
SOX2 Is a Univocal Marker for Human Oral Mucosa Epithelium Useful in Post-COMET Patient Characterization

 ,
,  ,
,  ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
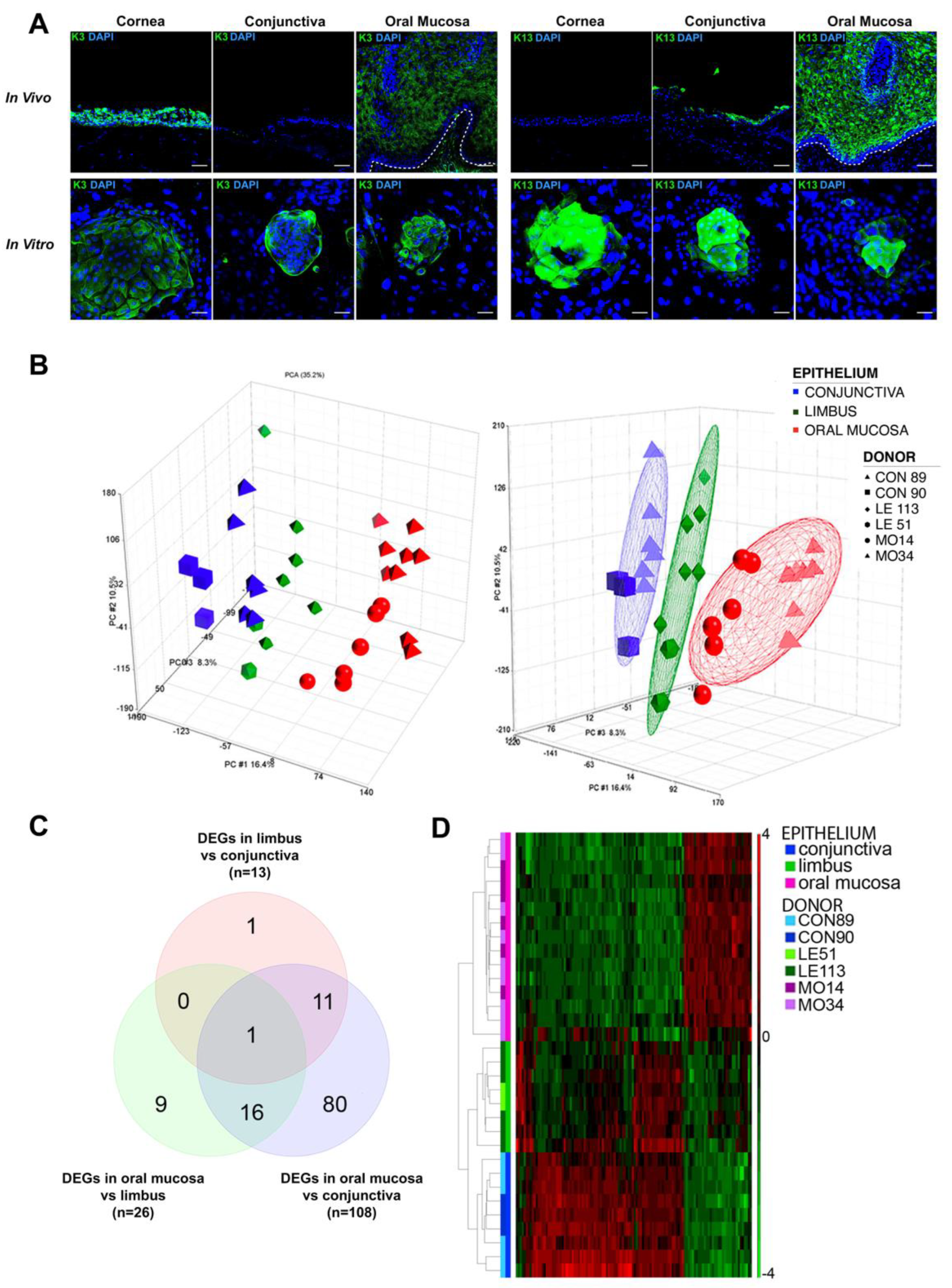
2.1. Keratins Expression in Wound Healing
2.2. Gene Expression Profiling of Oral Mucosa, Conjunctiva and Limbal Holoclones
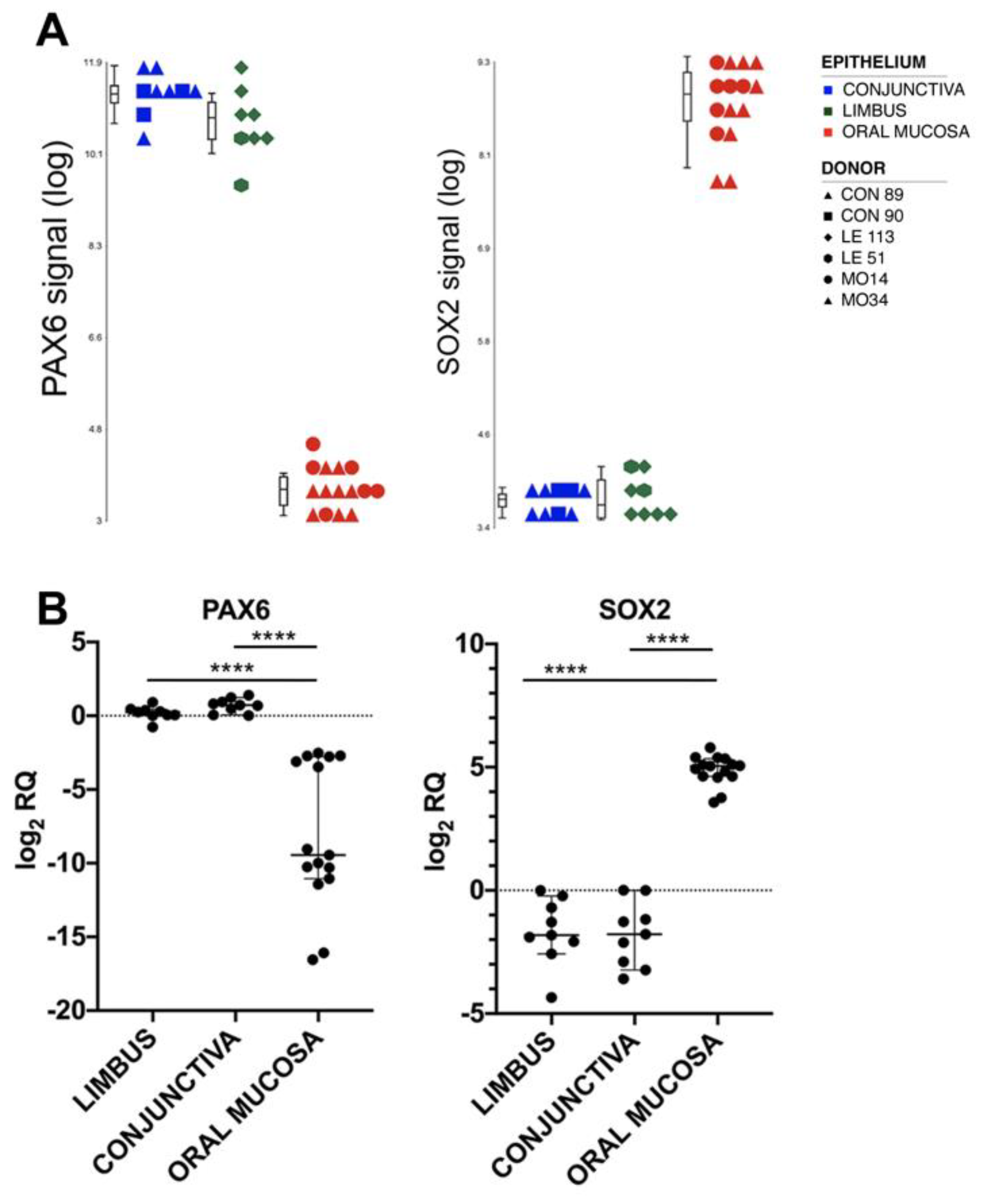
2.3. Markers for Oral Mucosa Compared to Ocular Surface Tissues
2.4. Validation of the Results by Real-Time RT-PCR
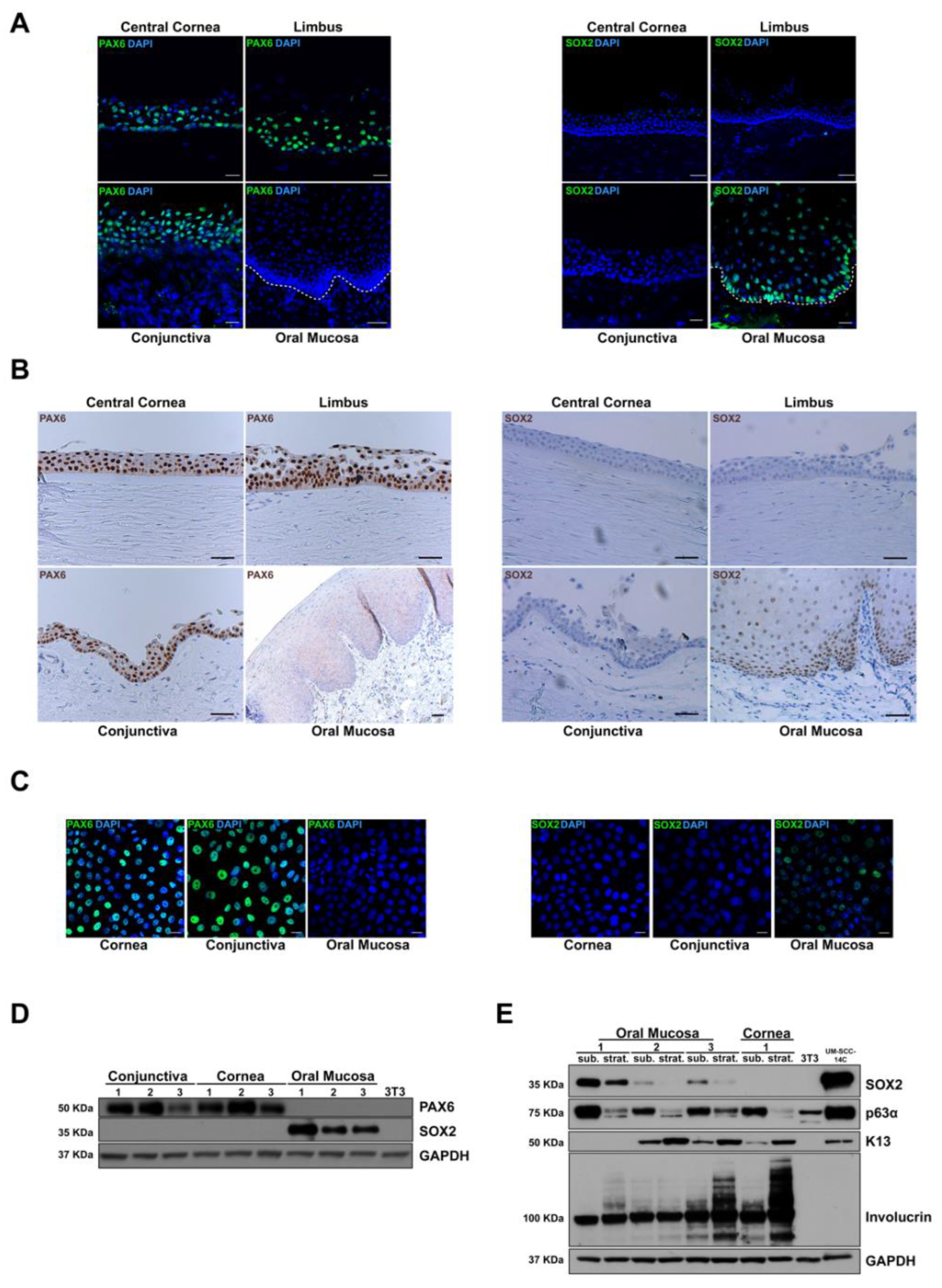
2.5. Validation of the Results by Immunofluorescence, Immunohistochemistry and Western Blot
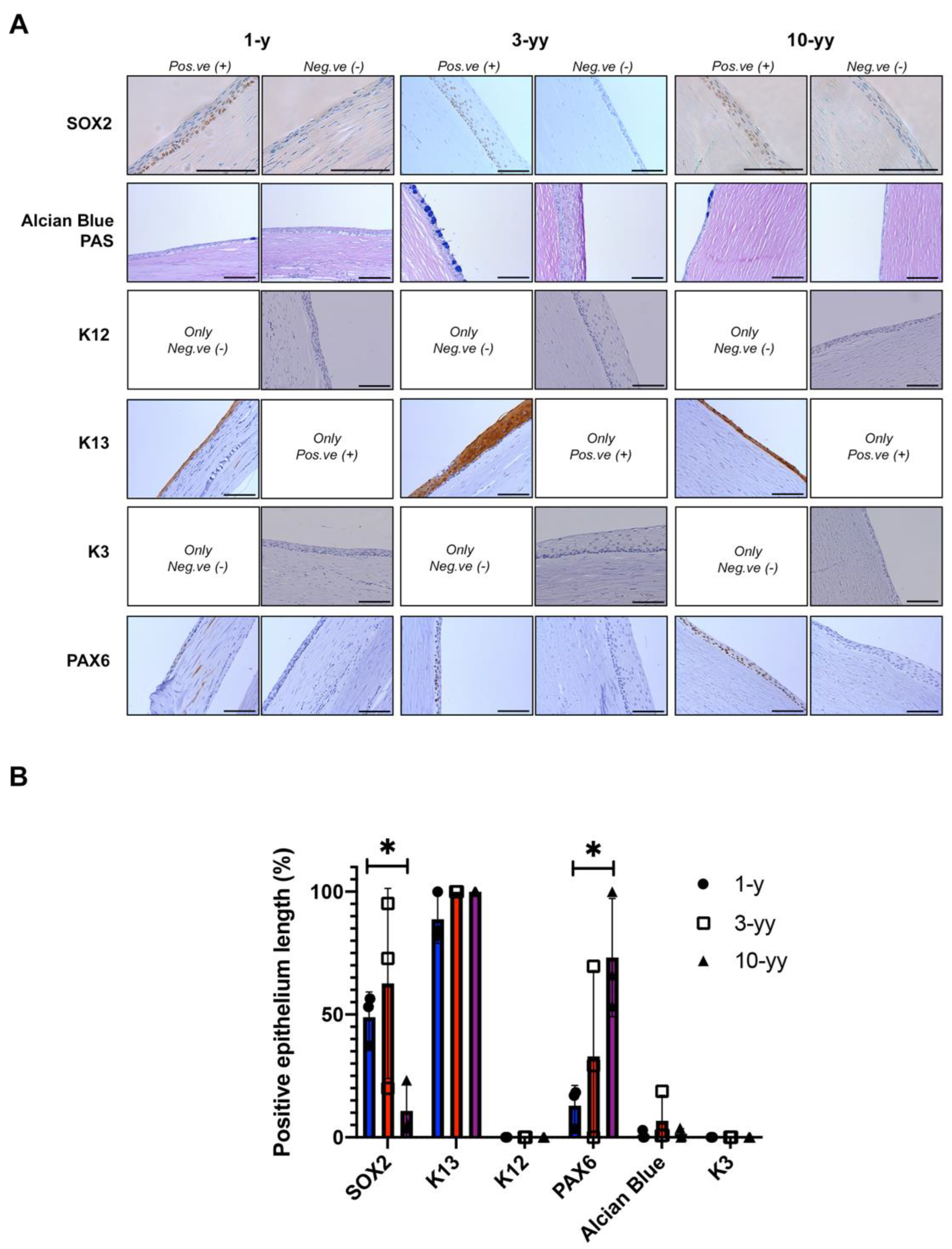
2.6. SOX2 Is Negatively Correlated to Oral Mucosa Stratification
2.7. Phenotypic Characterization of Patient after COMET
3. Discussion
4. Materials and Methods
4.1. Patients and Specimens
4.2. COMET Transplantation
4.3. Cell Cultures, Clonal Analysis and Colony Forming Efficiency
4.4. Microarray Analyses and Real-Time RT-PCR
4.5. Immunofluorescence and Immunohistochemistry
4.6. Western Blot Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chiavelli, C.; Attico, E.; Sceberras, V.; Fantacci, M.; Melonari, M.; Pellegrini, G. Stem Cells and Ocular Regeneration. In Encyclopedia of Tissue Engineering and Regenerative Medicine; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar] [CrossRef]
- Niederkorn, J.Y.; Kaplan, H.J. Rationale for immune response and the eye. Chem. Immunol. Allergy 2007, 92, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Schermer, A.; Galvin, S.; Sun, T.-T. Differentiation-related expression of a major 64K corneal keratin in vivo and in culture suggests limbal location of corneal epithelial stem cells. J. Cell Biol. 1986, 103, 49–62. [Google Scholar] [CrossRef] [PubMed]
- A Shanmuganathan, V.; Foster, T.; Kulkarni, B.B.; Hopkinson, A.; Gray, T.; Powe, D.G.; Lowe, J.; Dua, H.S. Morphological characteristics of the limbal epithelial crypt. Br. J. Ophthalmol. 2007, 91, 514–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, S.X.; Borderie, V.; Chan, C.C.; Dana, R.; Figueiredo, F.C.; Gomes, J.A.P.; Pellegrini, G.; Shimmura, S.; Kruse, F.E. Global Consensus on Definition, Classification, Diagnosis, and Staging of Limbal Stem Cell Deficiency. Cornea 2019, 38, 364–375. [Google Scholar] [CrossRef]
- Haagdorens, M.; Van Acker, S.I.; Van Gerwen, V.; Dhubhghaill, S.N.; Koppen, C.; Tassignon, M.-J.; Zakaria, N. Limbal Stem Cell Deficiency: Current Treatment Options and Emerging Therapies. Stem Cells Int. 2016, 2016, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Banayan, N.; Georgeon, C.; Grieve, K.; Ghoubay, D.; Baudouin, F.; Borderie, V. In vivo confocal microscopy and optical coherence tomography as innovative tools for the diagnosis of limbal stem cell deficiency. J. Fr. Ophtalmol. 2018, 41, e395–e406. [Google Scholar] [CrossRef]
- Orphanet: Limbal Stem Cell Deficiency. Available online: https://www.orpha.net/consor/cgi-bin/OC_Exp.php?lng=EN&Expert=171673 (accessed on 28 April 2022).
- Pellegrini, G.; Traverso, C.E.; Franzi, A.T.; Zingirian, M.; Cancedda, R.; De Luca, M. Long-term restoration of damaged corneal surfaces with autologous cultivated corneal epithelium. Lancet 1997, 349, 990–993. [Google Scholar] [CrossRef]
- Holoclar|European Medicines Agency. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/holoclar (accessed on 28 April 2022).
- Holan, V.; Javorkova, E. Mesenchymal Stem Cells, Nanofiber Scaffolds and Ocular Surface Reconstruction. Stem Cell Rev. Rep. 2013, 9, 609–619. [Google Scholar] [CrossRef]
- Nakamura, T.; Kinoshita, S. Ocular Surface Reconstruction Using Cultivated Mucosal Epithelial Stem Cells. Cornea 2003, 22, S75–S80. [Google Scholar] [CrossRef]
- Nakamura, T.; Takeda, K.; Inatomi, T.; Sotozono, C.; Kinoshita, S. Long-term results of autologous cultivated oral mucosal epithelial transplantation in the scar phase of severe ocular surface disorders. Br. J. Ophthalmol. 2010, 95, 942–946. [Google Scholar] [CrossRef] [Green Version]
- Nishida, K.; Yamato, M.; Hayashida, Y.; Watanabe, K.; Yamamoto, K.; Adachi, E.; Nagai, S.; Kikuchi, A.; Maeda, N.; Watanabe, H.; et al. Corneal Reconstruction with Tissue-Engineered Cell Sheets Composed of Autologous Oral Mucosal Epithelium. N. Engl. J. Med. 2004, 351, 1187–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burillon, C.; Huot, L.; Justin, V.; Nataf, S.; Chapuis, F.; Decullier, E.; Damour, O. Cultured Autologous Oral Mucosal Epithelial Cell Sheet (CAOMECS) Transplantation for the Treatment of Corneal Limbal Epithelial Stem Cell Deficiency. Investig. Opthalmol. Vis. Sci. 2012, 53, 1325–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, H.; Yamato, M.; Nishida, K.; Okano, T. Evidence of the Survival of Ectopically Transplanted Oral Mucosal Epithelial Stem Cells After Repeated Wounding of Cornea. Mol. Ther. 2014, 22, 1544–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, G. Changing the Cell Source in Cell Therapy? New Engl. J. Med. 2004, 351, 1170–1172. [Google Scholar] [CrossRef]
- Attico, E.; Galaverni, G.; Pellegrini, G. Clinical Studies of COMET for Total LSCD: A Review of the Methods and Molecular Markers for Follow-Up Characterizations. Curr. Ophthalmol. Rep. 2021, 9, 25–37. [Google Scholar] [CrossRef]
- Williams, K.A.; Brereton, H.M.; Aggarwal, R.; Sykes, P.J.; Turner, D.R.; Russ, G.R.; Coster, D.J. Use of DNA Polymorphisms and the Polymerase Chain Reaction to Examine the Survival of a Human Limbal Stem Cell Allograft. Am. J. Ophthalmol. 1995, 120, 342–350. [Google Scholar] [CrossRef]
- Henderson, T.R.M.; Coster, D.J.; A Williams, K. The long term outcome of limbal allografts: The search for surviving cells. Br. J. Ophthalmol. 2001, 85, 604–609. [Google Scholar] [CrossRef] [Green Version]
- Moll, R.; Divo, M.; Langbein, L. The human keratins: Biology and pathology. Histochem. Cell Biol. 2008, 129, 705–733. [Google Scholar] [CrossRef] [Green Version]
- Merjava, S.; Neuwirth, A.; Tanzerova, M.; Jirsova, K. The spectrum of cytokeratins expressed in the adult human cornea, limbus and perilimbal conjunctiva. Histol. Histopathol. 2011, 26, 323–331. [Google Scholar] [CrossRef]
- Chen, H.-C.J.; Chen, H.-L.; Lai, J.-Y.; Chen, C.-C.; Tsai, Y.-J.; Kuo, M.-T.; Chu, P.-H.; Sun, C.-C.; Chen, J.-K.; Ma, D.H.-K. Persistence of Transplanted Oral Mucosal Epithelial Cells in Human Cornea. Investig. Opthalmology Vis. Sci. 2009, 50, 4660–4668. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Inatomi, T.; Cooper, L.J.; Rigby, H.; Fullwood, N.J.; Kinoshita, S. Phenotypic Investigation of Human Eyes with Transplanted Autologous Cultivated Oral Mucosal Epithelial Sheets for Severe Ocular Surface Diseases. Ophthalmology 2007, 114, 1080–1088. [Google Scholar] [CrossRef] [PubMed]
- Vemuganti, G.; Gaddipati, S.; Muralidhar, R.; Sangwan, V.; Mariappan, I.; Balasubramanian, D. Oral epithelial cells transplanted on to corneal surface tend to adapt to the ocular phenotype. Indian J. Ophthalmol. 2014, 62, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yin, M.; Zhang, L.-J. Keratin 6, 16 and 17—Critical Barrier Alarmin Molecules in Skin Wounds and Psoriasis. Cells 2019, 8, 807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzalupo, S.; Wong, P.; Martin, P.; Coulombe, P.A. Role for keratins 6 and 17 during wound closure in embryonic mouse skin. Dev. Dyn. 2003, 226, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Freedberg, I.M.; Tomic-Canic, M.; Komine, M.; Blumenberg, M. Keratins and the Keratinocyte Activation Cycle. J. Investig. Dermatol. 2001, 116, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.; Piatigorsky, J. Overexpression of Pax6 in Mouse Cornea Directly Alters Corneal Epithelial Cells: Changes in Immune Function, Vascularization, and Differentiation. Investig. Opthalmology Vis. Sci. 2011, 52, 4158–4168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sceberras, V.; Attico, E.; Bianchi, E.; Galaverni, G.; Melonari, M.; Corradini, F.; Fantacci, M.; Ribbene, A.; Losi, L.; Balò, S.; et al. Preclinical study for treatment of hypospadias by advanced therapy medicinal products. World J. Urol. 2019, 38, 2115–2122. [Google Scholar] [CrossRef]
- Barrandon, Y.; Green, H. Three clonal types of keratinocyte with different capacities for multiplication. Proc. Natl. Acad. Sci. USA 1987, 84, 2302–2306. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Li, J. Corneal neovascularization suppressed by TIMP2 released from human amniotic membranes. Yan Ke Xue Bao 2005, 21, 56–61. [Google Scholar]
- Iglesias-Bartolome, R.; Uchiyama, A.; Molinolo, A.A.; Abusleme, L.; Brooks, S.R.; Callejas-Valera, J.L.; Edwards, D.; Doci, C.; Asselin-Labat, M.-L.; Onaitis, M.W.; et al. Transcriptional signature primes human oral mucosa for rapid wound healing. Sci. Transl. Med. 2018, 10, 855–862. [Google Scholar] [CrossRef] [Green Version]
- Brysk, M.M.; Arany, I.; Brysk, H.; Chen, S.-H.; Calhoun, K.H.; Tyring, S.K. Gene expression of markers associated with proliferation and differentiation in human keratinocytes cultured from epidermis and from buccal mucosa. Arch. Oral Biol. 1995, 40, 855–862. [Google Scholar] [CrossRef]
- Henderson, T.R.M.; McCall, S.H.; Taylor, G.R.; A Noble, B. Do transplanted corneal limbal stem cells survive in vivo long term? Possible techniques to detect donor cell survival by polymerase chain reaction with the amelogenin gene and Y-specific probes. Eye 1997, 11, 779–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utheim, T.P.; Utheim, A.; Khan, Q.-E.; Sehic, A. Culture of Oral Mucosal Epithelial Cells for the Purpose of Treating Limbal Stem Cell Deficiency. J. Funct. Biomater. 2016, 7, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazirani, J.; Nair, D.; Shanbhag, S.; Wurity, S.; Ranjan, A.; Sangwan, V. Limbal Stem Cell Deficiency—Demography and Underlying Causes. Am. J. Ophthalmol. 2018, 188, 99–103. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, L.; Carulli, S.; Cocchiarella, F.; Quaglino, D.; Enzo, E.; Franchini, E.; Giannetti, A.; DE Santis, G.; Recchia, A.; Pellegrini, G.; et al. Long-Term Stability and Safety of Transgenic Cultured Epidermal Stem Cells in Gene Therapy of Junctional Epidermolysis Bullosa. Stem Cell Rep. 2014, 2, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; He, J.; Wei, J.; Cho, W.C.; Liu, X. Diversity of Epithelial Stem Cell Types in Adult Lung. Stem Cells Int. 2015, 2015, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Bianco, P.; Cao, X.; Frenette, P.S.; Mao, J.J.; Robey, P.G.; Simmons, P.J.; Wang, C.-Y. The meaning, the sense and the significance: Translating the science of mesenchymal stem cells into medicine. Nat. Med. 2013, 19, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Presland, R.B.; Dale, B.A. Epithelial Structural Proteins of the Skin and Oral Cavity: Function in Health and Disease. Crit. Rev. Oral Biol. Med. 2000, 11, 383–408. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Serror, L.; Nir, E.; Dhiraj, D.; Altshuler, A.; Khreish, M.; Tiosano, B.; Hasson, P.; Panman, L.; Luxenburg, C.; et al. SOX2 Regulates P63 and Stem/Progenitor Cell State in the Corneal Epithelium. Stem Cells 2018, 37, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Pedrotti, E.; Passilongo, M.; Fasolo, A.; Nubile, M.; Parisi, G.; Mastropasqua, R.; Ficial, S.; Bertolin, M.; Di Iorio, E.; Ponzin, D.; et al. In Vivo Confocal Microscopy 1 Year after Autologous Cultured Limbal Stem Cell Grafts. Ophthalmology 2015, 122, 1660–1668. [Google Scholar] [CrossRef] [PubMed]
- Mastropasqua, R.; Agnifili, L.; Fasanella, V.; Lappa, A.; Brescia, L.; Lanzini, M.; Oddone, F.; Perri, P.; Mastropasqua, L. In Vivo Distribution of Corneal Epithelial Dendritic Cells in Patients With Glaucoma. Investig. Opthalmology Vis. Sci. 2016, 57, 5996–6002. [Google Scholar] [CrossRef]
- Latta, L.; Viestenz, A.; Stachon, T.; Colanesi, S.; Szentmáry, N.; Seitz, B.; Käsmann-Kellner, B. Human aniridia limbal epithelial cells lack expression of keratins K3 and K12. Exp. Eye Res. 2018, 167, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Auw-Haedrich, C.; Agrawal, M.; Gabbert, H.E.; Meyer, P.; Arnold, N.; Reinhard, T. Immunohistochemical expression of epithelial cell markers in corneas with congenital aniridia and ocular cicatrizing pemphigoid. Acta Ophthalmol. 2011, 89, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Tanioka, H.; Kawasaki, S.; Yamasaki, K.; Ang, L.P.K.; Koizumi, N.; Nakamura, T.; Yokoi, N.; Komuro, A.; Inatomi, T.; Kinoshita, S. Establishment of a Cultivated Human Conjunctival Epithelium as an Alternative Tissue Source for Autologous Corneal Epithelial Transplantation. Investig. Opthalmology Vis. Sci. 2006, 47, 3820–3827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contaldo, M.; Agozzino, M.; Moscarella, E.; Esposito, S.; Serpico, R.; Ardigò, M. In VivoCharacterization of Healthy Oral Mucosa by Reflectance Confocal Microscopy: A Translational Research for Optical Biopsy. Ultrastruct. Pathol. 2013, 37, 151–158. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Attico, E.; Galaverni, G.; Bianchi, E.; Losi, L.; Manfredini, R.; Lambiase, A.; Rama, P.; Pellegrini, G. SOX2 Is a Univocal Marker for Human Oral Mucosa Epithelium Useful in Post-COMET Patient Characterization. Int. J. Mol. Sci. 2022, 23, 5785. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105785
Attico E, Galaverni G, Bianchi E, Losi L, Manfredini R, Lambiase A, Rama P, Pellegrini G. SOX2 Is a Univocal Marker for Human Oral Mucosa Epithelium Useful in Post-COMET Patient Characterization. International Journal of Molecular Sciences. 2022; 23(10):5785. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105785
Chicago/Turabian StyleAttico, Eustachio, Giulia Galaverni, Elisa Bianchi, Lorena Losi, Rossella Manfredini, Alessandro Lambiase, Paolo Rama, and Graziella Pellegrini. 2022. "SOX2 Is a Univocal Marker for Human Oral Mucosa Epithelium Useful in Post-COMET Patient Characterization" International Journal of Molecular Sciences 23, no. 10: 5785. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105785