
Venom Peptide Toxins Targeting the Outer Pore Region of Transient Receptor Potential Vanilloid 1 in Pain: Implications for Analgesic Drug Development

 , , , ,
, , , ,
Abstract
:1. Introduction
2. General Characteristics of TRPV1
2.1. The TRP Superfamily and TRPV Family
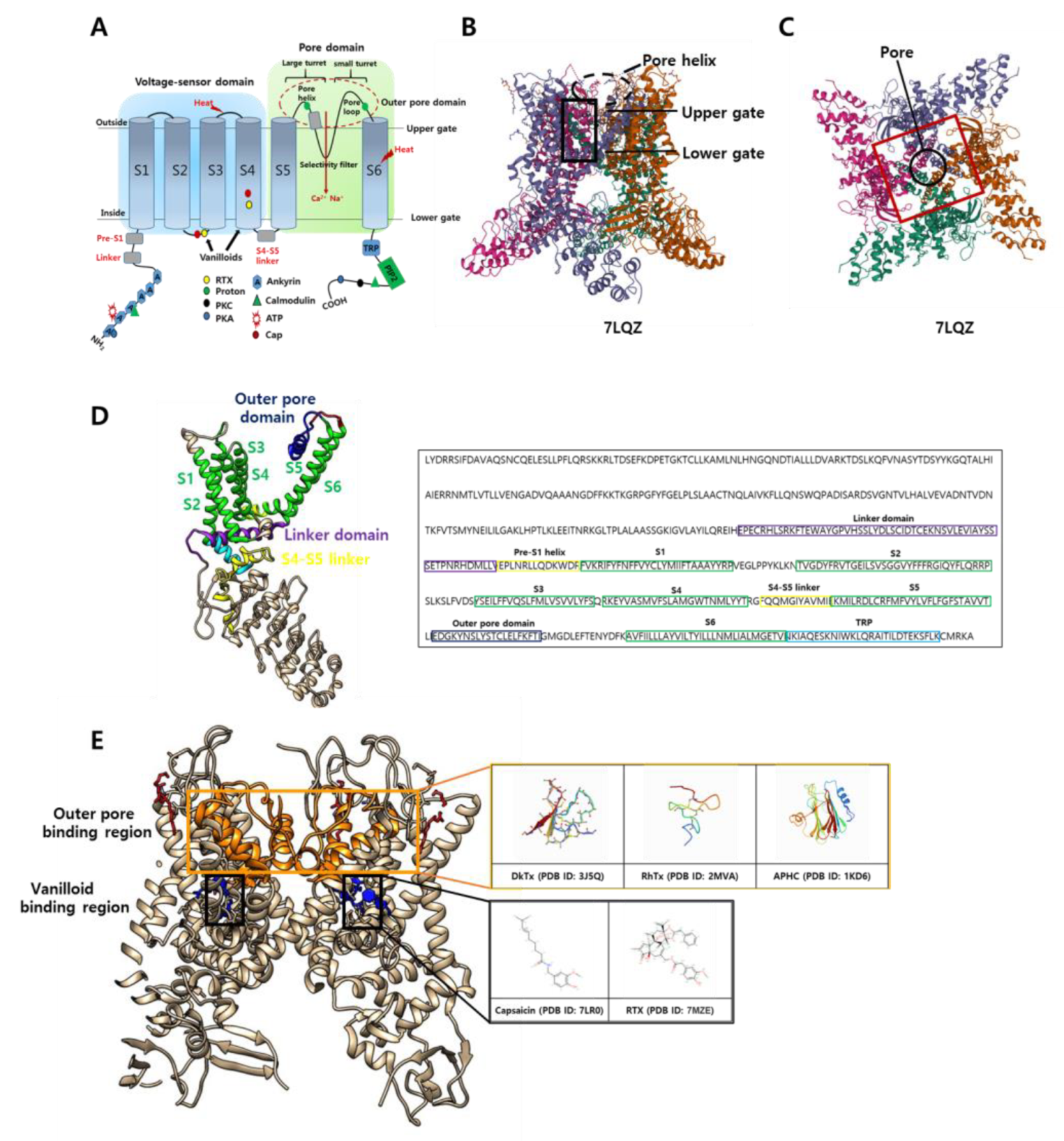
2.2. The Structure of TRPV1 Channel
2.3. The Function of the TRPV1 Channel
2.4. The Mechanism of Heat-Dependent Opening of TRPV1
2.5. The TRPV1 Pore
3. Functional Regulation of TRPV1 by Venom Peptide
3.1. Venom Peptides
3.1.1. Relevance to Pain and Analgesia
3.1.2. The Inhibitor Cystine Knot Domain of Venom Peptides
3.2. TRPV1 Activation by Venom Peptides
3.2.1. DkTx
3.2.2. Vanillotoxins (VaTx1–3)
3.2.3. RhTx
3.2.4. BmP01
3.3. TRPV1 Inhibition by Venom Peptides
3.3.1. Analgesic Polypeptide Heteractis crispa Toxin
3.3.2. Heteractis crispa RG 21
3.4. Complex between TRPV1 and Peptides (Toxins)
3.4.1. Open and Closed State: Dual Gating Mechanism
3.4.2. The Outer Pore Region of TRPV1
3.4.3. The Allosteric Coupling of TRPV1: Upper and Lower Gates
3.4.4. The Two Distinct Binding Sites of TRPV1
4. Functional Regulation of TRPV1 as a Venom Peptide Target
4.1. Side Effects of TRPV1 Inhibition as a Drug Target
4.2. Minimizing Side Effects of TRPV1 Inhibition as a Drug Target
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Benitez-Angeles, M.; Morales-Lazaro, S.L.; Juarez-Gonzalez, E.; Rosenbaum, T. TRPV1: Structure, Endogenous Agonists, and Mechanisms. Int. J. Mol. Sci. 2020, 21, 3421. [Google Scholar] [CrossRef] [PubMed]
- Hazan, A.; Kumar, R.; Matzner, H.; Priel, A. The pain receptor TRPV1 displays agonist-dependent activation stoichiometry. Sci. Rep. 2015, 5, 12278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, R.; Sheth, S.; Mukherjea, D.; Rybak, L.P.; Ramkumar, V. TRPV1: A Potential Drug Target for Treating Various Diseases. Cells 2014, 3, 517–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szolcsanyi, J.; Sandor, Z. Multisteric TRPV1 nocisensor: A target for analgesics. Trends. Pharm. Sci. 2012, 33, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yang, F.; Zhang, B.; Lee, B.H.; Li, B.; Luo, L.; Zheng, J.; Lai, R. A bimodal activation mechanism underlies scorpion toxin-induced pain. Sci. Adv. 2017, 3, e1700810. [Google Scholar] [CrossRef] [Green Version]
- Geron, M.; Hazan, A.; Priel, A. Animal Toxins Providing Insights into TRPV1 Activation Mechanism. Toxins 2017, 9, 326. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Jara-Oseguera, A.; Chang, T.H.; Bae, C.; Hanson, S.M.; Swartz, K.J. Heat activation is intrinsic to the pore domain of TRPV1. Proc. Natl. Acad. Sci. USA 2018, 115, E317–E324. [Google Scholar] [CrossRef] [Green Version]
- Liang, R.; Kawabata, Y.; Kawabata, F.; Nishimura, S.; Tabata, S. Differences in the acidic sensitivity of transient receptor potential vanilloid 1 (TRPV1) between chickens and mice. Biochem. Biophys. Res. Commun. 2019, 515, 386–393. [Google Scholar] [CrossRef]
- Kumar, R.; Geron, M.; Hazan, A.; Priel, A. Endogenous and Exogenous Vanilloids Evoke Disparate TRPV1 Activation to Produce Distinct Neuronal Responses. Front. Pharm. 2020, 11, 903. [Google Scholar] [CrossRef]
- Szallasi, A.; Cruz, F.; Geppetti, P. TRPV1: A therapeutic target for novel analgesic drugs? Trends Mol. Med. 2006, 12, 545–554. [Google Scholar] [CrossRef]
- Ryu, S.; Liu, B.; Yao, J.; Fu, Q.; Qin, F. Uncoupling proton activation of vanilloid receptor TRPV1. J. Neurosci. 2007, 27, 12797–12807. [Google Scholar] [CrossRef] [PubMed]
- Utkin, Y.N. Animal venom studies: Current benefits and future developments. World J. Biol. Chem. 2015, 6, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Xu, S.; Zhang, Y.; Wang, F. Animal protein toxins: Origins and therapeutic applications. Biophys. Rep. 2018, 4, 233–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohlen, C.J.; Julius, D. Receptor-targeting mechanisms of pain-causing toxins: How ow? Toxicon 2012, 60, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Hardy, M.C.; Cochrane, J.; Allavena, R.E. Venomous and poisonous Australian animals of veterinary importance: A rich source of novel therapeutics. Biomed. Res. Int. 2014, 2014, 671041. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, S.M.; Silva, C.R.; Trevisan, G.; Villarinho, J.G.; Cordeiro, M.N.; Richardson, M.; Borges, M.H.; Castro, C.J., Jr.; Gomez, M.V.; Ferreira, J. Antinociceptive effect of a novel armed spider peptide Tx3-5 in pathological pain models in mice. Pflug. Arch. 2016, 468, 881–894. [Google Scholar] [CrossRef]
- Siemens, J.; Zhou, S.; Piskorowski, R.; Nikai, T.; Lumpkin, E.A.; Basbaum, A.I.; King, D.; Julius, D. Spider toxins activate the capsaicin receptor to produce inflammatory pain. Nature 2006, 444, 208–212. [Google Scholar] [CrossRef]
- Rigo, F.K.; Rossato, M.F.; Trevisan, G.; De Pra, S.D.; Ineu, R.P.; Duarte, M.B.; de Castro, C.J., Jr.; Ferreira, J.; Gomez, M.V. PhKv a toxin isolated from the spider venom induces antinociception by inhibition of cholinesterase activating cholinergic system. Scand. J. Pain 2017, 17, 203–210. [Google Scholar]
- Patapoutian, A.; Tate, S.; Woolf, C.J. Transient receptor potential channels: Targeting pain at the source. Nat. Rev. Drug Discov. 2009, 8, 55–68. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.J.; Garcia, M.L. Therapeutic potential of venom peptides. Nat. Rev. Drug Discov. 2003, 2, 790–802. [Google Scholar] [CrossRef]
- Vidya, V.; Achar, R.R.; Himathi, M.U.; Akshita, N.; Kameshwar, V.H.; Byrappa, K.; Ramadas, D. Venom peptides—A comprehensive translational perspective in pain management. Curr. Res. Toxicol. 2021, 2, 329–340. [Google Scholar]
- Laps, S.; Atamleh, F.; Kamnesky, G.; Sun, H.; Brik, A. General synthetic strategy for regioselective ultrafast formation of disulfide bonds in peptides and proteins. Nat. Commun. 2021, 12, 870. [Google Scholar] [CrossRef] [PubMed]
- Nicke, A.; Ulens, C.; Rolland, J.F.; Tsetlin, V.I. Editorial: From Peptide and Protein Toxins to Ion Channel Structure/Function and Drug Design. Front. Pharm. 2020, 11, 548366. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, M.; Undheim, E.A.B.; Mobli, M. Secreted Cysteine-Rich Repeat Proteins “SCREPs”: A Novel Multi-Domain Architecture. Front. Pharm. 2018, 9, 1333. [Google Scholar] [CrossRef]
- Carnevale, V.; Rohacs, T. TRPV1: A Target for Rational Drug Design. Pharmaceuticals 2016, 9, 52. [Google Scholar] [CrossRef]
- Hwang, S.M.; Lee, J.Y.; Park, C.K.; Kim, Y.H. The Role of TRP Channels and PMCA in Brain Disorders: Intracellular Calcium and pH Homeostasis. Front. Cell Dev. Biol. 2021, 9, 584388. [Google Scholar] [CrossRef]
- Brederson, J.D.; Kym, P.R.; Szallasi, A. Targeting TRP channels for pain relief. Eur. J. Pharm. 2013, 716, 61–76. [Google Scholar] [CrossRef]
- Broad, L.M.; Mogg, A.J.; Beattie, R.E.; Ogden, A.M.; Blanco, M.J.; Bleakman, D. TRP channels as emerging targets for pain therapeutics. Expert Opin. Targets 2009, 13, 69–81. [Google Scholar] [CrossRef]
- Gonzalez-Ramirez, R.; Chen, Y.; Liedtke, W.B.; Morales-Lazaro, S.L. TRP Channels and Pain. In Neurobiology of TRP Channels; Emir, T.L.R., Ed.; NCBI: Boca Raton, FL, USA, 2017; pp. 125–147. [Google Scholar]
- Marwaha, L.; Bansal, Y.; Singh, R.; Saroj, P.; Bhandari, R.; Kuhad, A. TRP channels: Potential drug target for neuropathic pain. Inflammopharmacology 2016, 24, 305–317. [Google Scholar] [CrossRef]
- Karki, T.; Tojkander, S. TRPV Protein Family-From Mechanosensing to Cancer Invasion. Biomolecules 2021, 11, 1019. [Google Scholar] [CrossRef]
- Bang, S.; Yoo, S.; Oh, U.; Hwang, S.W. Endogenous lipid-derived ligands for sensory TRP ion channels and their pain modulation. Arch. Pharm. Res. 2010, 33, 1509–1520. [Google Scholar] [CrossRef] [PubMed]
- Cortright, D.N.; Szallasi, A. TRP channels and pain. Curr. Pharm. Des. 2009, 15, 1736–1749. [Google Scholar] [CrossRef]
- Julius, D. TRP channels and pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, J.D.; Alessandri-Haber, N. TRP channels: Targets for the relief of pain. Biochim. Biophys. Acta 2007, 1772, 989–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Duo, L.; Hu, L.; Tian, N.; Cheng, G.; Wang, H.; Lin, Z.; Wang, Y.; Yang, Y. TRPV1 gain-of-function mutation impairs pain and itch sensations in mice. Mol. Pain 2018, 14, 1744806918762031. [Google Scholar] [CrossRef]
- Abbas, M.A. Modulation of TRPV1 channel function by natural products in the treatment of pain. Chem. Biol. Interact. 2020, 330, 109178. [Google Scholar] [CrossRef]
- Rohacs, T. Phosphoinositide regulation of TRPV1 revisited. Pflug. Arch. 2015, 467, 1851–1869. [Google Scholar] [CrossRef] [Green Version]
- Leffler, A.; Fischer, M.J.; Rehner, D.; Kienel, S.; Kistner, K.; Sauer, S.K.; Gavva, N.R.; Reeh, P.W.; Nau, C. The vanilloid receptor TRPV1 is activated and sensitized by local anesthetics in rodent sensory neurons. J. Clin. Investig. 2008, 118, 763–776. [Google Scholar] [CrossRef]
- Jardin, I.; Lopez, J.J.; Diez, R.; Sanchez-Collado, J.; Cantonero, C.; Albarran, L.; Woodard, G.E.; Redondo, P.C.; Salido, G.M.; Smani, T.; et al. TRPs in Pain Sensation. Front. Physiol. 2017, 8, 392. [Google Scholar] [CrossRef] [Green Version]
- Storozhuk, M.V.; Moroz, O.F.; Zholos, A.V. Multifunctional TRPV1 Ion Channels in Physiology and Pathology with Focus on the Brain, Vasculature, and Some Visceral Systems. Biomed. Res. Int. 2019, 2019, 5806321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balleza-Tapia, H.; Crux, S.; Andrade-Talavera, Y.; Dolz-Gaiton, P.; Papadia, D.; Chen, G.; Johansson, J.; Fisahn, A. TrpV1 receptor activation rescues neuronal function and network gamma oscillations from Abeta-induced impairment in mouse hippocampus in vitro. Elife 2018, 7, e37703. [Google Scholar] [CrossRef] [PubMed]
- Marrone, M.C.; Morabito, A.; Giustizieri, M.; Chiurchiu, V.; Leuti, A.; Mattioli, M.; Marinelli, S.; Riganti, L.; Lombardi, M.; Murana, E.; et al. TRPV1 channels are critical brain inflammation detectors and neuropathic pain biomarkers in mice. Nat. Commun. 2017, 8, 15292. [Google Scholar] [CrossRef] [PubMed]
- Silverman, H.A.; Chen, A.; Kravatz, N.L.; Chavan, S.S.; Chang, E.H. Involvement of Neural Transient Receptor Potential Channels in Peripheral Inflammation. Front. Immunol. 2020, 11, 590261. [Google Scholar] [CrossRef] [PubMed]
- Gavva, N.R. Body-temperature maintenance as the predominant function of the vanilloid receptor TRPV1. Trends Pharm. Sci. 2008, 29, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Du, G.; Tian, Y.; Yao, Z.; Vu, S.; Zheng, J.; Chai, L.; Wang, K.; Yang, S. A specialized pore turret in the mammalian cation channel TRPV1 is responsible for distinct and species-specific heat activation thresholds. J. Biol. Chem. 2020, 295, 9641–9649. [Google Scholar] [CrossRef] [PubMed]
- Yonghak, P.; Miyata, S.; Kurganov, E. TRPV1 is crucial for thermal homeostasis in the mouse by heat loss behaviors under warm ambient temperature. Sci. Rep. 2020, 10, 8799. [Google Scholar] [CrossRef]
- Xia, R.; Dekermendjian, K.; Lullau, E.; Dekker, N. TRPV1: A therapy target that attracts the pharmaceutical interests. Adv. Exp. Med. Biol. 2011, 704, 637–665. [Google Scholar]
- Mishra, S.K.; Tisel, S.M.; Orestes, P.; Bhangoo, S.K.; Hoon, M.A. TRPV1-lineage neurons are required for thermal sensation. EMBO J. 2011, 30, 582–593. [Google Scholar] [CrossRef] [Green Version]
- Kwon, D.H.; Zhang, F.; Suo, Y.; Bouvette, J.; Borgnia, M.J.; Lee, S.Y. Heat-dependent opening of TRPV1 in the presence of capsaicin. Nat. Struct. Mol. Biol. 2021, 28, 554–563. [Google Scholar] [CrossRef]
- Kasimova, M.A.; Yazici, A.; Yudin, Y.; Granata, D.; Klein, M.L.; Rohacs, T.; Carnevale, V. Ion Channel Sensing: Are Fluctuations the Crux of the Matter? J. Phys. Chem. Lett. 2018, 9, 1260–1264. [Google Scholar] [CrossRef]
- Kasimova, M.A.; Yazici, A.T.; Yudin, Y.; Granata, D.; Klein, M.L.; Rohacs, T.; Carnevale, V. A hypothetical molecular mechanism for TRPV1 activation that invokes rotation of an S6 asparagine. J. Gen. Physiol. 2018, 150, 1554–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geron, M.; Kumar, R.; Zhou, W.; Faraldo-Gomez, J.D.; Vasquez, V.; Priel, A. TRPV1 pore turret dictates distinct DkTx and capsaicin gating. Proc. Natl. Acad. Sci. USA 2018, 115, E11837–E11846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellmich, U.A.; Gaudet, R. High-resolution views of TRPV1 and their implications for the TRP channel superfamily. Handb. Exp. Pharm. 2014, 223, 991–1004. [Google Scholar]
- Gladkikh, I.N.; Sintsova, O.V.; Leychenko, E.V.; Kozlov, S.A. TRPV1 Ion Channel: Structural Features, Activity Modulators, and Therapeutic Potential. Biochemistry 2021, 86 (Suppl. 1), S50–S70. [Google Scholar] [CrossRef]
- Winter, Z.; Buhala, A.; Otvos, F.; Josvay, K.; Vizler, C.; Dombi, G.; Szakonyi, G.; Olah, Z. Functionally important amino acid residues in the transient receptor potential vanilloid 1 (TRPV1) ion channel—An overview of the current mutational data. Mol. Pain 2013, 9, 30. [Google Scholar] [CrossRef] [Green Version]
- King, G.F. Venoms as a platform for human drugs: Translating toxins into therapeutics. Expert Opin. Biol. 2011, 11, 1469–1484. [Google Scholar] [CrossRef]
- Chen, Y.N.; Li, K.C.; Li, Z.; Shang, G.W.; Liu, D.N.; Lu, Z.M.; Zhang, J.W.; Ji, Y.H.; Gao, G.D.; Chen, J. Effects of bee venom peptidergic components on rat pain-related behaviors and inflammation. Neuroscience 2006, 138, 631–640. [Google Scholar] [CrossRef]
- Herzig, V.; Cristofori-Armstrong, B.; Israel, M.R.; Nixon, S.A.; Vetter, I.; King, G.F. Animal toxins—Nature’s evolutionary-refined toolkit for basic research and drug discovery. Biochem. Pharm. 2020, 181, 114096. [Google Scholar] [CrossRef]
- Pineda, S.S.; Chin, Y.K.; Undheim, E.A.B.; Senff, S.; Mobli, M.; Dauly, C.; Escoubas, P.; Nicholson, G.M.; Kaas, Q.; Guo, S.; et al. Structural venomics reveals evolution of a complex venom by duplication and diversification of an ancient peptide-encoding gene. Proc. Natl. Acad. Sci. USA 2020, 117, 11399–11408. [Google Scholar] [CrossRef]
- Craik, D.J.; Daly, N.L.; Waine, C. The cystine knot motif in toxins and implications for drug design. Toxicon 2001, 39, 43–60. [Google Scholar] [CrossRef]
- Menez, A. Functional architectures of animal toxins: A clue to drug design? Toxicon 1998, 36, 1557–1572. [Google Scholar] [CrossRef]
- Rosengren, K.J.; Daly, N.L.; Plan, M.R.; Waine, C.; Craik, D.J. Twists, knots, and rings in proteins. Structural definition of the cyclotide framework. J. Biol. Chem. 2003, 278, 8606–8616. [Google Scholar] [CrossRef] [Green Version]
- Iyer, S.; Acharya, K.R. Tying the knot: The cystine signature and molecular-recognition processes of the vascular endothelial growth factor family of angiogenic cytokines. FEBS J. 2011, 278, 4304–4322. [Google Scholar] [CrossRef] [Green Version]
- Kimura, T. Stability and Safety of Inhibitor Cystine Knot Peptide, GTx1-15, from the Tarantula Spider Grammostola rosea. Toxins 2021, 13, 621. [Google Scholar] [CrossRef]
- Wu, T.; Wang, M.; Wu, W.; Luo, Q.; Jiang, L.; Tao, H.; Deng, M. Spider venom peptides as potential drug candidates due to their anticancer and antinociceptive activities. J. Venom. Anim. Toxins Incl. Trop. Dis. 2019, 25, e146318. [Google Scholar] [CrossRef]
- Bae, C.; Anselmi, C.; Kalia, J.; Jara-Oseguera, A.; Schwieters, C.D.; Krepkiy, D.; Won Lee, C.; Kim, E.H.; Kim, J.I.; Faraldo-Gomez, J.D.; et al. Structural insights into the mechanism of activation of the TRPV1 channel by a membrane-bound tarantula toxin. Elife 2016, 5, e11273. [Google Scholar] [CrossRef]
- Bae, C.; Kalia, J.; Song, I.; Yu, J.; Kim, H.H.; Swartz, K.J.; Kim, J.I. High yield production and refolding of the double-knot toxin, an activator of TRPV1 channels. PLoS ONE 2012, 7, e51516. [Google Scholar] [CrossRef] [Green Version]
- Bohlen, C.J.; Priel, A.; Zhou, S.; King, D.; Siemens, J.; Julius, D. A bivalent tarantula toxin activates the capsaicin receptor, TRPV1, by targeting the outer pore domain. Cell 2010, 141, 834–845. [Google Scholar] [CrossRef] [Green Version]
- Mihailescu, M.; Krepkiy, D.; Milescu, M.; Gawrisch, K.; Swartz, K.J.; White, S. Structural interactions of a voltage sensor toxin with lipid membranes. Proc. Natl. Acad. Sci. USA 2014, 111, E5463–E5470. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Xu, L.; Lee, B.H.; Xiao, X.; Yarov-Yarovoy, V.; Zheng, J. An Unorthodox Mechanism Underlying Voltage Sensitivity of TRPV1 Ion Channel. Adv. Sci. 2020, 7, 2000575. [Google Scholar] [CrossRef]
- Elokely, K.; Velisetty, P.; Delemotte, L.; Palovcak, E.; Klein, M.L.; Rohacs, T.; Carnevale, V. Understanding TRPV1 activation by ligands: Insights from the binding modes of capsaicin and resiniferatoxin. Proc. Natl. Acad. Sci. USA 2016, 113, E137–E145. [Google Scholar] [CrossRef] [Green Version]
- Vu, S.; Singh, V.; Wulff, H.; Yarov-Yarovoy, V.; Zheng, J. New capsaicin analogs as molecular rulers to define the permissive conformation of the mouse TRPV1 ligand-binding pocket. Elife 2020, 9, e62039. [Google Scholar] [CrossRef]
- Chu, Y.; Cohen, B.E.; Chuang, H.H. A single TRPV1 amino acid controls species sensitivity to capsaicin. Sci. Rep. 2020, 10, 8038. [Google Scholar] [CrossRef]
- Nadezhdin, K.D.; Neuberger, A.; Nikolaev, Y.A.; Murphy, L.A.; Gracheva, E.O.; Bagriantsev, S.N.; Sobolevsky, A.I. Extracellular cap domain is an essential component of the TRPV1 gating mechanism. Nat. Commun. 2021, 12, 2154. [Google Scholar] [CrossRef]
- Gao, Y.; Cao, E.; Julius, D.; Cheng, Y. TRPV1 structures in nanodiscs reveal mechanisms of ligand and lipid action. Nature 2016, 534, 347–351. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, D.; Singh, Y.; Kalia, J. Protein-Lipid Interfaces Can Drive the Functions of Membrane-Embedded Protein-Protein Complexes. ACS Chem. Biol. 2018, 13, 2689–2698. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Yang, F.; Wei, N.; Hong, J.; Li, B.; Luo, L.; Rong, M.; Yarov-Yarovoy, V.; Zheng, J.; Wang, K.; et al. A pain-inducing centipede toxin targets the heat activation machinery of nociceptor TRPV1. Nat. Commun. 2015, 6, 8297. [Google Scholar] [CrossRef] [Green Version]
- Zhu, A.; Aierken, A.; Yao, Z.; Vu, S.; Tian, Y.; Zheng, J.; Yang, S.; Yang, F. A centipede toxin causes rapid desensitization of nociceptor TRPV1 ion channel. Toxicon 2020, 178, 41–49. [Google Scholar] [CrossRef]
- Zheng, J.; Ma, L. Structure and function of the thermoTRP channel pore. Curr. Top. Membr. 2014, 74, 233–257. [Google Scholar]
- Hakim, M.A.; Jiang, W.; Luo, L.; Li, B.; Yang, S.; Song, Y.; Lai, R. Scorpion Toxin, BmP01, Induces Pain by Targeting TRPV1 Channel. Toxins 2015, 7, 3671–3687. [Google Scholar] [CrossRef] [Green Version]
- Nikolaev, M.V.; Dorofeeva, N.A.; Komarova, M.S.; Korolkova, Y.V.; Andreev, Y.A.; Mosharova, I.V.; Grishin, E.V.; Tikhonov, D.B.; Kozlov, S.A. TRPV1 activation power can switch an action mode for its polypeptide ligands. PLoS ONE 2017, 12, e0177077. [Google Scholar] [CrossRef]
- Andreev, Y.A.; Kozlov, S.A.; Koshelev, S.G.; Ivanova, E.A.; Monastyrnaya, M.M.; Kozlovskaya, E.P.; Grishin, E.V. Analgesic compound from sea anemone Heteractis crispa is the first polypeptide inhibitor of vanilloid receptor 1 (TRPV1). J. Biol. Chem. 2008, 283, 23914–23921. [Google Scholar] [CrossRef] [Green Version]
- Andreev, Y.A.; Kozlov, S.A.; Korolkova, Y.V.; Dyachenko, I.A.; Bondarenko, D.A.; Skobtsov, D.I.; Murashev, A.N.; Kotova, P.D.; Rogachevskaja, O.A.; Kabanova, N.V.; et al. Polypeptide modulators of TRPV1 produce analgesia without hyperthermia. Mar. Drugs 2013, 11, 5100–5115. [Google Scholar] [CrossRef] [Green Version]
- Dyachenko, I.A.; Andreev, Y.A.; Logashina, Y.A.; Murashev, A.N.; Grishin, E.V. Biological activity of a polypeptide modulator of TRPV1 receptor. Dokl. Biol. Sci. 2015, 465, 279–281. [Google Scholar] [CrossRef]
- Monastyrnaya, M.; Peigneur, S.; Zelepuga, E.; Sintsova, O.; Gladkikh, I.; Leychenko, E.; Isaeva, M.; Tytgat, J.; Kozlovskaya, E. Kunitz-Type Peptide HCRG21 from the Sea Anemone Heteractis crispa Is a Full Antagonist of the TRPV1 Receptor. Mar. Drugs 2016, 14, 229. [Google Scholar] [CrossRef]
- Sintsova, O.; Gladkikh, I.; Klimovich, A.; Palikova, Y.; Palikov, V.; Styshova, O.; Monastyrnaya, M.; Dyachenko, I.; Kozlov, S.; Leychenko, E. TRPV1 Blocker HCRG21 Suppresses TNF-alpha Production and Prevents the Development of Edema and Hypersensitivity in Carrageenan-Induced Acute Local Inflammation. Biomedicines 2021, 9, 716. [Google Scholar] [CrossRef]
- Peigneur, S.; Tytgat, J. Toxins in Drug Discovery and Pharmacology. Toxins 2018, 10, 126. [Google Scholar] [CrossRef] [Green Version]
- Diochot, S. Pain-related toxins in scorpion and spider venoms: A face to face with ion channels. J. Venom. Anim. Toxins Incl. Trop. Dis. 2021, 27, e20210026. [Google Scholar] [CrossRef]
- Karbat, I.; Altman-Gueta, H.; Fine, S.; Szanto, T.; Hamer-Rogotner, S.; Dym, O.; Frolow, F.; Gordon, D.; Panyi, G.; Gurevitz, M.; et al. Pore-modulating toxins exploit inherent slow inactivation to block K(+) channels. Proc. Natl. Acad. Sci. USA 2019, 116, 18700–18709. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; Li, Y.; Liu, L.; Ren, S.; Tian, Y.; Yang, F. Molecular mechanism underlying modulation of TRPV1 heat activation by polyols. J. Biol. Chem. 2021, 297, 100806. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zheng, J. Understand spiciness: Mechanism of TRPV1 channel activation by capsaicin. Protein Cell 2017, 8, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, H.; Zheng, W. Decrypting the Heat Activation Mechanism of TRPV1 Channel by Molecular Dynamics Simulation. Biophys. J. 2018, 114, 40–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Franulic, I.; Caceres-Molina, J.; Sepulveda, R.V.; Gonzalez-Nilo, F.; Latorre, R. Structure-Driven Pharmacology of Transient Receptor Potential Channel Vanilloid 1. Mol. Pharm. 2016, 90, 300–308. [Google Scholar] [CrossRef]
- Cromer, B.A.; McIntyre, P. Painful toxins acting at TRPV1. Toxicon 2008, 51, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Premkumar, L.S. Targeting TRPV1 as an alternative approach to narcotic analgesics to treat chronic pain conditions. AAPS J. 2010, 12, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Cortright, D.N.; Krause, J.E.; Broom, D.C. TRP channels and pain. Biochim. Biophys. Acta 2007, 1772, 978–988. [Google Scholar] [CrossRef] [Green Version]
- Boukalova, S.; Teisinger, J.; Vlachova, V. Protons stabilize the closed conformation of gain-of-function mutants of the TRPV1 channel. Biochim. Biophys. Acta 2013, 1833, 520–528. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Xiao, X.; Lee, B.H.; Vu, S.; Yang, W.; Yarov-Yarovoy, V.; Zheng, J. The conformational wave in capsaicin activation of transient receptor potential vanilloid 1 ion channel. Nat. Commun. 2018, 9, 2879. [Google Scholar] [CrossRef]
- Zheng, W.; Wen, H. Heat activation mechanism of TRPV1: New insights from molecular dynamics simulation. Temperature 2019, 6, 120–131. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; Qiu, P.; Yu, R. Centipede Venom Peptides Acting on Ion Channels. Toxins 2020, 12, 230. [Google Scholar] [CrossRef] [Green Version]
- Velisetty, P.; Stein, R.A.; Sierra-Valdez, F.J.; Vasquez, V.; Cordero-Morales, J.F. Expression and Purification of the Pain Receptor TRPV1 for Spectroscopic Analysis. Sci. Rep. 2017, 7, 9861. [Google Scholar] [CrossRef] [PubMed]
- Szallasi, A.; Cortright, D.N.; Blum, C.A.; Eid, S.R. The vanilloid receptor TRPV1: 10 years from channel cloning to antagonist proof-of-concept. Nat. Rev. Drug Discov. 2007, 6, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Goswami, C.; Schmidt, H.; Hucho, F. TRPV1 at nerve endings regulates growth cone morphology and movement through cytoskeleton reorganization. FEBS J. 2007, 274, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Frias, B.; Merighi, A. Capsaicin, Nociception and Pain. Molecules 2016, 21, 797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koivisto, A.P.; Belvisi, M.G.; Gaudet, R.; Szallasi, A. Advances in TRP channel drug discovery: From target validation to clinical studies. Nat. Rev. Drug. Discov. 2021, 21, 1–19. [Google Scholar] [CrossRef]
- Huang, S.; Szallasi, A. Transient Receptor Potential (TRP) Channels in Drug Discovery: Old Concepts & New Thoughts. Pharmaceuticals 2017, 10, 64. [Google Scholar]
- Szymaszkiewicz, A.; Wlodarczyk, J.; Wasilewski, A.; Di Marzo, V.; Storr, M.; Fichna, J.; Zielinska, M. Desensitization of transient receptor potential vanilloid type-1 (TRPV1) channel as promising therapy of irritable bowel syndrome: Characterization of the action of palvanil in the mouse gastrointestinal tract. Naunyn. Schmiedebergs. Arch. Pharm. 2020, 393, 1357–1364. [Google Scholar] [CrossRef] [Green Version]
- Szallasi, A.; Sheta, M. Targeting TRPV1 for pain relief: Limits, losers and laurels. Expert Opin. Investig. Drugs 2012, 21, 1351–1369. [Google Scholar] [CrossRef]
- Gunthorpe, M.J.; Chizh, B.A. Clinical development of TRPV1 antagonists: Targeting a pivotal point in the pain pathway. Drug Discov. Today 2009, 14, 56–67. [Google Scholar] [CrossRef]
- Gavva, N.R.; Bannon, A.W.; Hovland, D.N., Jr.; Lehto, S.G.; Klionsky, L.; Surapaneni, S.; Immke, D.C.; Henley, C.; Arik, L.; Bak, A.; et al. Repeated administration of vanilloid receptor TRPV1 antagonists attenuates hyperthermia elicited by TRPV1 blockade. J. Pharm. Exp. 2007, 323, 128–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garami, A.; Shimansky, Y.P.; Pakai, E.; Oliveira, D.L.; Gavva, N.R.; Romanovsky, A.A. Contributions of different modes of TRPV1 activation to TRPV1 antagonist-induced hyperthermia. J. Neurosci. 2010, 30, 1435–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garami, A.; Shimansky, Y.P.; Rumbus, Z.; Vizin, R.C.L.; Farkas, N.; Hegyi, J.; Szakacs, Z.; Solymar, M.; Csenkey, A.; Chiche, D.A.; et al. Hyperthermia induced by transient receptor potential vanilloid-1 (TRPV1) antagonists in human clinical trials: Insights from mathematical modeling and meta-analysis. Pharmacol. Ther. 2020, 208, 107474. [Google Scholar] [CrossRef] [PubMed]
- Garami, A.; Pakai, E.; McDonald, H.A.; Reilly, R.M.; Gomtsyan, A.; Corrigan, J.J.; Pinter, E.; Zhu, D.X.D.; Lehto, S.G.; Gavva, N.R.; et al. TRPV1 antagonists that cause hypothermia, instead of hyperthermia, in rodents: Compounds’ pharmacological profiles, in vivo targets, thermoeffectors recruited and implications for drug development. Acta. Physiol. 2018, 223, e13038. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Ligands | Binding Region | Binding Type | Pain Condition | Reference |
|---|---|---|---|---|
| Capsaicin | Vanilloid-binding pocket | Hydrogen, Van der Waals bond | Pain sensation | [73,74,75] |
| RTX | Vanilloid-binding pocket | Hydrogen bond | Pain sensation | [73,76] |
| Proton | Outer pore domain | Hydrogen bond | Pain sensation | [36] |
| Double-knot toxin (DkTx) | Outer pore domain | Hydrophobic interaction | Pain sensation | [6,54,68,70,71,77,78] |
| Vanillotoxins (VaTx1–3) | Outer pore domain | Electrostatic interaction | Pain sensation | [5,6,14] |
| Scolopendra subspinipes mutilans toxin RhTx | Outer pore domain | Electrostatic and hydrophobic interactions | Pain sensation | [2,79,80] |
| Pain-inducing peptide (BmP01) | Outer pore domain | Electrostatic interaction | Pain sensation | [5,81,82] |
| Analgesic polypeptide Heteractis crispa (APHC) toxin | Outer pore domain | No direct evidence | Analgesic action | [83,84,85,86] |
| Heteractis crispa RG 21 (HCRG21) | Outer pore domain | No direct evidence | Analgesic action | [87,88] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, S.-M.; Jo, Y.-Y.; Cohen, C.F.; Kim, Y.-H.; Berta, T.; Park, C.-K. Venom Peptide Toxins Targeting the Outer Pore Region of Transient Receptor Potential Vanilloid 1 in Pain: Implications for Analgesic Drug Development. Int. J. Mol. Sci. 2022, 23, 5772. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105772
Hwang S-M, Jo Y-Y, Cohen CF, Kim Y-H, Berta T, Park C-K. Venom Peptide Toxins Targeting the Outer Pore Region of Transient Receptor Potential Vanilloid 1 in Pain: Implications for Analgesic Drug Development. International Journal of Molecular Sciences. 2022; 23(10):5772. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105772
Chicago/Turabian StyleHwang, Sung-Min, Youn-Yi Jo, Cinder Faith Cohen, Yong-Ho Kim, Temugin Berta, and Chul-Kyu Park. 2022. "Venom Peptide Toxins Targeting the Outer Pore Region of Transient Receptor Potential Vanilloid 1 in Pain: Implications for Analgesic Drug Development" International Journal of Molecular Sciences 23, no. 10: 5772. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105772