
Inhibitory Effects of Saururus chinensis Extract on Receptor for Advanced Glycation End-Products-Dependent Inflammation and Diabetes-Induced Dysregulation of Vasodilation


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
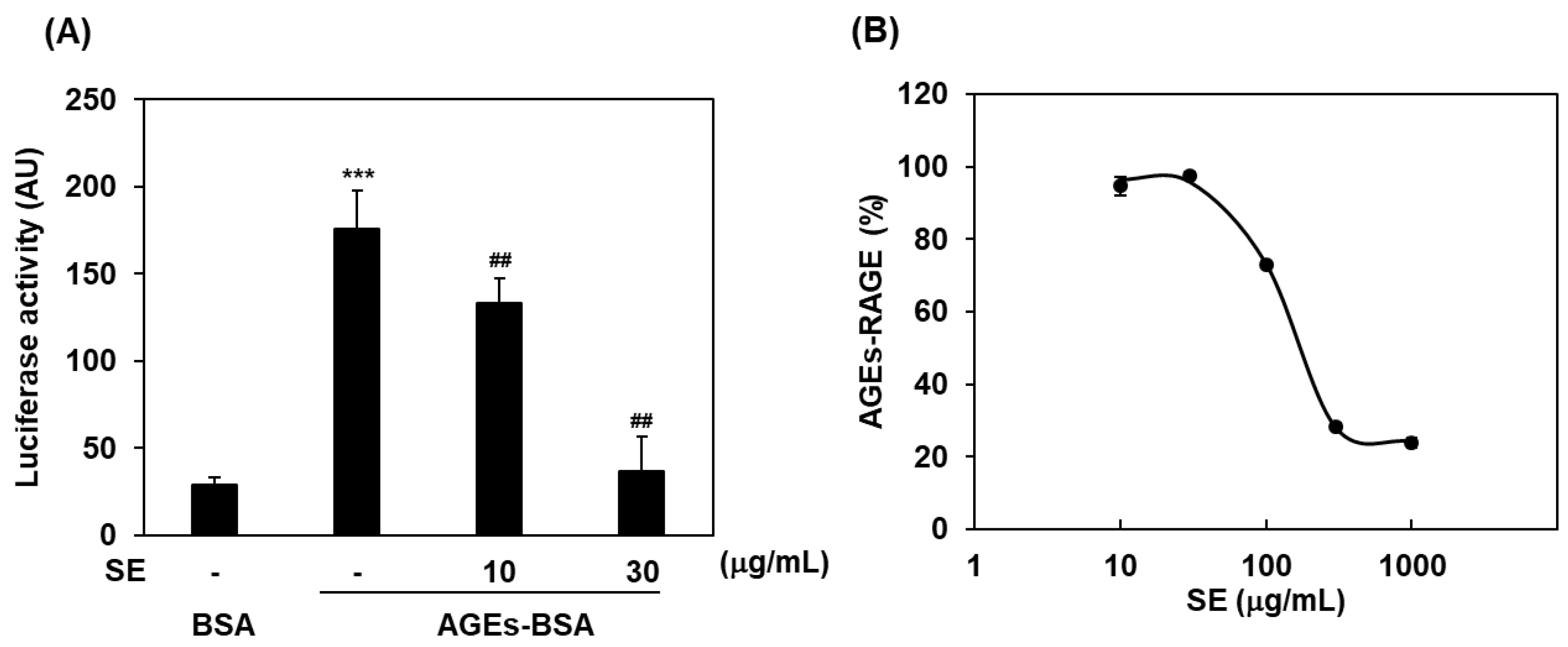
2.1. Inhibitory Effects of Saururus chinensis Extracts on RAGE Signaling
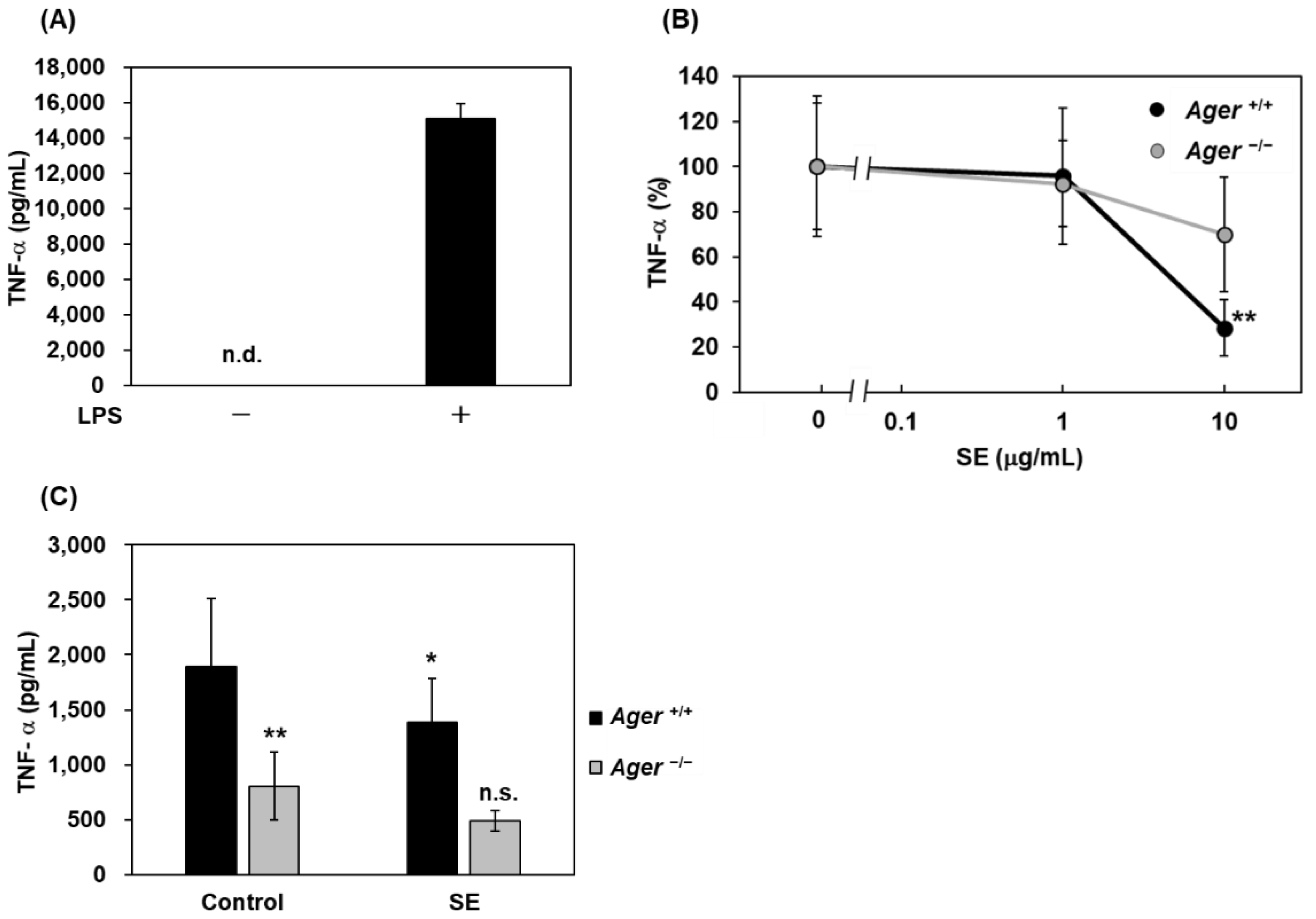
2.2. Inhibitory Effects of Saururus chinensis Extracts on LPS-Induced Cytokine Release
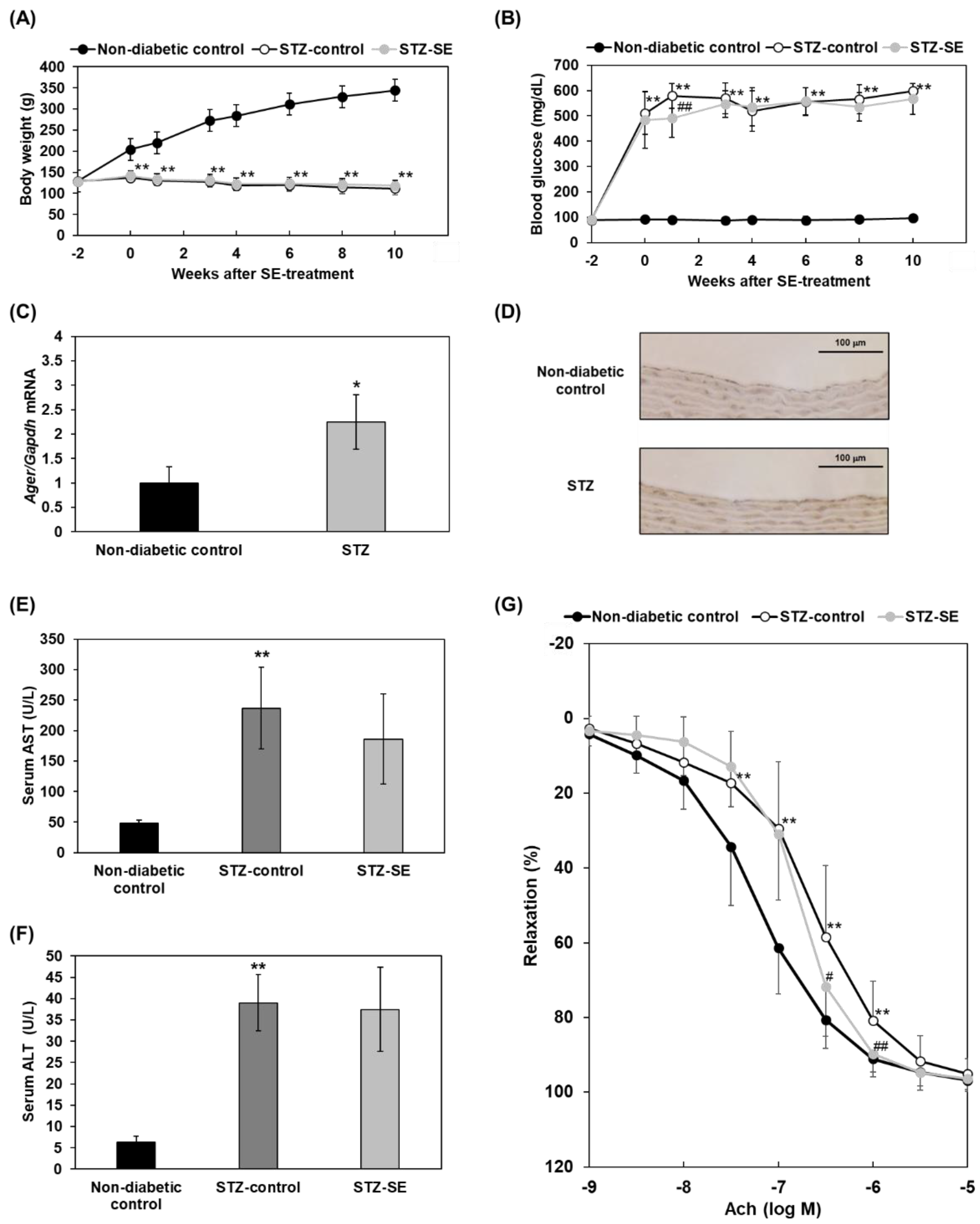
2.3. Effects of Saururus chinensis Extracts on Impairment of Acetylcholine-Induced Vasodilatation in Diabetes
3. Discussion
4. Materials and Methods
4.1. Sample Preparation
4.2. Animals
4.3. LPS-Induced Septic Mouse Model
4.4. STZ-Induced Diabetic Rat Model
4.5. NF-κB Luciferase Assay
4.6. Plate-Binding Assay
4.7. Real-Time Reverse Transcription PCR
4.8. Immunohistochemistry
4.9. Isolation of Peritoneal Macrophages
4.10. ELISA for TNF-α
4.11. Measurements of AST and ALT
4.12. Measurement of Vascular Relaxation
4.13. Statistical Analysis
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagai, R.; Matsumoto, K.; Ling, X.; Suzuki, H.; Araki, T.; Horiuchi, S. Glycolaldehyde, a Reactive Intermediate for Advanced Glycation End Products, Plays an Important Role in the Generation of an Active Ligand for the Macrophage Scavenger Receptor. Diabetes 2000, 49, 1714–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced Glycation End Products and Diabetic Complications. Korean J. Physiol. Pharmacol. 2014, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Yamamoto, H. Controlling the Receptor for Advanced Glycation End-Products to Conquer Diabetic Vascular Complications. J. Diabetes Investig. 2012, 3, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegab, Z.; Gibbons, S.; Neyses, L.; Mamas, M.A. Role of Advanced Glycation End Products in Cardiovascular Disease. World J. Cardiol. 2012, 4, 90–102. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.M.; Vianna, M.; Gerlach, M.; Brett, J.; Ryan, J.; Kao, J.; Esposito, C.; Hegarty, H.; Hurley, W.; Clauss, M. Isolation and Characterization of Two Binding Proteins for Advanced Glycosylation End Products from Bovine Lung Which Are Present on the Endothelial Cell Surface. J. Biol. Chem. 1992, 267, 14987–14997. [Google Scholar] [CrossRef]
- Hofmann, M.A.; Drury, S.; Fu, C.; Qu, W.; Taguchi, A.; Lu, Y.; Avila, C.; Kambham, N.; Bierhaus, A.; Nawroth, P.; et al. RAGE Mediates a Novel Proinflammatory Axis: A Central Cell Surface Receptor for S100/Calgranulin Polypeptides. Cell 1999, 97, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Hori, O.; Brett, J.; Slattery, T.; Cao, R.; Zhang, J.; Chen, J.X.; Nagashima, M.; Lundh, E.R.; Vijay, S.; Nitecki, D. The Receptor for Advanced Glycation End Products (RAGE) Is a Cellular Binding Site for Amphoterin. Mediation of Neurite Outgrowth and Co-Expression of Rage and Amphoterin in the Developing Nervous System. J. Biol. Chem. 1995, 270, 25752–25761. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Harashima, A.; Saito, H.; Tsuneyama, K.; Munesue, S.; Motoyoshi, S.; Han, D.; Watanabe, T.; Asano, M.; Takasawa, S.; et al. Septic Shock Is Associated with Receptor for Advanced Glycation End Products Ligation of LPS. J. Immunol. 2011, 186, 3248–3257. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.D.; Schmidt, A.M.; Anderson, G.M.; Zhang, J.; Brett, J.; Zou, Y.S.; Pinsky, D.; Stern, D. Enhanced Cellular Oxidant Stress by the Interaction of Advanced Glycation End Products with Their Receptors/Binding Proteins. J. Biol. Chem. 1994, 269, 9889–9897. [Google Scholar] [CrossRef]
- Lander, H.M.; Tauras, J.M.; Ogiste, J.S.; Hori, O.; Moss, R.A.; Schmidt, A.M. Activation of the Receptor for Advanced Glycation End Products Triggers a P21(Ras)-Dependent Mitogen-Activated Protein Kinase Pathway Regulated by Oxidant Stress. J. Biol. Chem. 1997, 272, 17810–17814. [Google Scholar] [CrossRef] [Green Version]
- Leerach, N.; Munesue, S.; Harashima, A.; Kimura, K.; Oshima, Y.; Kawano, S.; Tanaka, M.; Niimura, A.; Sakulsak, N.; Yamamoto, H.; et al. RAGE Signaling Antagonist Suppresses Mouse Macrophage Foam Cell Formation. Biochem. Biophys. Res. Commun. 2021, 555, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Liang, M.; Munesue, S.; Deguchi, K.; Harashima, A.; Furuhara, K.; Yuhi, T.; Zhong, J.; Akther, S.; Goto, H.; et al. Vascular RAGE Transports Oxytocin into the Brain to Elicit Its Maternal Bonding Behaviour in Mice. Commun. Biol. 2019, 2, 76. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Higashida, H. RAGE Regulates Oxytocin Transport into the Brain. Commun. Biol. 2020, 3, 70. [Google Scholar] [CrossRef] [PubMed]
- Munesue, S.-I.; Liang, M.; Harashima, A.; Zhong, J.; Furuhara, K.; Boitsova, E.B.; Cherepanov, S.M.; Gerasimenko, M.; Yuhi, T.; Yamamoto, Y.; et al. Transport of Oxytocin to the Brain after Peripheral Administration by Membrane-Bound or Soluble Forms of Receptors for Advanced Glycation End-Products. J. Neuroendocrinol. 2021, 33, e12963. [Google Scholar] [CrossRef]
- Basta, G.; Lazzerini, G.; Massaro, M.; Simoncini, T.; Tanganelli, P.; Fu, C.; Kislinger, T.; Stern, D.M.; Schmidt, A.M.; De Caterina, R. Advanced Glycation End Products Activate Endothelium through Signal-Transduction Receptor RAGE: A Mechanism for Amplification of Inflammatory Responses. Circulation 2002, 105, 816–822. [Google Scholar] [CrossRef] [Green Version]
- Esposito, C.; Gerlach, H.; Brett, J.; Stern, D.; Vlassara, H. Endothelial Receptor-Mediated Binding of Glucose-Modified Albumin Is Associated with Increased Monolayer Permeability and Modulation of Cell Surface Coagulant Properties. J. Exp. Med. 1989, 170, 1387–1407. [Google Scholar] [CrossRef] [Green Version]
- Bierhaus, A.; Illmer, T.; Kasper, M.; Luther, T.; Quehenberger, P.; Tritschler, H.; Wahl, P.; Ziegler, R.; Müller, M.; Nawroth, P.P. Advanced Glycation End Product (AGE)-Mediated Induction of Tissue Factor in Cultured Endothelial Cells Is Dependent on RAGE. Circulation 1997, 96, 2262–2271. [Google Scholar] [CrossRef]
- Rojas, A.; Romay, S.; González, D.; Herrera, B.; Delgado, R.; Otero, K. Regulation of Endothelial Nitric Oxide Synthase Expression by Albumin-Derived Advanced Glycosylation End Products. Circ. Res. 2000, 86, e50–e54. [Google Scholar] [CrossRef] [Green Version]
- Soro-Paavonen, A.; Watson, A.M.D.; Li, J.; Paavonen, K.; Koitka, A.; Calkin, A.C.; Barit, D.; Coughlan, M.T.; Drew, B.G.; Lancaster, G.I.; et al. Receptor for Advanced Glycation End Products (RAGE) Deficiency Attenuates the Development of Atherosclerosis in Diabetes. Diabetes 2008, 57, 2461–2469. [Google Scholar] [CrossRef] [Green Version]
- Myint, K.-M.; Yamamoto, Y.; Doi, T.; Kato, I.; Harashima, A.; Yonekura, H.; Watanabe, T.; Shinohara, H.; Takeuchi, M.; Tsuneyama, K.; et al. RAGE Control of Diabetic Nephropathy in a Mouse Model: Effects of RAGE Gene Disruption and Administration of Low-Molecular Weight Heparin. Diabetes 2006, 55, 2510–2522. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Rho, Y.; Kim, M.-Y.; Cho, J.Y. TAK1 in the AP-1 Pathway Is a Critical Target of Saururus chinensis (Lour.) Baill in Its Anti-Inflammatory Action. J. Ethnopharmacol. 2021, 279, 114400. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-W.; Koppula, S.; Park, S.-Y.; Hwang, J.-W.; Park, P.-J.; Lim, J.-H.; Choi, D.-K. Attenuation of Inflammatory-Mediated Neurotoxicity by Saururus chinensis Extract in LPS-Induced BV-2 Microglia Cells via Regulation of NF-ΚB Signaling and Anti-Oxidant Properties. BMC Complement. Altern. Med. 2014, 14, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, S.H.; Lee, E.J.; Cho, J.H.; Kim, H.S.; Kim, Y.C. Sauchinone, a Lignan from Saururus chinensis, Attenuates CCl4-Induced Toxicity in Primary Cultures of Rat Hepatocytes. Biol. Pharm. Bull. 2000, 23, 666–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, B.Y.; Lee, J.H.; Nam, J.B.; Hong, Y.S.; Lee, J.J. Lignans from Saururus chinensis Inhibiting the Transcription Factor NF-KappaB. Phytochemistry 2003, 64, 765–771. [Google Scholar] [CrossRef]
- Min, H.J.; Won, H.Y.; Kim, Y.C.; Sung, S.H.; Byun, M.R.; Hwang, J.-H.; Hong, J.-H.; Hwang, E.S. Suppression of Th2-Driven, Allergen-Induced Airway Inflammation by Sauchinone. Biochem. Biophys. Res. Commun. 2009, 385, 204–209. [Google Scholar] [CrossRef]
- Moon, T.C.; Kim, J.C.; Song, S.-E.; Suh, S.-J.; Seo, C.-S.; Kim, Y.K.; Jin, M.; Yang, J.H.; Son, J.-K.; Jahng, Y.; et al. Saucerneol D, a Naturally Occurring Sesquilignan, Inhibits LPS-Induced INOS Expression in RAW264.7 Cells by Blocking NF-KappaB and MAPK Activation. Int. Immunopharmacol. 2008, 8, 1395–1400. [Google Scholar] [CrossRef]
- Nho, J.-H.; Lee, H.-J.; Jung, H.-K.; Jang, J.-H.; Lee, K.-H.; Kim, A.-H.; Sung, T.-K.; Cho, H.-W. Effect of Saururus chinensis Leaves Extract on Type II Collagen-Induced Arthritis Mouse Model. BMC Complement. Altern. Med. 2019, 19, 2. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, A.; Yamamoto, Y.; Munesue, S.; Harashima, A.; Watanabe, T.; Yonekura, H.; Yamamoto, H.; Tsuchiya, H. Low Molecular Weight Heparin Suppresses Receptor for Advanced Glycation End Products-Mediated Expression of Malignant Phenotype in Human Fibrosarcoma Cells. Cancer Sci. 2013, 104, 740–749. [Google Scholar] [CrossRef] [Green Version]
- El-Far, A.H.A.M.; Munesue, S.; Harashima, A.; Sato, A.; Shindo, M.; Nakajima, S.; Inada, M.; Tanaka, M.; Takeuchi, A.; Tsuchiya, H.; et al. In Vitro Anticancer Effects of a RAGE Inhibitor Discovered Using a Structure-Based Drug Design System. Oncol. Lett. 2018, 15, 4627–4634. [Google Scholar] [CrossRef]
- Elçioğlu, H.K.; Kabasakal, L.; Özkan, N.; Çelikel, Ç.; Ayanoğlu-Dülger, G. A Study Comparing the Effects of Rosiglitazone and/or Insulin Treatments on Streptozotocin Induced Diabetic (Type I Diabetes) Rat Aorta and Cavernous Tissues. Eur. J. Pharmacol. 2011, 660, 476–484. [Google Scholar] [CrossRef]
- Topal, G.; Koç, E.; Karaca, C.; Altuğ, T.; Ergin, B.; Demirci, C.; Melikoğlu, G.; Meriçli, A.H.; Kucur, M.; Ozdemir, O.; et al. Effects of Crataegus Microphylla on Vascular Dysfunction in Streptozotocin-Induced Diabetic Rats. Phytother. Res. 2013, 27, 330–337. [Google Scholar] [CrossRef]
- Calles-Escandon, J.; Cipolla, M. Diabetes and Endothelial Dysfunction: A Clinical Perspective. Endocr. Rev. 2001, 22, 36–52. [Google Scholar] [CrossRef]
- Zemse, S.M.; Chiao, C.W.; Hilgers, R.H.P.; Webb, R.C. Interleukin-10 Inhibits the in Vivo and in Vitro Adverse Effects of TNF-Alpha on the Endothelium of Murine Aorta. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1160–H1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spillmann, F.; Van Linthout, S.; Miteva, K.; Lorenz, M.; Stangl, V.; Schultheiss, H.-P.; Tschöpe, C. LXR Agonism Improves TNF-α-Induced Endothelial Dysfunction in the Absence of Its Cholesterol-Modulating Effects. Atherosclerosis 2014, 232, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hallam, K.M.; Li, Q.; Ananthakrishnan, R.; Kalea, A.; Zou, Y.S.; Vedantham, S.; Schmidt, A.M.; Yan, S.F.; Ramasamy, R. Aldose Reductase and AGE-RAGE Pathways: Central Roles in the Pathogenesis of Vascular Dysfunction in Aging Rats. Aging Cell 2010, 9, 776–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Q.; Wang, B.; Zhang, X.-F.; Ma, Y.-P.; Liu, J.-D.; Wang, X.-Z. Contribution of Receptor for Advanced Glycation End Products to Vasculature-Protecting Effects of Exercise Training in Aged Rats. Eur. J. Pharmacol. 2014, 741, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Satheesan, S.; Figarola, J.L.; Dabbs, T.; Rahbar, S.; Ermel, R. Effects of a New Advanced Glycation Inhibitor, LR-90, on Mitigating Arterial Stiffening and Improving Arterial Elasticity and Compliance in a Diabetic Rat Model: Aortic Impedance Analysis. Br. J. Pharmacol. 2014, 171, 3103–3114. [Google Scholar] [CrossRef] [Green Version]
- Yonekura, H.; Yamamoto, Y.; Sakurai, S.; Petrova, R.G.; Abedin, M.J.; Li, H.; Yasui, K.; Takeuchi, M.; Makita, Z.; Takasawa, S.; et al. Novel Splice Variants of the Receptor for Advanced Glycation End-Products Expressed in Human Vascular Endothelial Cells and Pericytes, and Their Putative Roles in Diabetes-Induced Vascular Injury. Biochem. J. 2003, 370, 1097–1109. [Google Scholar] [CrossRef]
- Rendon-Mitchell, B.; Ochani, M.; Li, J.; Han, J.; Wang, H.; Yang, H.; Susarla, S.; Czura, C.; Mitchell, R.A.; Chen, G.; et al. IFN-Gamma Induces High Mobility Group Box 1 Protein Release Partly through a TNF-Dependent Mechanism. J. Immunol. 2003, 170, 3890–3897. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayashi, K.; Sato, K.; Ochi, S.; Kawano, S.; Munesue, S.; Harashima, A.; Oshima, Y.; Kimura, K.; Kyoi, T.; Yamamoto, Y. Inhibitory Effects of Saururus chinensis Extract on Receptor for Advanced Glycation End-Products-Dependent Inflammation and Diabetes-Induced Dysregulation of Vasodilation. Int. J. Mol. Sci. 2022, 23, 5757. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105757
Hayashi K, Sato K, Ochi S, Kawano S, Munesue S, Harashima A, Oshima Y, Kimura K, Kyoi T, Yamamoto Y. Inhibitory Effects of Saururus chinensis Extract on Receptor for Advanced Glycation End-Products-Dependent Inflammation and Diabetes-Induced Dysregulation of Vasodilation. International Journal of Molecular Sciences. 2022; 23(10):5757. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105757
Chicago/Turabian StyleHayashi, Kenjiro, Koichi Sato, Seishi Ochi, Shuhei Kawano, Seiichi Munesue, Ai Harashima, Yu Oshima, Kumi Kimura, Takashi Kyoi, and Yasuhiko Yamamoto. 2022. "Inhibitory Effects of Saururus chinensis Extract on Receptor for Advanced Glycation End-Products-Dependent Inflammation and Diabetes-Induced Dysregulation of Vasodilation" International Journal of Molecular Sciences 23, no. 10: 5757. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105757