
Effects of Methoxyfenozide-Loaded Fluorescent Mesoporous Silica Nanoparticles on Plutella xylostella (L.) (Lepidoptera: Plutellidae) Mortality and Detoxification Enzyme Levels Activities



Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Preparation of Fluorescent Silica Nanoparticles (FL-SiO2 NPs)
2.3. Preparation of Insecticide-Loaded Nanoparticles HCl
2.4. Sample Characterization
2.5. Methoxyfenozide Loading Content
2.6. In Vitro Release Experiment
2.7. Insecticidal Activity of Me@FL-SiO2-70 nm and -150 nm NPs against P. xylostella
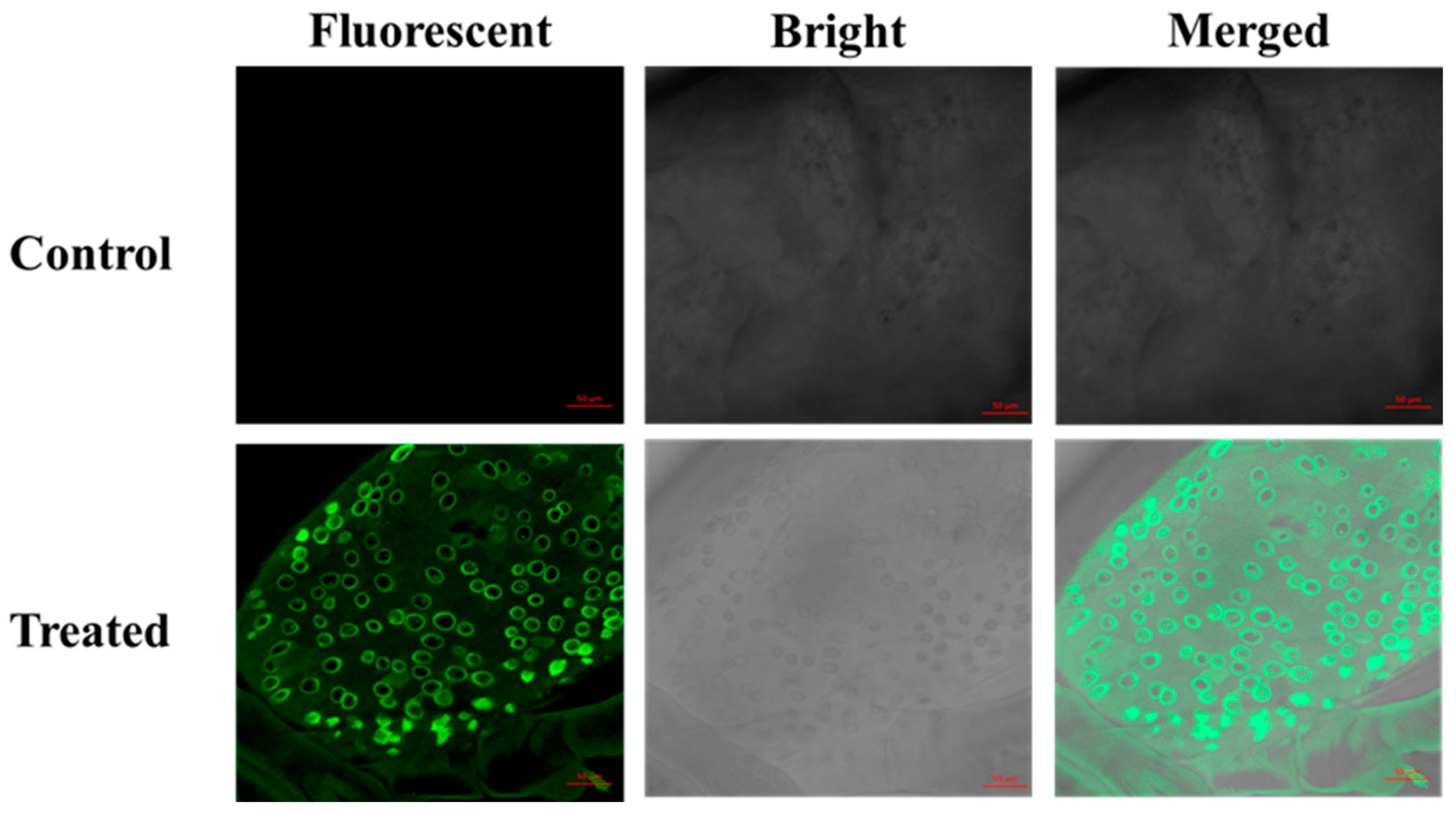
2.8. Confocal Study of FL-SiO2 NPs in the Midgut of P. xylostella
2.9. Detoxification Enzymes
2.9.1. Preparation of P. xylostella Samples
2.9.2. Activity of Glutathione-S-Transferase (GST)
2.9.3. Activity of Carboxylesterase (CarE)
2.9.4. P450 Activity
2.9.5. AChE Activity
2.9.6. Statistical Analysis
3. Results
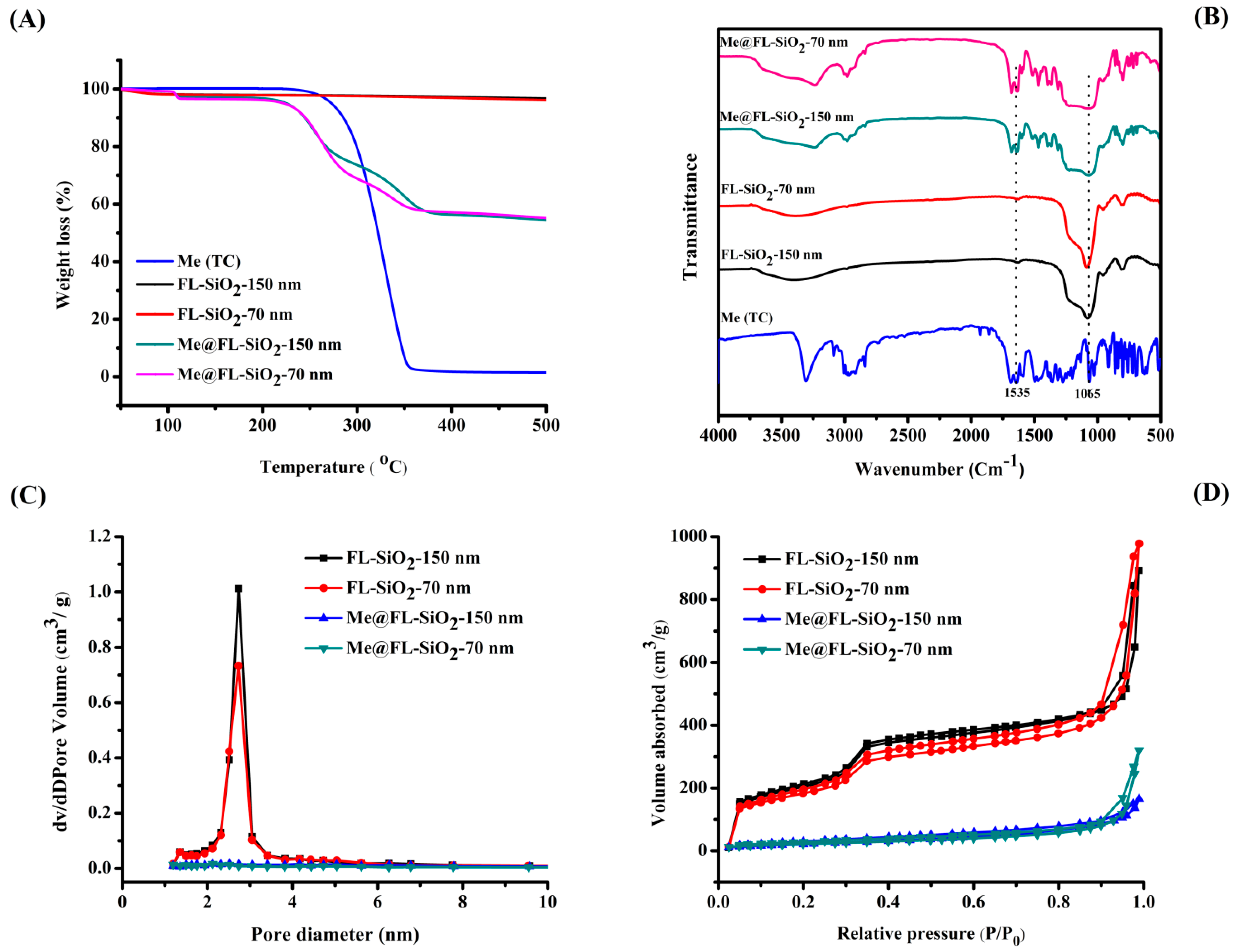
3.1. Morphological and Structural Characterization
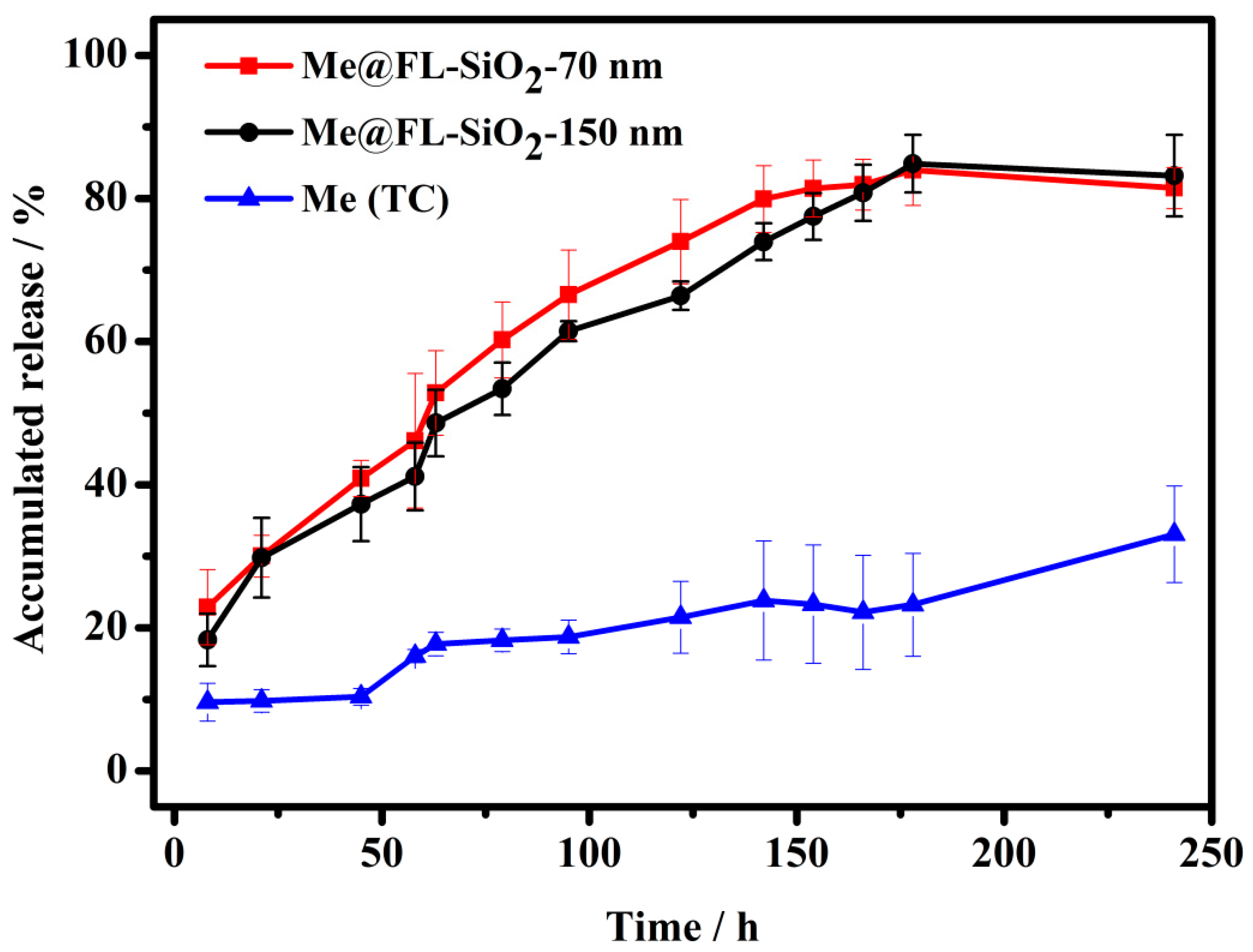
3.2. Controlled Release of Methoxyfenozide
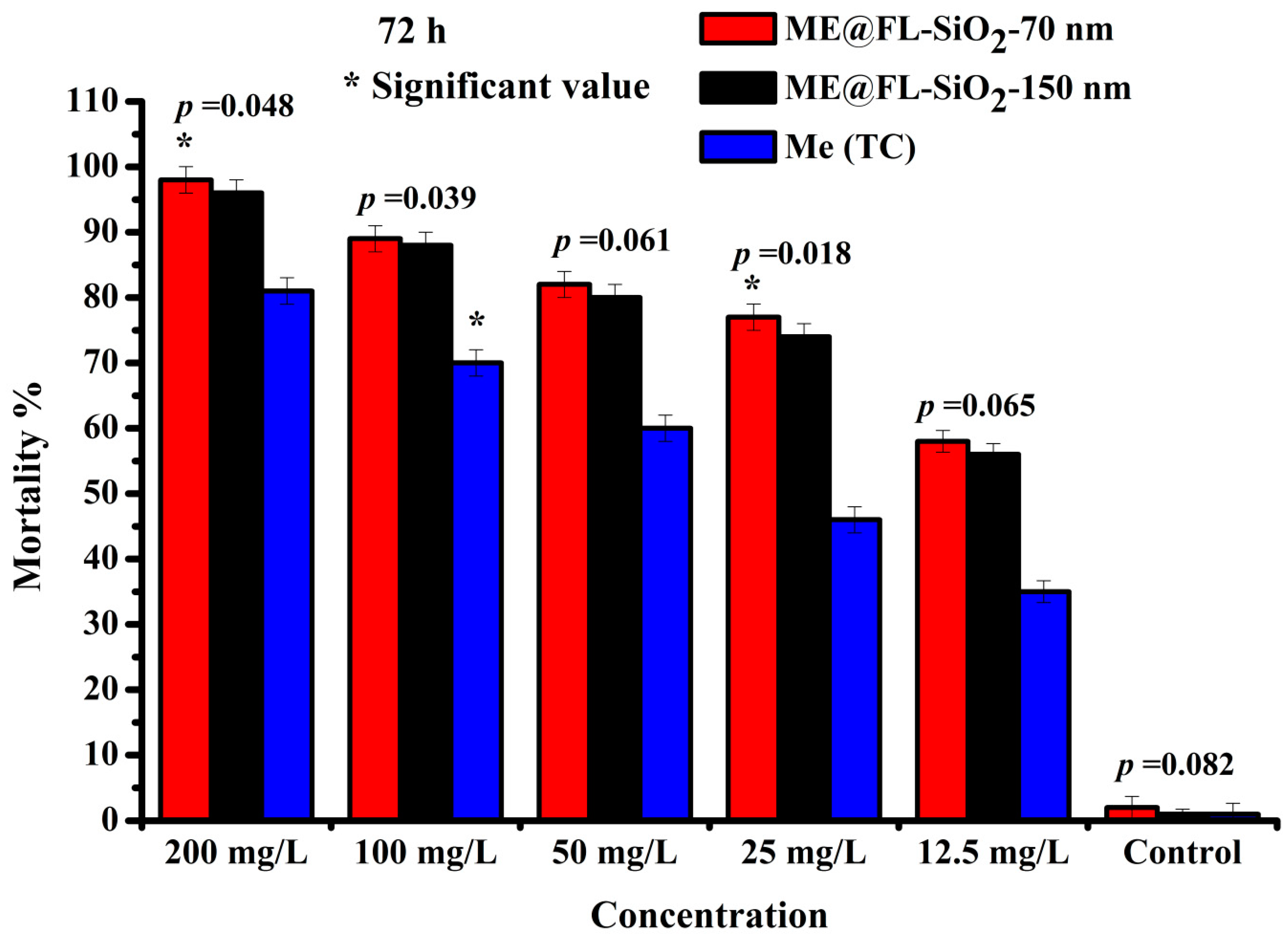
3.3. Bioactivity Study of Me@FL-SiO2 NPs
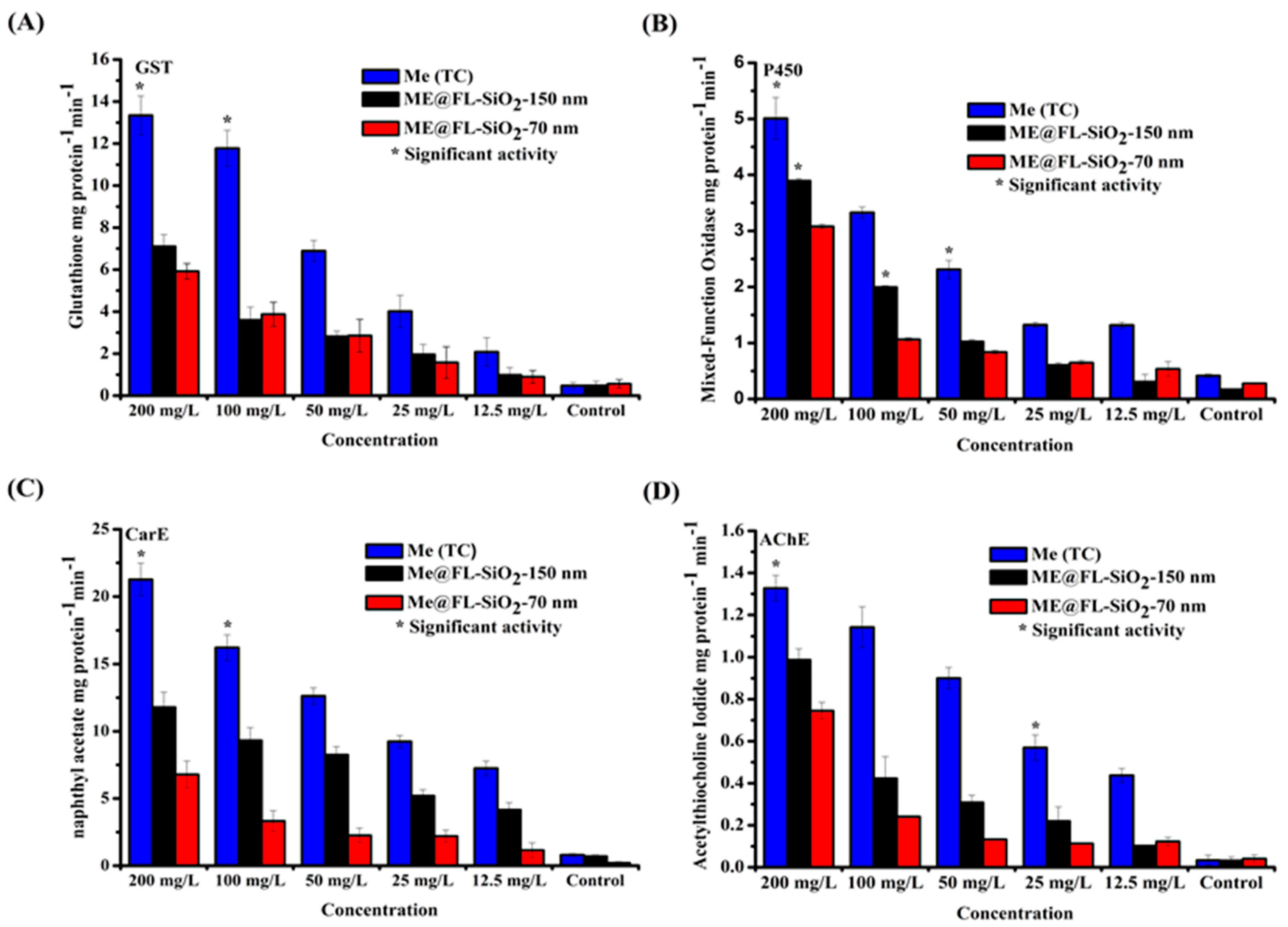
3.4. Effect of Me@FL-SiO2 NPs (70 nm and 150 nm) and Methoxyfenozide TC on Detoxification Enzyme Activity
3.5. Confocal Microscopy of Fluorescent Silica Nanoparticles in the P. xylostella
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shan, J.; Sun, X.; Li, R.; Zhu, B.; Liang, P.; Gao, X. Identification of ABCG transporter genes associated with chlorantraniliprole resistance in Plutella xylostella (L.). Pest Manag. Sci. 2021, 77, 3491–3499. [Google Scholar] [CrossRef] [PubMed]
- Abram, P.K.; Thiessen, J.; Clarke, P.; Gillespie, D.R.; Fernández-Triana, J.L.; Bennett, A.M.; Gibson, G.A.; Huber, J.T.; Mason, P.G.; Landry, J.-F. Natural History of Plutella armoraciae Busck, 1912, A Sympatric Congener of the Diamondback Moth, Plutella xylostella (L., 1758), in Southwestern Canada. J. Lepid. Soc. 2022, 76, 25–39. [Google Scholar] [CrossRef]
- Chen, M.Z.; Cao, L.J.; Li, B.Y.; Chen, J.C.; Gong, Y.J.; Yang, Q.; Schmidt, T.L.; Yue, L.; Zhu, J.Y.; Li, H. Migration trajectories of the diamondback moth Plutella xylostella in China inferred from population genomic variation. Pest Manag. Sci. 2020, 77, 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lv, W.; Yuan, Y.; Zhang, T.; Teng, H.; Losey, J.E.; Chang, X. Effects of insecticides on malacostraca when managing diamondback moth (Plutella xylostella) in combination planting-rearing fields. Ecotoxicol. Environ. Saf. 2021, 229, 113090. [Google Scholar] [CrossRef]
- Liang, J.; Yu, M.; Guo, L.; Cui, B.; Zhao, X.; Sun, C.; Wang, Y.; Liu, G.; Cui, H.; Zeng, Z. Bioinspired Development of P(St–MAA)–Avermectin nanoparticles with high affinity for foliage to enhance folia retention. J. Agric. Food Chem. 2017, 66, 6578–6584. [Google Scholar] [CrossRef]
- Eyer, P. The role of oximes in the management of organophosphorus pesticide poisoning. Toxicol. Rev. 2003, 22, 165–190. [Google Scholar] [CrossRef]
- Bradberry, S.M.; Proudfoot, A.T.; Vale, J.A. Poisoning due to chlorophenoxy herbicides. Toxicol. Rev. 2004, 23, 65–73. [Google Scholar] [CrossRef]
- Eddleston, M.; Eyer, P.; Worek, F.; Mohamed, F.; Senarathna, L.; von Meyer, L.; Juszczak, E.; Hittarage, A.; Azhar, S.; Dissanayake, W.; et al. Differences between organophosphorus insecticides in human self-poisoning: A prospective cohort study. Lancet 2005, 366, 1452–1459. [Google Scholar] [CrossRef]
- Eddleston, M.; Worek, F.; Eyer, P.; Thiermann, H.; Von Meyer, L.; Jeganathan, K.; Sheriff, M.H.R.; Dawson, A.; Buckley, N. Poisoning with the S-Alkyl organophosphorus insecticides profenofos and prothiofos. QJM Int. J. Med. 2009, 102, 785–792. [Google Scholar] [CrossRef]
- Fenner, K.; Canonica, S.; Wackett, L.P.; Elsner, M. Evaluating pesticide degradation in the environment: Blind spots and emerging opportunities. Science 2013, 341, 752–758. [Google Scholar] [CrossRef] [Green Version]
- Köhler, H.-R.; Triebskorn, R. Wildlife ecotoxicology of pesticides: Can we track effects to the population level and beyond? Science 2013, 341, 759–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malaj, E.; Peter, C.; Grote, M.; Kühne, R.; Mondy, C.P.; Usseglio-Polatera, P.; Brack, W.; Schäfer, R.B. Organic chemicals jeopardize the health of freshwater ecosystems on the continental scale. Proc. Natl. Acad. Sci. USA 2014, 111, 9549–9554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuruzzaman, M.; Rahman, M.M.; Liu, Y.; Naidu, R. Nanoencapsulation, nano-guard for pesticides: A new window for safe application. J. Agric. Food Chem. 2016, 64, 1447–1483. [Google Scholar] [CrossRef]
- Meshram, A.T.; Vanalkar, A.V.; Kalambe, K.B.; Badar, A.M. Pesticide spraying robot for precision agriculture: A categorical literature review and future trends. J. Field Robot. 2021, 39, 153–171. [Google Scholar] [CrossRef]
- Wang, B.; Song, J.; Zeng, A.; Liu, Y.; Zhang, J.; He, X. Effects of formulations and surfactants on the behavior of pesticide liquid spreading in the plant leaves. Chin. J. Pestic. Sci. 2012, 14, 334–340. [Google Scholar]
- Sharma, B.; Lakra, U.; Sharma, R.; Sharma, S.R. A comprehensive review on nanopesticides and nanofertilizers—A boon for agriculture. Nano-Enabled Agrochem. Agric. 2022, 1, 273–290. [Google Scholar]
- Pandey, A.; Srivastava, S.; Aggarwal, N.; Srivastava, C.; Adholeya, A.; Kochar, M. Assessment of the pesticidal behaviour of diacyl hydrazine-based ready-to-use nanoformulations. Chem. Biol. Technol. Agric. 2020, 7, 1–15. [Google Scholar] [CrossRef]
- Prasad, R.; Bhattacharyya, A.; Nguyen, Q.D. Nanotechnology in sustainable agriculture: Recent developments, challenges, and perspectives. Front. Microbiol. 2017, 8, 1014. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, N.; Ranjan, S.; Ramalingam, C. Applications of nanotechnology in agriculture and water quality management. Environ. Chem. Lett. 2017, 15, 591–605. [Google Scholar] [CrossRef]
- Khot, L.R.; Sankaran, S.; Maja, J.M.; Ehsani, R.; Schuster, E.W. Applications of nanomaterials in agricultural production and crop protection: A review. Crop Prot. 2012, 35, 64–70. [Google Scholar] [CrossRef]
- Jameel, M.; Shoeb, M.; Khan, M.T.; Ullah, R.; Mobin, M.; Farooqi, M.K.; Adnan, S.M. Enhanced insecticidal activity of thiamethoxam by zinc oxide nanoparticles: A novel nanotechnology approach for pest control. ACS Omega 2020, 5, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Nehra, M.; Dilbaghi, N.; Marrazza, G.; Kaushik, A.; Sonne, C.; Kim, K.-H.; Kumar, S. Emerging nanobiotechnology in agriculture for the management of pesticide residues. J. Hazard. Mater. 2020, 401, 123369. [Google Scholar] [CrossRef] [PubMed]
- Rohela, G.K.; Srinivasulu, Y.; Rathore, M.S. A review paper on recent trends in bio-nanotechnology: Implications and potentials. Nanoscience 2019, 9, 12–20. [Google Scholar] [CrossRef]
- Maroofpour, N.; Mousavi, M.; Hejazi, M.J.; Iranipour, S.; Hamishehkar, H.; Desneux, N.; Biondi, A.; Haddi, K. Comparative selectivity of nano and commercial formulations of pirimicarb on a target pest, Brevicoryne brassicae, and its predator Chrysoperla carnea. Ecotoxicology 2021, 30, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Johnston, H.J.; Hutchison, G.R.; Christensen, F.M.; Peters, S.; Hankin, S.; Stone, V. Identification of the mechanisms that drive the toxicity of TiO2 particulates: The contribution of physicochemical characteristics. Part. Fibre Toxicol. 2009, 6, 1–27. [Google Scholar] [CrossRef]
- Stadler, T.; Buteler, M.; Weaver, D.K. Novel use of nanostructured alumina as an insecticide. Pest Manag. Sci. 2010, 66, 577–579. [Google Scholar] [CrossRef]
- Langa, T.P.; Dantas, K.C.T.; Pereira, D.L.; de Oliveira, M.; Ribeiro, L.M.S.; Siqueira, H.A.A. Basis and monitoring of methoxyfenozide resistance in the South American tomato pinworm Tuta absoluta. J. Pest Sci. 2021, 95, 351–364. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, F.; Yu, G.; Shi, J.; Li, C.; Dai, A.; Liu, Z.; Xu, J.; Wang, F.; Wu, J. Synthesis and insecticidal activity of diacyl hydrazine derivatives containing a 3-bromo-1-(3 chloropyridin-2-yl)-1H-pyrazole scafold. Chem. Cent. J. 2017, 11, 50. [Google Scholar] [CrossRef] [Green Version]
- Fahrbach, S.E.; Velarde, R.A. Nervous System Actions of Insect Developmental Hormones in Adult Insects. Hormones 2009, 2, 943–965. [Google Scholar]
- Smagghe, G.; Gomez, L.E.; Dhadialla, T.S. Bisacylhydrazine insecticides for selective pest control. Adv. Insect Physiol. 2012, 43, 163–249. [Google Scholar]
- Yu, L.; Lin, H.; Lu, X.; Chen, Y. Multifunctional Mesoporous Silica Nanoprobes: Material Chemistry—Based Fabrication and Bio-Imaging Functionality. Adv. Ther. 2018, 1, 1800078. [Google Scholar] [CrossRef]
- Bilal, M.; Xu, C.; Cao, L.; Zhao, P.; Cao, C.; Li, F.; Huang, Q. Indoxacarb-Loaded fluorescent mesoporous silica nanoparticles for effective control of Plutella xylostella L. with decreased detoxification enzymes activities. Pest Manag. Sci. 2020, 76, 3749–3758. [Google Scholar] [CrossRef]
- Chen, H.; Zhen, Z.; Tang, W.; Todd, T.; Chuang, Y.-J.; Wang, L.; Pan, Z.; Xie, J. Label-free luminescent mesoporous silica nanoparticles for imaging and drug delivery. Theranostics 2013, 3, 650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walton, K.S.; Snurr, R.Q. Applicability of the BET method for determining surface areas of microporous Metal—Organic frameworks. J. Am. Chem. Soc. 2007, 129, 8552–8556. [Google Scholar] [CrossRef] [PubMed]
- Choma, J.; Jaroniec, M.; Burakiewicz-Mortka, W.; Kloske, M. Critical appraisal of classical methods for determination of mesopore size distributions of MCM-41 materials. App. Surf. Sci. 2002, 196, 216–223. [Google Scholar] [CrossRef]
- Gomes, F.M.; Carvalho, D.B.; Machado, E.A.; Miranda, K. Ultrastructural and functional analysis of secretory goblet cells in the midgut of the lepidopteran Anticarsia gemmatalis. Cell Tissue Res. 2013, 352, 313–326. [Google Scholar] [CrossRef]
- Ye, X.; Liu, M.; Zhao, N.; Xiao, C.; Xu, H.; Jia, J. Targeted delivery of emamectin benzoate by functionalized polysuccinimide nanoparticles for the flowering cabbage and controlling Plutella xylostella. Pest Manag. Sci. 2021, 78, 758–769. [Google Scholar] [CrossRef]
- Shabbir, M.; Afsar, T.; Razak, S.; Almajwal, A.; Khan, M.R. Phytochemical analysis and Evaluation of hepatoprotective effect of Maytenus royleanus leaves extract against anti-tuberculosis drug induced liver injury in mice. Lipids Health Dis. 2020, 19, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Bilal, M.; Freed, S.; Ashraf, M.Z.; Rehan, A. Resistance and detoxification enzyme activities to bifenthrin in Oxycarenus hyalinipennis (Hemiptera: Lygaeidae). Crop Prot. 2018, 111, 17–22. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kristensen, M. Glutathione S-transferase and insecticide resistance in laboratory strains and field populations of Musca domestica. J. Econ. Entomol. 2005, 98, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Li, Z.; Li, D.; Wang, R.; Zhang, S.; You, H.; Li, J. Biochemical mechanisms, cross-resistance and stability of resistance to metaflumizone in Plutella xylostella. Insects 2020, 11, 311. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Wang, Q.; Qi, H.; Wang, Q.; Yuan, H.; Rui, C. Resistance selection of indoxacarb in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae): Cross-resistance, biochemical mechanisms and associated fitness costs. Pest Manag. Sci. 2018, 74, 2636–2644. [Google Scholar] [CrossRef]
- Zibaee, A.; Bandani, A.R.; Tork, M. Effect of the entomopathogenic fungus, Beauveria bassiana, and its secondary metabolite on detoxifying enzyme activities and acetylcholinesterase (AChE) of the Sunn pest, Eurygaster integriceps (Heteroptera: Scutellaridae). Biocontrol Sci. Technol. 2009, 19, 485–498. [Google Scholar] [CrossRef]
- Ahmed, R.; Freed, S. Biochemical resistance mechanisms against chlorpyrifos, imidacloprid and lambda-cyhalothrin in Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae). Crop Prot. 2021, 143, 105568. [Google Scholar] [CrossRef]
- He, Q.; Shi, J. Mesoporous silica nanoparticle based nano drug delivery systems: Synthesis, controlled drug release and delivery, pharmacokinetics and biocompatibility. J. Mater. Chem. 2011, 21, 5845–5855. [Google Scholar] [CrossRef]
- Colilla, M.; González, B.; Vallet-Regí, M. Mesoporous silica nanoparticles for the design of smart delivery nanodevices. Biomat. Sci. 2013, 1, 114–134. [Google Scholar] [CrossRef]
- Cao, L.; Zhang, H.; Zhou, Z.; Xu, C.; Shan, Y.; Lin, Y.; Huang, Q. Fluorophore-free luminescent double-shelled hollow mesoporous silica nanoparticles as pesticide delivery vehicles. Nanoscale 2018, 10, 20354–20365. [Google Scholar] [CrossRef]
- Lu, K.; Li, Z.; Hu, Q.; Sun, J.; Chen, M. CRPC Membrane-Camouflaged, Biomimetic Nanosystem for Overcoming Castration-Resistant Prostate Cancer by Cellular Vehicle-Aided Tumor Targeting. Int. J. Mol. Sci. 2022, 23, 3623. [Google Scholar] [CrossRef]
- Hauck, E.S.; Hecker, J.G. Non-Viral Delivery of RNA Gene Therapy to the Central Nervous System. Pharmaceutics 2022, 14, 165. [Google Scholar] [CrossRef]
- Yilmaz, B.; Ozay, O. Synthesis of antibiotic-modified silica nanoparticles and their use as a controlled drug release system with antibacterial properties. Phosphorus Sulfur Silicon Relat. Elem. 2022, 4, 1–9. [Google Scholar] [CrossRef]
- Zhang, M.; Feng, J.; Zhong, Y.; Luo, J.; Zhao, Y.; Yang, Y.; Song, Y.; Lin, X.; Yang, Y.; Song, H.; et al. In-Situ Synthesis of Drug-Containing Bactericidal Rough Silica Nanoparticles for Antibacterial Coating. Chem. Eng. J. 2022, 440, 135837. [Google Scholar] [CrossRef]
- Hassan, Y.M.; Guan, B.H.; Chuan, L.K.; Hamza, M.F.; Adil, M.; Adam, A.A. The synergistic effect of Fe2O3/SiO2 nanoparticles concentration on rheology, wettability, and brine-oil interfacial tension. J. Pet. Sci. Eng. 2022, 210, 110059. [Google Scholar] [CrossRef]
- Xu, C.; Shan, Y.; Bilal, M.; Xu, B.; Cao, L.; Huang, Q. Copper ions chelated mesoporous silica nanoparticles via dopamine chemistry for controlled pesticide release regulated by coordination bonding. Chem. Engin. J. 2020, 395, 125093. [Google Scholar] [CrossRef]
- Xu, C.; Cao, L.; Zhao, P.; Zhou, Z.; Cao, C.; Li, F.; Huang, Q. Emulsion-based synchronous pesticide encapsulation and surface modification of mesoporous silica nanoparticles with carboxymethyl chitosan for controlled azoxystrobin release. Chem. Eng. J. 2018, 348, 244–254. [Google Scholar] [CrossRef]
- Alizadeh, M.; Sheikhi-Garjan, A.; Ma’Mani, L.; Salekdeh, G.H.; Bandehagh, A. Ethology of Sunn-pest oviposition in interaction with deltamethrin loaded on mesoporous silica nanoparticles as a nanopesticide. Chem. Biol. Technol. Agric. 2022, 9, 1–13. [Google Scholar] [CrossRef]
- Xu, L.; Qin, J.; Fu, W.; Wang, S.; Wu, Q.; Zhou, X.; Crickmore, N.; Guo, Z.; Zhang, Y. MAP4K4 controlled transcription factor POUM1 regulates PxABCG1 expression influencing Cry1Ac resistance in Plutella xylostella (L.). Pestic. Biochem. Physiol. 2022, 182, 105053. [Google Scholar] [CrossRef]
- Meng, X.; Wu, Z.; Jiang, C.; Guan, D.; Zhang, N.; Jiang, H.; Shen, Q.; Qian, K.; Wang, J. Identification and characterization of glutathione S-transferases and their potential roles in detoxification of abamectin in the rice stem borer, Chilo suppressalis. Pestic. Biochem. Physiol. 2022, 182, 105050. [Google Scholar] [CrossRef]
- Wang, R.; Qu, C.; Wang, Z.; Yang, G. Cross-resistance, biochemical mechanism and fitness costs of laboratory-selected resistance to pyridalyl in diamondback moth, Plutella xylostella. Pestic. Biochem. Physiol. 2019, 163, 8–13. [Google Scholar] [CrossRef]
- Sayani, Z.; Mikani, A.; Mosallanejad, H. Biochemical resistance mechanisms to fenvalerate in Plutella xylostella (Lepidoptera: Plutellidae). J. Econ. Entomol. 2019, 112, 1372–1377. [Google Scholar] [CrossRef]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Harold, J.A.; Ottea, J.A. Characterization of esterases associated with profenofos resistance in the tobacco budworm, Heliothis virescens (F.). Arch. Insect Biochem. Physiol. 2000, 45, 47–59. [Google Scholar] [CrossRef]
- Kerns, D.D.; Yang, F.; Kerns, D.L.; Stewart, S.D. Evaluation of Bt resistance in Helicoverpa zea (Lepidoptera: Noctuidae) strains using various Bt cotton plant tissues. Pest Manag. Sci. 2022, 78, 95–103. [Google Scholar] [CrossRef]
- Monteiro, R.A.; Camara, M.C.; de Oliveira, J.L.; Campos, E.V.R.; Carvalho, L.B.; Proença, P.L.D.F.; Guilger-Casagrande, M.; Lima, R.; Nascimento, J.D.; Gonçalves, K.C.; et al. Zein based-nanoparticles loaded botanical pesticides in pest control: An enzyme stimuli-responsive approach aiming sustainable agriculture. J. Hazard. Mater. 2021, 417, 126004. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Kim, J.-E.; Lee, Y.-W.; Son, H. Fungal cytochrome P450s and the P450 complement (CYPome) of Fusarium graminearum. Toxins 2018, 10, 112. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.; Choi, B.; Park, W.; Kim, Y.; Kim, B.; Mun, S.; Choi, H.; Kim, D. Nuclear receptor HR96 up regulates cytochrome P450 for insecticide detoxification in Tribolium castaneum. Pest Manag. Sci. 2021, 78, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Rathore, S.; Varshney, A.; Mohan, S.; Dahiya, P. An innovative approach of bioremediation in enzymatic degradation of xenobiotics. Biotechnol. Genet. Eng. Rev. 2022, 38, 1–32. [Google Scholar] [CrossRef]
- Rosenberry, T.L. Catalysis by acetylcholinesterase: Evidence that the rate-limiting step for acylation with certain substrates precedes general acid-base catalysis. Proc. Natl. Acad. Sci. USA 1975, 72, 3834–3838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nematollahi, A.; Rezaei, F.; Afsharian, Z.; Mollakhalili-Meybodi, N. Diazinon reduction in food products: A comprehensive review of conventional and emerging processing methods. Environ. Sci. Pollut. Res. 2022, 23, 1–16. [Google Scholar] [CrossRef]
- Reynoso, E.C.; Romero-Guido, C.; Rebollar-Pérez, G.; Torres, E. Enzymatic biosensors for the detection of water pollutants. In Nanomaterials for Biocatalysis; Elsevier: Amsterdam, The Netherlands, 2022; pp. 463–511. [Google Scholar]
- Intirach, J.; Junkum, A.; Lumjuan, N.; Chaithong, U.; Somboon, P.; Jitpakdi, A.; Riyong, D.; Champakaew, D.; Muangmoon, R.; Chansang, A.; et al. Biochemical effects of Petroselinum crispum (Umbellifereae) essential oil on the pyrethroid resistant strains of Aedes aegypti (Diptera: Culicidae). Insects 2018, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Chandra, S.; Chauhan, L.K.S.; Narayan, G.; Chowdhuri, D.K. Cellular internalization and stress response of ingested amorphous silica nanoparticles in the midgut of Drosophila melanogaster. Biochim. et Biophys. Acta (BBA)-Gen. Subj. 2012, 1830, 2256–2266. [Google Scholar] [CrossRef] [PubMed]
- Carotenuto, R.; Tussellino, M.; Ronca, R.; Benvenuto, G.; Fogliano, C.; Fusco, S.; Netti, P.A. Toxic effects of SiO2NPs in early embryogenesis of Xenopus laevis. Chemosphere 2021, 289, 133233. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Yu, A.; Wang, G.; Zheng, F.; Jia, J.; Xu, H. Chitosan-based nanoparticles of avermectin to control pine wood nematodes. Int. J. Biol. Macromol. 2018, 112, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Novak, S.; Drobne, D.; Menard, A. Prolonged feeding of terrestrial isopod (Porcellio scaber, Isopoda, Crustacea) on TiO2 nanoparicles. Absence of toxic effect. ZooKeys 2012, 176, 261. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Size (nm) a | Size (nm) b | SBET (m2/g) | DBJH (nm) | Vt (cm3/g) | Zeta (mV) |
|---|---|---|---|---|---|---|
| FL-SiO2-70 nm | 102 | 312 | 881 | 2.74 | 1.5 | −8.2 |
| FL-SiO2-150 nm | 168 | 387 | 969 | 2.73 | 1.4 | −8.2 |
| Me@FL-SiO2-70 nm | 113 | 534 | 121 | 2.25 | 0.5 | −8.4 |
| Me@FL-SiO2-150 nm | 180 | 602 | 149 | 2.19 | 0.2 | −8.5 |
| Insecticide/Insecticide@FL-SiO2 NPs | LC90 mg/L | LC50 mg/L | FL a 95% CL a mg/L | Slope (±SE b) | Chi c | N |
|---|---|---|---|---|---|---|
| Me@FL-SiO2 NPs-70 nm | 71 | 14.4 | 10.5–18.01 | 1.024 ± 0.170 | 1.673 | 540 |
| Me@FL-SiO2 NPs-150 nm | 49 | 15.2 | 10.8–20.22 | 1.087 ± 0.155 | 1.032 | 540 |
| Methoxyfenozide (TC) | 1339 | 24.74 | 18.2–31.33 | 1.393 ± 0.190 | 0.603 | 540 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilal, M.; Sial, M.U.; Cao, L.; Huang, Q. Effects of Methoxyfenozide-Loaded Fluorescent Mesoporous Silica Nanoparticles on Plutella xylostella (L.) (Lepidoptera: Plutellidae) Mortality and Detoxification Enzyme Levels Activities. Int. J. Mol. Sci. 2022, 23, 5790. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105790
Bilal M, Sial MU, Cao L, Huang Q. Effects of Methoxyfenozide-Loaded Fluorescent Mesoporous Silica Nanoparticles on Plutella xylostella (L.) (Lepidoptera: Plutellidae) Mortality and Detoxification Enzyme Levels Activities. International Journal of Molecular Sciences. 2022; 23(10):5790. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105790
Chicago/Turabian StyleBilal, Muhammad, Muhammad Umair Sial, Lidong Cao, and Qiliang Huang. 2022. "Effects of Methoxyfenozide-Loaded Fluorescent Mesoporous Silica Nanoparticles on Plutella xylostella (L.) (Lepidoptera: Plutellidae) Mortality and Detoxification Enzyme Levels Activities" International Journal of Molecular Sciences 23, no. 10: 5790. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105790