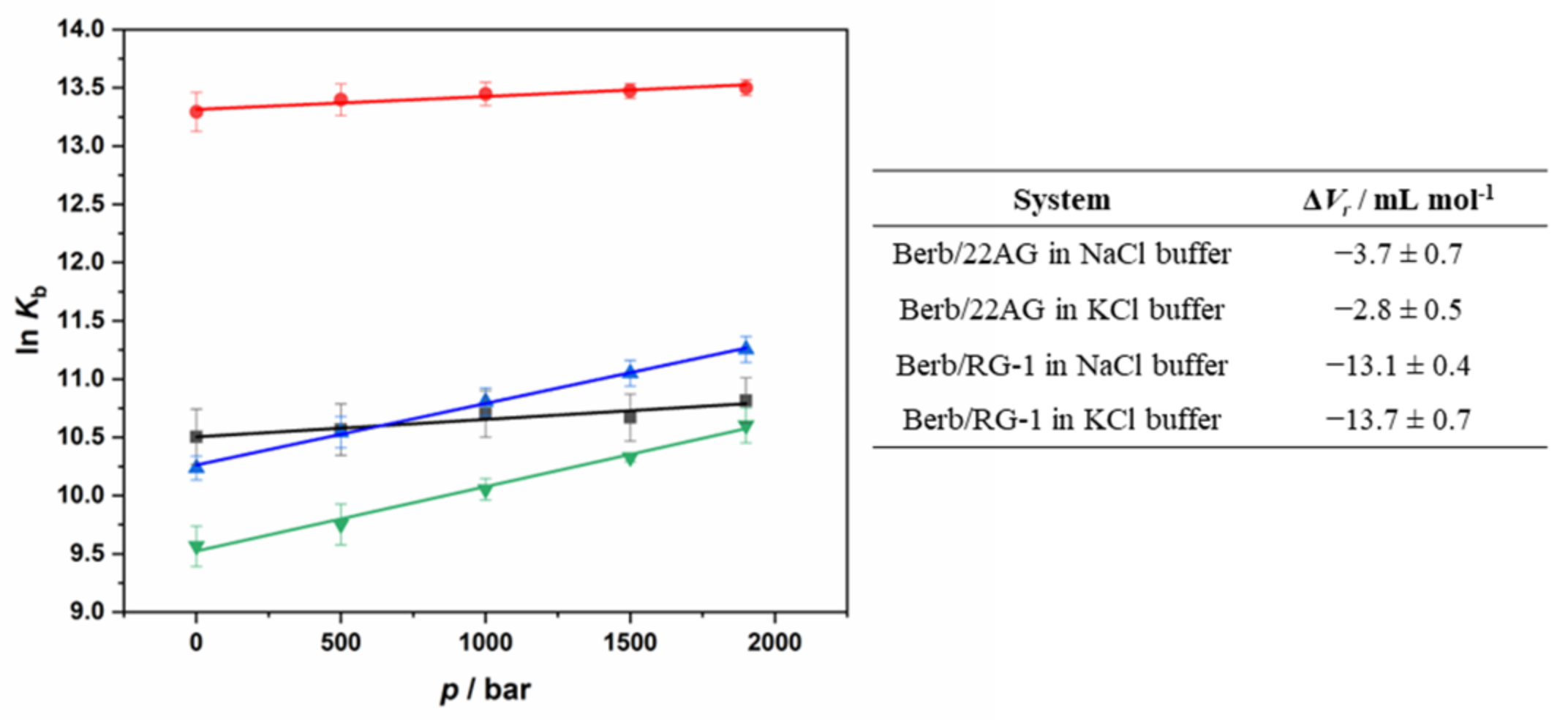
2.3. Probing Volumetric and Hydration Properties upon Complex Formation between Berberine and G4Qs Using Pressure Modulation Spectroscopy
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Sample Preparation
4.3. UV/Vis Spectrometry
4.4. Steady-State Fluorescence Spectroscopy
4.5. Job’s Plot
4.6. Fluorescence Resonance Energy Transfer
4.7. Circular Dichroism Spectroscopy
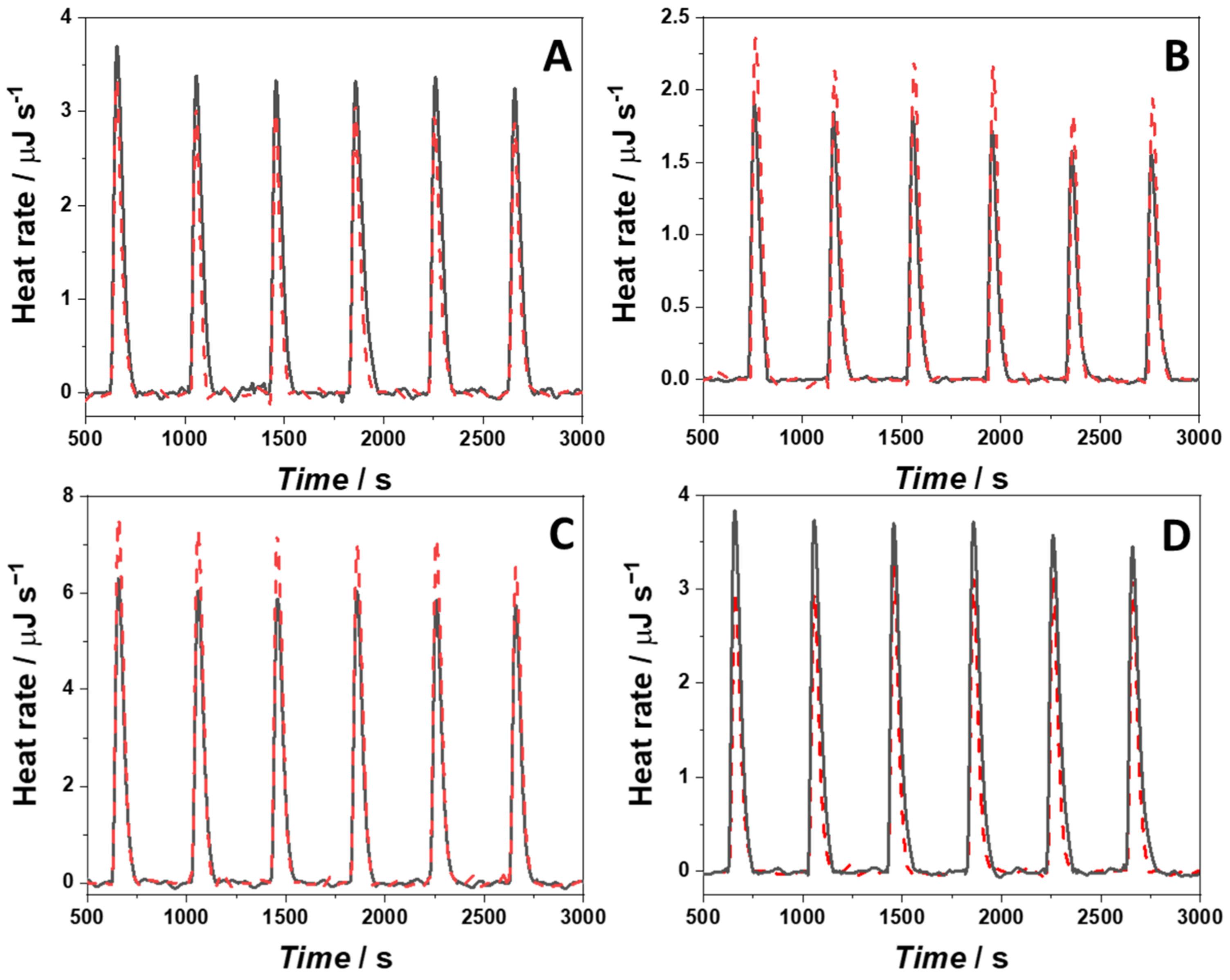
4.8. Isothermal Titration Calorimetry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Synowiec, A.; Szczepański, A.; Barreto-Duran, E.; Lie, L.K.; Pyrc, K. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): A Systemic Infection. Clin. Microbiol. Rev. 2021, 34, e00133-20. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xiao, Z.; Ye, K.; He, X.; Sun, B.; Qin, Z.; Yu, J.; Yao, J.; Wu, Q.; Bao, Z.; et al. SARS-CoV-2: Characteristics and current advances in research. Virol. J. 2020, 17, 117. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, U.M.; Abokor, A.A.; Edwards, J.M.; Waigi, E.W.; Royfman, R.S.; Hasan, S.A.-M.; Smedlund, K.B.; Hardy, A.M.G.; Chakravarti, R.; Koch, L.G. SARS-CoV-2, ACE2 expression, and systemic organ invasion. Physiol. Genom. 2021, 53, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Gil, C.; Ginex, T.; Maestro, I.; Nozal, V.; Barrado-Gil, L.; Cuesta-Geijo, M.; Urquiza, J.; Ramírez, D.; Alonso, C.; Campillo, N.E.; et al. COVID-19: Drug Targets and Potential Treatments. J. Med. Chem. 2020, 63, 12359–12386. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, A.; Mohammad, T.; Anwar, S.; Amani, S.; Khan, M.S.; Husain, F.M.; Rehman, T.; Islam, A.; Hassan, I. Potential drug targets of SARS-CoV-2: From genomics to therapeutics. Int. J. Biol. Macromol. 2021, 177, 1–9. [Google Scholar] [CrossRef]
- Zhang, Y.; Geng, X.; Tan, Y.; Li, Q.; Xu, C.; Xu, J.; Hao, L.; Zeng, Z.; Luo, X.; Liu, F.; et al. New understanding of the damage of SARS-CoV-2 infection outside the respiratory system. Biomed. Pharmacother. 2020, 127, 110195. [Google Scholar] [CrossRef]
- Su, H.; Zhou, F.; Huang, Z.; Ma, X.; Natarajan, K.; Zhang, M.; Huang, Y.; Su, H. Molecular Insights into Small-Molecule Drug Discovery for SARS-CoV-2. Angew. Chem. Int. Ed. 2021, 60, 9789–9802. [Google Scholar] [CrossRef]
- Zhao, C.; Qin, G.; Niu, J.; Wang, Z.; Wang, C.; Ren, J.; Qu, X. Targeting RNA G-Quadruplex in SARS-CoV-2: A Promising Therapeutic Target for COVID-19? Angew. Chem. Int. Ed. 2021, 60, 432–438. [Google Scholar] [CrossRef]
- Sreeramulu, S.; Richter, C.; Berg, H.; Martin, M.A.W.; Ceylan, B.; Matzel, T.; Adam, J.; Altincekic, N.; Azzaoui, K.; Bains, J.K.; et al. Exploring the Druggability of Conserved RNA Regulatory Elements in the SARS-CoV-2 Genome. Angew. Chem. Int. Ed. 2021, 60, 19191–19200. [Google Scholar] [CrossRef]
- Bezzi, G.; Piga, E.; Binolfi, A.; Armas, P. CNBP Binds and Unfolds In Vitro G-Quadruplexes Formed in the SARS-CoV-2 Positive and Negative Genome Strands. Int. J. Mol. Sci. 2021, 22, 2614. [Google Scholar] [CrossRef]
- Varshney, D.; Spiegel, J.; Zyner, K.; Tannahill, D.; Balasubramanian, S. The regulation and functions of DNA and RNA G-quadruplexes. Nat. Rev. Mol. Cell Biol. 2020, 21, 459–474. [Google Scholar] [CrossRef]
- Dumas, L.; Herviou, P.; Dassi, E.; Cammas, A.; Millevoi, S. G-Quadruplexes in RNA Biology: Recent Advances and Future Directions. Trends Biochem. Sci. 2021, 46, 270–283. [Google Scholar] [CrossRef]
- Rhodes, D.; Lipps, H.J. G-quadruplexes and their regulatory roles in biology. Nucleic Acids Res. 2015, 43, 8627–8637. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, S.; Kotar, A.; Tateishi-Karimata, H.; Bhowmik, S.; Wang, Z.-F.; Chang, T.-C.; Sato, S.; Takenaka, S.; Plavec, J.; Sugimoto, N. Chemical Modulation of DNA Replication along G-Quadruplex Based on Topology-Dependent Ligand Binding. J. Am. Chem. Soc. 2021, 143, 16458–16469. [Google Scholar] [CrossRef]
- Takahashi, S.; Sugimoto, N. Watson–Crick versus Hoogsteen Base Pairs: Chemical Strategy to Encode and Express Genetic Information in Life. Acc. Chem. Res. 2021, 54, 2110–2120. [Google Scholar] [CrossRef]
- Mitteaux, J.; Lejault, P.; Wojciechowski, F.; Joubert, A.; Boudon, J.; Desbois, N.; Gros, C.P.; Hudson, R.H.E.; Boulé, J.-B.; Granzhan, A.; et al. Identifying G-Quadruplex-DNA-Disrupting Small Molecules. J. Am. Chem. Soc. 2021, 143, 12567–12577. [Google Scholar] [CrossRef]
- Sugimoto, N. Chemistry and Biology of Non-Canonical Nucleic Acids, 1st ed.; Wiley: Hoboken, NJ, USA, 2020; ISBN 978-3-527-81786-3. [Google Scholar]
- Belmonte-Reche, E.; Serrano-Chacón, I.; Gonzalez, C.; Gallo, J.; Bañobre-López, M. Potential G-quadruplexes and i-Motifs in the SARS-CoV-2. PLoS ONE 2021, 16, e0250654. [Google Scholar] [CrossRef]
- Liu, G.; Du, W.; Sang, X.; Tong, Q.; Wang, Y.; Chen, G.; Yuan, Y.; Jiang, L.; Cheng, W.; Liu, D.; et al. RNA G-quadruplex in TMPRSS2 reduces SARS-CoV-2 infection. Nat. Commun. 2022, 13, 1444. [Google Scholar] [CrossRef]
- Wang, Z.; Li, K.; Maskey, A.R.; Huang, W.; Toutov, A.A.; Yang, N.; Srivastava, K.; Geliebter, J.; Tiwari, R.; Miao, M.; et al. A small molecule compound berberine as an orally active therapeutic candidate against COVID-19 and SARS: A computational and mechanistic study. FASEB J. 2021, 35, e21360. [Google Scholar] [CrossRef]
- Varghese, F.; van Woudenbergh, E.; Overheul, G.; Eleveld, M.; Kurver, L.; van Heerbeek, N.; van Laarhoven, A.; Miesen, P.; Hartog, G.D.; de Jonge, M.; et al. Berberine and Obatoclax Inhibit SARS-CoV-2 Replication in Primary Human Nasal Epithelial Cells In Vitro. Viruses 2021, 13, 282. [Google Scholar] [CrossRef]
- Miclot, T.; Hognon, C.; Bignon, E.; Terenzi, A.; Marazzi, M.; Barone, G.; Monari, A. Structure and Dynamics of RNA Guanine Quadruplexes in SARS-CoV-2 Genome. Original Strategies against Emerging Viruses. J. Phys. Chem. Lett. 2021, 12, 10277–10283. [Google Scholar] [CrossRef]
- Arora, A.; Balasubramanian, C.; Kumar, N.; Agrawal, S.; Ojha, R.P.; Maiti, S. Binding of Berberine to Human Telomeric Quadruplex—Spectroscopic, Calorimetric and Molecular Modeling Studies: Telomeric Quadruplex-Berberine Interaction. FEBS J. 2008, 275, 3971–3983. [Google Scholar] [CrossRef]
- Bazzicalupi, C.; Ferraroni, M.; Bilia, A.R.; Scheggi, F.; Gratteri, P. The crystal structure of human telomeric DNA complexed with berberine: An interesting case of stacked ligand to G-tetrad ratio higher than 1:1. Nucleic Acids Res. 2013, 41, 632–638. [Google Scholar] [CrossRef]
- Sharma, N.K.; Lunawat, P.; Dixit, M. Binding Studies of Natural Product Berberine with DNA G-Quadruplex. Am. J. Biochem. Biotechnol. 2011, 7, 130–134. [Google Scholar] [CrossRef]
- Connors, K.A. Binding Constants: The Measurement of Molecular Complex Stability; Wiley: New York, NY, USA, 1987; ISBN 978-0-471-83083-2. [Google Scholar]
- Ulatowski, F.; Dabrowa, K.; Bałakier, T.; Jurczak, J. Recognizing the Limited Applicability of Job Plots in Studying Host–Guest Interactions in Supramolecular Chemistry. J. Org. Chem. 2016, 81, 1746–1756. [Google Scholar] [CrossRef]
- Zhang, W.-J.; Ou, T.-M.; Lu, Y.-J.; Huang, Y.-Y.; Wu, W.-B.; Huang, Z.-S.; Zhou, J.-L.; Wong, K.-Y.; Gu, L.-Q. 9-Substituted berberine derivatives as G-quadruplex stabilizing ligands in telomeric DNA. Bioorganic Med. Chem. 2007, 15, 5493–5501. [Google Scholar] [CrossRef]
- Wickhorst, P.; Ihmels, H. Berberrubine Phosphate: A Selective Fluorescent Probe for Quadruplex DNA. Molecules 2021, 26, 2566. [Google Scholar] [CrossRef]
- Oliva, R.; Banerjee, S.; Cinar, H.; Ehrt, C.; Winter, R. Alteration of Protein Binding Affinities by Aqueous Two-Phase Systems Revealed by Pressure Perturbation. Sci. Rep. 2020, 10, 8074. [Google Scholar] [CrossRef]
- Del Villar-Guerra, R.; Trent, J.O.; Chaires, J.B. G-Quadruplex Secondary Structure Obtained from Circular Dichroism Spectroscopy. Angew. Chem. Int. Ed. 2018, 57, 7171–7175. [Google Scholar] [CrossRef]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2005, 1751, 119–139. [Google Scholar] [CrossRef]
- Oliva, R.; Mukherjee, S.; Winter, R. Unraveling the binding characteristics of small ligands to telomeric DNA by pressure modulation. Sci. Rep. 2021, 11, 9714. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.K.; Knop, J.; Winter, R. Modulation of the Conformational Space of SARS-CoV-2 RNA Quadruplex RG-1 by Cellular Components and the Amyloidogenic Peptides α-Synuclein and hIAPP. Chem. A Eur. J. 2022, 28, e202104182. [Google Scholar] [CrossRef] [PubMed]
- Oliva, R.; Jahmidi-Azizi, N.; Mukherjee, S.; Winter, R. Harnessing Pressure Modulation for Exploring Ligand Binding Reac-tions in Cosolvent Solutions. J. Phys. Chem. B 2021, 125, 539–546. [Google Scholar] [CrossRef] [PubMed]
- De Rache, A.; Mergny, J.-L. Assessment of selectivity of G-quadruplex ligands via an optimised FRET melting assay. Biochimie 2015, 115, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.D.; Trent, J.O.; Chaires, J.B. Folding and Unfolding Pathways of the Human Telomeric G-Quadruplex. J. Mol. Biol. 2014, 426, 1629–1650. [Google Scholar] [CrossRef] [Green Version]
- Moraca, F.; Amato, J.; Ortuso, F.; Artese, A.; Pagano, B.; Novellino, E.; Alcaro, S.; Parrinello, M.; Limongelli, V. Ligand binding to telomeric G-quadruplex DNA investigated by funnel-metadynamics simulations. Proc. Natl. Acad. Sci. USA 2017, 114, E2136–E2145. [Google Scholar] [CrossRef] [Green Version]
- Chaires, J.B. Human Telomeric G-Quadruplex: Thermodynamic and Kinetic Studies of Telomeric Quadruplex Stability: Te-lomeric Quadruplex Stability. FEBS J. 2010, 277, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.Y.; Shek, Y.L.; Amiri, A.; Dubins, D.N.; Heerklotz, H.; Macgregor, J.R.B.; Chalikian, T.V. Volumetric Characterization of Sodium-Induced G-Quadruplex Formation. J. Am. Chem. Soc. 2011, 133, 4518–4526. [Google Scholar] [CrossRef]
- Son, I.; Shek, Y.L.; Dubins, D.N.; Chalikian, T.V. Hydration Changes Accompanying Helix-to-Coil DNA Transitions. J. Am. Chem. Soc. 2014, 136, 4040–4047. [Google Scholar] [CrossRef]
- Chalikian, T.V.; Macgregor, R.B. On empirical decomposition of volumetric data. Biophys. Chem. 2019, 246, 8–15. [Google Scholar] [CrossRef]
- Liu, L.; Scott, L.; Tariq, N.; Kume, T.; Dubins, D.N.; Macgregor, R.B.; Chalikian, T.V. Volumetric Interplay between the Con-formational States Adopted by Guanine-Rich DNA from the c-MYC Promoter. J. Phys. Chem. B 2021, 125, 7406–7416. [Google Scholar] [CrossRef]
- Mohanty, J.; Barooah, N.; Dhamodharan, V.; Harikrishna, S.; Pradeepkumar, P.I.; Bhasikuttan, A.C. Thioflavin T as an Efficient Inducer and Selective Fluorescent Sensor for the Human Telomeric G-Quadruplex DNA. J. Am. Chem. Soc. 2013, 135, 367–376. [Google Scholar] [CrossRef]
- Kibbe, W.A. OligoCalc: An online oligonucleotide properties calculator. Nucleic Acids Res. 2007, 35, W43–W46. [Google Scholar] [CrossRef]
- Díaz, M.S.; Freile, M.L.; Gutiérrez, M.I. Solvent effect on the UV/Vis absorption and fluorescence spectroscopic properties of berberine. Photochem. Photobiol. Sci. 2009, 8, 970–974. [Google Scholar] [CrossRef]
- Lee, J.; Vogt, C.E.; McBrairty, M.; Al-Hashimi, H.M. Influence of Dimethylsulfoxide on RNA Structure and Ligand Binding. Anal. Chem. 2013, 85, 9692–9698. [Google Scholar] [CrossRef] [Green Version]
- Oliva, R.; Battista, F.; Cozzolino, S.; Notomista, E.; Winter, R.; Del Vecchio, P.; Petraccone, L. Encapsulating properties of sulfobutylether-β-cyclodextrin toward a thrombin-derived antimicrobial peptide. J. Therm. Anal. Calorim. 2019, 138, 3249–3256. [Google Scholar] [CrossRef]
- Patra, S.; Schuabb, V.; Kiesel, I.; Knop, J.-M.; Oliva, R.; Winter, R. Exploring the effects of cosolutes and crowding on the volumetric and kinetic profile of the conformational dynamics of a poly dA loop DNA hairpin: A single-molecule FRET study. Nucleic Acids Res. 2019, 47, 981–996. [Google Scholar] [CrossRef]
- Brynn Hibbert, D.; Thordarson, P. The Death of the Job Plot, Transparency, Open Science and Online Tools, Uncertainty Estimation Methods and Other Developments in Supramolecular Chemistry Data Analysis. Chem. Commun. 2016, 52, 12792–12805. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Kb/M−1 ∙ 105 | 1 ΔG°b /kJ mol−1 | 2 ΔH°b /kJ mol−1 | 3 TΔS°b /kJ mol−1 | 4 Berb:G4Q |
|---|---|---|---|---|---|
| Berb/22AG in NaCl buffer | 0.37 ± 0.09 | −26.0 ± 0.6 | −15.5 ± 2.8 | 10.5 ± 3.4 | 2:1 |
| Berb/22AG in KCl buffer | 5.9 ± 1.0 | −32.9 ± 0.4 | 19.5 ± 3.8 | 52.4 ± 4.2 | 2:1 |
| Berb/RG-1 in NaCl buffer | 0.28 ± 0.03 | −25.4 ± 0.3 | 75.9 ± 7.6 | 101.3 ± 7.2 | 2:1 |
| Berb/RG-1 in KCl buffer | 0.14 ± 0.03 | −23.7 ± 0.4 | −92.2 ± 12.0 | −66.9 ± 8.8 | 2:1 |
| Berb/22AG NaCl (100 mM) | Berb/22AG KCl (100 mM) | ||||
| p/bar | Kb/M−1 ∙ 105 | 1 Berb:G4Q | p/bar | Kb/M−1 ∙ 105 | 1 Berb:G4Q |
| 1 | 0.37 ± 0.09 | 2:1 | 1 | 5.9 ± 1.0 | 2:1 |
| 500 | 0.39 ± 0.07 | 2:1 | 500 | 6.6 ± 0.9 | 2:1 |
| 1000 | 0.44 ± 0.06 | 2:1 | 1000 | 6.9 ± 0.7 | 2:1 |
| 1500 | 0.43 ± 0.09 | 3:1 | 1500 | 7.1 ± 0.4 | 2:1 |
| 1900 | 0.50 ± 0.09 | 3:1 | 1900 | 7.3 ± 0.5 | 2:1 |
| Berb/RG-1 NaCl (100 mM) | Berb/RG-1 KCl (100 mM) | ||||
| p/bar | Kb/M−1 ∙ 105 | 1 Berb:G4Q | p/bar | Kb/M−1 ∙ 105 | 1 Berb:G4Q |
| 1 | 0.28 ± 0.03 | 2:1 | 1 | 0.14 ± 0.03 | 2:1 |
| 500 | 0.38 ± 0.05 | 2:1 | 500 | 0.17 ± 0.03 | 2:1 |
| 1000 | 0.49 ± 0.06 | 2:1 | 1000 | 0.23 ± 0.02 | 2:1 |
| 1500 | 0.63 ± 0.07 | 2:1 | 1500 | 0.31 ± 0.01 | 2:1 |
| 1900 | 0.77 ± 0.09 | 2:1 | 1900 | 0.40 ± 0.06 | 2:1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliva, R.; Mukherjee, S.; Manisegaran, M.; Campanile, M.; Del Vecchio, P.; Petraccone, L.; Winter, R. Binding Properties of RNA Quadruplex of SARS-CoV-2 to Berberine Compared to Telomeric DNA Quadruplex. Int. J. Mol. Sci. 2022, 23, 5690. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105690
Oliva R, Mukherjee S, Manisegaran M, Campanile M, Del Vecchio P, Petraccone L, Winter R. Binding Properties of RNA Quadruplex of SARS-CoV-2 to Berberine Compared to Telomeric DNA Quadruplex. International Journal of Molecular Sciences. 2022; 23(10):5690. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105690
Chicago/Turabian StyleOliva, Rosario, Sanjib Mukherjee, Magiliny Manisegaran, Marco Campanile, Pompea Del Vecchio, Luigi Petraccone, and Roland Winter. 2022. "Binding Properties of RNA Quadruplex of SARS-CoV-2 to Berberine Compared to Telomeric DNA Quadruplex" International Journal of Molecular Sciences 23, no. 10: 5690. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105690