
Metabolic Profiles Reveal Changes in the Leaves and Roots of Rapeseed (Brassica napus L.) Seedlings under Nitrogen Deficiency

Abstract
:1. Introduction
2. Results
2.1. Phenotypic Characterization of Rapeseed between the HN and LN Treatments
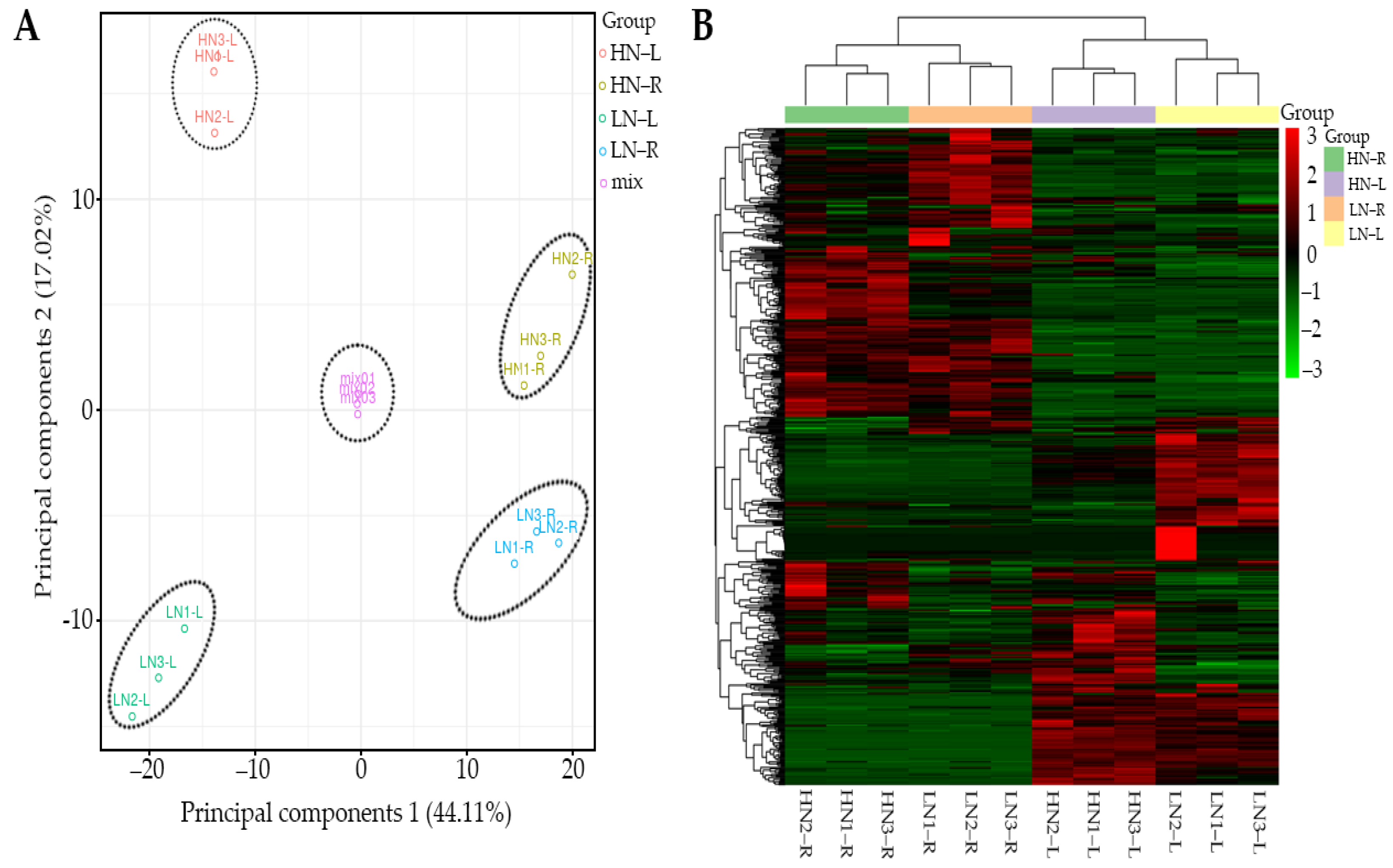
2.2. Multivariate Analysis of Identified Metabolites
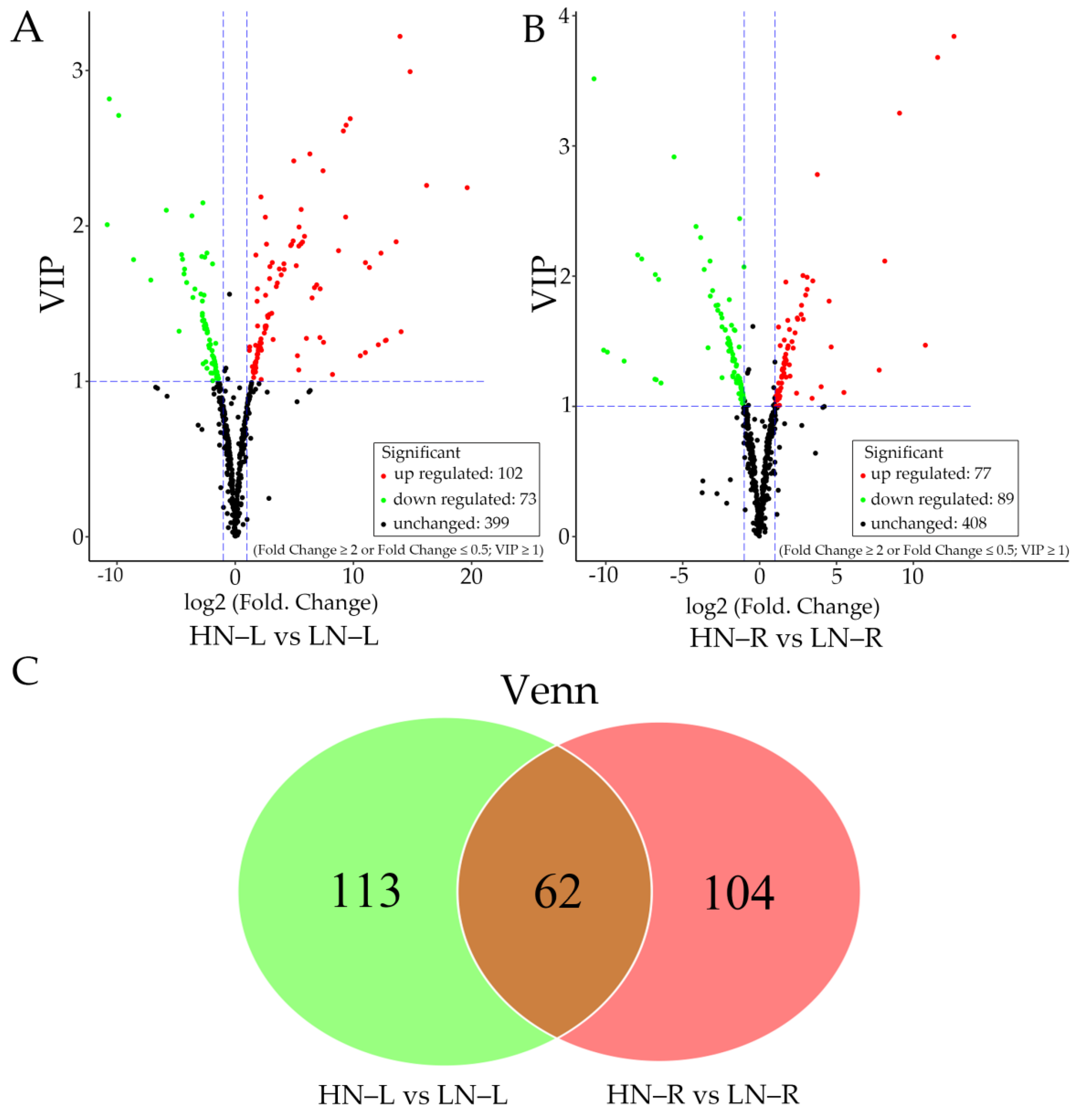
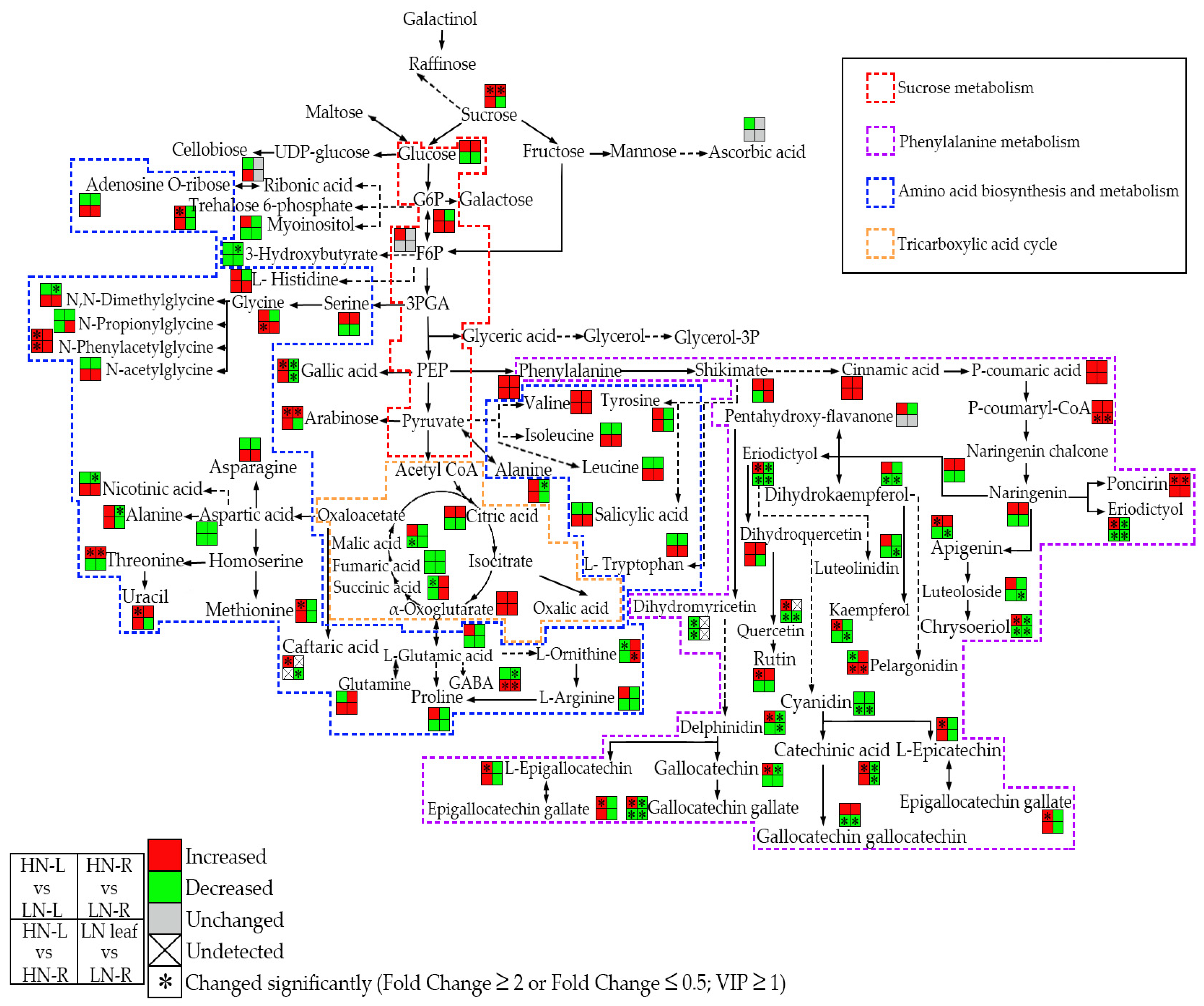
2.3. Metabolic Changes in the Leaves and Roots of Rapeseed during the HN and LN Treatments
2.4. KEGG Analysis
2.5. LN Treatment Increased Flavonoid and Sugar Accumulation in Rapeseed
2.6. LN Treatment Affects Amino Acids and Their Derivative Metabolites in Rapeseed
2.7. LN Treatment Alter Phytohormone Accumulation in Rapeseed
2.8. LN Treatment Can Modulate the Carbon-Nitrogen Metabolite Balance in the Leaves and Roots of Rapeseed and Increase the Accumulation of Multiple Stress-Related Metabolites
3. Discussion
3.1. N Deficiency Led to Architectural Changes in Rapeseed Roots
3.2. N Deficiency Led to Substantial Changes in Metabolite Contents in the Leaves and Roots of Rapeseed
3.3. Flavonoids Play an Important Protective Role in Rapeseed under N Starvation Conditions
3.4. Amino Acids and Their Derivative Metabolites May Enhance Rapeseed Resistance to N Starvation Conditions
3.5. Plant Hormones Function Synergistically to Regulate and Maintain Rapeseed Growth under N Starvation
4. Conclusions
5. Materials and Methods
5.1. Rapeseed Seedlings and N-Deficient Treatment
5.2. Phenotypic Characterization
5.3. Plant Sample Preparation and Extraction of Secondary Metabolites
5.4. LC–MS Analysis
5.5. Metabolite Profiling
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| C | Carbon |
| N | Nitrogen |
| HN | Normal nitrogen |
| LN | Nitrogen deficient |
| VIP | Variable importance in projection |
| PLS-DA | Partial least squares discriminant analysis |
| OPLS-DA | Orthogonal partial least squares discriminant analysis |
| DAMs | Differentially accumulated metabolites |
| HCA | Hierarchical cluster analysis |
| CEPs | C-terminally encoded peptides |
| PCC | Pearson correlation coefficients |
| PCA | Principal component analysis |
| IAA | Indole-3-acetic acid |
| ICA | Indole-carboxylic acid |
| ZT | Trans-zeatin 9-O-glucoside |
| IAA | Indole 3-acetic acid |
| GA | Gibberellin |
| JA | Jasmonic acid |
| SA | Salicylic acid |
| ABA | Abscisic acid |
| ROS | Reactive oxygen species |
| KEGG | Kyoto encyclopedia of genes and genomes |
| RL | Root length |
| RS | Root surface area |
| RV | Root volume |
| RD | Root average diameter |
| LC-MS | Liquid chromatographic mass spectrometry |
References
- Schluter, U.; Mascher, M.; Colmsee, C.; Scholz, U.; Brautigam, A.; Fahnenstich, H.; Sonnewald, U. Maize Source Leaf Adaptation to Nitrogen Deficiency Affects Not Only Nitrogen and Carbon Metabolism But Also Control of Phosphate Homeostasis. Plant Physiol. 2012, 160, 1384–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kant, S. Understanding Nitrate Uptake, Signaling and Remobilisation for Improving Plant Nitrogen Use Efficiency. Semin. Cell Dev. Biol. 2018, 74, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Walk, T.C.; Han, P.; Chen, L.; Zhang, S.; Li, Y.; Hu, X.; Xie, L.; Yang, Y.; Liu, J.; et al. Adaption of Roots to Nitrogen Deficiency Revealed by 3D Quantification and Proteomic Analysis. Plant Physiol. 2018, 179, 329–347. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Zheng, H.; Wu, Y.; Xiao, Y.; Du, Y.; Xu, W.; Lu, F.; Wang, X.; Ouyang, Z. Changes in Nitrogen Budget and Potential Risk to the Environment Over 20 Years (1990–2010) in the Agroecosystems of the Haihe Basin, China. J. Environ. Sci. 2015, 28, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Giehl, R.; Meyer, R.C.; Altmann, T.; Von Wirén, N. Natural Variation of BSK3 Tunes Brassinosteroid Signaling to Regulate Root Foraging under Low Nitrogen. Nat. Commun. 2019, 10, 2378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flis, A.; Mengin, V.; Ivakov, A.A.; Mugford, S.T.; Hubberten, H.-M.; Encke, B.; Krohn, N.; Höhne, M.; Feil, R.; Hoefgen, R.; et al. Multiple Circadian Clock Outputs Regulate Diel Turnover of Carbon and Nitrogen Reserves. Plant Cell Environ. 2018, 42, 549–573. [Google Scholar] [CrossRef]
- Patzke, K.; Prananingrum, P.; Klemens, P.A.W.; Trentmann, O.; Martins Rodrigues, C.; Keller, I.; Fernie, A.R.; Geigenberger, P.; Bölter, B.; Lehmann, M.; et al. The Plastidic Sugar Transporter pSuT Influences Flowering and Affects Cold Responses. Plant Physiol. 2018, 179, 569–587. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Constabel, C.P. MYB Repressors as Regulators of Phenylpropanoid Metabolism in Plants. Trends Plant Sci. 2019, 24, 275–289. [Google Scholar] [CrossRef]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.-Y.; Li, J.; Wang, P.-Y.; Qin, F.; et al. Plant Abiotic Stress Response and Nutrient Use Efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Mounier, E.; Pervent, M.; Ljung, K.; Gojon, A.; Nacry, P. Cell and Environment: Auxin-Mediated Nitrate Signalling by NRT1.1 Participates in the Adaptive Response of Arabidopsis Root Architecture to the Spatial Heterogeneity of Nitrate Availability. Plant Cell Environ. 2013, 37, 162–174. [Google Scholar] [CrossRef]
- Kajikawa, M.; Sawaragi, Y.; Shinkawa, H.; Yamano, T.; Ando, A.; Kato, M.; Hirono, M.; Sato, N.; Fukuzawa, H. Algal Dual-Specificity Tyrosine Phosphorylation-Regulated Kinase, Triacylglycerol Accumulation Regulator1, Regulates Accumulation of Triacylglycerol in Nitrogen or Sulfur Deficiency. Plant Physiol. 2015, 168, 752–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaporn, O.; Michael, R.; Arjen, V.D.; Schuurink, R.C.; Van Loon, J.J.A.; Dicke, M. Modulation of Flavonoid Metabolites in Arabidopsis thaliana through Overexpression of the MYB75 Transcription Factor: Role of Kaempferol-3,7-dirhamnoside in Resistance to the Specialist Insect Herbivore Pieris brassicae. J. Exp. Bot. 2014, 65, 2203–2217. [Google Scholar] [CrossRef] [Green Version]
- Ohkubo, Y.; Tanaka, M.; Tabata, R.; Ogawa-Ohnishi, M.; Matsubayashi, Y. Shoot-to-Root Mobile Polypeptides Involved in Systemic Regulation of Nitrogen Acquisition. Nat. Plants 2017, 3, 17029. [Google Scholar] [CrossRef] [PubMed]
- Taleski, M.; Imin, N.; Djordjevic, M.A. CEP Peptide Hormones: Key Players in Orchestrating Nitrogen-Demand Signalling, Root Nodulation, and Lateral Root Development. J. Exp. Bot. 2018, 69, 1829–1836. [Google Scholar] [CrossRef] [Green Version]
- Mohd-Radzman, N.A.; Binos, S.; Truong, T.T.; Imin, N.; Mariani, M.; Djordjevic, M.A. Novel MtCEP1 Peptides Produced in Vivo Differentially Regulate Root Development in Medicago truncatula. J. Exp. Bot. 2015, 17, 5289–5300. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Huang, Z.; Li, X.; Ma, L.; Gu, Q.; Wu, H.; Liu, J.; Borriss, R.; Wu, Z.; Gao, X. Stomatal Closure and SA-, JA/ET- Signaling Pathways are Essential for Bacillus amyloliquefaciens FZB42 to restrict leaf disease caused by Phytophthora nicotianae in Nicotiana benthamiana. Front. Microbiol. 2018, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.J.; Kai, Z.; Liu, L.; Zhang, K.; Li, T. A Role for PacMYBA in ABA-Regulated Anthocyanin Biosynthesis in Red-Colored Sweet Cherry cv. Hong Deng (Prunus avium L.). Plant Cell Physiol. 2014, 55, 862–880. [Google Scholar] [CrossRef]
- Tan, H.; Man, C.; Xie, Y.; Yan, J.; Chu, J.; Huang, J. A Crucial Role of GA-Regulated Flavonol Biosynthesis in Root Growth of Arabidopsis. Mol. Plant. 2019, 12, 521–537. [Google Scholar] [CrossRef] [Green Version]
- Buer, C.S.; Muday, G.K.; Djordjevic, M.A. Flavonoids Are Differentially Taken Up and Transported Long Distances in Arabidopsis. Plant Physiol. 2007, 145, 478–490. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, X.; Zhang, Y.; Chen, X. Antioxidant Activities and Major Anthocyanins of Myrobalan Plum (Prunus cerasifera Ehrh.). J. Food Sci. 2012, 77, C388–C393. [Google Scholar] [CrossRef]
- Misyura, M.; Colasanti, J.; Rothstein, S.J. Physiological and Genetic Analysis of Arabidopsis thaliana Anthocyanin Biosynthesis Mutants under Chronic Adverse Environmental Conditions. J. Exp. Bot. 2012, 64, 229–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eom, S.H.; Baek, S.-A.; Kim, J.K.; Hyun, T.K. Transcriptome Analysis in Chinese Cabbage (Brassica rapa ssp. pekinensis) Provides the Role of Glucosinolate Metabolism in Response to Drought Stress. Molecules 2018, 23, 1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chhajed, S.; Mostafa, I.; He, Y.; Abou-Hashem, M.; El-Domiaty, M.; Chen, S. Glucosinolate Biosynthesis and the Glucosinolate–Myrosinase System in Plant Defense. Agronomy 2020, 10, 1786. [Google Scholar] [CrossRef]
- Fang, C.; Luo, J. Metabolic GWAS-based Dissection of Genetic Bases Underlying the Diversity of Plant Metabolism. Plant J. 2019, 97, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Allan, A.C.; Hellens, R.P.; Laing, W.A. MYB Transcription Factors that Colour Our Fruit. Trends Plant Sci. 2008, 13, 99–102. [Google Scholar] [CrossRef]
- Espley, R.V.; Brendolise, C.; Chagne, D.; Kutty-Amma, S.; Green, S.; Volz, R.; Putterill, J.; Schouten, H.J.; Gardiner, S.E.; Hellens, R.P.; et al. Multiple Repeats of a Promoter Segment Causes Transcription Factor Autoregulation in Red Apples. Plant Cell 2009, 21, 168–183. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Guo, X.; Guo, X.; Zhao, D.; Zhao, W.; Chen, J.; Li, T. PacMYBA, a Sweet Cherry R2R3-MYB Transcription Factor, is a Positive Regulator of Salt Stress Tolerance and Pathogen Resistance. Plant Physiol. Biochem. 2017, 112, 302–311. [Google Scholar] [CrossRef]
- Akhter, D.; Qin, R.; Nath, U.K.; Eshag, J.; Jin, X.; Shi, C. A rice gene, OsPL, Encoding a MYB Family Transcription Factor Confers Anthocyanin Synthesis, Heat Stress Response and Hormonal Signaling. Gene 2019, 699, 62–72. [Google Scholar] [CrossRef]
- Usenik, V.; Fabcic, J.; Stampar, F. Sugars, Organic acids, Phenolic Composition and Antioxidant Activity of Sweet Cherry (Prunus avium L.). Food Chem. 2008, 107, 185–192. [Google Scholar] [CrossRef]
- Yang, X.; Pang, X.; Xu, L.; Fang, R.; Huang, X.; Guan, P.; Lu, W.; Zhang, Z. Accumulation of Soluble Sugars in Peel at High Temperature Leads to Stay-Green Ripe Banana Fruit. J. Exp. Bot. 2009, 60, 4051–4062. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, T.; Xing, C.; Dong, H.; Qi, K.; Gao, J.; Tao, S.; Wu, J.; Wu, J.; Zhang, S.; et al. The β-amylase PbrBAM3 from Pear (Pyrus betulaefolia) Regulates Soluble Sugar Accumulation and ROS Homeostasis in Response to Cold Stress. Plant Sci. 2019, 287, 110184. [Google Scholar] [CrossRef] [PubMed]
- Fichtner, F.; Olas, J.J.; Feil, R.; Watanabe, M.; Krause, U.; Hoefgen, R.; Stitt, M.; Lunn, J. Functional Features of TREHALOSE-6-PHOSPHATE SYNTHASE1—An Essential Enzyme in Arabidopsis thaliana. Plant Cell 2020, 32, 1949–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Lin, T.; Qin, M.; Peng, M.; Yang, C.; et al. Rewiring of the Fruit Metabolome in Tomato Breeding. Cell 2018, 172, 249–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Alseekh, S.; Fernie, A.R.; Luo, J. The Structure and Function of Major Plant Metabolite Modifications. Mol. Plant 2019, 12, 899–919. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wang, D.; Gong, L.; Lyu, Y.; Guo, H.; Chen, W.; Jin, C.; Liu, X.; Fang, C.; Luo, J. Comparative Analysis of Metabolome of Rice Seeds at Three Developmental Stages Using a Recombinant Inbred Line Population. Plant J. 2019, 100, 908–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panida, Y.; Suchada, S.; Muenduen, P. Isolation, Separation and Purification of Rutin from Banana Leaves (Musa balbisiana)—ScienceDirect. Ind. Crops Prod. 2020, 149, 112307. [Google Scholar] [CrossRef]
- Hussain, J.; Bassal, M.; Sarhan, H.; Aga, M. Qualitative and Quantitative Comparison of Rutin, Quercetin and Gallic Acid Concentrations in Syrian Capparis spinosa L. Leaves. J. Pharmacogn. Phytochem. 2017, 6, 407–415. [Google Scholar]
- Diab, H.; Limami, A.M. Reconfiguration of N Metabolism upon Hypoxia Stress and Recovery: Roles of Alanine Aminotransferase (AlaAT) and Glutamate Dehydrogenase (GDH). Plants 2016, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Nostadt, R.; Hilbert, M.; Nizam, S.; Rovenich, H.; Wawra, S.; Martin, J.; Küpper, H.; Mijovilovich, A.; Ursinus, A.; Langen, G.; et al. A Secreted Fungal Histidineand Alanine-Rich Protein Regulates Metal Ion Homeostasis and Oxidative Stress. New Phytol. 2020, 227, 1174–1188. [Google Scholar] [CrossRef] [Green Version]
- Raimbault, A.-K.; Zuily-Fodil, Y.; Soler, A.; de Carvalhoa, M.H.C. A Novel Aspartic Acid Protease Gene from Pineapple Fruit (Ananas comosus): Cloning, Characterization and Relation to Postharvest Chilling Stress Resistance. J. Plant Physiol. 2013, 170, 1536–1540. [Google Scholar] [CrossRef]
- Guo, R.; Zhao, J.; Wang, X.; Guo, C.; Li, Z.; Wang, Y.; Wang, X. Constitutive Expression of a Grape Aspartic Protease Gene in Transgenic Arabidopsis Confers Osmotic Stress Tolerance. Plant Cell Tissue Organ Cult. 2015, 121, 275–287. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Zaheer, A.M.; Shakoor, M.B.; Mahmood, A.; Ishaque, W.; Hussain, A. Foliar Application of Aspartic Acid Lowers Cadmium Uptake. and Cd-Induced Oxidative Stress in Rice under Cd Stress. Environ. Sci. Pollut. Res. 2017, 24, 21938–21947. [Google Scholar] [CrossRef] [PubMed]
- Wingler, A.; Tijero, V.; Müller, M.; Yuan, B.; Munné-Bosch, S. Interactions Between Sucrose and Jasmonate Signalling in the Response to Cold Stress. BMC Plant Biol. 2020, 20, 176. [Google Scholar] [CrossRef] [PubMed]
- Forcat, S.; Bennett, M.; Grant, M.; Mansfield, J. Rapid Linkage of Indole Carboxylic Acid to the Plant Cell Wall Identified as a Component of Basal Defence in Arabidopsis against hrp Mutant Bacteria. Phytochemistry 2010, 71, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; An, J.P.; Li, C.Y.; Shen, X.N.; Liu, Y.J.; Wang, D.R.; Ji, X.L.; Hao, Y.J.; You, C.X. The Apple C2H2-Type Zinc Finger Transcription Factor MdZAT10 Positively Regulates JA-Induced Leaf Senescence by Interacting with MdBT2. Hortic. Res. 2021, 8, 159. [Google Scholar] [CrossRef]
- Jibran, R.; Hunter, D.; Dijkwel, P. Hormonal Regulation of Leaf Senescence through Integration of Developmental and Stress Signals. Plant Mol. Biol. 2013, 82, 547–561. [Google Scholar] [CrossRef]
- Dun, X.; Shi, J.; Liu, H.; Wang, J.; Wang, X.; Wang, H. Genetic Dissection of Root Morphological Traits as Related to Potassium Use Efficiency in Rapeseed under Two Contrasting Potassium Levels by Hydroponics. Sci. China Life Sci. 2019, 62, 746–757. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HN-L vs. LN-L | ||||
| ID | Class | Fold Change | Compounds | Type |
| Bra0152 | Anthocyanins | 3.728 | Delphinidin 3-O-glucoside (Mirtillin) | up |
| * Bra0166 | Anthocyanins | 17.390 | Petunidin 3-O-glucoside | up |
| * Bra0194 | Anthocyanins | 7.650 | Delphinidin | up |
| Bra0232 | Flavone | 29.870 | Selgin 5-O-hexoside | up |
| * Bra0256 | Flavone | 177.398 | Apigenin 7-rutinoside (Isorhoifolin) | up |
| * Bra0282 | Flavone | 4.529 | Chrysoeriol 7-O-hexoside | up |
| Bra0284 | Flavone | 7.519 | Selgin O-malonylhexoside | up |
| Bra0593 | Flavone | 6.265 | Acacetin O-acetyl hexoside | up |
| Bra0636 | Flavone | 3.709 | Velutin O-glucuronic acid | up |
| Bra0641 | Flavone | 47.791 | Chrysoeriol 5-O-hexoside | up |
| Bra0648 | Flavone | 649.644 | Chrysoeriol 7-O-rutinoside | up |
| Bra0692 | Flavone | 3.292 | Butin | up |
| Bra0230 | Flavonol | 41.507 | Quercetin 3-O-rutinoside (Rutin) | up |
| * Bra0245 | Flavonol | 143.514 | Kaempferol 3-O-robinobioside (Biorobin) | up |
| Bra0249 | Flavonol | 38.440 | Quercetin 7-O-rutinoside | up |
| * Bra0255 | Flavonol | 145.810 | Kaempferol 3-O-rutinoside (Nicotiflorin) | up |
| Bra0258 | Flavonol | 172.283 | methylQuercetin O-hexoside | up |
| * Bra0261 | Flavonol | 27.174 | Isorhamnetin 5-O-hexoside | up |
| Bra0286 | Flavonol | 42.274 | Isorhamnetin O-hexoside | up |
| Bra0314 | Flavonol | 13.310 | Kaempferol | up |
| Bra0629 | Flavonol | 51.798 | Quercetin 3-O-glucoside (Isotrifoliin) | up |
| Bra0630 | Flavonol | 428.117 | Quercetin 7-O-β-D-Glucuronide | up |
| * Bra0658 | Flavonol | 58.168 | Quercetin 4′-O-glucoside (Spiraeoside) | up |
| Bra0680 | Flavonol | 35.875 | Quercetin | up |
| Bra0682 | Flavonol | 79.744 | Morin | up |
| * Bra0178 | Anthocyanins | 0.146 | Pelargonidin 3-O-beta-D-glucoside (Callistephin chloride) | down |
| Bra0277 | Flavone | 0.317 | Apigenin 7-O-glucoside (Cosmosiin) | down |
| Bra0320 | Flavonol | 0.141 | Isorhamnetin | down |
| Bra0614 | Flavonol | 0.001 | Dihydromyricetin | down |
| Bra0370 | Flavanone | 0.320 | Xanthohumol | down |
| Bra0611 | Flavanone | 0.315 | Afzelechin (3,5,7,4′-Tetrahydroxyflavan) | down |
| Bra0678 | Flavanone | 0.191 | Liquiritigenin | down |
| Bra0342 | Isoflavone | 0.007 | Biochanin A | down |
| Bra0364 | Isoflavone | 0.037 | Rotenone | down |
| HN-R vs. LN-R | ||||
| ID | Class | Fold Change | Compounds | Type |
| Bra0554 | Anthocyanins | 6.529 | Cyanidin O-syringic acid | up |
| Bra0151 | Anthocyanins | 2.347 | Cyanidin 3,5-O-diglucoside (Cyanin) | up |
| * Bra0265 | Flavone | 1759.926 | Tricin 5-O-rutinoside | up |
| * Bra0245 | Flavonol | 282.185 | Kaempferol 3-O-robinobioside (Biorobin) | up |
| * Bra0255 | Flavonol | 219.556 | Kaempferol 3-O-rutinoside (Nicotiflorin) | up |
| Bra0626 | Flavonol | 6.621 | Fustin | up |
| * Bra0239 | Flavonol | 3.191 | Isorhamnetin 3-O-neohesperidoside | up |
| * Bra0658 | Flavonol | 2.211 | Quercetin 4′-O-glucoside (Spiraeoside) | up |
| Bra0283 | Flavone C-glycosides | 2.323 | Apigenin 8-C-pentoside | up |
| Bra0652 | Flavanone | 25.210 | Hesperetin 7-rutinoside (Hesperidin) | up |
| Bra0663 | Flavanone | 16.040 | Hesperetin 7-O-neohesperidoside (Neohesperidin) | up |
| * Bra0677 | Flavanone | 10.653 | Isosakuranetin-7-neohesperidoside (Poncirin) | up |
| Bra0647 | Flavanone | 3.206 | Hesperetin O-malonylhexoside | up |
| * Bra0194 | Anthocyanins | 0.0105 | Delphinidin | down |
| * Bra0178 | Anthocyanins | 0.213 | Pelargonidin 3-O-beta-D-glucoside (Callistephin chloride) | down |
| * Bra0166 | Anthocyanins | 0.282 | Petunidin 3-O-glucoside | down |
| Bra0301 | Flavone | 0.0005 | Tricetin O-malonylhexoside | down |
| * Bra0282 | Flavone | 0.0117 | Chrysoeriol 7-O-hexoside | down |
| Bra0664 | Flavone | 0.184 | Tricin 7-O-hexoside | down |
| Bra0293 | Flavone | 0.237 | Apigenin O-malonylhexoside | down |
| * Bra0261 | Flavonol | 0.009 | Isorhamnetin 5-O-hexoside | down |
| Bra0182 | Flavone C-glycosides | 0.009 | Eriodictiol 6-C-hexoside 8-C-hexoside-O-hexoside | down |
| Bra0197 | Flavone C-glycosides | 0.009 | Eriodictiol C-hexosyl-O-hexoside | down |
| * Bra0229 | Flavone C-glycosides | 0.255 | Vitexin 2″-O-beta-L-rhamnoside | down |
| Bra0612 | Flavone C-glycosides | 0.329 | Luteolin C-hexoside | down |
| HN-L vs. LN-L | ||||
| ID | Class | Fold Change | Compounds | Type |
| * Bra0436 | Carbohydrates | 25.867 | D-(+)-Sucrose | up |
| Bra0437 | Carbohydrates | 31.017 | L-Gulonic-γ-lactone | up |
| * Bra0443 | Carbohydrates | 8.515 | D-glucoronic acid | up |
| * Bra0445 | Carbohydrates | 5.894 | DL-Arabinose | up |
| Bra0449 | Carbohydrates | 3.364 | Trehalose 6-phosphate | up |
| Bra0453 | Carbohydrates | 3.854 | L-Fucose | up |
| Bra0456 | Carbohydrates | 0.160 | 2-Deoxyribose 1-phosphate | down |
| HN-R vs. LN-R | ||||
| ID | Class | Fold Change | Compounds | Type |
| * Bra0443 | Carbohydrates | 11.008 | D-glucoronic acid | up |
| * Bra0436 | Carbohydrates | 2.702 | D-(+)-Sucrose | up |
| * Bra0445 | Carbohydrates | 2.286 | DL-Arabinose | up |
| Bra0447 | Carbohydrates | 2.212 | D-(+)-Glucono-1,5-lactone | up |
| HN-L vs. LN-L | ||||
| ID | Class | Fold Change | Compounds | Type |
| * Bra0056 | Amino acids | 2.922 | 2-Aminoadipic acid (L-Homoglutamic acid) | up |
| Bra0082 | Amino acids | 3.617 | L-Methionine | up |
| * Bra0421 | Amino acids | 4.185 | L-Threonine | up |
| Bra0435 | Amino acids | 104.407 | L-Cysteine | up |
| * Bra0057 | Amino acids | 41.650 | Aspartic acid | up |
| Bra0222 | Amino acids derivative | 3.361 | N-Phenylacetylglycine | up |
| Bra0440 | Amino acids derivative | 17.621 | L-Saccharopine | up |
| * Bra0451 | Amino acids derivative | 5.508 | Allysine(6-Oxo DL-Norleucine) | up |
| Bra0483 | Amino acids derivative | 3.663 | γ-Glu-Cys | up |
| * Bra0487 | Amino acids derivative | 90.556 | (−)-3-(3,4-Dihydroxyphenyl)-2-methylalanine | up |
| * Bra0515 | Amino acids derivative | 6.176 | 3-Hydroxy-3-methylpentane-1,5-dioic acid | up |
| * Bra0656 | Amino acids derivative | 3.674 | N-(3-Indolylacetyl)-L-alanine | up |
| Bra0009 | Amino acids | 0.018 | L(+)-Ornithine | down |
| * Bra0431 | Amino acids | 0.001 | L-(−)-Cystine | down |
| Bra0101 | Amino acids derivative | 0.196 | N-Acetyl-L-glutamic acid | down |
| Bra0439 | Amino acids derivative | 0.300 | L-Glutamine O-hexside | down |
| * Bra0470 | Amino acids derivative | 0.179 | Glutathione oxidized | down |
| * Bra0510 | Amino acids derivative | 0.146 | L-alanine | down |
| * Bra0553 | Amino acids derivative | 0.263 | Asp-phe | down |
| * Bra0560 | Amino acids derivative | 0.079 | N-Acetylmethionine | down |
| Bra0561 | Amino acids derivative | 0.372 | 2,3-dimethylsuccinic acid | down |
| HN-R vs. LN-R | ||||
| ID | Class | Fold Change | Compounds | Type |
| Bra0421 | Amino acids | 5.571 | L-Threonine | up |
| Bra0043 | Amino acids | 3.255 | L-Homocitrulline | up |
| Bra0426 | Amino acids | 3.105 | L-Citrulline | up |
| * Bra0056 | Amino acids | 2.208 | 2-Aminoadipic acid (L-Homoglutamic acid) | up |
| Bra0503 | Amino acid derivatives | 13.495 | S-(5’-Adenosy)-L-homocysteine | up |
| * Bra0487 | Amino acid derivatives | 8.523 | (−)-3-(3,4-Dihydroxyphenyl)-2-methylalanine | up |
| Bra0050 | Amino acid derivatives | 5.462 | Methionine sulfoxide | up |
| Bra0613 | Amino acid derivatives | 3.527 | Phe-Phe | up |
| * Bra0451 | Amino acid derivatives | 3.329 | Allysine(6-Oxo DL-Norleucine) | up |
| * Bra0510 | Amino acid | 3.095 | L-alanine | up |
| * Bra0656 | Amino acid derivatives | 2.534 | N-(3-Indolylacetyl)-L-alanine | up |
| * Bra0515 | Amino acid derivatives | 2.247 | 3-Hydroxy-3-methylpentane-1,5-dioic acid | up |
| Bra0031 | Amino acids | 0.320 | L-alanine | down |
| * Bra0431 | Amino acids | 0.407 | L-(−)-Cystine | down |
| Bra0011 | Amino acids | 0.469 | L-(+)-Lysine | down |
| Bra0497 | Amino acid derivatives | 0.272 | S-(methyl)glutathione | down |
| * Bra0057 | Amino acid | 0.353 | Aspartic acid | down |
| Bra0638 | Amino acid derivatives | 0.381 | Acetyl tryptophan | down |
| Bra0474 | Amino acid derivatives | 0.418 | N-Acetylaspartate | down |
| * Bra0560 | Amino acid derivatives | 0.429 | N-Acetylmethionine | down |
| Bra0022 | Amino acid derivatives | 0.433 | 2,6-Diaminooimelic acid | down |
| * Bra0470 | Amino acid derivatives | 0.445 | Glutathione oxidized | down |
| Bra0071 | Amino acid derivatives | 0.448 | (5-L-Glutamyl)-L-amino acid | down |
| Bra0072 | Amino acid derivatives | 0.457 | Nα-Acetyl-L-arginine | down |
| * Bra0553 | Amino acid derivatives | 0.483 | Asp-phe | down |
| Bra0045 | Amino acid derivatives | 0.497 | N,N-Dimethylglycine | down |
| HN-L vs. LN-L | ||||
| ID | Class | Fold Change | Compounds | Type |
| * Bra0177 | Phytohormones | 300.296 | Indole carboxylic acid | up |
| Bra0329 | Phytohormones | 5.791 | Methyl indole-3-acetate | up |
| Bra0303 | Phytohormones | 0.188 | Indole 3-acetic acid (IAA) | down |
| Bra0679 | Phytohormones | 0.168 | Gibberellin A20 | down |
| Bra0701 | Phytohormones | 0.153 | N-[(−)-Jasmonoyl]-(L)-Isoleucine (JA-L-Ile) | down |
| HN-R vs. LN-R | ||||
| ID | Class | Fold Change | Compounds | Type |
| * Bra0177 | Phytohormones | 0.057 | Indole carboxylic acid | down |
| Bra0156 | Phytohormones | 0.418 | trans-zeatin 9-O-glucoside | down |
| Bra0529 | Phytohormones | 0.431 | Salicylic acid O-glucoside | down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, X.; Yang, L.; Han, P.; Gu, C.; Li, Y.; Liao, X.; Qin, L. Metabolic Profiles Reveal Changes in the Leaves and Roots of Rapeseed (Brassica napus L.) Seedlings under Nitrogen Deficiency. Int. J. Mol. Sci. 2022, 23, 5784. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105784
Shen X, Yang L, Han P, Gu C, Li Y, Liao X, Qin L. Metabolic Profiles Reveal Changes in the Leaves and Roots of Rapeseed (Brassica napus L.) Seedlings under Nitrogen Deficiency. International Journal of Molecular Sciences. 2022; 23(10):5784. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105784
Chicago/Turabian StyleShen, Xinjie, Ling Yang, Peipei Han, Chiming Gu, Yinshui Li, Xing Liao, and Lu Qin. 2022. "Metabolic Profiles Reveal Changes in the Leaves and Roots of Rapeseed (Brassica napus L.) Seedlings under Nitrogen Deficiency" International Journal of Molecular Sciences 23, no. 10: 5784. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105784