
MDM2-Mediated Ubiquitination of RXRβ Contributes to Mitochondrial Damage and Related Inflammation in Atherosclerosis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
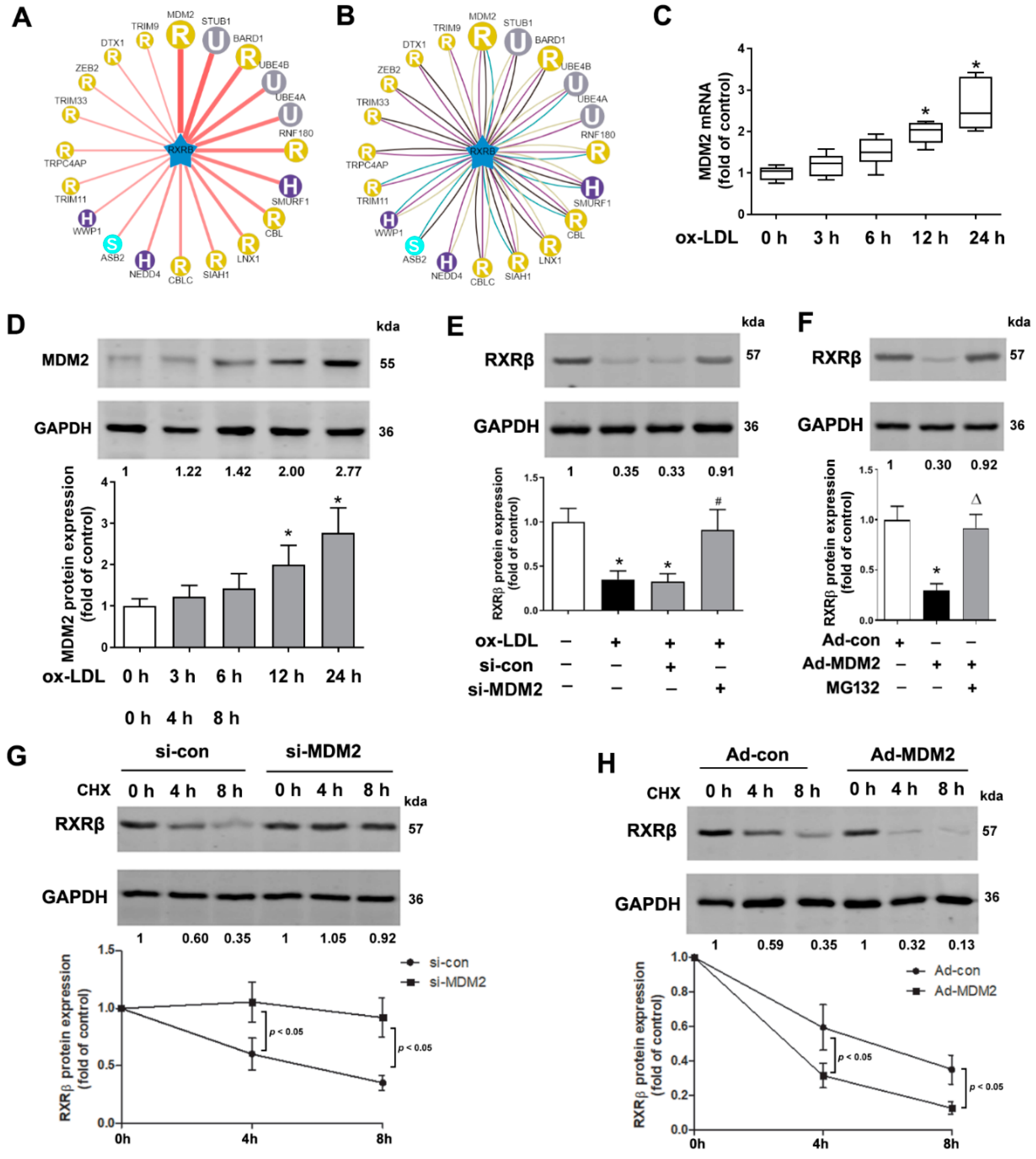
2.1. RXRβ Is Ubiquitinated in ox-LDL-Induced HAECs
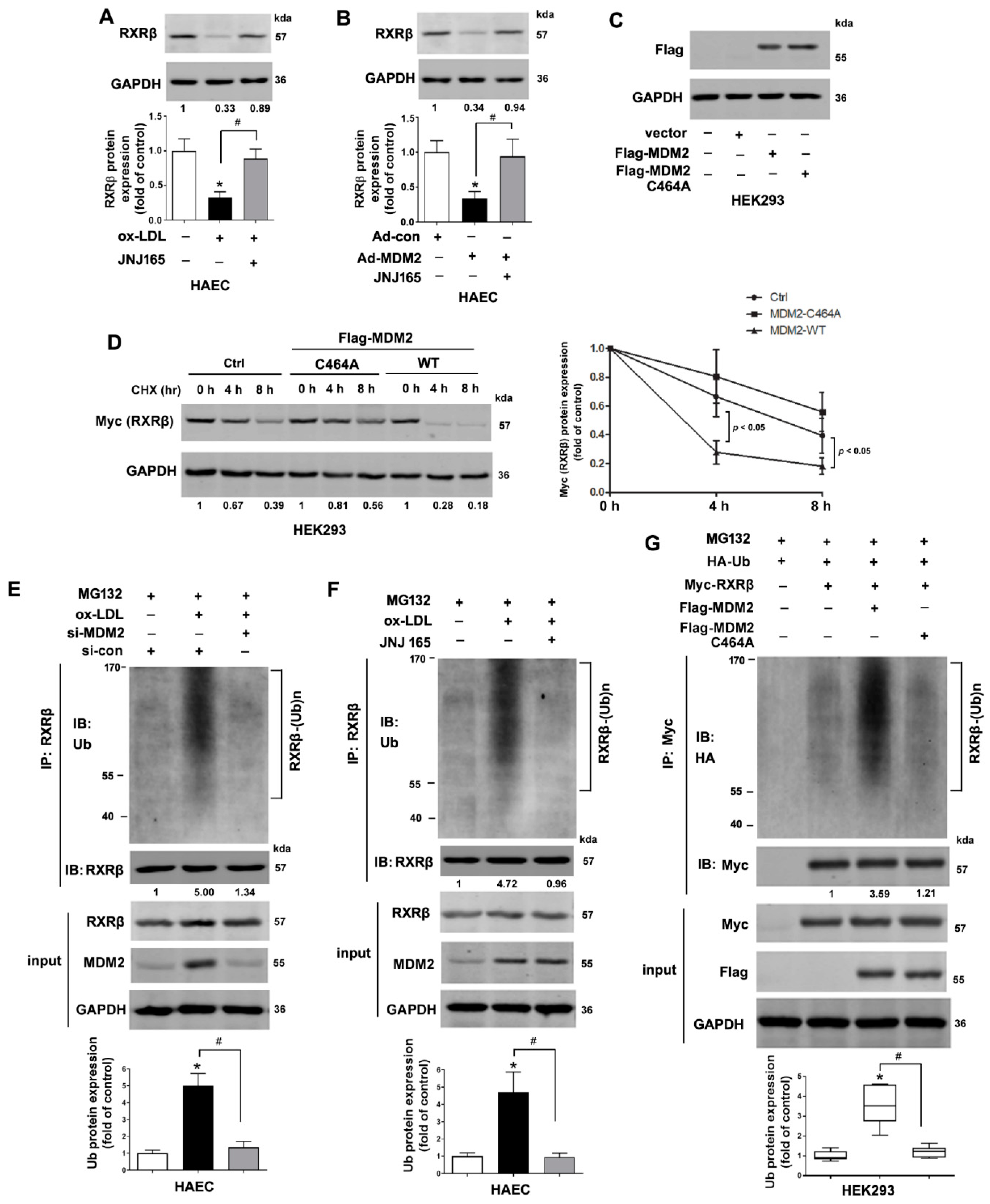
2.2. MDM2 Can Affect the Stability of RXRβ
2.3. MDM2 Mediates the Poly-Ubiquitination of RXRβ
2.4. MDM2 Interacts with RXRβ
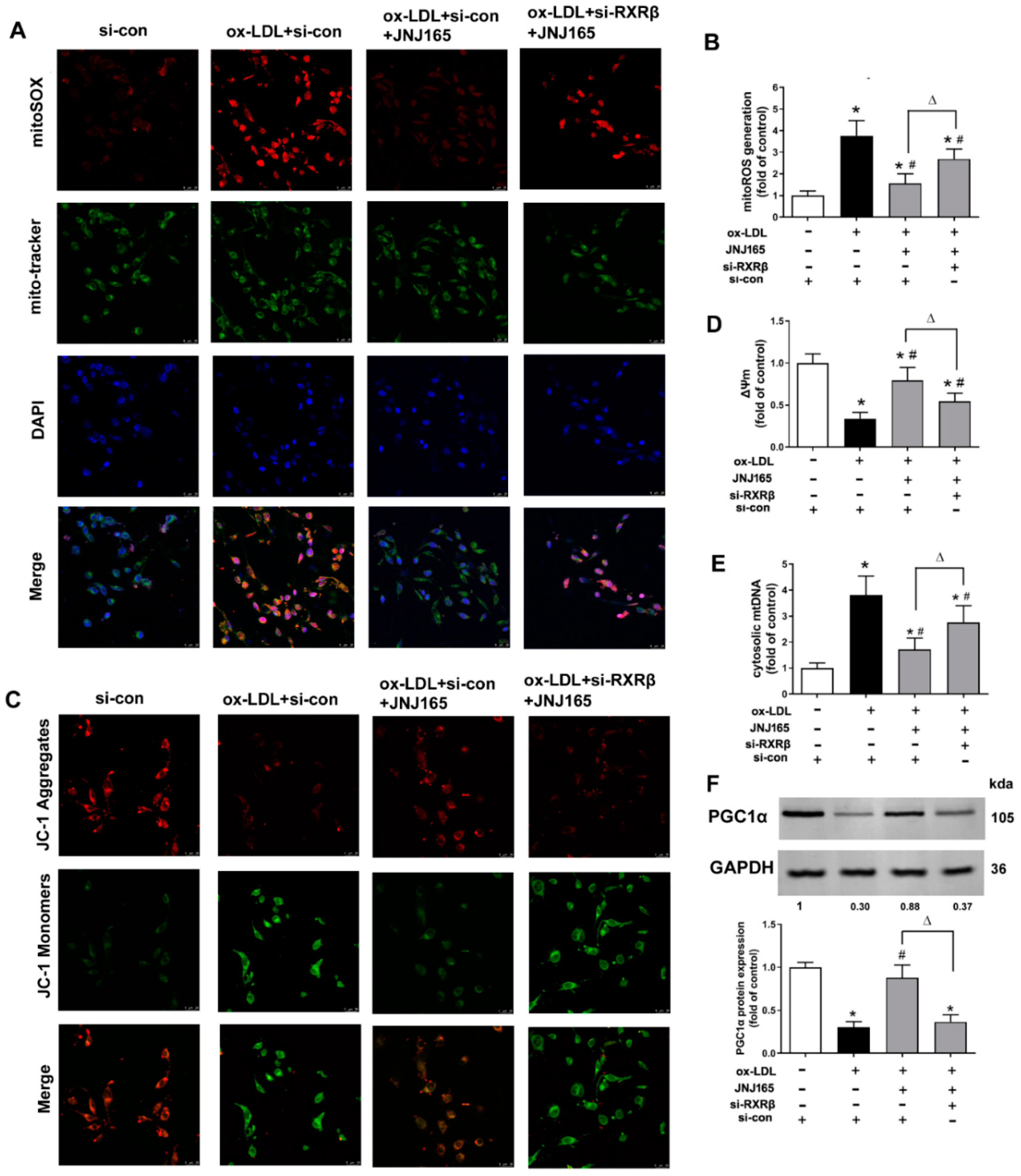
2.5. MDM2 Inhibitor JNJ-165 Provides Protection against ox-LDL-Induced Mitochondrial Damage via RXRβ
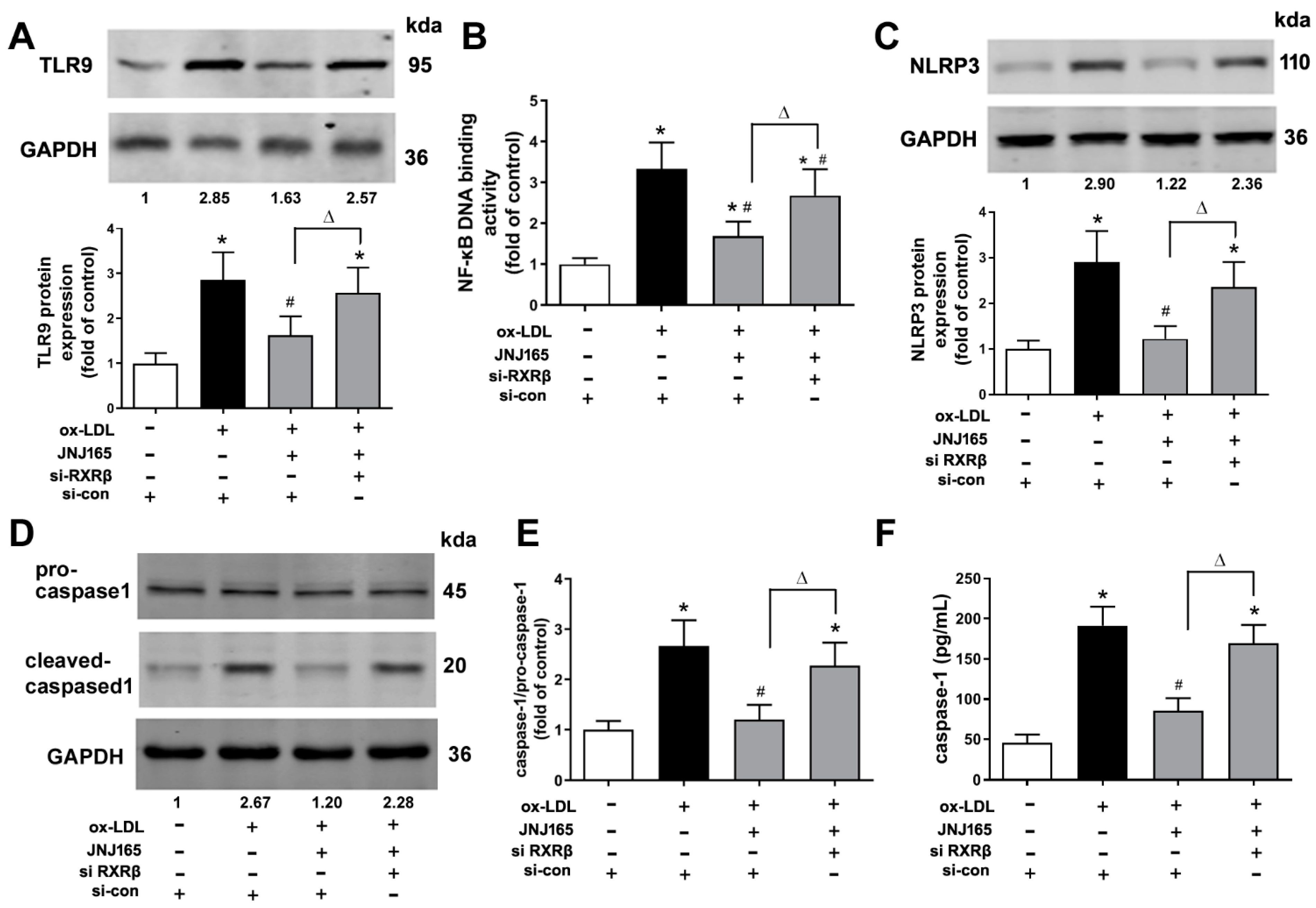
2.6. MDM2 Inhibitor JNJ-165 Provides Protection against ox-LDL-Induced Mitochondrial Related Inflammation via RXRβ
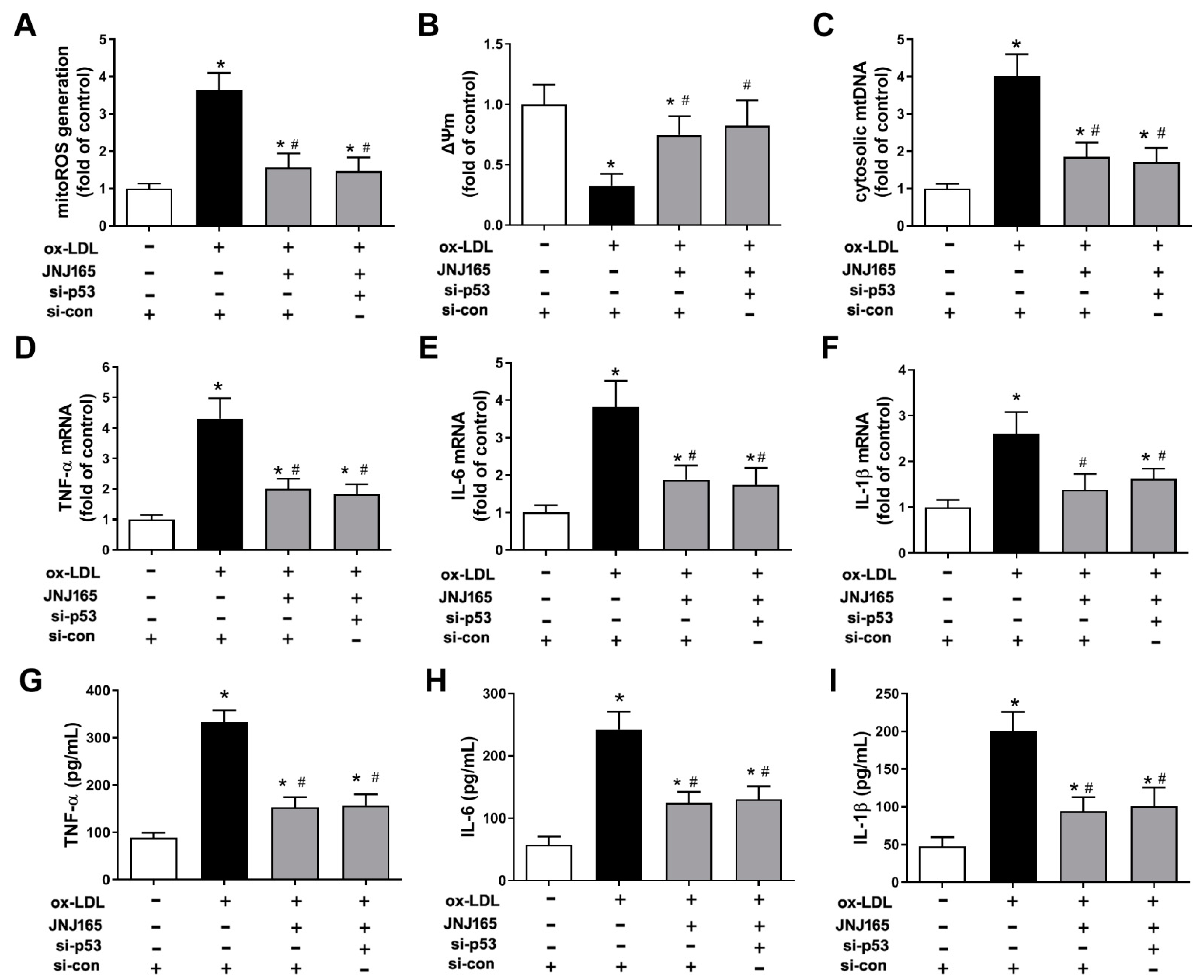
2.7. MDM2 Inhibitor JNJ-165 Provides Protection against ox-LDL-Induced Mitochondrial Damage and Related Inflammation Independent of p53
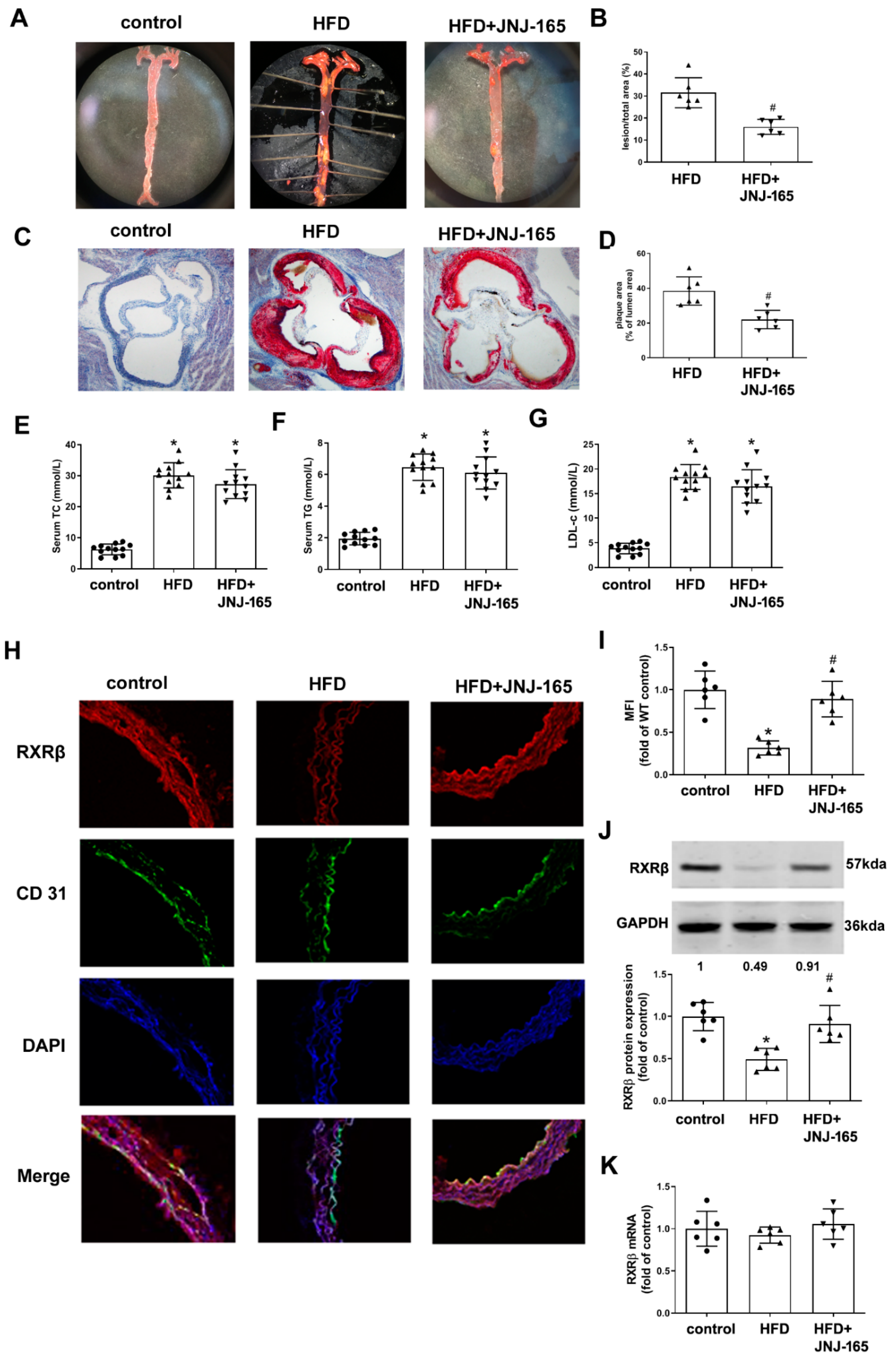
2.8. MDM2 Inhibition Alleviates Atherosclerotic Lesion Formation and RXRβ Protein Expression in HFD-Fed LDLr-/- Mice
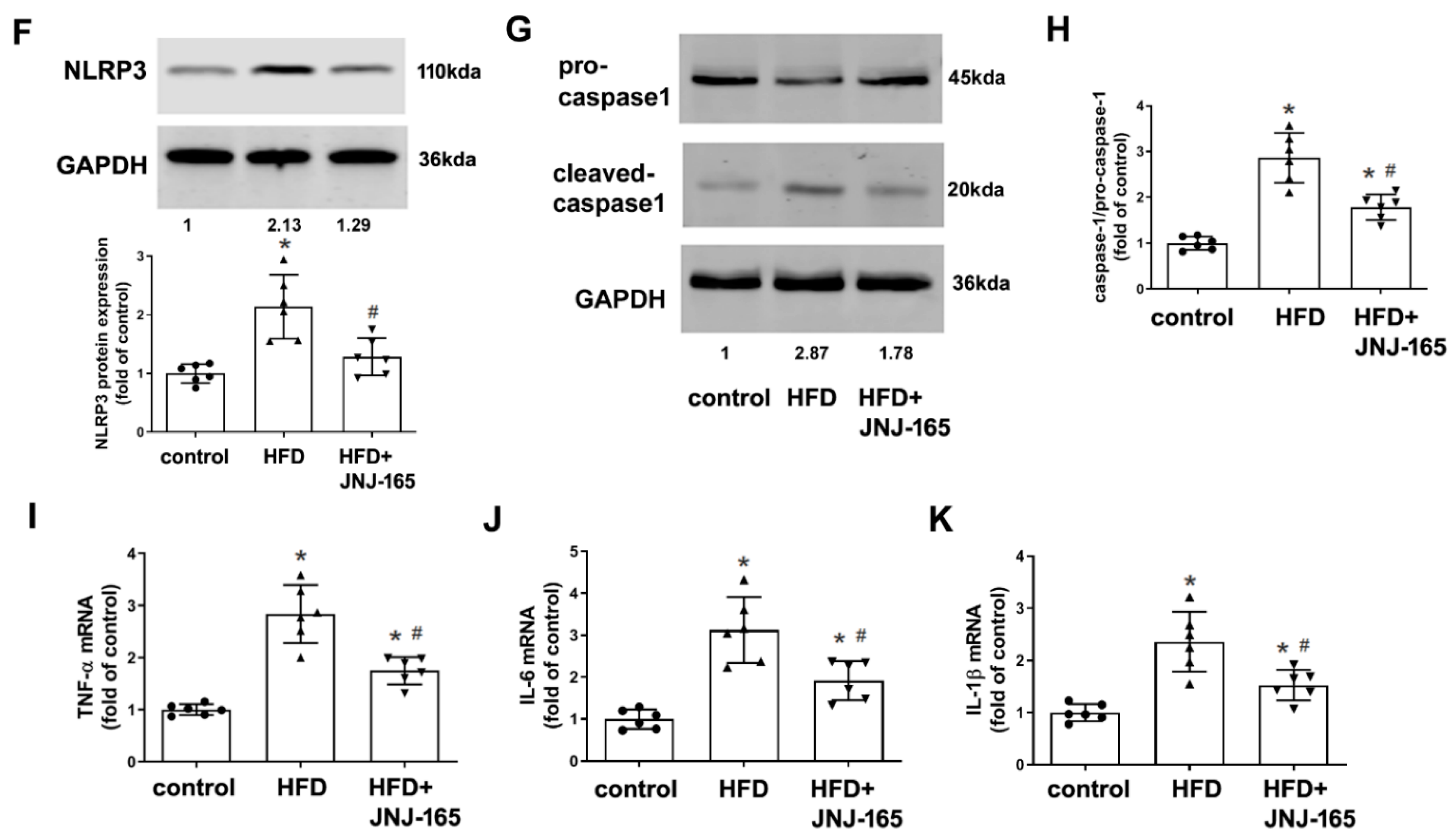
2.9. MDM2 Inhibition Alleviates Mitochondrial Damage and Related Inflammation in HFD-Fed LDLr-/- Mice
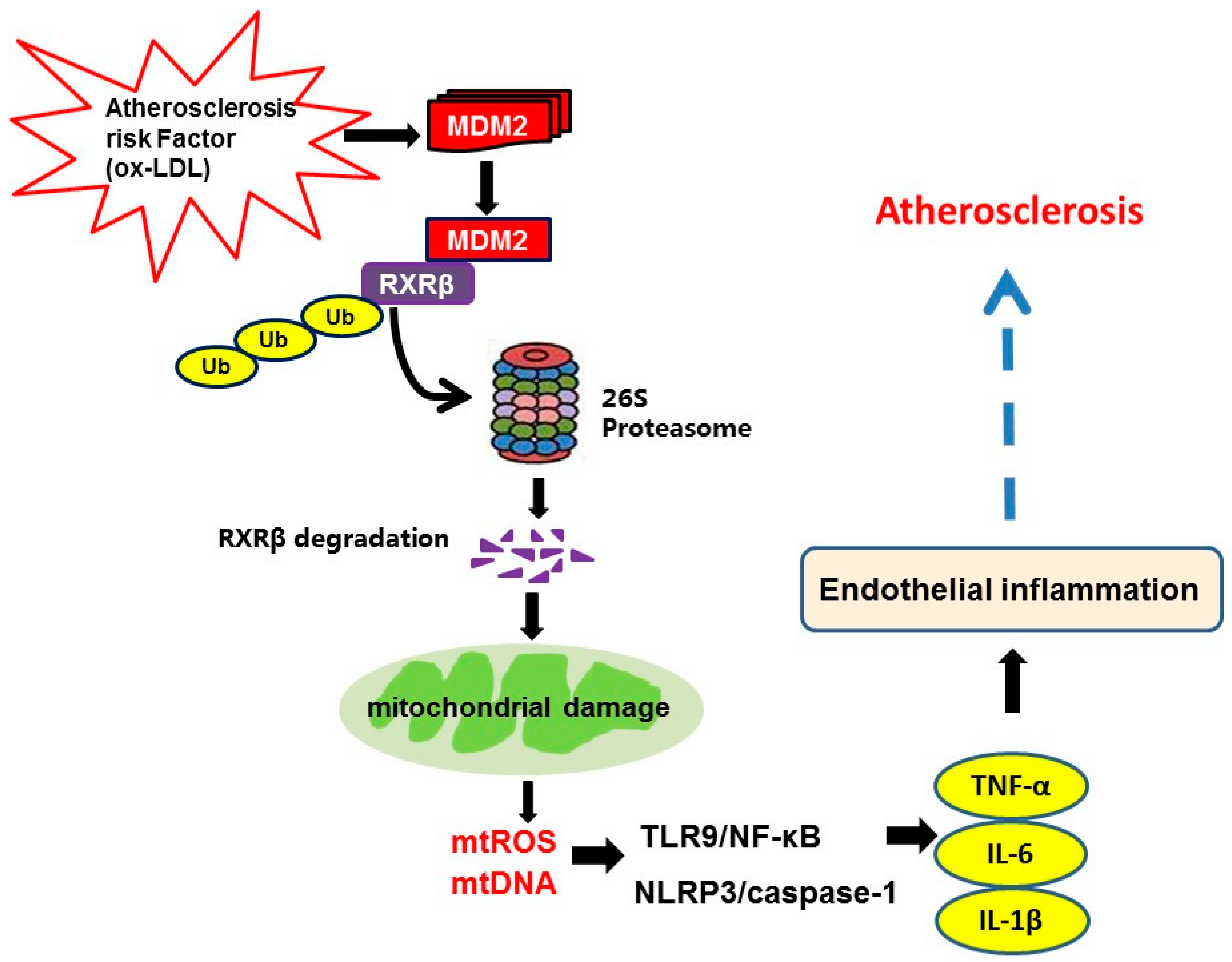
3. Discussion
4. Materials and Methods
Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Negre-Salvayre, A.; Guerby, P.; Gayral, S.; Laffargue, M.; Salvayre, R. Role of reactive oxygen species in atherosclerosis: Lessons from murine genetic models. Free Radic. Biol. Med. 2020, 149, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Khatana, C.; Saini, N.K.; Chakrabarti, S.; Saini, V.; Sharma, A.; Saini, R.V.; Saini, A.K. Mechanistic Insights into the Oxidized Low-Density Lipoprotein-Induced Atherosclerosis. Oxidative Med. Cell. Longev. 2020, 2020, 5245308. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, P.; Benomar, Y.; Staels, B. Retinoid X receptors: Common heterodimerization partners with distinct functions. Trends Endocrinol. Metab. 2010, 21, 676–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, M.; Lu, L.; Wang, Q.; Ma, L.; Tian, X.; Li, C.; Li, C.; Guo, D.; Wang, Q.; Wang, W.; et al. The multi-faceted role of retinoid X receptor in cardiovascular diseases. Biomed. Pharmacother. 2021, 137, 111264. [Google Scholar] [CrossRef]
- Nohara, A.; Kobayashi, J.; Mabuchi, H. Retinoid X receptor heterodimer variants and cardiovascular risk factors. J. Atheroscler. Thromb. 2009, 16, 303–318. [Google Scholar] [CrossRef] [Green Version]
- Staels, B. Regulation of lipid and lipoprotein metabolism by retinoids. J. Am. Acad. Dermatol. 2001, 45, S158–S167. [Google Scholar] [CrossRef]
- Lüsebrink, E.; Warm, V.; Pircher, J.; Ehrlich, A.; Zhang, Z.; Strecker, J.; Chambon, P.; Massberg, S.; Schulz, C.; Petzold, T. Role of RXRβ in platelet function and arterial thrombosis. J. Thromb. Haemost. 2019, 17, 1489–1499. [Google Scholar] [CrossRef]
- Claudel, T.; Leibowitz, M.D.; Fiévet, C.; Tailleux, A.; Wagner, B.; Repa, J.J.; Torpier, G.; Lobaccaro, J.M.; Paterniti, J.R.; Mangelsdorf, D.J.; et al. Reduction of atherosclerosis in apolipoprotein E knockout mice by activation of the retinoid X receptor. Proc. Natl. Acad. Sci. USA 2001, 98, 2610–2615. [Google Scholar] [CrossRef] [Green Version]
- Lalloyer, F.; Fiévet, C.; Lestavel, S.; Torpier, G.; van der Veen, J.; Touche, V.; Bultel, S.; Yous, S.; Kuipers, F.; Paumelle, R.; et al. The RXR agonist bexarotene improves cholesterol homeostasis and inhibits atherosclerosis progression in a mouse model of mixed dyslipidemia. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2731–2737. [Google Scholar] [CrossRef]
- Lin, L.M.; Peng, F.; Liu, Y.P.; Chai, D.J.; Ning, R.B.; Xu, C.D.; Lin, J.X. Coadministration of VDR and RXR agonists synergistically alleviates atherosclerosis through inhibition of oxidative stress: An in vivo and in vitro study. Atherosclerosis 2016, 251, 273–281. [Google Scholar] [CrossRef]
- Rőszer, T.; Menéndez-Gutiérrez, M.P.; Cedenilla, M.; Ricote, M. Retinoid X receptors in macrophage biology. Trends Endocrinol. Metab. 2013, 24, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Shen, L.H.; Hu, L.H.; Ge, H.; Pu, J.; Chai, D.J.; Shao, Q.; Wang, L.i.; Zeng, J.Z.; He, B. Retinoid X receptor agonists inhibit phorbol-12-myristate-13-acetate (PMA)-induced differentiation of monocytic THP-1 cells into macrophages. Mol. Cell. Biochem. 2010, 335, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Sun, Z.; Nie, P.; Yuan, R.; Cai, Z.; Wu, C.; Hu, L.; Jin, S.; Zhou, H.; Zhang, X.; et al. Sulindac-derived retinoid X receptor-α modulator attenuates atherosclerotic plaque progression and destabilization in ApoE-/- mice. Br. J. Pharmacol. 2019, 176, 2559–2572. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Wang, C.Y.; Xu, J.; Xu, X.L. Overexpression of retinoid X receptor beta provides protection against oxidized low-density lipoprotein-induced inflammation via regulating PGC1α-dependent mitochondrial homeostasis in endothelial cells. Biochem. Pharmacol. 2021, 188, 114559. [Google Scholar] [CrossRef] [PubMed]
- Fåhraeus, R.; Olivares-Illana, V. MDM2’s social network. Oncogene 2014, 33, 4365–4376. [Google Scholar] [CrossRef] [PubMed]
- Lam, B.; Roudier, E. Considering the Role of Murine Double Minute 2 in the Cardiovascular System? Front. Cell Dev. Biol. 2019, 7, 320. [Google Scholar] [CrossRef] [PubMed]
- Ihling, C.; Haendeler, J.; Menzel, G.; Hess, R.D.; Fraedrich, G.; Schaefer, H.E.; Zeiher, A.M. Co-expression of p53 and MDM2 in human atherosclerosis: Implications for the regulation of cellularity of atherosclerotic lesions. J. Pathol. 1998, 185, 303–312. [Google Scholar] [CrossRef]
- Kwon, D.H.; Eom, G.H.; Ko, J.H.; Shin, S.; Joung, H.; Choe, N.; Nam, Y.S.; Min, H.K.; Kook, T.; Yoon, S.; et al. MDM2 E3 ligase-mediated ubiquitination and degradation of HDAC1 in vascular calcification. Nat. Commun. 2016, 7, 10492. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Zhang, J.; Wang, C.; Jain, P.P.; Xiong, M.; Shi, X.; Lei, Y.; Chen, S.; Yin, Q.; Thistlethwaite, P.A.; et al. MDM2-Mediated Ubiquitination of Angiotensin-Converting Enzyme 2 Contributes to the Development of Pulmonary Arterial Hypertension. Circulation 2020, 142, 1190–1204. [Google Scholar] [CrossRef]
- Hashimoto, T.; Ichiki, T.; Ikeda, J.; Narabayashi, E.; Matsuura, H.; Miyazaki, R.; Inanaga, K.; Takeda, K.; Sunagawa, K. Inhibition of MDM2 attenuates neointimal hyperplasia via suppression of vascular proliferation and inflammation. Cardiovasc. Res. 2011, 91, 711–719. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Xu, J.; Hua, Y.Q.; Peng, Y.; Xu, X.L. MDM2 contributes to oxidized low-density lipoprotein-induced inflammation through modulation of mitochondrial damage in endothelial cells. Atherosclerosis 2020, 305, 1–9. [Google Scholar] [CrossRef]
- Deshaies, R.J.; Joazeiro, C.A. RING domain E3 ubiquitin ligases. Annu. Rev. Biochem. 2009, 78, 399–434. [Google Scholar] [CrossRef]
- Fang, S.; Jensen, J.P.; Ludwig, R.L.; Vousden, K.H.; Weissman, A.M. Mdm2 is a RING finger-dependent ubiquitin protein ligase for itself and p53. J. Biol. Chem. 2000, 275, 8945–8951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Ko, Y.S.; Park, S.W.; Kim, H.J. P2Y (2)R activation by ATP induces oxLDL-mediated inflammasome activation through modulation of mitochondrial damage in human endothelial cells. Free Radic. Biol. Med. 2019, 136, 109–117. [Google Scholar] [CrossRef]
- Gopinathan, L.; Hannon, D.B.; Peters, J.M.; Heuvel, J.P.V. Regulation of peroxisome proliferator-activated receptor-alpha by MDM2. Toxicol. Sci. 2009, 108, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Metzger, M.B.; Pruneda, J.N.; Klevit, R.E.; Weissman, A.M. RING-type E3 ligases: Master manipulators of E2 ubiquitin-conjugating enzymes and ubiquitination. Biochim. Biophys. Acta. 2014, 1843, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Sdek, P.; Ying, H.; Chang, D.L.F.; Qiu, W.; Zheng, H.; Touitou, R.; Allday, M.J.; Xiao, Z.X. MDM2 promotes proteasome-dependent ubiquitin-independent degradation of retinoblastoma protein. Mol. Cell. 2005, 20, 699–708. [Google Scholar] [CrossRef]
- Min, X.; Zhang, X.; Sun, N.; Acharya, S.; Kim, K.-M. Mdm2-mediated ubiquitination of PKCβII in the nucleus mediates clathrin-mediated endocytic activity. Biochem. Pharmacol. 2019, 170, 113675. [Google Scholar] [CrossRef] [PubMed]
- Roubín, S.R.; Cordero, A. The Two-way Relationship Between Cancer and Atherosclerosis. Rev. Esp. Cardiol. 2019, 72, 487–494. [Google Scholar]
- Boucherat, O.; Vitry, G.; Trinh, I.; Paulin, R.; Provencher, S.; Bonnet, S. The cancer theory of pulmonary arterial hypertension. Pulm. Circ. 2017, 7, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Qin, J.-J.; Rajaei, M.; Li, X.; Yu, X.; Hunt, C.; Zhang, R. Targeting MDM2 for novel molecular therapy: Beyond oncology. Med. Res. Rev. 2020, 40, 856–880. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Patiño, C.; Trotta, A.P.; Chipuk, J.E. MDM2 and mitochondrial function: One complex intersection. Biochem. Pharmacol. 2019, 162, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Arena, G.; Cissé, M.Y.; Pyrdziak, S.; Chatre, L.; Riscal, R.; Fuentes, M.; Arnold, J.J.; Kastner, M.; Gayte, L.; Bertrand-Gaday, C.; et al. Mitochondrial MDM2 Regulates Respiratory Complex I Activity Independently of p53. Mol. Cell 2018, 69, 594–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkholi, R.; Abraham-Enachescu, I.; Trotta, A.P.; Rubio-Patiño, C.; Mohammed, J.N. MDM2 Integrates Cellular Respiration and Apoptotic Signaling through NDUFS1 and the Mitochondrial Network. Mol. Cell 2019, 74, 452–465. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Pan, G.; Chen, Y.; Guo, X.; Liu, T.; Zhang, J.; Yang, X.; Cheng, M.; Gao, H.; Jiang, F. The p53 pathway in vasculature revisited: A therapeutic target for pathological vascular remodeling? Pharmacol. Res. 2021, 169, 105683. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Xu, J.; Zeng, Y.; Chen, L.; Xu, X.L. Polydatin attenuates atherosclerosis in apolipoprotein E-deficient mice: Role of reverse cholesterol transport, Polydatin attenuates atherosclerosis in apolipoprotein E-deficient mice: Role of reverse cholesterol transport. Phytomedicine 2019, 62, 152935. [Google Scholar] [CrossRef]
- Liu, T.T.; Zeng, Y.; Tang, K.; Chen, X.M.; Zhang, W.; Xu, X.L. Dihydromyricetin ameliorates atherosclerosis in LDL receptor deficient mice. Atherosclerosis 2017, 262, 39–50. [Google Scholar] [CrossRef]
- Ding, Y.; Gong, W.W.; Zhang, S.; Shen, J.; Liu, X.; Wang, Y.Q.; Chen, Y.; Meng, G.L. Protective role of sirtuin3 against oxidative stress and NLRP3 inflammasome in cholesterol accumulation and foam cell formation of macrophages with ox-LDL-stimulation. Biochem. Pharmacol. 2021, 192, 114665. [Google Scholar] [CrossRef]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Y.; Cao, J.; Li, C.-X.; Wang, C.-Y.; Wu, R.-M.; Xu, X.-L. MDM2-Mediated Ubiquitination of RXRβ Contributes to Mitochondrial Damage and Related Inflammation in Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 5766. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105766
Zeng Y, Cao J, Li C-X, Wang C-Y, Wu R-M, Xu X-L. MDM2-Mediated Ubiquitination of RXRβ Contributes to Mitochondrial Damage and Related Inflammation in Atherosclerosis. International Journal of Molecular Sciences. 2022; 23(10):5766. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105766
Chicago/Turabian StyleZeng, Yi, Ji Cao, Chun-Xia Li, Chun-Yan Wang, Ruo-Man Wu, and Xiao-Le Xu. 2022. "MDM2-Mediated Ubiquitination of RXRβ Contributes to Mitochondrial Damage and Related Inflammation in Atherosclerosis" International Journal of Molecular Sciences 23, no. 10: 5766. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105766