
Current Insights on the Impact of Proteomics in Respiratory Allergies

 , , ,
, , ,
Abstract
:1. Introduction
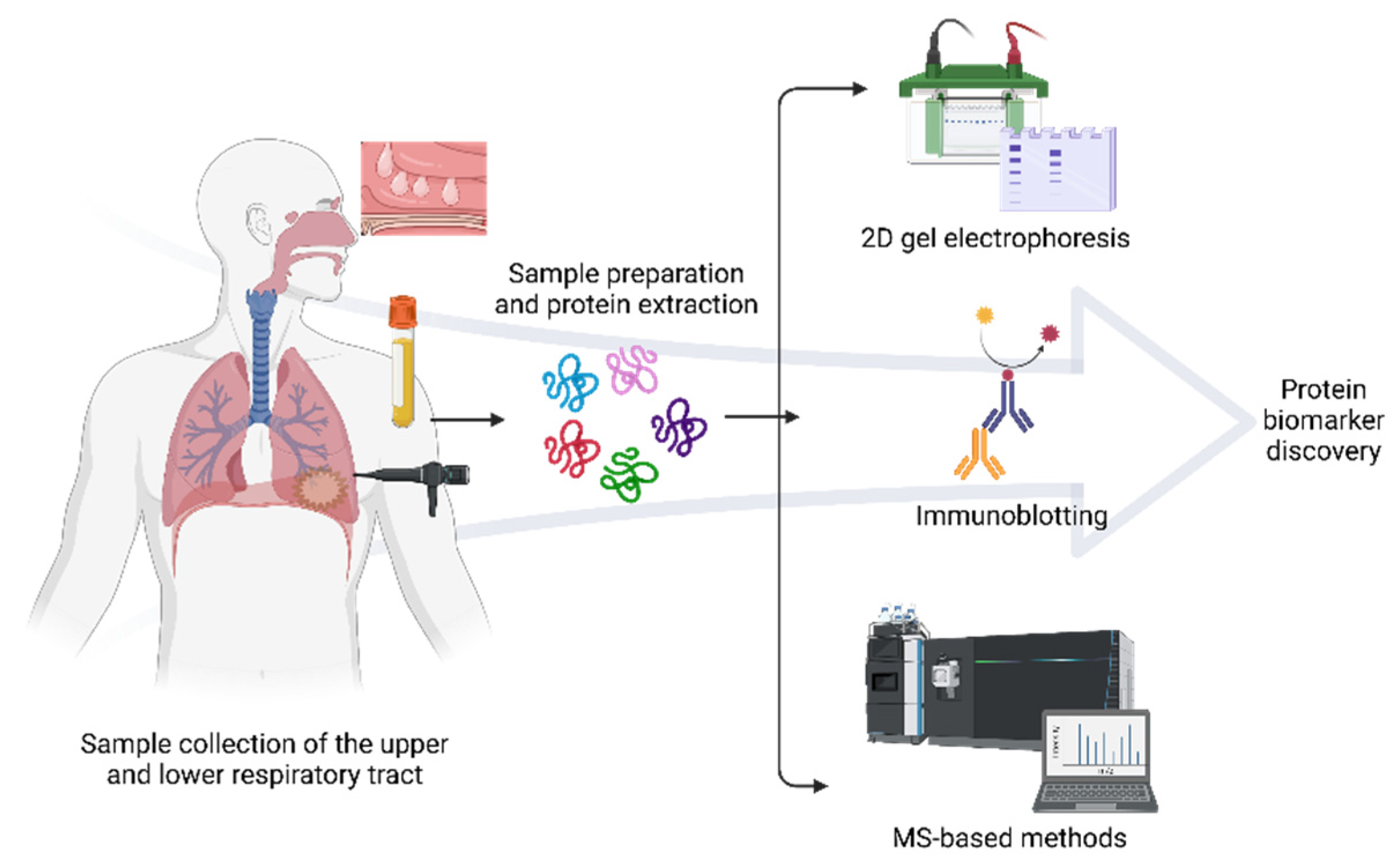
2. MS-Based Proteomics
3. Proteomics Approaches in Respiratory Allergy
3.1. Upper Respiratory
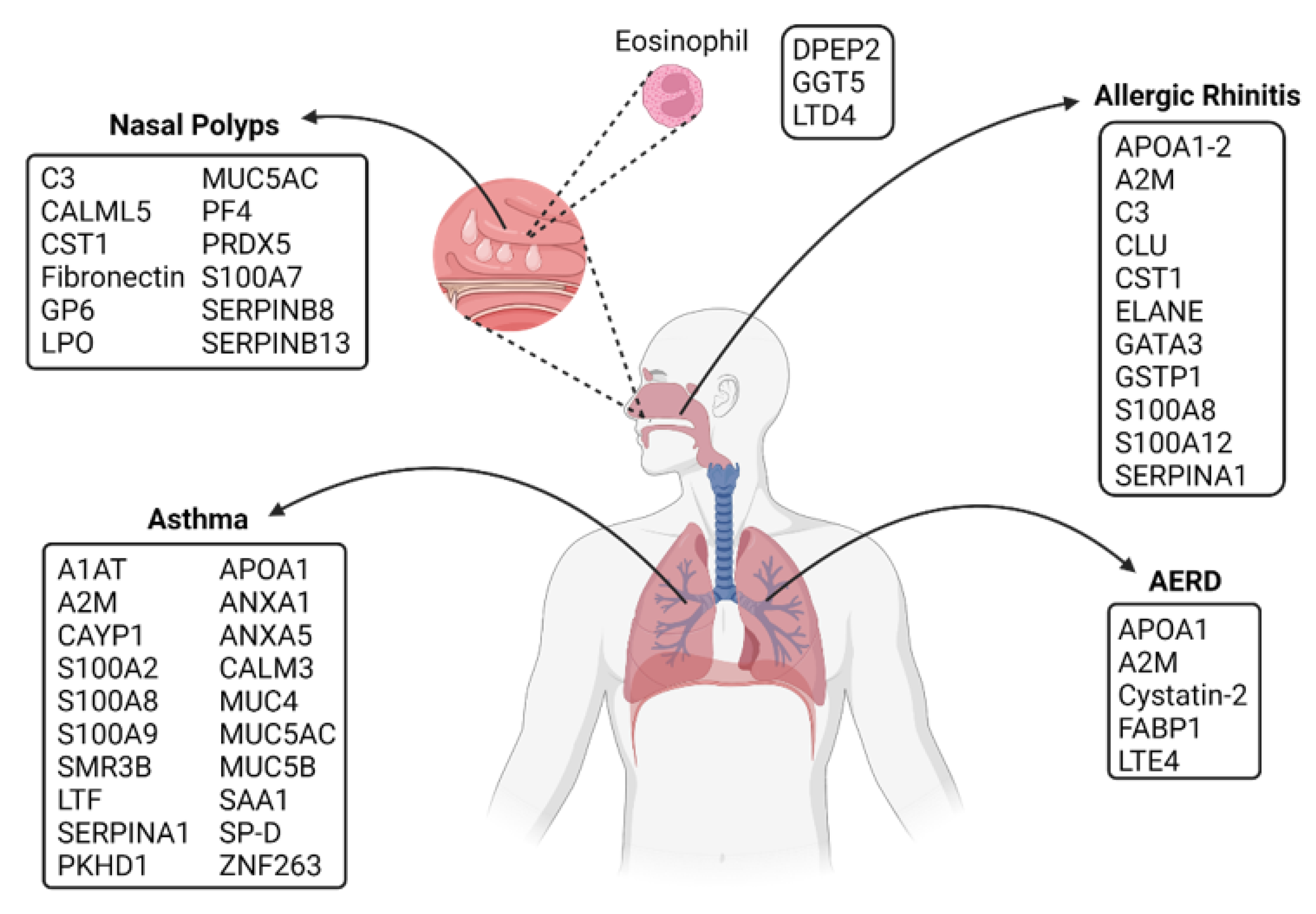
3.1.1. Allergic Rhinitis (AR)
3.1.2. Nasal Polyps (NPs)
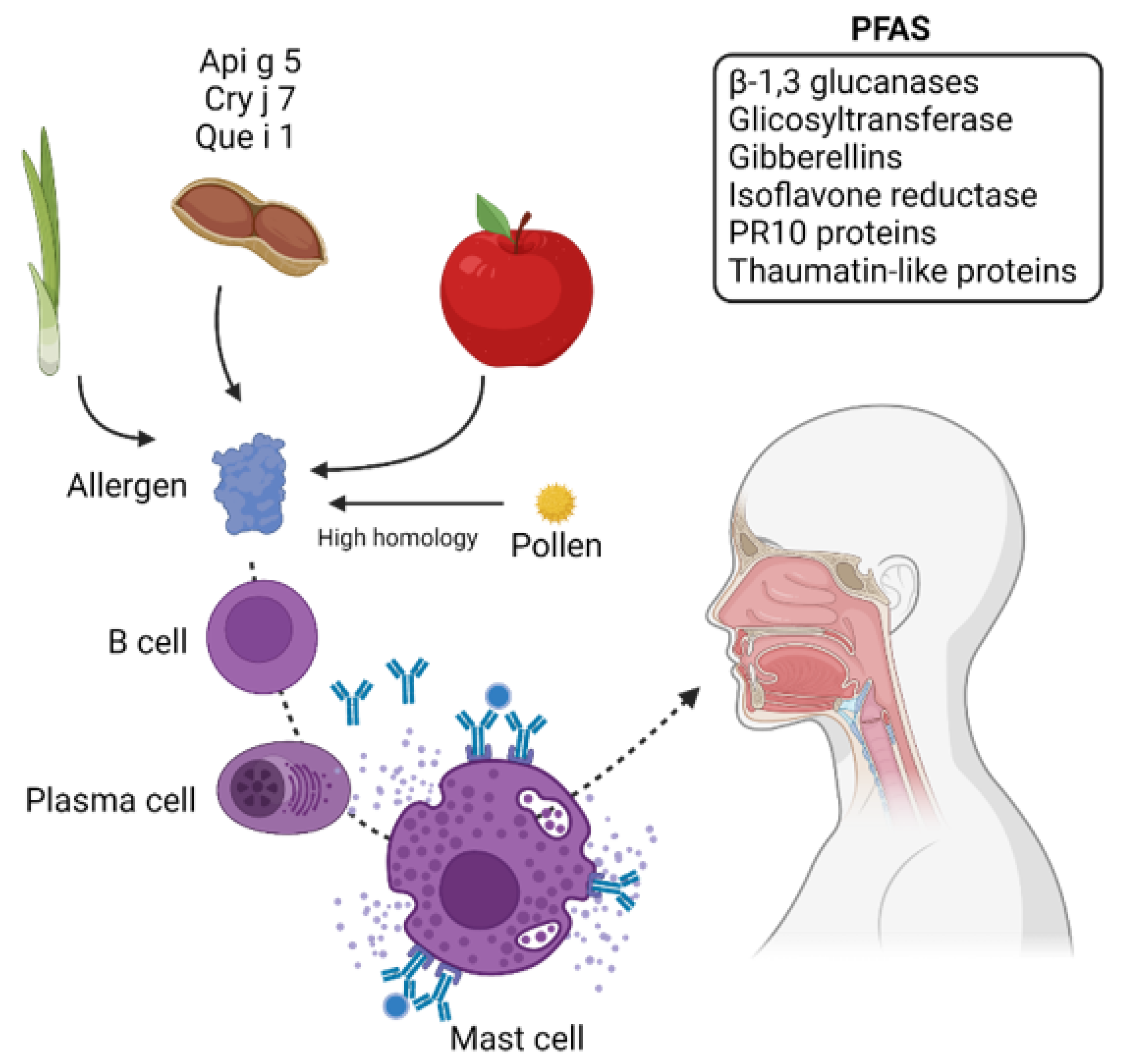
3.1.3. Pollen Food Allergy Syndrome (PFAS)
3.2. Lower Respiratory
3.2.1. Asthma
3.2.2. Aspirin Exacerbated Respiratory Disease (AERD)
4. Proteomics in Allergy Diagnostics and Therapeutics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, Y.-G.; Lee, P.-H.; Choi, S.-M.; An, M.-H.; Jang, A.-S. Effects of Air Pollutants on Airway Diseases. Int. J. Environ. Res. Public Health 2021, 18, 9905. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.-H.; Park, S.; Lee, Y.-G.; Choi, S.-M.; An, M.-H.; Jang, A.-S. The Impact of Environmental Pollutants on Barrier Dysfunction in Respiratory Disease. Allergy Asthma Immunol. Res. 2021, 13, 850. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, G.; Pawankar, R.; Vitale, C.; Lanza, M.; Molino, A.; Stanziola, A.; Sanduzzi, A.; Vatrella, A.; D’Amato, M. Climate Change and Air Pollution: Effects on Respiratory Allergy. Allergy Asthma Immunol. Res. 2016, 8, 391–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, F.-D. Cross-reactivity between aeroallergens and food allergens. World J. Methodol. 2015, 5, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Vaillant, A.A.J.; Vashisht, R.; Zito, P.M. Immediate Hypersensitivity Reactions. StatPearls 2021. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK513315/ (accessed on 10 May 2022).
- Alexovič, M.; Urban, P.L.; Tabani, H.; Sabo, J. Recent advances in robotic protein sample preparation for clinical analysis and other biomedical applications. Clin. Chim. Acta 2020, 507, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, W.; Chen, J. Recent progress in mass spectrometry proteomics for biomedical research. Sci. China Life Sci. 2017, 60, 1093–1113. [Google Scholar] [CrossRef]
- Aslam, B.; Basit, M.; Nisar, M.A.; Khurshid, M.; Rasool, M.H. Proteomics: Technologies and Their Applications. J. Chromatogr. Sci. 2017, 55, 182–196. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-S.; Pinto, S.M.; Getnet, D.; Nirujogi, R.S.; Manda, S.S.; Chaerkady, R.; Madugundu, A.K.; Kelkar, D.S.; Isserlin, R.; Jain, S.; et al. A draft map of the human proteome. Nature 2014, 509, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Overall, C.M. The Human Proteome: 90% in the Light, 10% on the Dark Side. J. Proteome Res. 2020, 19, 4731–4734. [Google Scholar] [CrossRef]
- Lössl, P.; Van De Waterbeemd, M.; Heck, A.J. The diverse and expanding role of mass spectrometry in structural and molecular biology. EMBO J. 2016, 35, 2634–2657. [Google Scholar] [CrossRef]
- Fenn, J.B.; Mann, M.; Meng, C.K.; Wong, S.F.; Whitehouse, C.M. Electrospray Ionization for Mass Spectrometry of Large Biomolecules. Science 1989, 246, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Waki, H.; Ido, Y.; Akita, S.; Yoshida, Y.; Yoshida, T.; Matsuo, T. Protein and polymer analyses up tom/z 100 000 by laser ionization time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 1988, 2, 151–153. [Google Scholar] [CrossRef]
- Fujii, K.; Nakamura, H.; Nishimura, T. Recent mass spectrometry-based proteomics for biomarker discovery in lung cancer, COPD, and asthma. Expert Rev. Proteom. 2017, 14, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Murray, K.K. Glossary of terms for separations coupled to mass spectrometry. J. Chromatogr. A 2010, 1217, 3922–3928. [Google Scholar] [CrossRef]
- Rockwood, A.L. The Expanding Role of Mass Spectrometry in Biotechnology. Gary Siuzdak. San Diego, CA: MCC Press, 2003, 286 pp., $49.00, softcover. ISBN 0-9742451-0-0. Clin. Chem. 2004, 50, 1108–1109. [Google Scholar] [CrossRef] [Green Version]
- Brewis, I.A.; Brennan, P. Proteomics technologies for the global identification and quantification of proteins. Adv. Protein Chem. Struct. Biol. 2010, 80, 1–44. [Google Scholar] [CrossRef]
- Marcus, K.; Lelong, C.; Rabilloud, T. What Room for Two-Dimensional Gel-Based Proteomics in a Shotgun Proteomics World? Proteomes 2020, 8, 17. [Google Scholar] [CrossRef]
- Durbin, K.R.; Fornelli, L.; Fellers, R.T.; Doubleday, P.F.; Narita, M.; Kelleher, N.L. Quantitation and Identification of Thousands of Human Proteoforms below 30 kDa. J. Proteome Res. 2016, 15, 976–982. [Google Scholar] [CrossRef] [Green Version]
- Fornelli, L.; Toby, T.K.; Schachner, L.F.; Doubleday, P.F.; Srzentić, K.; DeHart, C.J.; Kelleher, N.L. Top-down Proteomics: Where We Are, Where We Are Going? J. Proteom. 2018, 175, 3. [Google Scholar] [CrossRef]
- Zhang, Y.; Fonslow, B.R.; Shan, B.; Baek, M.-C.; Yates, J.R., 3rd. Protein Analysis by Shotgun/Bottom-up Proteomics. Chem. Rev. 2013, 113, 2343–2394. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Xu, Z.; Xu, X.; Zhao, X.; Huang, C.; Wei, Y. Quantitative Proteomics for Cancer Biomarker Discovery. Comb. Chem. High Throughput Screen. 2012, 15, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, A.; Geiger, T. Super-SILAC: Current trends and future perspectives. Expert Rev. Proteom. 2014, 12, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Fontarigo, J.J.; González-Barcala, F.J.; Andrade-Bulos, L.J.; San-José, M.E.; Cruz, M.J.; Valdés-Cuadrado, L.; Crujeiras, R.M.; Arias, P.; Salgado, F.J. iTRAQ-Based proteomic analysis reveals potential serum biomarkers of allergic and nonallergic asthma. Allergy 2020, 75, 3171–3183. [Google Scholar] [CrossRef]
- O’Connell, J.D.; Paulo, J.A.; O’Brien, J.J.; Gygi, S.P. Proteome-Wide Evaluation of Two Common Protein Quantification Methods. J. Proteome Res. 2018, 17, 1934–1942. [Google Scholar] [CrossRef] [PubMed]
- Priyadharshini, V.; Teran, L.M. Role of respiratory proteomics in precision medicine. Precis. Med. Investig. Pract. Provid. 2020, 1, 255–261. [Google Scholar] [CrossRef]
- Lindemann, C.; Thomanek, N.; Hundt, F.; Lerari, T.; Meyer, H.E.; Wolters, D.; Marcus, K. Strategies in relative and absolute quantitative mass spectrometry based proteomics. Biol. Chem. 2017, 398, 687–699. [Google Scholar] [CrossRef]
- Rabilloud, T.; Lelong, C. Two-dimensional gel electrophoresis in proteomics: A tutorial. J. Proteom. 2011, 74, 1829–1841. [Google Scholar] [CrossRef]
- Meleady, P. Two-Dimensional Gel Electrophoresis and 2D-DIGE. Differ. Gel Electrophor. 2017, 1664, 3–14. [Google Scholar] [CrossRef]
- Teran, L.M.; Montes-Vizuet, R.; Li, X.; Franz, T. Respiratory Proteomics: From Descriptive Studies to Personalized Medicine. J. Proteome Res. 2014, 14, 38–50. [Google Scholar] [CrossRef]
- Priyadharshini, V.S.; Teran, L.M. Personalized Medicine in Respiratory Disease: Role of Proteomics. Adv. Protein Chem. Struct. Biol. 2016, 102, 115–146. [Google Scholar]
- Nony, E.; Le Mignon, M.; Brier, S.; Martelet, A.; Moingeon, P. Proteomics for Allergy: From Proteins to the Patients. Curr. Allergy Asthma Rep. 2016, 16, 64. [Google Scholar] [CrossRef] [PubMed]
- Okubo, K.; Kurono, Y.; Ichimura, K.; Enomoto, T.; Okamoto, Y.; Kawauchi, H.; Suzaki, H.; Fujieda, S.; Masuyama, K. Japanese Guidelines for Allergic Rhinitis. Allergol. Int. 2020, 69, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Xu, R.; Xu, G. Recent Advances in Allergic Rhinitis. J. Clin. Otorhinolaryngol. Head Neck Surg. 2015, 29, 202–206. [Google Scholar]
- Diamant, Z.; Boot, J.D.; Mantzouranis, E.; Flohr, R.; Sterk, P.J.; Gerth van Wijk, R. Biomarkers in Asthma and Allergic Rhinitis. Pulm. Pharmacol. Ther. 2010, 23, 468–481. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Gottfries, J.; Barrenäs, F.; Benson, M. Identification of Novel Biomarkers in Seasonal Allergic Rhinitis by Combining Proteomic, Multivariate and Pathway Analysis. PLoS ONE 2011, 6, e23563. [Google Scholar] [CrossRef] [Green Version]
- Sivaprasad, U.; Askew, D.J.; Ericksen, M.B.; Gibson, A.M.; Stier, M.T.; Brandt, E.; Bass, S.A.; Daines, M.O.; Chakir, J.; Stringer, K.F.; et al. A nonredundant role for mouse Serpinb3a in the induction of mucus production in asthma. J. Allergy Clin. Immunol. 2011, 127, 254–261.e1–6. [Google Scholar] [CrossRef] [Green Version]
- Kelly-Robinson, G.; Reihill, J.; Lundy, F.; McGarvey, L.; Lockhart, J.; Litherland, G.; Thornbury, K.; Martin, S. The Serpin Superfamily and Their Role in the Regulation and Dysfunction of Serine Protease Activity in COPD and Other Chronic Lung Diseases. Int. J. Mol. Sci. 2021, 22, 6351. [Google Scholar] [CrossRef]
- Turato, C.; Pontisso, P. SERPINB3 (Serpin Peptidase Inhibitor, Clade B (Ovalbumin), Member 3). Atlas Genet. Cytogenet. Oncol. Haematol. 2015, 19, 202–209. [Google Scholar] [CrossRef]
- Shamji, M.H.; Temblay, J.N.; Cheng, W.; Byrne, S.M.; Macfarlane, E.; Switzer, A.R.; Francisco, N.D.; Olexandra, F.; Jacubczik, F.; Durham, S.R.; et al. Antiapoptotic serine protease inhibitors contribute to survival of allergenic T H 2 cells. J. Allergy Clin. Immunol. 2017, 142, 569–581.e5. [Google Scholar] [CrossRef] [Green Version]
- Lü, F.X.; Esch, R.E. Novel nasal secretion collection method for the analysis of allergen specific antibodies and inflammatory biomarkers. J. Immunol. Methods 2010, 356, 6–17. [Google Scholar] [CrossRef]
- Zorampari, C.; Prakash, A.; Rehan, H.S.; Gupta, L.K. Serum dipeptidyl peptidase-4 and eosinophil cationic protein levels in patients of bronchial asthma. Pulm. Pharmacol. Ther. 2021, 72, 102109. [Google Scholar] [CrossRef] [PubMed]
- Rydell, N.; Nagao, M.; Movérare, R.; Ekoff, H.; Sjölander, A.; Borres, M.P.; Fujisawa, T. Serum Eosinophilic Cationic Protein Is a Reliable Biomarker for Childhood Asthma. Int. Arch. Allergy Immunol. 2022, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Poposki, J.A.; Klingler, A.I.; Stevens, W.W.; Suh, L.A.; Tan, B.K.; Peters, A.T.; Abdala-Valencia, H.; Grammer, L.C.; Welch, K.C.; Smith, S.S.; et al. Elevation of activated neutrophils in chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2021, 149, 1666–1674. [Google Scholar] [CrossRef] [PubMed]
- London, N.R.; Ramanathan, M. The Role of the Sinonasal Epithelium in Allergic Rhinitis. Otolaryngol. Clin. N. Am. 2017, 50, 1043–1050. [Google Scholar] [CrossRef]
- Husna, S.M.N.; Tan, H.-T.T.; Shukri, N.M.; Ashari, N.S.M.; Wong, K.K. Nasal Epithelial Barrier Integrity and Tight Junctions Disruption in Allergic Rhinitis: Overview and Pathogenic Insights. Front. Immunol. 2021, 12, 1659. [Google Scholar] [CrossRef]
- Tomazic, P.V.; Birner-Gruenberger, R.; Leitner, A.; Obrist, B.; Spoerk, S.; Lang-Loidolt, D. Nasal mucus proteomic changes reflect altered immune responses and epithelial permeability in patients with allergic rhinitis. J. Allergy Clin. Immunol. 2014, 133, 741–750. [Google Scholar] [CrossRef]
- Tomazic, P.V.; Birner-Gruenberger, R.; Leitner, A.; Darnhofer, B.; Spoerk, S.; Lang-Loidolt, D. Apolipoproteins have a potential role in nasal mucus of allergic rhinitis patients: A proteomic study. Laryngoscope 2014, 125, E91–E96. [Google Scholar] [CrossRef]
- Tomazic, P.V.; Birner-Gruenberger, R.; Leitner, A.; Spoerk, S.; Lang-Loidolt, D. Seasonal proteome changes of nasal mucus reflect perennial inflammatory response and reduced defence mechanisms and plasticity in allergic rhinitis. J. Proteom. 2016, 133, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Zhong, J.; Liao, W.; Hu, R.; Chen, L.; Wu, X.; Liu, S. iTRAQ-Based proteomic analysis reveals potential regulatory networks in dust mite-related asthma treated with subcutaneous allergen immunotherapy. Mol. Med. Rep. 2020, 22, 3607–3620. [Google Scholar] [CrossRef]
- Suárez-Lorenzo, I.; De Castro, F.R.; Cruz-Niesvaara, D.; Herrera-Ramos, E.; Rodríguez-Gallego, C.; Carrillo-Diaz, T. Alpha 1 antitrypsin distribution in an allergic asthmatic population sensitized to house dust mites. Clin. Transl. Allergy 2018, 8, 44. [Google Scholar] [CrossRef]
- Miravitlles, M.; Vilà, S.; Torrella, M.; Balcells, E.; Rodríguez-Frías, F.; LA Roza, C.; Jardí, R.; Vidal, R. Influence of deficient α1-anti-trypsin phenotypes on clinical characteristics and severity of asthma in adults. Respir. Med. 2002, 96, 186–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khirfan, G.; Li, M.; Wang, X.; DiDonato, J.A.; Dweik, R.A.; Heresi, G.A. Abnormal levels of apolipoprotein A-I in chronic thromboembolic pulmonary hypertension. Pulm. Circ. 2021, 11, 20458940211010371. [Google Scholar] [CrossRef] [PubMed]
- Trakaki, A.; Marsche, G. High-Density Lipoprotein (HDL) in Allergy and Skin Diseases: Focus on Immunomodulating Functions. Biomedicines 2020, 8, 558. [Google Scholar] [CrossRef] [PubMed]
- Wittenberg, M.; Nassiri, M.; Francuzik, W.; Lehmann, K.; Babina, M.; Worm, M. Serum levels of 9α,11β-PGF2 and apolipoprotein A1 achieve high predictive power as biomarkers of anaphylaxis. Allergy 2017, 72, 1801–1805. [Google Scholar] [CrossRef] [Green Version]
- Preianò, M.; Maggisano, G.; Murfuni, M.S.; Villella, C.; Colica, C.; Fregola, A.; Pelaia, C.; Lombardo, N.; Pelaia, G.; Savino, R.; et al. Rapid Detection and Identification of Antimicrobial Peptide Fingerprints of Nasal Fluid by Mesoporous Silica Particles and MALDI-TOF/TOF Mass Spectrometry: From the Analytical Approach to the Diagnostic Applicability in Precision Medicine. Int. J. Mol. Sci. 2018, 19, 4005. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Xie, Z.; Lv, Y.; Tang, Q.; Zhang, H.; Zhang, J.Y.; Wu, B.; Jiang, W. A proteomics analysis reveals that A2M might be regulated by STAT3 in persistent allergic rhinitis. Clin. Exp. Allergy 2016, 46, 813–824. [Google Scholar] [CrossRef]
- Pullerits, T.; Rönmark, E.P.; Ekerljung, L.; Palmqvist, M.A.; Arvidsson, M.; Mincheva, R.; Backman, H.; Kankaanranta, H.; Ilmarinen, P.; Rådinger, M.; et al. The triad of current asthma, rhinitis and eczema is uncommon among adults: Prevalence, sensitization profiles, and risk factors. Respir. Med. 2020, 176, 106250. [Google Scholar] [CrossRef]
- Aguilar, D.; Pinart, M.; Koppelman, G.; Saeys, Y.; Nawijn, M.C.; Postma, D.S.; Akdis, M.; Auffray, C.; Ballereau, S.; Benet, M.; et al. Computational analysis of multimorbidity between asthma, eczema and rhinitis. PLoS ONE 2017, 12, e0179125. [Google Scholar] [CrossRef] [Green Version]
- Koch, S.; Sopel, N.; Finotto, S. Th9 and other IL-9-producing cells in allergic asthma. Semin. Immunopathol. 2016, 39, 55–68. [Google Scholar] [CrossRef]
- Ndika, J.; Airaksinen, L.; Suojalehto, H.; Karisola, P.; Fyhrquist, N.; Puustinen, A.; Alenius, H. Epithelial proteome profiling suggests the essential role of interferon-inducible proteins in patients with allergic rhinitis. J. Allergy Clin. Immunol. 2017, 140, 1288–1298. [Google Scholar] [CrossRef] [Green Version]
- Käck, U.; Einarsdottir, E.; van Hage, M.; Asarnoj, A.; James, A.; Nopp, A.; Krjutškov, K.; Katayama, S.; Kere, J.; Lilja, G.; et al. Nasal upregulation of CST1 in dog-sensitised children with severe allergic airway disease. ERJ Open Res. 2021, 7, 00917-02020. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.-T.; Cao, M.-D.; Yu, R.-L.; Shi, H.-Y.; Yan, W.-J.; Liu, J.-G.; Pan, C.; Sun, J.; Wei, Q.-Y.; Wang, D.-Y.; et al. Leukotriene A4 Hydrolase Is a Candidate Predictive Biomarker for Successful Allergen Immunotherapy. Front. Immunol. 2020, 11, 3003. [Google Scholar] [CrossRef] [PubMed]
- Di Cicco, M.E.; Bizzoco, F.; Morelli, E.; Seccia, V.; Ragazzo, V.; Peroni, D.G.; Comberiati, P. Nasal Polyps in Children: The Early Origins of a Challenging Adulthood Condition. Children 2021, 8, 997. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Yang, G.; Zhang, R.; Xu, G.; Zhang, L.; Wen, W.; Lu, J.; Liu, J.; Yu, Y. Altered microRNA Expression Profiles of Extracellular Vesicles in Nasal Mucus from Patients With Allergic Rhinitis. Allergy Asthma Immunol. Res. 2015, 7, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Tan, K.S.; Guan, W.-J.; Jiang, L.-J.; Deng, J.; Gao, W.-X.; Lee, Y.M.; Xu, Z.-F.; Luo, X.; Liu, C.; et al. Proteomics profiling of epithelium-derived exosomes from nasal polyps revealed signaling functions affecting cellular proliferation. Respir. Med. 2020, 162, 105871. [Google Scholar] [CrossRef]
- Wang, L.-F.; Lee, C.-H.; Liang, S.-S.; Hung, C.-C.; Wu, Y.-R.; Chien, C.-Y.; Chen, J.-F. Mucin 5AC is significantly upregulated in exosomes from the nasal lavage fluid and may promote the expression of COX-2, VEGF and MMP-9: An implication in nasal polyp pathogenesis. Rhinol. J. 2021, 59, 328–336. [Google Scholar] [CrossRef]
- Tajiri, T.; Matsumoto, H.; Jinnai, M.; Kanemitsu, Y.; Nagasaki, T.; Iwata, T.; Inoue, H.; Nakaji, H.; Oguma, T.; Ito, I.; et al. Pathophysiological relevance of sputum MUC5AC and MUC5B levels in patients with mild asthma. Allergol. Int. 2021, 71, 193–199. [Google Scholar] [CrossRef]
- Yan, D.; Ye, Y.; Zhang, J.; Zhao, J.; Yu, J.; Luo, Q. Human Neutrophil Elastase Induces MUC5AC Overexpression in Chronic Rhinosinusitis Through miR-146a. Am. J. Rhinol. Allergy 2019, 34, 59–69. [Google Scholar] [CrossRef]
- Mueller, S.; Ms, A.L.N.; Dillon, S.T.; Gu, X.; Wendler, O.; Otu, H.; Libermann, T.A.; Bleier, B.S. Noninvasive exosomal proteomic biosignatures, including cystatin SN, peroxiredoxin-5, and glycoprotein VI, accurately predict chronic rhinosinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2018, 9, 177–186. [Google Scholar] [CrossRef]
- Yan, Z.; Liu, L.; Jiao, L.; Wen, X.; Liu, J.; Wang, N. Bioinformatics Analysis and Identification of Underlying Biomarkers Potentially Linking Allergic Rhinitis and Asthma. Med. Sci. Monit. 2020, 26, e924934. [Google Scholar] [CrossRef]
- Wang, W.; He, Y.; Zhao, Q.; Zhao, X.; Li, Z. Identification of potential key genes in gastric cancer using bioinformatics analysis. Biomed. Rep. 2020, 12, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Liu, Y.; Zhang, K.; Chen, L. CST1 Promoted Gastric Cancer Migration and Invasion Through Activating Wnt Pathway. Cancer Manag. Res. 2021, 13, 1901–1907. [Google Scholar] [CrossRef] [PubMed]
- Szeliga, M. Comprehensive analysis of the expression levels and prognostic values of PRDX family genes in glioma. Neurochem. Int. 2021, 153, 105256. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zou, S.; Zhang, Y.; Lin, R.; Duan, Y.; He, W.; Yang, Z. Discovery of Antitumor Active Peptides Derived from Peroxiredoxin. ChemMedChem 2021, 16, 3477–3483. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhou, L.; Zhang, Q.; Qian, F.; Yin, K.; Huang, M.; Zhang, X. Increased RhoGDI2 and Peroxiredoxin 5 Levels in Asthmatic Murine Model of Β2-Adrenoceptor Desensitization: A Proteomics Approach. Chin. Med. J. 2008, 121, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.K.; Nocera, A.L.; Dillon, S.T.; Wu, D.; Libermann, T.A.; Bleier, B.S. Highly multiplexed proteomic analysis reveals significant tissue and exosomal coagulation pathway derangement in chronic rhinosinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2018, 8, 1438–1444. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, J.K.; Patel, K.; Williamson, T.; Reaves, N.; Carroll, W.; Stephenson, S.E.; Gao, P.; Drake, R.R.; Neely, B.A.; Tomlinson, S.; et al. C3a receptor antagonism as a novel therapeutic target for chronic rhinosinusitis. Mucosal Immunol. 2018, 11, 1375–1385. [Google Scholar] [CrossRef] [Green Version]
- Miyata, J.; Fukunaga, K.; Kawashima, Y.; Watanabe, T.; Saitoh, A.; Hirosaki, T.; Araki, Y.; Kikawada, T.; Betsuyaku, T.; Ohara, O.; et al. Dysregulated fatty acid metabolism in nasal polyp-derived eosinophils from patients with chronic rhinosinusitis. Allergy 2019, 74, 1113–1124. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Han, D.; Kim, J.; Kim, D.W.; Kim, Y.-M.; Mo, J.-H.; Choi, H.-G.; Park, J.-W.; Shin, H.-W. In-Depth, Proteomic Analysis of Nasal Secretions from Patients with Chronic Rhinosinusitis and Nasal Polyps. Allergy Asthma Immunol. Res. 2019, 11, 691–708. [Google Scholar] [CrossRef]
- Chen, T.-Y.; Zhou, M.; Lin, M.-Q.; Liang, S.-T.; Yan, Y.; Wang, S.-M.; Fang, C.-S.; Li, D.; Ruan, Y. Research Progress on the SERPINE1 Protein and Chronic Inflammatory Diseases of the Upper Respiratory Tract: A Literature Review. Int. Arch. Allergy Immunol. 2021, 182, 1097–1102. [Google Scholar] [CrossRef]
- Sumsion, J.S.; Pulsipher, A.; Alt, J.A. Differential expression and role of S100 proteins in chronic rhinosinusitis. Curr. Opin. Allergy Clin. Immunol. 2020, 20, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Workman, A.D.; Miyake, M.M.; Ms, A.L.N.; Mueller, S.; Finn, K.; Otu, H.H.; Libermann, T.A.; Bleier, B.S. Unexpected effects of systemic steroids on the CRSwNP proteome: Is protein upregulation more important than inhibition? Int. Forum Allergy Rhinol. 2020, 10, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Lin, X.-S.; Yao, D.-M.; Zhuang, Y.-Y.; Wen, G.-F.; Shi, J.; Sun, Y.-Q. Increased serum amyloid A in nasal polyps is associated with systemic corticosteroid insensitivity in patients with chronic rhinosinusitis with nasal polyps: A pilot study. Eur. Arch. Oto-Rhino-Laryngol. 2017, 275, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Ta, V.; Chin, W.K.; Wineinger, N.E.; Scott, D.R.; Kelso, J.M.; White, A.A. Differential skin test reactivity to pollens in pollen food allergy syndrome versus allergic rhinitis. Allergy Asthma Proc. 2015, 36, 379–385. [Google Scholar] [CrossRef]
- Carlson, G.; Coop, C. Pollen food allergy syndrome (PFAS): A review of current available literature. Ann. Allergy Asthma Immunol. 2019, 123, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Poncet, P.; Sénéchal, H.; Charpin, D. Update on Pollen-Food Allergy Syndrome. Expert Rev. Clin. Immunol. 2020, 16, 561–578. [Google Scholar] [CrossRef]
- Bedolla-Barajas, M.; Kestler-Gramajo, A.; Alcalá-Padilla, G.; Morales-Romero, J. Prevalence of oral allergy syndrome in children with allergic diseases. Allergol. Immunopathol. 2017, 45, 127–133. [Google Scholar] [CrossRef]
- Borghesan, F.; Mistrello, G.; Amato, S.; Giuffrida, M.G.; Villalta, D.; Asero, R. Mugwort-fennel-allergy-syndrome associated with sensitization to an allergen homologous to Api g 5. Eur. Ann. Allergy Clin. Immunol. 2013, 45, 130–137. [Google Scholar]
- Biagioni, B.; Tomei, L.; Valleriani, C.; Liccioli, G.; Barni, S.; Sarti, L.; Citera, F.; Giovannini, M.; Mori, F. Allergy to Gibberellin-Regulated Proteins (Peamaclein) in Children. Int. Arch. Allergy Immunol. 2021, 182, 1194–1199. [Google Scholar] [CrossRef]
- Asero, R.; Abbadessa, S.; Aruanno, A.; Barilaro, G.; Barzaghi, C.; Bignardi, D.; Bilò, M.; Borro, M.; Bresciani, M.; Busa, M.; et al. Detection of Gibberellin-Regulated Protein (Peamaclein) Sensitization among Italian Cypress Pollen-Sensitized Patients. J. Investig. Allergy Clin. Immunol. 2021, 32, 40–47. [Google Scholar] [CrossRef]
- Mori, Y.; Okazaki, F.; Momma, K.; Narita, H.; Kondo, Y. Investigation of the sensitization rate for gibberellin-regulated protein in patients with Japanese cedar pollinosis. Allergol. Immunopathol. 2022, 50, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Klingebiel, C.; Chantran, Y.; Arif-Lusson, R.; Ehrenberg, A.E.; Östling, J.; Poisson, A.; Liabeuf, V.; Agabriel, C.; Birnbaum, J.; Porri, F.; et al. Pru p 7 sensitization is a predominant cause of severe, cypress pollen-associated peach allergy. Clin. Exp. Allergy 2019, 49, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Inomata, N.; Miyakawa, M.; Ikeda, N.; Oda, K.; Aihara, M. Identification of gibberellin-regulated protein as a new allergen in orange allergy. Clin. Exp. Allergy 2018, 48, 1509–1520. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, T.; Takei, M.; Saito, Y.; Rumi, F.; Zheng, J.; Lu, X.; Chafey, P.; Broussard, C.; Guilloux-Assalet, L.; Charpin, D.; et al. Gibberellin-regulated protein sensitization in Japanese cedar (Cryptomeria japonica) pollen allergic Japanese cohorts. Allergy 2021, 76, 2297–2302. [Google Scholar] [CrossRef]
- Carrón-Herrero, A.; Fernández-Lozano, C.; Peracho, L.; Martínez-Alonso, E.; Alcázar, A.; Martínez-Botas, J.; De-La-Hoz-Caballer, B.; González-De-Olano, D. Custard Apple Allergy with Glycosyltransferase as the Allergen Involved. J. Investig. Allergy Clin. Immunol. 2021, 32, 73–74. [Google Scholar] [CrossRef]
- Khan, I.M.; Ulrich, B.J.; Nelson, A.S.; Sehra, S.; Kansas, G.S.; Kaplan, M.H. Selectin Dependence of Allergic Skin Inflammation Is Diminished by Maternal Atopy. ImmunoHorizons 2021, 5, 703–710. [Google Scholar] [CrossRef]
- Pedrosa, M.; Guerrero-Sanchez, V.M.; Canales-Bueno, N.; Loli-Ausejo, D.; Castillejo, M.; Quirce, S.; Jorrin-Novo, J.V.; Rodriguez-Perez, R. Quercus ilexpollen allergen, Que i 1, responsible for pollen food allergy syndrome caused by fruits in Spanish allergic patients. Clin. Exp. Allergy 2020, 50, 815–823. [Google Scholar] [CrossRef]
- Zeindl, R.; Tollinger, M. NMR resonance assignments of the PR-10 allergens Act c 8 and Act d 8 from golden and green kiwifruit. Biomol. NMR Assign. 2021, 15, 367–371. [Google Scholar] [CrossRef]
- Bastiaan-Net, S.; Pérez, M.C.P.; Dekkers, B.J.W.; Westphal, A.H.; America, A.H.P.; Ariëns, R.M.C.; de Jong, N.W.; Wichers, H.; Mes, J.J. Identification and in silico bioinformatics analysis ofPR10proteins in cashew nut. Protein Sci. 2020, 29, 1581–1595. [Google Scholar] [CrossRef] [Green Version]
- Sim, S.; Choi, Y.; Park, H.-S. Potential Metabolic Biomarkers in Adult Asthmatics. Metabolites 2021, 11, 430. [Google Scholar] [CrossRef]
- Karaulov, A.V.; Garib, V.; Garib, F.; Valenta, R. Protein Biomarkers in Asthma. Int. Arch. Allergy Immunol. 2018, 175, 189–208. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Clark, T.J.H.; Hurd, S.; Khaltaev, N.; Lenfant, C.; O’Byrne, P.; Sheffer, A. GINA Guidelines on Asthma and Beyond. Allergy Eur. J. Allergy Clin. Immunol. 2007, 62, 102–112. [Google Scholar] [CrossRef] [PubMed]
- di Palmo, E.; Cantarelli, E.; Catelli, A.; Ricci, G.; Gallucci, M.; Miniaci, A.; Pession, A. The Predictive Role of Biomarkers and Genetics in Childhood Asthma Exacerbations. Int. J. Mol. Sci. 2021, 22, 4651. [Google Scholar] [CrossRef] [PubMed]
- Bich, T.C.T.; Quoc, Q.L.; Choi, Y.; Yang, E.-M.; Trinh, H.K.T.; Shin, Y.S.; Park, H.-S. Serum Amyloid A1: A Biomarker for Neutrophilic Airway Inflammation in Adult Asthmatic Patients. Allergy Asthma Immunol. Res. 2022, 14, 40–58. [Google Scholar] [CrossRef]
- Smole, U.; Gour, N.; Phelan, J.; Hofer, G.; Köhler, C.; Kratzer, B.; Tauber, P.A.; Xiao, X.; Yao, N.; Dvorak, J.; et al. Serum amyloid A is a soluble pattern recognition receptor that drives type 2 immunity. Nat. Immunol. 2020, 21, 756–765. [Google Scholar] [CrossRef]
- Li, M.; Wu, M.; Qin, Y.; Liu, H.; Tu, C.; Shen, B.; Xu, X.; Chen, H. Differentially expressed serum proteins in children with or without asthma as determined using isobaric tags for relative and absolute quantitation proteomics. PeerJ 2020, 8, e9971. [Google Scholar] [CrossRef]
- Weitoft, M.; Kadefors, M.; Stenberg, H.; Tufvesson, E.; Diamant, Z.; Enes, S.R.; Bjermer, L.; Rosmark, O.; Westergren-Thorsson, G. Plasma proteome changes linked to late phase response after inhaled allergen challenge in asthmatics. Respir. Res. 2022, 23, 50. [Google Scholar] [CrossRef]
- Gharib, S.A.; Nguyen, E.; Lai, Y.; Plampin, J.D.; Goodlett, D.R.; Hallstrand, T.S. Induced sputum proteome in healthy subjects and asthmatic patients. J. Allergy Clin. Immunol. 2011, 128, 1176–1184.e6. [Google Scholar] [CrossRef] [Green Version]
- Quoc, Q.L.; Choi, Y.; Bich, T.C.T.; Yang, E.-M.; Shin, Y.S.; Park, H.-S. S100A9 in adult asthmatic patients: A biomarker for neutrophilic asthma. Exp. Mol. Med. 2021, 53, 1170–1179. [Google Scholar] [CrossRef]
- Lee, T.-H.; Jang, A.-S.; Park, J.-S.; Kim, T.-H.; Choi, Y.S.; Shin, H.-R.; Park, S.-W.; Uh, S.-T.; Choi, J.-S.; Kim, Y.H.; et al. Elevation of S100 calcium binding protein A9 in sputum of neutrophilic inflammation in severe uncontrolled asthma. Ann. Allergy Asthma Immunol. 2013, 111, 268–275.e1. [Google Scholar] [CrossRef]
- Chen, M.; Ge, Y.; Lin, W.; Ying, H.; Zhang, W.; Yu, X.; Li, C.; Cao, C. Clinical features and nasal inflammation in asthma and allergic rhinitis. Clin. Exp. Immunol. 2022, 208, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Chakraborty, R.; Saha, B.; Suri, H.; Singh, P.; Raj, A.; Taneja, B.; Dash, D.; Sengupta, S.; Agrawal, A. Sputum Protein Biomarkers in Airway Diseases: A Pilot Study. Int. J. Chronic Obstr. Pulm. Dis. 2021, 16, 2203–2215. [Google Scholar] [CrossRef] [PubMed]
- Hastie, A.T.; Steele, C.; Dunaway, C.W.; Moore, W.C.; Rector, B.M.; Ampleford, E.; Li, H.; Denlinger, L.C.; Jarjour, N.; Meyers, D.A.; et al. Complex association patterns for inflammatory mediators in induced sputum from subjects with asthma. Clin. Exp. Allergy 2018, 48, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Coakley, R.C.; Mascenik, T.; Rowell, T.R.; Davis, E.S.; Rogers, K.; Webster, M.J.; Dang, H.; Herring, L.E.; Sassano, M.F.; et al. Chronic E-Cigarette Exposure Alters the Human Bronchial Epithelial Proteome. Am. J. Respir. Crit. Care Med. 2018, 198, 67–76. [Google Scholar] [CrossRef]
- Tarran, R.; Ghosh, A.; Alexis, N. Reply to Shields et Al. Am. J. Respir. Crit. Care Med. 2018, 198, 1351–1352. [Google Scholar] [CrossRef]
- Tong, J.; Gu, Q. Expression and Clinical Significance of Mucin Gene in Chronic Rhinosinusitis. Curr. Allergy Asthma Rep. 2020, 20, 63. [Google Scholar] [CrossRef]
- Ye, Y.; Zhao, J.; Ye, J.; Jiang, X.; Liu, H.; Xie, Y.; Zhang, J.; Luo, Q. The role of autophagy in the overexpression of MUC5AC in patients with chronic rhinosinusitis. Int. Immunopharmacol. 2019, 71, 169–180. [Google Scholar] [CrossRef]
- Li, C.-X.; Zhang, L.; Yan, Y.-R.; Ding, Y.-J.; Lin, Y.-N.; Zhou, J.-P.; Li, N.; Li, H.-P.; Li, S.-Q.; Sun, X.-W.; et al. A narrative review of exploring potential salivary biomarkers in respiratory diseases: Still on its way. J. Thorac. Dis. 2021, 13, 4541–4553. [Google Scholar] [CrossRef]
- Zamora-Mendoza, B.; Espinosa-Tanguma, R.; Ramírez-Elías, M.; Cabrera-Alonso, R.; Montero-Moran, G.; Portales-Pérez, D.; Rosales-Romo, J.; Gonzalez, J.; Gonzalez, C. Surface-enhanced raman spectroscopy: A non-invasive alternative procedure for early detection in childhood asthma biomarkers in saliva. Photodiagnosis Photodyn. Ther. 2019, 27, 85–91. [Google Scholar] [CrossRef]
- Okazaki, S.; Murai, H.; Kidoguchi, S.; Nomura, E.; Itoh, N.; Hashimoto, N.; Hamada, T.; Kawakita, A.; Yasutomi, M.; Ohshima, Y. The Biomarker Salivary SP-D May Indicate Small Airway Inflammation and Asthma Exacerbation. J. Investig. Allergy Clin. Immunol. 2017, 27, 305–312. [Google Scholar] [CrossRef]
- Mackay, R.-M.A.; Grainge, C.; Lau, L.C.; Barber, C.; Clark, H.W.; Howarth, P.H. Airway Surfactant Protein D Deficiency in Adults with Severe Asthma. Chest 2016, 149, 1165–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poachanukoon, O.; Roytrakul, S.; Koontongkaew, S. A shotgun proteomic approach reveals novel potential salivary protein biomarkers for asthma. J. Asthma 2020, 59, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Steinke, J.W.; Lawrence, M.G.; Teague, W.G.; Braciale, T.J.; Patrie, J.T.; Borish, L. Bronchoalveolar lavage cytokine patterns in children with severe neutrophilic and paucigranulocytic asthma. J. Allergy Clin. Immunol. 2021, 147, 686–693.e3. [Google Scholar] [CrossRef] [PubMed]
- O‘Neil, S.E.; Sitkauskiene, B.; Babusyte, A.; Krisiukeniene, A.; Stravinskaite-Bieksiene, K.; Sakalauskas, R.; Sihlbom, C.; Ekerljung, L.; Carlsohn, E.; Lötvall, J. Network analysis of quantitative proteomics on asthmatic bronchi: Effects of inhaled glucocorticoid treatment. Respir. Res. 2011, 12, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-H.; Lee, P.-H.; Kim, B.-G.; Hong, J.; Jang, A.-S. Annexin A5 Protein as a Potential Biomarker for the Diagnosis of Asthma. Lung 2018, 196, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Lee, P.-H.; Kim, B.-G.; Seo, H.-J.; Baek, A.-R.; Park, J.-S.; Lee, J.-H.; Park, S.-W.; Kim, D.-J.; Park, C.-S.; et al. Annexin A1 in plasma from patients with bronchial asthma: Its association with lung function. BMC Pulm. Med. 2018, 18, 1. [Google Scholar] [CrossRef] [Green Version]
- Bouter, A.; Carmeille, R.; Gounou, C.; Bouvet, F.; Degrelle, S.; Evain-Brion, D.; Brisson, A. Review: Annexin-A5 and cell membrane repair. Placenta 2015, 36, S43–S49. [Google Scholar] [CrossRef]
- Winter, N.A.; Gibson, P.G.; Fricker, M.; Simpson, J.L.; Wark, P.A.; McDonald, V.M. Hemopexin: A Novel Anti-inflammatory Marker for Distinguishing COPD From Asthma. Allergy Asthma Immunol. Res. 2021, 13, 450–467. [Google Scholar] [CrossRef]
- Landi, C.; Vantaggiato, L.; Shaba, E.; Cameli, P.; Carleo, A.; D’Alessandro, M.; Bergantini, L.; Bargagli, E.; Bini, L. Differential redox proteomic profiles of serum from severe asthma patients after one month of benralizumab and mepolizumab treatment. Pulm. Pharmacol. Ther. 2021, 70, 102060. [Google Scholar] [CrossRef]
- Vantaggiato, L.; Perruzza, M.; Refini, R.M.; Bergantini, L.; D’Alessandro, M.; Cameli, P.; Perruzza, D.; Bini, L.; Bargagli, E.; Landi, C. Mepolizumab and Benralizumab in Severe Eosinophilic Asthma: Preliminary Results of a Proteomic Study. Lung 2020, 198, 761–765. [Google Scholar] [CrossRef]
- Takahashi, K.; Pavlidis, S.; Kwong, F.N.K.; Hoda, U.; Rossios, C.; Sun, K.; Loza, M.; Baribaud, F.; Chanez, P.; Fowler, S.; et al. Sputum proteomics and airway cell transcripts of current and ex-smokers with severe asthma in U-BIOPRED: An exploratory analysis. Eur. Respir. J. 2018, 51, 1702173. [Google Scholar] [CrossRef] [PubMed]
- Brasier, A.; Victor, S.; Ju, H.; Busse, W.W.; Curran-Everett, U.; Bleecker, E.; Castro, M.; Chung, K.F.; Gaston, B.; Israel, E.; et al. Predicting Intermediate Phenotypes in Asthma Using Bronchoalveolar Lavage-Derived Cytokines. Clin. Transl. Sci. 2010, 3, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Brasier, A.; Victor, S.; Boetticher, G.; Ju, H.; Lee, C.; Bleecker, E.R.; Castro, M.; Busse, W.W.; Calhoun, W.J. Molecular phenotyping of severe asthma using pattern recognition of bronchoalveolar lavage–derived cytokines. J. Allergy Clin. Immunol. 2008, 121, 30–37.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wills, T.A.; Soneji, S.S.; Choi, K.; Jaspers, I.; Tam, E.K. E-cigarette use and respiratory disorders: An integrative review of converging evidence from epidemiological and laboratory studies. Eur. Respir. J. 2020, 57, 1901815. [Google Scholar] [CrossRef]
- Bircan, E.; Bezirhan, U.; Porter, A.; Fagan, P.; Orloff, M.S. Electronic cigarette use and its association with asthma, chronic obstructive pulmonary disease (COPD) and asthma- COPD overlap syndrome among never cigarette smokers. Tob. Induc. Dis. 2021, 19, 23. [Google Scholar] [CrossRef]
- Roberts, J.; Chow, J.; Trivedi, K. Adult-Onset Asthma Associated with E-Cigarette Use. Cureus 2021, 13, e19190. [Google Scholar] [CrossRef]
- Clapp, P.; Jaspers, I. Electronic Cigarettes: Their Constituents and Potential Links to Asthma. Curr. Allergy Asthma Rep. 2017, 17, 79. [Google Scholar] [CrossRef]
- Nair, V.; Tran, M.; Behar, R.Z.; Zhai, S.; Cui, X.; Phandthong, R.; Wang, Y.; Pan, S.; Luo, W.; Pankow, J.F.; et al. Menthol in electronic cigarettes: A contributor to respiratory disease? Toxicol. Appl. Pharmacol. 2020, 407, 115238. [Google Scholar] [CrossRef]
- Rodríguez-Jiménez, J.C.; Moreno-Paz, F.J.; Teran, L.M.; Guaní-Guerra, E. Aspirin exacerbated respiratory disease: Current topics and trends. Respir. Med. 2018, 135, 62–75. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, H.; Fukutomi, Y.; Mitsui, C.; Kajiwara, K.; Watai, K.; Kamide, Y.; Nakamura, Y.; Hamada, Y.; Tomita, Y.; Sekiya, K.; et al. Omalizumab for Aspirin Hypersensitivity and Leukotriene Overproduction in Aspirin-exacerbated Respiratory Disease. A Randomized Controlled Trial. Am. J. Respir. Crit. Care Med. 2020, 201, 1488–1498. [Google Scholar] [CrossRef]
- Potaczek, D.P.; Trąd, G.; Sanak, M.; Garn, H.; Mastalerz, L. Local and Systemic Production of Pro-Inflammatory Eicosanoids Is Inversely Related to Sensitization to Aeroallergens in Patients with Aspirin-Exacerbated Respiratory Disease. J. Pers. Med. 2022, 12, 447. [Google Scholar] [CrossRef] [PubMed]
- Bochenek, G.; Stachura, T.; Szafraniec, K.; Plutecka, H.; Sanak, M.; Nizankowska-Mogilnicka, E.; Sladek, K. Diagnostic Accuracy of Urinary LTE4 Measurement to Predict Aspirin-Exacerbated Respiratory Disease in Patients with Asthma. J. Allergy Clin. Immunol. Pract. 2018, 6, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Choby, G.; Low, C.M.; Levy, J.M.; Stokken, J.K.; Pinheiro-Neto, C.; Bartemes, K.; Marino, M.; Han, J.K.; Divekar, R.; O’Brien, E.K.; et al. Urine Leukotriene E4: Implications as a Biomarker in Chronic Rhinosinusitis. Otolaryngol. Neck Surg. 2021, 166, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.-S.; Kim, J.-H.; Shin, Y.-S.; Ye, Y.-M.; Kim, S.-H.; Park, H.-S. Eosinophil activation and novel mediators in the aspirin-induced nasal response in AERD. Clin. Exp. Allergy 2013, 43, 730–740. [Google Scholar] [CrossRef]
- Kim, T.-H.; Lee, J.-Y.; Park, J.-S.; Park, S.-W.; Jang, A.-S.; Lee, J.-Y.; Byun, J.-Y.; Uh, S.-T.; Koh, E.-S.; Chung, I.Y.; et al. Fatty Acid Binding Protein 1 Is Related with Development of Aspirin-Exacerbated Respiratory Disease. PLoS ONE 2011, 6, e22711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawicki, L.R.; Arias, N.M.B.; Scaglia, N.; Lockhart, L.J.F.; Franchini, G.R.; Storch, J.; Córsico, B. FABP1 knockdown in human enterocytes impairs proliferation and alters lipid metabolism. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2017, 1862, 1587–1594. [Google Scholar] [CrossRef]
- Miyake, M.M.; Workman, A.D.; Ms, A.L.N.; Wu, D.; Mueller, S.K.; Finn, K.; Amiji, M.M.; Bleier, B.S. Discriminant analysis followed by unsupervised cluster analysis including exosomal cystatins predict presence of chronic rhinosinusitis, phenotype, and disease severity. Int. Forum Allergy Rhinol. 2019, 9, 1069–1076. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source | Biomarker | Proteomic Technology | Reference |
|---|---|---|---|
| NLF before treatment with glucocorticoids | ↓ORM, APOH, FGA, CTSD, SERPINB3 | LC-MS/MS | [41] |
| Nasal lavages | HSA, ECP, tryptase, cytokines, and total Igs | Bio-Plex suspension array Luminex xMAP system ELISA | [41] |
| Nasal mucus | ↑ALB, IgA, BPIFBI, APOA2, A2M, APOA1, SERPINA1 and C3 ↓LTF, LYZ, SLPI, WFDC2, IGJ, Ig hc V-III region BRO | LC-MS/MS | [47] |
| Nasal mucus | ↑APOA1, APOA2, APOA4 and B-100 | LC-MS/MS | [41] |
| Nasal mucus (Pollen season) | ↑CLU, IGKC ↓GSTP1, ELANE, HIST1H2BK, S100A8, S100A12 and ARHGDIB | LC-MS/MS | [41] |
| Nasal fluids (NF) | HNPs, Statherin, Thymosin-β4, P-D peptide, II-2, β-MSP, SLPI, Lysozyme-C, and their proteo-form | LC-MS/MS. | [56] |
| Serum | A2M, STAT3, p-STAT3 and IL-17 | iTRAQ, SCX, and LC-MS/MS. | [57] |
| Nasal brush samples | CST1, PRNT3, IFIT1, IFIT3 | Orbitrap-based, bottom-up label-free quantitative proteomic. LC-MS/MS | [41] |
| Serum after allergen immunotherapy (AIT) | LTA4H | Nanoflow (LC-MS/MS) | [41] |
| Source | Biomarker | Proteomic Technology | Reference |
|---|---|---|---|
| Exosomes from the NLF | MUC5AC and MUC5B | LC-MS/MS | [67,68] |
| CST1, PRDX5, and GP6 | SOMAscanTM | [70] | |
| Before steroid treatment ↓LPO, CAIII, PF4, PLAT ↑α2AP, LILRB2, CD209, APOE2 | SOMAscanTM | [83] | |
| After steroid treatment ↑APOL1, CSNK2A2, LPO, ANG | |||
| Eosinophils from nasal polyps | ↑GGT5 ↓DPEP2 | LC-MS/MS | [79] |
| Nasal secretions | ↑FTL, FTH1, GAA ↓ S100A7, SERPINB13, SERPINB8, CALML5, and RNASE3 | LC-MS/MS | [80] |
| Allergen | Cross-Reaction Protein | Food | Proteomic Technology | Reference |
|---|---|---|---|---|
| Api g 5 (Celery) | Phosphoglyceromutase | Mugwort Fennel | Immunoblots and MS | [89] |
| Cry j 7 (Japanese cedar) | Gibberellins | Peach, citrus, and apple | 2-DE and MS | [95] |
| Que i 1 | PR10 family | Banana, melon, apple, watermelon, pear, kiwi | MALDI-TOF/TOF and Orbitrap LC-MSMS | [98] |
| Source | Biomarker | Proteomic Technology | Reference |
|---|---|---|---|
| Serum | ↑IGKV2-40, IGHV3-74, IGKV1-27, V1-19, IGLC-7, APP, IGKV1-16, PIP ↓APOD, ACAN, CNTN1, C1S, AOC3, LRP1, COL10A1, ITGB1, PTPRG, ADAMTS13, DPP4, IFNa2, HSPA1A, APOB, NCAM2, TNXB, ACTB, CACNA2D1, POSTN, ALP, PK, LTF, ELANE, CTSG, MPO, G6PD, PFN1epl | LC-MS/MS | [107] |
| Serum during treatment | Mepolizumab: APOA1, CAYP1, A1AT and A2M | SWISS2DPAGE software | [131] |
| Benralizumab: CERU, CAYP1, A1AT, and A2M | |||
| Sputum | ↑HP, SERPINA1, PR4. ↓S100A9, S100A8, IGL, HTN1, SCGB1A1, SMR3B | LC-MS/MS | [109] |
| ↑CALM3 ↓LTF, MUC5AC, MUC5B, BPIFB, SFN | LC-MS/MS | [113] | |
| CSA-NH/NSA-NH: CSF, CXCL8, AGR ESA-NH/NSA-NH: AZU, ELANE, CFP, CXCL | SOMAscanTM | [132] | |
| Saliva | ↑SP-D | ELISA | [121] |
| PKHD1, ZNF263, DSG2, S100A2 | Shotgun proteomics | [123] | |
| Endobronchial biopsies | ANXA5, DPT, HIST1H2AH, LMNA, PPIA, RPBL7, and RPBL8 | NanoLC-LTQ Orbitrap mass spectrometer | [125] |
| Bronchial epithelial after chronic E-Cigarette exposure | CYP1B1, MUC5AC, and MUC4 | LC-MS/MS | [116] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizuet-de-Rueda, J.C.; Montero-Vargas, J.M.; Galván-Morales, M.Á.; Porras-Gutiérrez-de-Velasco, R.; Teran, L.M. Current Insights on the Impact of Proteomics in Respiratory Allergies. Int. J. Mol. Sci. 2022, 23, 5703. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105703
Vizuet-de-Rueda JC, Montero-Vargas JM, Galván-Morales MÁ, Porras-Gutiérrez-de-Velasco R, Teran LM. Current Insights on the Impact of Proteomics in Respiratory Allergies. International Journal of Molecular Sciences. 2022; 23(10):5703. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105703
Chicago/Turabian StyleVizuet-de-Rueda, Juan Carlos, Josaphat Miguel Montero-Vargas, Miguel Ángel Galván-Morales, Raúl Porras-Gutiérrez-de-Velasco, and Luis M. Teran. 2022. "Current Insights on the Impact of Proteomics in Respiratory Allergies" International Journal of Molecular Sciences 23, no. 10: 5703. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105703