
First Report on Genome Editing via Ribonucleoprotein (RNP) in Castanea sativa Mill.

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
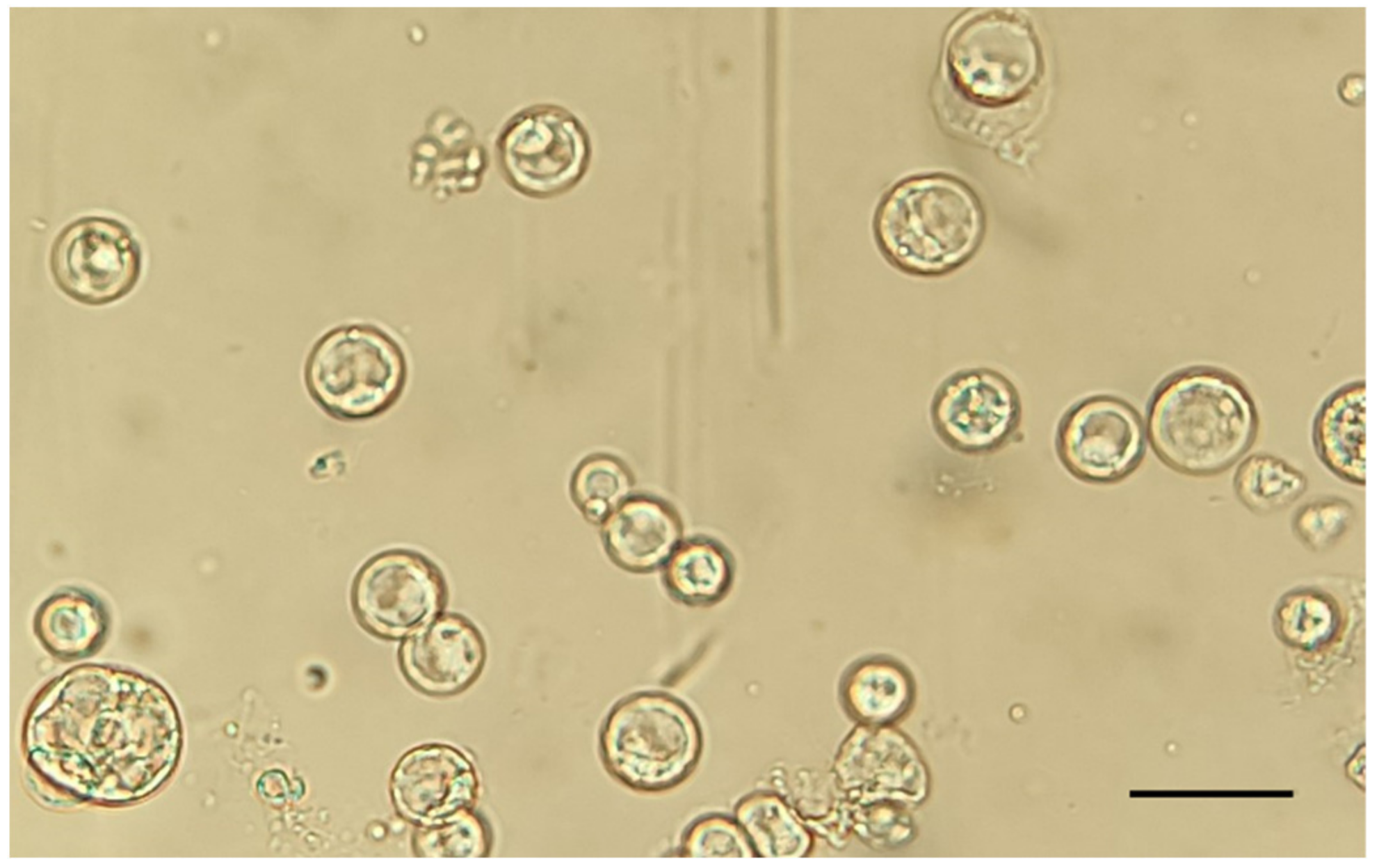
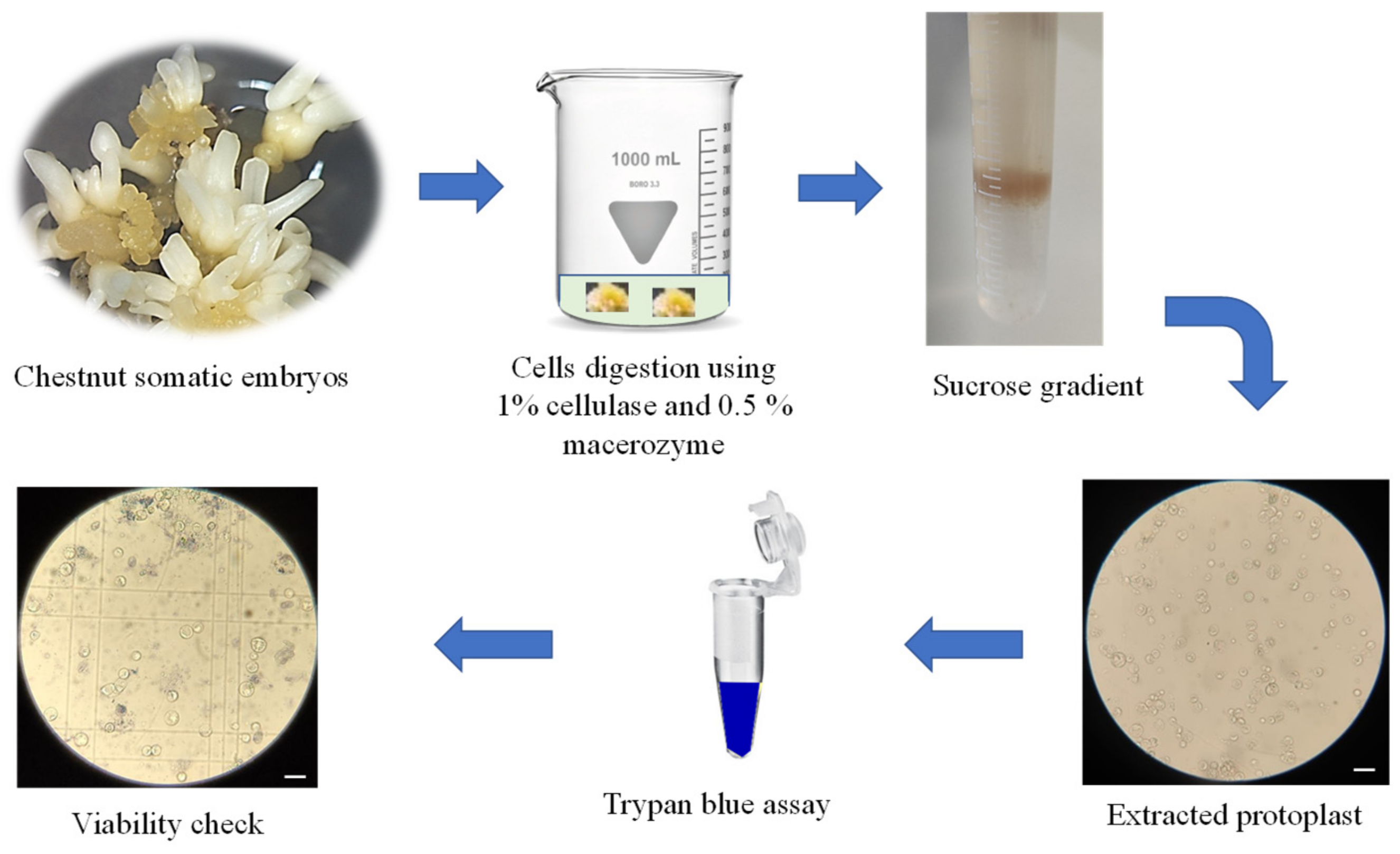
2.1. Protoplast Isolation from Somatic Embryos
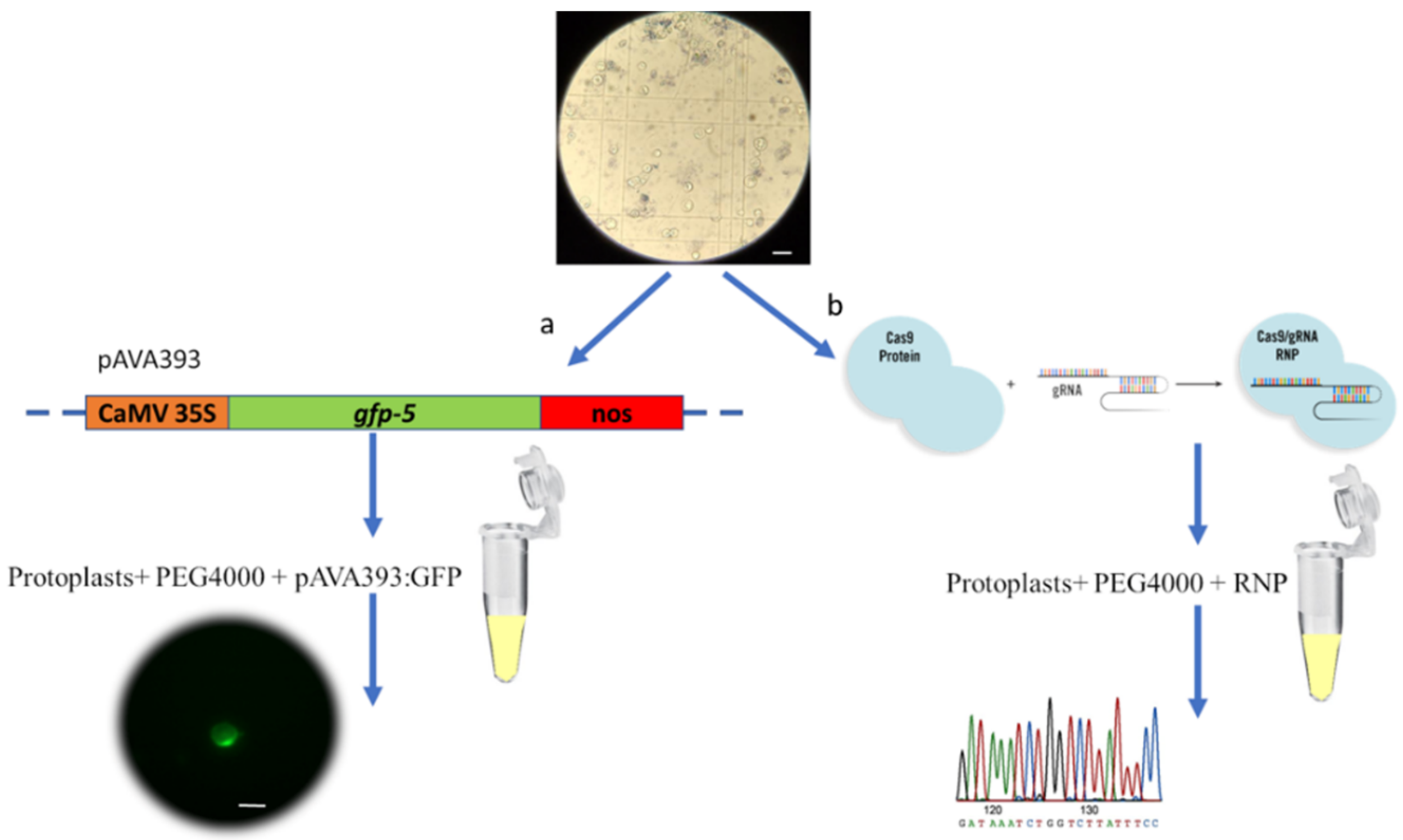
2.2. Protoplast Transfection with GFP Vector
2.3. Protoplast Transfection with CRISPR RNPs
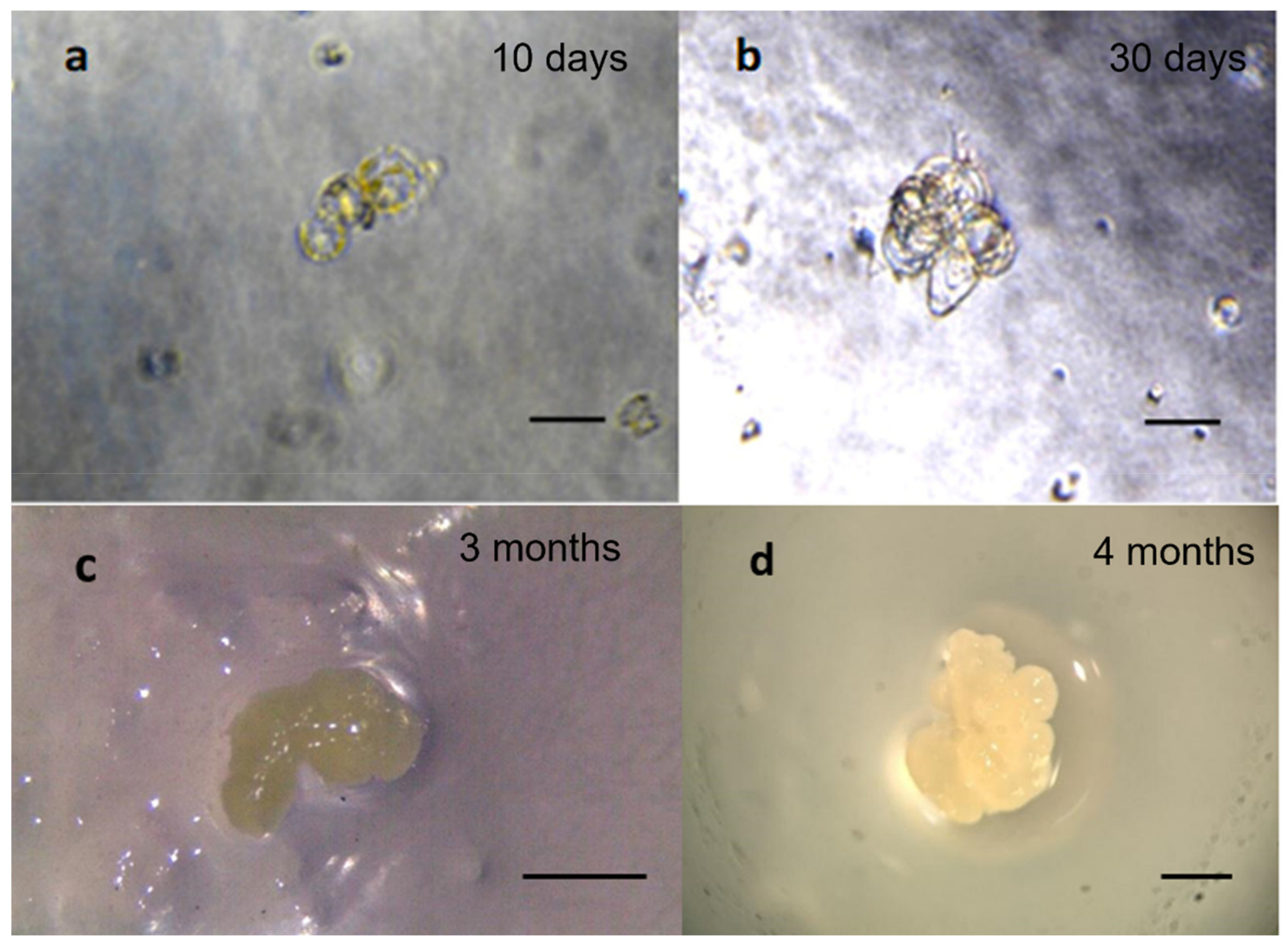
2.4. Regeneration of Protoplasts
3. Material and Methods
3.1. Plant Material
3.2. Protoplast Isolation
3.3. Protoplast Transfection with GFP Vector
3.4. Protoplast Transfection with CRISPR RNPs
3.5. Protoplast Culture and Regeneration
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fernández-López, J.; Alía, R. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Chestnut (Castanea sativa); International Plant Genetic Resources Institute: Rome, Italy, 2003; p. 6. [Google Scholar]
- Conedera, M.; Krebs, P. History, Present Situation and Perspective of Chestnut Cultivation in Europe. Acta Hortic. 2008, 784, 23–28. [Google Scholar] [CrossRef]
- Pavese, V.; Moglia, A.; Gonthier, P.; Torello Marinoni, D.; Cavalet-Giorsa, E.; Botta, R. Identification of susceptibility genes in Castanea sativa and their transcription dynamics following pathogen infection. Plants 2021, 10, 913. [Google Scholar] [CrossRef] [PubMed]
- Brussino, G.; Bosio, M.; Giordano, R.; Ramello, F.; Melika, G. A dangerous exotic insect threating European chestnut. Inf. Agrar. 2002, 58, 59–61. [Google Scholar]
- Aebi, A.; Schönrogge, K.; Melika, G.; Alma, A.; Bosio, G.; Quacchia, A.; Picciau, L.; Abe, Y.; Moriya, S.; Yara, K.; et al. Parasitoid recruitment to the globally invasive chestnut gall wasp Dryocosmus kuriphilus. In Ecology and Evolution of Galling Arthropods and Their Associates; Ozaki, K., Yukawa, J., Ohgushi, T., Price, P.W., Eds.; Springer: Tokyo, Japan, 2006; pp. 103–121. [Google Scholar]
- Lione, G.; Giordano, L.; Turina, M.; Gonthier, P. Hail-nduced Infections of the Chestnut Blight Pathogen Cryphonectria parasitica Depend on Wound Size and May Lead to Severe Diebacks. Phytopathology 2020, 110, 1280–1293. [Google Scholar] [CrossRef] [PubMed]
- Limera, C.; Sabbadini, S.; Sweet, J.B.; Mezzetti, B. New biotechnological tools for the genetic improvement of major woody fruit species. Front. Plant Sci. 2017, 8, 1418. [Google Scholar] [CrossRef] [PubMed]
- Ahmar, S.; Gill, R.A.; Jung, K.-H.; Faheem, A.; Qasim, M.U.; Mubeen, M.; Zhou, W. Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook. Int. J. Mol. Sci. 2020, 21, 2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osakabe, Y.; Liang, Z.; Ren, C.; Nishitani, C.; Osakabe, K.; Wada, M.; Komori, S.; Malnoy, M.; Velasco, R.; Poli, M.; et al. CRISPR–Cas9-mediated genome editing in apple and grapevine. Nat. Protoc. 2018, 13, 2844–2863. [Google Scholar] [CrossRef] [PubMed]
- Santillán Martínez, M.I.; Bracuto, V.; Koseoglou, E.; Appiano, M.; Jacobsen, E.; Visser, R.G.F.; Wolters, A.A.; Bai, Y. CRISPR/Cas9-targeted mutagenesis of the tomato susceptibility gene PMR4 for resistance against powdery mildew. BMC Plant Biol. 2020, 20, 284. [Google Scholar] [CrossRef]
- Corredoira, E.; Montenegro, D.; San José, M.C.; Vieitez, A.M.; Ballester, A. Agrobacterium-mediated transformation of European chestnut embryogenic cultures. Plant Cell Rep. 2004, 23, 311–318. [Google Scholar] [CrossRef]
- Corredoira, E.; Valladares, S.; Allona, I.; Aragoncillo, C.; Vieitez, A.M.; Ballester, A. Genetic transformation of European chestnut somatic embryos with a native thaumatin-like protein (CsTL1) gene isolated from Castanea sativa seeds. Tree Physiol. 2012, 32, 1389–1402. [Google Scholar] [CrossRef] [Green Version]
- Corredoira, E.; San José, M.C.; Vieitez, A.M.; Allona, I.; Aragoncillo, C.; Ballester, A. Agrobacterium-mediated transformation of European chestnut somatic embryos with a Castanea sativa (Mill.) endochitinase gene. New For. 2016, 47, 669–684. [Google Scholar] [CrossRef]
- Pavese, V.; Moglia, A.; Corredoira, E.; Martínez, M.T.; Torello Marinoni, D.; Botta, R. First report of CRISPR/Cas9 Gene Editing in Castanea sativa Mill. Front. Plant Sci. 2021, 12, 728516. [Google Scholar] [CrossRef]
- Scintilla, S.; Salvagnin, U.; Giacomelli, L.; Zeilmaker, T.; Malnoy, M.A.; van der Voort, J.R.; Moser, C. Regeneration of Plants from DNA-free Edited Grapevine Protoplasts. bioRxiv 2021. [Google Scholar] [CrossRef]
- Jia, H.; Wang, N. Targeted genome editing of sweet orange using cas9/sgrna. PLoS ONE 2014, 9, e93806. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Jacobs, T.B.; Xue, L.; Harding, S.A.; Tsai, C.J. Exploiting SNPs for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-coumarate: CoA ligase specificity and redundancy. New Phytol. 2015, 208, 298–301. [Google Scholar] [CrossRef]
- Xing, S.; Jia, M.; Wei, L.; Mao, W.; Abbasi, U.A.; Zhao, Y.; Chen, Y.; Cao, M.; Zhang, K.; Dai, Z.; et al. CRISPR/Cas9-introduced single and multiple mutagenesis in strawberry. J. Genet. Genom. 2018, 45, 685–687. [Google Scholar] [CrossRef]
- Nishitani, C.; Hirai, N.; Komori, S.; Wada, M.; Okada, K.; Osakabe, K.; Yamamoto, T.; Osakabe, Y. Efficient genome editing in apple using a CRISPR/Cas9 system. Sci. Rep. 2016, 6, 31481. [Google Scholar] [CrossRef]
- Charrier, A.; Vergne, E.; Dousset, N.; Richer, A.; Petiteau, A.; Chevreau, E. Efficient targeted mutagenesis in apple and first time edition of pear using the CRISPR-Cas9 System. Front. Plant Sci. 2019, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Fister, A.S.; Landherr, L.; Maximova, S.N.; Guiltinan, M.J. Transient expression of CRISPR/Cas9 machinery targeting TcNPR3 enhances defense response in Theobroma cacao. Front. Plant Sci. 2018, 9, 268. [Google Scholar] [CrossRef] [Green Version]
- Woo, J.W.; Kim, J.; Kwon, S.I.; Corvalán, C.; Cho, S.W.; Kim, H.; Kim, S.G.; Kim, S.T.; Choe, S.; Kim, J.S. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nat. Biotechnol. 2015, 33, 1162–1164. [Google Scholar] [CrossRef]
- Chen, G.; Abdeen, A.A.; Wang, Y.; Shahi, P.K.; Robertson, S.; Xie, R.; Suzuki, M.; Pattnaik, B.R.; Saha, K.; Gong, S. A biodegradable nanocapsule delivers a Cas9 ribonucleoprotein complex for in vivo genome editing. Nat. Nanotechnol. 2019, 14, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Demirer, G.S.; Landry, M.P. Delivering genes to plants. Chem. Eng. Prog. 2017, 113, 40–45. [Google Scholar]
- Fossi, M.; Amundson, K.; Kuppu, S.; Britt, A.; Comai, L. Regeneration of Solanum tuberosum Plants from Protoplasts Induces Widespread Genome Instability. Plant Physiol. 2019, 180, 78–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, J.S. Bypassing GMO regulations with CRISPR gene editing. Nat. Biotechnol. 2016, 34, 1014–1015. [Google Scholar] [CrossRef]
- Malnoy, M.; Viola, R.; Jung, M.-H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Nagamangala Kanchiswamy, C. DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef]
- Poovaiah, C.; Phillips, L.; Geddes, B.; Reeves, C.; Sorieul, M.; Thorlby, G. Genome editing with CRISPR/Cas9 in Pinus radiata (D. Don). BMC Plant Biol. 2021, 21, 363. [Google Scholar] [CrossRef]
- Bertini, E.; Tornielli, G.B.; Pezzotti, M.; Zenoni, S. Regeneration of plants from embryogenic callus-derived protoplasts of Garganega and Sangiovese grapevine (Vitis vinifera L.) cultivars. Plant Cell Tiss. Organ Cult. 2019, 138, 239–246. [Google Scholar] [CrossRef]
- Kuzminsky, E.; Meschini, R.; Terzoli, S.; Pavani, L.; Silvestri, C.; Choury, Z.; Scarascia-Mugnozza, G. Isolation of Mesophyll Protoplasts from Mediterranean Woody Plants for the Study of DNA Integrity under Abiotic Stress. Front. Plant Sci. 2016, 7, 1168. [Google Scholar] [CrossRef] [Green Version]
- Brandt, K.M.; Gunn, H.; Moretti, N.; Zemetra, R.S. A Streamlined Protocol for Wheat (Triticum aestivum) Protoplast Isolation and Transformation with CRISPR-Cas Ribonucleoprotein Complexes. Plant Sci. 2020, 11, 769. [Google Scholar] [CrossRef]
- Shen, J.; Fu, J.; Ma, J.; Wang, X.; Gao, C.; Zhuang, C.; Wan, J.; Jiang, L. Isolation, culture, and transient transformation of plant protoplasts. Curr. Protoc. Cell. Biol. 2014, 63, 2.8.1–2.8.17. Curr. Protoc. Cell. Biol. 2014, 63, 2.8.1–2.8.17. [Google Scholar]
- Coy, M.R.; Abbitt, S.E.; Frank, M.J. Protoplast Isolation and Transfection in Maize. In Protoplast Technology; Wang, K., Zhang, F., Eds.; Humana: New York, NY, USA, 2022; Volume 2464, pp. 91–104. [Google Scholar]
- Masani, M.Y.A.; Noll, G.A.; Parveez, G.K.A.; Sambanthamurthi, R.; Prufer, D. Efficient Transformation of Oil Palm Protoplasts by PEGMediated Transfection and DNA Microinjection. PLoS ONE 2014, 9, e96831. [Google Scholar]
- Sun, B.; Yuan, Q.; Zheng, H.; Hiang, S.; Jiang, M.; Wang, M.; Chen, Q.; Li, M.; Zhang, Y.; Luo, Y.; et al. An Efficient and Economical Protocol for Isolating, Purifying and PEG-Mediated Transient Gene Expression of Chinese Kale Hypocotyl Protoplasts. Plants 2019, 8, 385. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Wanga, Z.; Chenga, J.; Xhaoa, W.; Li, X.; Wanga, H.; Zhanga, Z.; Sui, X. An efficient cucumber (Cucumis sativus L.) protoplast isolation and transient expression system. Sci. Hortic. 2013, 105, 206–212. [Google Scholar] [CrossRef]
- Yoo, S.; Cho, Y.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [Green Version]
- Ohnuma, M.; Yokoyama, T.; Inouye, T.; Sekine, Y.; Tanaka, K. Polyethylene glycol (PEG)-mediated transient gene expression in a red alga, Cyanidioschyzon merolae 10D. Plant Cell Physiol. 2008, 49, 117–120. [Google Scholar] [CrossRef]
- Waltz, E. Gene-edited CRISPR mushroom escapes US regulation. Nature 2016, 532, 293. [Google Scholar] [CrossRef] [Green Version]
- Bortesi, L.; Zhu, C.; Zischewski, J.; Perez, L.; Bassié, L.; Nadi, R.; Forni, G.; Lade, S.B.; Soto, E.; Jin, X.; et al. Patterns of CRISPR/Cas9 activity in plants, animals and microbes. Plant Biotechnol. J. 2016, 14, 2203–2216. [Google Scholar] [CrossRef]
- Attree, S.M.; Dunstan, D.I.; Fowke, L.C. Plantlet Regeneration from Embryogenic Protoplasts of White Spruce (Picea glauca). Nat. Biotechnol. 1989, 7, 1060–1062. [Google Scholar] [CrossRef]
- Klimaszewska, K. Recovery of somatic embryos and plantlets from protoplast cultures of Larix × eurolepis. Plant Cell Rep. 1989, 8, 440–444. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Corredoira, E.; Ballester, A.; Vieitez, A.M. Proliferation, maturation and germination of Castanea sativa Mill. somatic embryos originated from leaf explants. Ann. Bot. 2003, 92, 129–136. [Google Scholar] [PubMed] [Green Version]
- Corredoira, E.; Ballester, A.; Vieitez, F.J.; Vieitez, A.M. Somatic Embryogenesis in Chestnut. In Somatic Embryogenesis. Plant Cell Monographs; Mujib, A., Šamaj, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 2, pp. 177–199. [Google Scholar]
- Nitsch, J.P.; Nitsch, C. Haploid plants from pollen grains. Science 1969, 163, 85–87. [Google Scholar] [CrossRef] [PubMed]
- Corredoira, E.; Merkle, S.A.; Martínez, M.T.; Toribio, M.; Canhoto, J.M.; Correia, S.I.; Ballester, A.; Vieitez, A.M. Non-zygotic embryogenesis in hardwood species. Crit. Rev. Plant Sci. 2019, 38, 29–97. [Google Scholar] [CrossRef]
- Corredoira, E.; Toribio, M.; Vieitez, A.M. Clonal propagation via somatic embryogenesis in Quercus spp. In Tree Biotechnology; Ramawhat, K.G., Mérillon, J.-M., Ahuja, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 262–302. [Google Scholar]
- Martínez, M.T.; San-José, M.C.; Arrillaga, I.; Cano, V.; Morcillo, M.; Cernadas, M.J.; Corredoira, E. Holm oak somatic embryogenesis: Current status and future perspectives. Front. Plant Sci. 2019, 10, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, J.M.; Bopaiah, A.K. Studies on the intiation [sic] of callusing and regeneration of plantlets in three different basal media with varied plant growth regulators for the micropropagation of Anthurium scherzeriaum [sic] using leaf and spathe as explants. Afr. J. Biotechnol. 2012, 11, 6259–6268. [Google Scholar]
- Kamlesh, R.P.; Narpat, S.S.; Graeme, P.B.; Trevor, A.T. Isolation and culture of protoplasts from cytoledons of Pinus coulteri D. Don. Plant Cell Tiss. Organ Cult. 1984, 3, 85–90. [Google Scholar]
- Ochatt, S.J.; Delaitre, C.; Lionneton, E.; Huchette, O.; Patat-Ochatt, E.M.; Kahane, R. One team, PCMV, and one approach, in vitro biotechnology, for one aim, the breeding of quality plants with a wide array of species. Crops Growth Qual. Biotechnol. 2005, 1038–1067. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples. | Efficiency (%) | R2 | Mutations |
|---|---|---|---|
| P1 | 21.4 | 0.96 | −3; −1; +1 |
| P2 | 17.9 | 0.95 | −1 |
| P3 | 14.6 | 0.97 | −1; +1 |
| Primers | |
|---|---|
| Name | Sequence |
| Seq_pds_gRNA1_F | TGGAAACTTTGGGTATGCATCC |
| Seq_pds_gRNA1_R | TTCTGTGATTGGTAGGCTTTCA |
| Media Components | C1 | C2 | C3 |
|---|---|---|---|
| Basal medium | MS | NN | MS |
| NAA (mg/L) | 1 | 1 | - |
| BAP (mg/L) | 0.5 | 0.5 | 0.2 |
| 2,4-D (mg/L) | - | - | 2 |
| Casein hydrolysate (g/L) | 0.5 | - | - |
| D-Mannitol (g/L) | - | 30 | 30 |
| Sucrose (g/L) | 30 | 5 | 5 |
| L-Glucose (g/L) | 50 | 50 | 50 |
| Plant agar (g/L) | 6 | 6 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavese, V.; Moglia, A.; Abbà, S.; Milani, A.M.; Torello Marinoni, D.; Corredoira, E.; Martínez, M.T.; Botta, R. First Report on Genome Editing via Ribonucleoprotein (RNP) in Castanea sativa Mill. Int. J. Mol. Sci. 2022, 23, 5762. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105762
Pavese V, Moglia A, Abbà S, Milani AM, Torello Marinoni D, Corredoira E, Martínez MT, Botta R. First Report on Genome Editing via Ribonucleoprotein (RNP) in Castanea sativa Mill. International Journal of Molecular Sciences. 2022; 23(10):5762. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105762
Chicago/Turabian StylePavese, Vera, Andrea Moglia, Silvia Abbà, Anna Maria Milani, Daniela Torello Marinoni, Elena Corredoira, Maria Teresa Martínez, and Roberto Botta. 2022. "First Report on Genome Editing via Ribonucleoprotein (RNP) in Castanea sativa Mill." International Journal of Molecular Sciences 23, no. 10: 5762. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105762