
Histone Modifications and Non-Coding RNAs: Mutual Epigenetic Regulation and Role in Pathogenesis

Abstract
:1. Introduction
2. Histone Modifications and ncRNAs: The Theory
2.1. Histone Methylation and Demethylation
2.2. Histone Acetylation and Deacetylation
2.3. Histone Phosphorylation and Dephosphorylation
2.4. Histone Ubiquitylation and Deubiquitylation
2.5. Histone Sumoylation and Desumoylation
2.6. Histone Biotinylation and Debiotinylation
3. Histone Modifications and ncRNAs: From Theory to Praxis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, J.-W.; Huang, K.; Yang, C.; Kang, C.-S. Non-Coding RNAs as Regulators in Epigenetics. Oncol. Rep. 2017, 37, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bure, I.V.; Nemtsova, M.V. Methylation and Noncoding RNAs in Gastric Cancer: Everything Is Connected. Int. J. Mol. Sci. 2021, 22, 5683. [Google Scholar] [CrossRef] [PubMed]
- Fuso, A.; Raia, T.; Orticello, M.; Lucarelli, M. The Complex Interplay between DNA Methylation and MiRNAs in Gene Expression Regulation. Biochimie 2020, 173, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Jeltsch, A.; Broche, J.; Bashtrykov, P. Molecular Processes Connecting DNA Methylation Patterns with DNA Methyltransferases and Histone Modifications in Mammalian Genomes. Genes 2018, 9, 566. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, M.V.C.; Bourc’his, D. The Diverse Roles of DNA Methylation in Mammalian Development and Disease. Nat. Rev. Mol. Cell Biol. 2019, 20, 590–607. [Google Scholar] [CrossRef]
- Li, J.; Li, W.X.; Bai, C.; Song, Y. Particulate Matter-Induced Epigenetic Changes and Lung Cancer: Particulate Matter, Epigenetics and Lung Cancer. Clin. Respir. J. 2017, 11, 539–546. [Google Scholar] [CrossRef]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Kouzarides, T. Chromatin Modifications and Their Function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Araki, Y.; Mimura, T. The Mechanisms Underlying Chronic Inflammation in Rheumatoid Arthritis from the Perspective of the Epigenetic Landscape. J. Immunol. Res. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Piletič, K.; Kunej, T. MicroRNA Epigenetic Signatures in Human Disease. Arch. Toxicol. 2016, 90, 2405–2419. [Google Scholar] [CrossRef]
- Gardner, K.E.; Allis, C.D.; Strahl, B.D. OPERating ON Chromatin, a Colorful Language Where Context Matters. J. Mol. Biol. 2011, 409, 36–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Zhang, Z.; Krause, H.M. Long Noncoding RNAs and Repetitive Elements: Junk or Intimate Evolutionary Partners? Trends Genet. 2019, 35, 892–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of Transcription in Human Cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, O.; Baccarelli, A.A. Environmental Health and Long Non-Coding RNAs. Curr. Environ. Health Rep. 2016, 3, 178–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taft, R.J.; Pang, K.C.; Mercer, T.R.; Dinger, M.; Mattick, J.S. Non-Coding RNAs: Regulators of Disease: Non-Coding RNAs: Regulators of Disease. J. Pathol. 2010, 220, 126–139. [Google Scholar] [CrossRef]
- Yao, Q.; Chen, Y.; Zhou, X. The Roles of MicroRNAs in Epigenetic Regulation. Curr. Opin. Chem. Biol. 2019, 51, 11–17. [Google Scholar] [CrossRef]
- Hammond, S.M. An Overview of MicroRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Arif, K.M.T.; Elliott, E.K.; Haupt, L.M.; Griffiths, L.R. Regulatory Mechanisms of Epigenetic MiRNA Relationships in Human Cancer and Potential as Therapeutic Targets. Cancers 2020, 12, 2922. [Google Scholar] [CrossRef]
- Moazed, D. Small RNAs in Transcriptional Gene Silencing and Genome Defence. Nature 2009, 457, 413–420. [Google Scholar] [CrossRef]
- Houwing, S.; Kamminga, L.M.; Berezikov, E.; Cronembold, D.; Girard, A.; van den Elst, H.; Filippov, D.V.; Blaser, H.; Raz, E.; Moens, C.B.; et al. A Role for Piwi and PiRNAs in Germ Cell Maintenance and Transposon Silencing in Zebrafish. Cell 2007, 129, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Lin, H. PiRNAs in the Germ Line. Science 2007, 316, 397. [Google Scholar] [CrossRef] [PubMed]
- Bure, I.V.; Kuznetsova, E.B.; Zaletaev, D.V. Long Noncoding RNAs and Their Role in Oncogenesis. Mol. Biol. 2018, 52, 787–798. [Google Scholar] [CrossRef]
- Peng, X.-F.; Huang, S.-F.; Chen, L.-J.; Xu, L.; Ye, W.-C. Targeting Epigenetics and LncRNAs in Liver Disease: From Mechanisms to Therapeutics. Pharmacol. Res. 2021, 172, 105846. [Google Scholar] [CrossRef] [PubMed]
- Sigova, A.A.; Mullen, A.C.; Molinie, B.; Gupta, S.; Orlando, D.A.; Guenther, M.G.; Almada, A.E.; Lin, C.; Sharp, P.A.; Giallourakis, C.C.; et al. Divergent Transcription of Long Noncoding RNA/MRNA Gene Pairs in Embryonic Stem Cells. Proc. Natl. Acad. Sci. USA 2013, 110, 2876–2881. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Song, M.; Qu, J.; Liu, G.-H. Epigenetic Modifications in Cardiovascular Aging and Diseases. Circ. Res. 2018, 123, 773–786. [Google Scholar] [CrossRef]
- Guenzl, P.M.; Barlow, D.P. Macro LncRNAs: A New Layer of Cis -Regulatory Information in the Mammalian Genome. RNA Biol. 2012, 9, 731–741. [Google Scholar] [CrossRef] [Green Version]
- Melissari, M.-T.; Grote, P. Roles for Long Non-Coding RNAs in Physiology and Disease. Pflüg. Arch. - Eur. J. Physiol. 2016, 468, 945–958. [Google Scholar] [CrossRef]
- Hu, X.; Sood, A.K.; Dang, C.V.; Zhang, L. The Role of Long Noncoding RNAs in Cancer: The Dark Matter Matters. Curr. Opin. Genet. Dev. 2018, 48, 8–15. [Google Scholar] [CrossRef]
- Hombach, S.; Kretz, M. Non-Coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar] [CrossRef]
- Golbabapour, S.; Abdulla, M.A.; Hajrezaei, M. A Concise Review on Epigenetic Regulation: Insight into Molecular Mechanisms. Int. J. Mol. Sci. 2011, 12, 8661–8694. [Google Scholar] [CrossRef]
- Stylianou, E. Epigenetics of Chronic Inflammatory Diseases. J. Inflamm. Res. 2018, 12, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harr, J.C.; Gonzalez-Sandoval, A.; Gasser, S.M. Histones and Histone Modifications in Perinuclear Chromatin Anchoring: From Yeast to Man. EMBO Rep. 2016, 17, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Rando, O.J. Combinatorial Complexity in Chromatin Structure and Function: Revisiting the Histone Code. Curr. Opin. Genet. Dev. 2012, 22, 148–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taudt, A.; Colomé-Tatché, M.; Johannes, F. Genetic Sources of Population Epigenomic Variation. Nat. Rev. Genet. 2016, 17, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Greer, E.L.; Shi, Y. Histone Methylation: A Dynamic Mark in Health, Disease and Inheritance. Nat. Rev. Genet. 2012, 13, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Al-Hasani, K.; Mathiyalagan, P.; El-Osta, A. Epigenetics, Cardiovascular Disease, and Cellular Reprogramming. J. Mol. Cell. Cardiol. 2019, 128, 129–133. [Google Scholar] [CrossRef]
- Paige, S.L.; Thomas, S.; Stoick-Cooper, C.L.; Wang, H.; Maves, L.; Sandstrom, R.; Pabon, L.; Reinecke, H.; Pratt, G.; Keller, G.; et al. A Temporal Chromatin Signature in Human Embryonic Stem Cells Identifies Regulators of Cardiac Development. Cell 2012, 151, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Fischle, W.; Tseng, B.S.; Dormann, H.L.; Ueberheide, B.M.; Garcia, B.A.; Shabanowitz, J.; Hunt, D.F.; Funabiki, H.; Allis, C.D. Regulation of HP1–Chromatin Binding by Histone H3 Methylation and Phosphorylation. Nature 2005, 438, 1116–1122. [Google Scholar] [CrossRef]
- Zippo, A.; Serafini, R.; Rocchigiani, M.; Pennacchini, S.; Krepelova, A.; Oliviero, S. Histone Crosstalk between H3S10ph and H4K16ac Generates a Histone Code That Mediates Transcription Elongation. Cell 2009, 138, 1122–1136. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. In Epigenetics in Allergy and Autoimmunity; Chang, C., Lu, Q., Eds.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2020; Volume 1253, pp. 3–55. ISBN 9789811534485. [Google Scholar]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. MiRBase: From MicroRNA Sequences to Function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Sætrom, P.; Snøve, O.; Rossi, J.J. Epigenetics and MicroRNAs. Pediatr. Res. 2007, 61, 17R–23R. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, J.; Li, Y.; Song, T.; Wu, Y.; Fang, S.; Bu, D.; Li, H.; Sun, L.; Pei, D.; et al. NONCODEV6: An Updated Database Dedicated to Long Non-Coding RNA Annotation in Both Animals and Plants. Nucleic Acids Res. 2021, 49, D165–D171. [Google Scholar] [CrossRef] [PubMed]
- Haberland, M.; Montgomery, R.L.; Olson, E.N. The Many Roles of Histone Deacetylases in Development and Physiology: Implications for Disease and Therapy. Nat. Rev. Genet. 2009, 10, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Xia, K.; Yu, L.-Y.; Huang, X.-Q.; Zhao, Z.-H.; Liu, J. Epigenetic Regulation by Long Noncoding RNAs in Osteo-/Adipogenic Differentiation of Mesenchymal Stromal Cells and Degenerative Bone Diseases. World J. Stem Cells 2022, 14, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Malumbres, M. MiRNAs and Cancer: An Epigenetics View. Mol. Aspects Med. 2013, 34, 863–874. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Chen, Q.; Zhang, X.; Yang, J.; Lin, K.; Ji, C.; Xu, A.; Yang, L.; Miao, L. Long Noncoding RNA ANRIL Promotes the Malignant Progression of Cholangiocarcinoma by Epigenetically Repressing ERRFI1 Expression. Cancer Sci. 2020, 111, 2297–2309. [Google Scholar] [CrossRef]
- Ni, H.; Chai, P.; Yu, J.; Xing, Y.; Wang, S.; Fan, J.; Ge, S.; Wang, Y.; Jia, R.; Fan, X. LncRNA CANT1 Suppresses Retinoblastoma Progression by Repellinghistone Methyltransferase in PI3Kγ Promoter. Cell Death Dis. 2020, 11, 306. [Google Scholar] [CrossRef]
- Li, F.-P.; Ling, D.-Q.; Gao, L.-Y. Long Noncoding RNA Chaer Mediated Polycomb Repressor Complex 2 (PRC2) Activity to Promote Atherosclerosis through MTOR Signaling. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7639–7648. [Google Scholar] [CrossRef]
- Wang, Q.; Zhuo, Z.; Yu, H.; Wang, C.; Gao, L. Circ_0019435 Exerts Its Functions in the Cellular Process of Cervical Cancer via Epigenetically Silencing DKK1 and PTEN. Reprod. Sci. 2021, 28, 2989–2999. [Google Scholar] [CrossRef]
- Wang, X.-H.; Li, J. CircAGFG1 Aggravates the Progression of Cervical Cancer by Downregulating P53. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1704–1711. [Google Scholar] [CrossRef]
- Hatanaka, Y.; Niinuma, T.; Kitajima, H.; Nishiyama, K.; Maruyama, R.; Ishiguro, K.; Toyota, M.; Yamamoto, E.; Kai, M.; Yorozu, A.; et al. DLEU1 Promotes Oral Squamous Cell Carcinoma Progression by Activating Interferon-Stimulated Genes. Sci. Rep. 2021, 11, 20438. [Google Scholar] [CrossRef]
- Chen, N.; Guo, D.; Xu, Q.; Yang, M.; Wang, D.; Peng, M.; Ding, Y.; Wang, S.; Zhou, J. Long Non-Coding RNA FEZF1-AS1 Facilitates Cell Proliferation and Migration in Colorectal Carcinoma. Oncotarget 2016, 7, 11271–11283. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Yu, S.; Xue, Y.; Yu, H.; Chen, D.; Wei, X.; Liu, Y. Over-Expressed FEZF1 Predicts a Poor Prognosis in Glioma and Promotes Glioma Cell Malignant Biological Properties by Regulating Akt-ERK Pathway. J. Mol. Neurosci. 2018, 65, 411–419. [Google Scholar] [CrossRef]
- Liu, Y.; Xia, R.; Lu, K.; Xie, M.; Yang, F.; Sun, M.; De, W.; Wang, C.; Ji, G. LincRNAFEZF1-AS1 Represses P21 Expression to Promote Gastric Cancer Proliferation through LSD1-Mediated H3K4me2 Demethylation. Mol. Cancer 2017, 16, 39. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Bonora, G.; Lewandowski, J.P.; Thakur, J.; Filippova, G.N.; Henikoff, S.; Shendure, J.; Duan, Z.; Rinn, J.L.; Deng, X.; et al. Trans- and Cis-Acting Effects of Firre on Epigenetic Features of the Inactive X Chromosome. Nat. Commun. 2020, 11, 6053. [Google Scholar] [CrossRef]
- Pan, W.; Zhang, N.; Liu, W.; Liu, J.; Zhou, L.; Liu, Y.; Yang, M. The Long Noncoding RNA GAS8-AS1 Suppresses Hepatocarcinogenesis by Epigenetically Activating the Tumor Suppressor GAS8. J. Biol. Chem. 2018, 293, 17154–17165. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Ramnarine, V.R.; Song, J.H.; Pandey, R.; Padi, S.K.R.; Nouri, M.; Olive, V.; Kobelev, M.; Okumura, K.; McCarthy, D.; et al. The Long Noncoding RNA H19 Regulates Tumor Plasticity in Neuroendocrine Prostate Cancer. Nat. Commun. 2021, 12, 7349. [Google Scholar] [CrossRef]
- Yuan, Z.; Bian, Y.; Ma, X.; Tang, Z.; Chen, N.; Shen, M. LncRNA H19 Knockdown in Human Amniotic Mesenchymal Stem Cells Suppresses Angiogenesis by Associating with EZH2 and Activating Vasohibin-1. Stem Cells Dev. 2019, 28, 781–790. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.-C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long Non-Coding RNA HOTAIR Reprograms Chromatin State to Promote Cancer Metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Song, Y.; Wang, R.; Li, L.-W.; Liu, X.; Wang, Y.-F.; Wang, Q.-X.; Zhang, Q. Long Non-Coding RNA HOTAIR Mediates the Switching of Histone H3 Lysine 27 Acetylation to Methylation to Promote Epithelial-to-Mesenchymal Transition in Gastric Cancer. Int. J. Oncol. 2018, 54, 77–86. [Google Scholar] [CrossRef]
- Gao, S.; Zhou, B.; Li, H.; Huang, X.; Wu, Y.; Xing, C.; Yu, X.; Ji, Y. Long Noncoding RNA HOTAIR Promotes the Self-Renewal of Leukemia Stem Cells through Epigenetic Silencing of P15. Exp. Hematol. 2018, 67, 32–40.e3. [Google Scholar] [CrossRef]
- Xia, M.; Yao, L.; Zhang, Q.; Wang, F.; Mei, H.; Guo, X.; Huang, W. Long Noncoding RNA HOTAIR Promotes Metastasis of Renal Cell Carcinoma by Up-Regulating Histone H3K27 Demethylase JMJD3. Oncotarget 2017, 8, 19795–19802. [Google Scholar] [CrossRef] [Green Version]
- Yi, E.; Zhang, J.; Zheng, M.; Zhang, Y.; Liang, C.; Hao, B.; Hong, W.; Lin, B.; Pu, J.; Lin, Z.; et al. Long Noncoding RNA IL6-AS1 Is Highly Expressed in Chronic Obstructive Pulmonary Disease and Is Associated with Interleukin 6 by Targeting MiR-149-5p and Early B-cell Factor 1. Clin. Transl. Med. 2021, 11, e479. [Google Scholar] [CrossRef]
- Wang, W.-T.; Chen, T.-Q.; Zeng, Z.-C.; Pan, Q.; Huang, W.; Han, C.; Fang, K.; Sun, L.-Y.; Yang, Q.-Q.; Wang, D.; et al. The LncRNA LAMP5-AS1 Drives Leukemia Cell Stemness by Directly Modulating DOT1L Methyltransferase Activity in MLL Leukemia. J. Hematol. Oncol.J Hematol Oncol 2020, 13, 78. [Google Scholar] [CrossRef]
- Liu, J.; Feng, Y.; Zeng, X.; He, M.; Gong, Y.; Liu, Y. Extracellular Vesicles-encapsulated Let-7i Shed from Bone Mesenchymal Stem Cells Suppress Lung Cancer via KDM3A/DCLK1/FXYD3 Axis. J. Cell. Mol. Med. 2021, 25, 1911–1926. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, X.; Ma, L.; Ding, R.; Zhao, L.; Ma, F.; Deng, X. LINC01419 Facilitates Hepatocellular Carcinoma Growth and Metastasis through Targeting EZH2-Regulated RECK. Aging 2020, 12, 11071–11084. [Google Scholar] [CrossRef]
- Wu, L.; Gong, Y.; Yan, T.; Zhang, H. LINP1 Promotes the Progression of Cervical Cancer by Scaffolding EZH2, LSD1, and DNMT1 to Inhibit the Expression of KLF2 and PRSS8. Biochem. Cell Biol. 2020, 98, 591–599. [Google Scholar] [CrossRef]
- Chen, X.; An, N. Long Noncoding RNA ATB Promotes Ovarian Cancer Tumorigenesis by Mediating Histone H3 Lysine 27 Trimethylation through Binding to EZH2. J. Cell. Mol. Med. 2021, 25, 37–46. [Google Scholar] [CrossRef]
- Sun, Y.; Cai, M.; Zhong, J.; Yang, L.; Xiao, J.; Jin, F.; Xue, H.; Liu, X.; Liu, H.; Zhang, Y.; et al. The Long Noncoding RNA Lnc-Ob1 Facilitates Bone Formation by Upregulating Osterix in Osteoblasts. Nat. Metab. 2019, 1, 485–496. [Google Scholar] [CrossRef]
- Liu, F.; Song, D.-Y.; Huang, J.; Yang, H.-Q.; You, D.; Ni, J.-D. Long Non-Coding RNA CIR Inhibits Chondrogenic Differentiation of Mesenchymal Stem Cells by Epigenetically Suppressing ATOH8 via Methyltransferase EZH2. Mol. Med. 2021, 27, 12. [Google Scholar] [CrossRef]
- Hu, A.; Hong, F.; Li, D.; Jin, Y.; Kon, L.; Xu, Z.; He, H.; Xie, Q. Long Non-Coding RNA ROR Recruits Histone Transmethylase MLL1 to up-Regulate TIMP3 Expression and Promote Breast Cancer Progression. J. Transl. Med. 2021, 19, 95. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Wang, K.; Yeh, S.; Sun, Y.; Liang, L.; Xiao, Y.; Xu, W.; Niu, Y.; Cheng, L.; Maity, S.N.; et al. LncRNA-P21 Alters the Antiandrogen Enzalutamide-Induced Prostate Cancer Neuroendocrine Differentiation via Modulating the EZH2/STAT3 Signaling. Nat. Commun. 2019, 10, 2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, N.; Yao, Z.; Xu, M.; Chen, J.; Lu, Y.; Yuan, L.; Zhou, S.; Zou, X.; Xu, R. Long Noncoding RNA MALAT1 Potentiates Growth and Inhibits Senescence by Antagonizing ABI3BP in Gallbladder Cancer Cells. J. Exp. Clin. Cancer Res. 2019, 38, 244. [Google Scholar] [CrossRef]
- Qu, D.; Sun, W.-W.; Li, L.; Ma, L.; Sun, L.; Jin, X.; Li, T.; Hou, W.; Wang, J.-H. Long Noncoding RNA MALAT1 Releases Epigenetic Silencing of HIV-1 Replication by Displacing the Polycomb Repressive Complex 2 from Binding to the LTR Promoter. Nucleic Acids Res. 2019, 47, 3013–3027. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.; Thomas, A.A.; Chen, S.; Aref-Eshghi, E.; Feng, B.; Gonder, J.; Sadikovic, B.; Chakrabarti, S. MALAT1: An Epigenetic Regulator of Inflammation in Diabetic Retinopathy. Sci. Rep. 2018, 8, 6526. [Google Scholar] [CrossRef]
- Li, D.-Q.; Ding, Y.-R.; Che, J.-H.; Su, Z.; Yang, W.-Z.; Xu, L.; Li, Y.-J.; Wang, H.-H.; Zhou, W.-Y. Tumor Suppressive LncRNA MEG3 Binds to EZH2 and Enhances CXCL3 Methylation in Gallbladder Cancer. Neoplasma 2022, 210726N1046. [Google Scholar] [CrossRef]
- Zhang, W.; Cao, D.; Wang, Y.; Ren, W. LncRNA MEG8 Is Upregulated in Gestational Diabetes Mellitus (GDM) and Predicted Kidney Injury. J. Diabetes Complications 2021, 35, 107749. [Google Scholar] [CrossRef]
- Terashima, M.; Ishimura, A.; Wanna-udom, S.; Suzuki, T. MEG8 Long Noncoding RNA Contributes to Epigenetic Progression of the Epithelial-Mesenchymal Transition of Lung and Pancreatic Cancer Cells. J. Biol. Chem. 2018, 293, 18016–18030. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Jia, J.; Zang, Y.; Li, J.; Wan, B. MicroRNA-101 Regulates Autophagy, Proliferation and Apoptosis via Targeting EZH2 in Laryngeal Squamous Cell Carcinoma. Neoplasma 2019, 66, 507–515. [Google Scholar] [CrossRef]
- Liu, X.; Lei, Q.; Yu, Z.; Xu, G.; Tang, H.; Wang, W.; Wang, Z.; Li, G.; Wu, M. MiR-101 Reverses the Hypomethylation of the LMO3 Promoter in Glioma Cells. Oncotarget 2015, 6, 7930–7943. [Google Scholar] [CrossRef] [Green Version]
- Tian, Z.; Li, Z.; Zhu, Y.; Meng, L.; Liu, F.; Sang, M.; Wang, G. Hypermethylation-Mediated Inactivation of MiR-124 Predicts Poor Prognosis and Promotes Tumor Growth at Least Partially through Targeting EZH2/H3K27me3 in ESCC. Clin. Exp. Metastasis 2019, 36, 381–391. [Google Scholar] [CrossRef]
- Wang, Z.; Dai, J.; Yan, J.; Zhang, Y.; Yin, Z. Targeting EZH 2 as a Novel Therapeutic Strategy for Sorafenib-resistant Thyroid Carcinoma. J. Cell. Mol. Med. 2019, 23, 4770–4778. [Google Scholar] [CrossRef] [Green Version]
- Villeneuve, L.M.; Kato, M.; Reddy, M.A.; Wang, M.; Lanting, L.; Natarajan, R. Enhanced Levels of MicroRNA-125b in Vascular Smooth Muscle Cells of Diabetic Db/Db Mice Lead to Increased Inflammatory Gene Expression by Targeting the Histone Methyltransferase Suv39h1. Diabetes 2010, 59, 2904–2915. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yan, T.; Liu, Z.; Wang, J.; Lu, Y.; Li, D.; Liang, W. MicroRNA-137 Is Negatively Associated with Clinical Outcome and Regulates Tumor Development through EZH2 in Cervical Cancer. J. Cell. Biochem. 2018, 119, 938–947. [Google Scholar] [CrossRef]
- Huang, Y.; Zou, Y.; Zheng, R.; Ma, X. MiR-137 Inhibits Cell Proliferation in Acute Lymphoblastic Leukemia by Targeting JARID1B. Eur. J. Haematol. 2019, 103, 215–224. [Google Scholar] [CrossRef]
- Singh, S.K.; Bahal, R.; Rasmussen, T.P. Evidence That MiR-152-3p Is a Positive Regulator of SETDB1-Mediated H3K9 Histone Methylation and Serves as a Toggle between Histone and DNA Methylation. Exp. Cell Res. 2020, 395, 112216. [Google Scholar] [CrossRef]
- Brunty, S.; Ray Wright, K.; Mitchell, B.; Santanam, N. Peritoneal Modulators of EZH2-MiR-155 Cross-Talk in Endometriosis. Int. J. Mol. Sci. 2021, 22, 3492. [Google Scholar] [CrossRef]
- Sengupta, D.; Deb, M.; Kar, S.; Parbin, S.; Pradhan, N.; Patra, S.K. MiR-193a Targets MLL1 MRNA and Drastically Decreases MLL1 Protein Production: Ectopic Expression of the MiRNA Aberrantly Lowers H3K4me3 Content of the Chromatin and Hampers Cell Proliferation and Viability. Gene 2019, 705, 22–35. [Google Scholar] [CrossRef]
- Kang, W.-L.; Xu, G.-S. Atrasentan Increased the Expression of Klotho by Mediating MiR-199b-5p and Prevented Renal Tubular Injury in Diabetic Nephropathy. Sci. Rep. 2016, 6, 19979. [Google Scholar] [CrossRef]
- Bai, S.; Xiong, X.; Tang, B.; Ji, T.; Li, X.; Qu, X.; Li, W. Hsa-MiR-199b-3p Prevents the Epithelial-Mesenchymal Transition and Dysfunction of the Renal Tubule by Regulating E-Cadherin through Targeting KDM6A in Diabetic Nephropathy. Oxid. Med. Cell. Longev. 2021, 2021, 1–17. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Y.; Li, G.; Li, L.; Geng, P.; Song, H. MicroRNA-214 Suppresses the Growth of Cervical Cancer Cells by Targeting EZH2. Oncol. Lett. 2018, 16, 5679–5686. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Xu, T.; Fan, L.; Liu, K.; Li, G. MicroRNA-216b Enhances Cisplatin-Induced Apoptosis in Osteosarcoma MG63 and SaOS-2 Cells by Binding to JMJD2C and Regulating the HIF1α/HES1 Signaling Axis. J. Exp. Clin. Cancer Res. 2020, 39, 201. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Zhang, W.; Yang, Y.; Meng, Q.; Wang, C.; Xin, X.; Jiang, X.; Song, S.; Lu, Y.; et al. MiR24-2 Promotes Malignant Progression of Human Liver Cancer Stem Cells by Enhancing Tyrosine Kinase Src Epigenetically. Mol. Ther. 2020, 28, 572–586. [Google Scholar] [CrossRef]
- Lyu, G.; Guan, Y.; Zhang, C.; Zong, L.; Sun, L.; Huang, X.; Huang, L.; Zhang, L.; Tian, X.-L.; Zhou, Z.; et al. TGF-β Signaling Alters H4K20me3 Status via MiR-29 and Contributes to Cellular Senescence and Cardiac Aging. Nat. Commun. 2018, 9, 2560. [Google Scholar] [CrossRef]
- Wu, Y.; Shi, W.; Tang, T.; Wang, Y.; Yin, X.; Chen, Y.; Zhang, Y.; Xing, Y.; Shen, Y.; Xia, T.; et al. MiR-29a Contributes to Breast Cancer Cells Epithelial–Mesenchymal Transition, Migration, and Invasion via down-Regulating Histone H4K20 Trimethylation through Directly Targeting SUV420H2. Cell Death Dis. 2019, 10, 176. [Google Scholar] [CrossRef]
- Yu, N.; Huangyang, P.; Yang, X.; Han, X.; Yan, R.; Jia, H.; Shang, Y.; Sun, L. MicroRNA-7 Suppresses the Invasive Potential of Breast Cancer Cells and Sensitizes Cells to DNA Damages by Targeting Histone Methyltransferase SET8. J. Biol. Chem. 2013, 288, 19633–19642. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.Y.; Shin, H.J.; Bae, I.H. MiR-93-5p Suppresses Cellular Senescence by Directly Targeting Bcl-w and P21. Biochem. Biophys. Res. Commun. 2018, 505, 1134–1140. [Google Scholar] [CrossRef]
- Chen, Q.; Cai, J.; Wang, Q.; Wang, Y.; Liu, M.; Yang, J.; Zhou, J.; Kang, C.; Li, M.; Jiang, C. Long Noncoding RNA NEAT1, Regulated by the EGFR Pathway, Contributes to Glioblastoma Progression Through the WNT/β -Catenin Pathway by Scaffolding EZH2. Clin. Cancer Res. 2018, 24, 684–695. [Google Scholar] [CrossRef] [Green Version]
- Butler, A.A.; Johnston, D.R.; Kaur, S.; Lubin, F.D. Long Noncoding RNA NEAT1 Mediates Neuronal Histone Methylation and Age-Related Memory Impairment. Sci. Signal. 2019, 12, eaaw9277. [Google Scholar] [CrossRef]
- Hiura, K.; Strahan, R.; Uppal, T.; Prince, B.; Rossetto, C.C.; Verma, S.C. KSHV ORF59 and PAN RNA Recruit Histone Demethylases to the Viral Chromatin during Lytic Reactivation. Viruses 2020, 12, 420. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Wang, S.; Meng, J.; Lyu, G.; Ding, G.; Hu, Y.; Wang, L.; Wu, L.; Yang, W.; Lv, Y.; et al. Long Noncoding RNA PART1 Restrains Aggressive Gastric Cancer through the Epigenetic Silencing of PDGFB via the PLZF-Mediated Recruitment of EZH2. Oncogene 2020, 39, 6513–6528. [Google Scholar] [CrossRef]
- Fu, Y.; Zhang, X.; Liu, X.; Wang, P.; Chu, W.; Zhao, W.; Wang, Y.; Zhou, G.; Yu, Y.; Zhang, H. The DNMT1-PAS1-PH20 Axis Drives Breast Cancer Growth and Metastasis. Signal Transduct. Target. Ther. 2022, 7, 81. [Google Scholar] [CrossRef]
- Chu, W.; Zhang, X.; Qi, L.; Fu, Y.; Wang, P.; Zhao, W.; Du, J.; Zhang, J.; Zhan, J.; Wang, Y.; et al. The EZH2–PHACTR2–AS1–Ribosome Axis Induces Genomic Instability and Promotes Growth and Metastasis in Breast Cancer. Cancer Res. 2020, 80, 2737–2750. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.-J.; Cheng, Y.-M.; Wang, C.-L. LncRNA PVT1 Epigenetically Silences MiR-195 and Modulates EMT and Chemoresistance in Cervical Cancer Cells. J. Drug Target. 2017, 25, 637–644. [Google Scholar] [CrossRef]
- Li, Z.; Guo, X.; Wu, S. Epigenetic Silencing of KLF2 by Long Non-Coding RNA SNHG1 Inhibits Periodontal Ligament Stem Cell Osteogenesis Differentiation. Stem Cell Res. Ther. 2020, 11, 435. [Google Scholar] [CrossRef]
- Xiang, W.; Lyu, L.; Huang, T.; Zheng, F.; Yuan, J.; Zhang, C.; Jiang, G. The Long Non-coding RNA SNHG1 Promotes Bladder Cancer Progression by Interacting with MiR-143-3p and EZH2. J. Cell. Mol. Med. 2020, 24, 11858–11873. [Google Scholar] [CrossRef]
- Mao, X.; Ji, T.; Liu, A.; Weng, Y. ELK4-Mediated LncRNA SNHG22 Promotes Gastric Cancer Progression through Interacting with EZH2 and Regulating MiR-200c-3p/Notch1 Axis. Cell Death Dis. 2021, 12, 957. [Google Scholar] [CrossRef]
- Chi, C.; Li, M.; Hou, W.; Chen, Y.; Zhang, Y.; Chen, J. Long Noncoding RNA SNHG7 Activates Wnt/β-Catenin Signaling Pathway in Cervical Cancer Cells by Epigenetically Silencing DKK1. Cancer Biother. Radiopharm. 2020, 35, 329–337. [Google Scholar] [CrossRef]
- Qu, X.; Li, Y.; Wang, L.; Yuan, N.; Ma, M.; Chen, Y. LncRNA SNHG8 Accelerates Proliferation and Inhibits Apoptosis in HPV-induced Cervical Cancer through Recruiting EZH2 to Epigenetically Silence RECK Expression. J. Cell. Biochem. 2020, 121, 4120–4129. [Google Scholar] [CrossRef]
- Wang, B.; Zhao, L.; Chi, W.; Cao, H.; Cui, W.; Meng, W. Aberrant Methylation-Mediated Downregulation of LncRNA SSTR5-AS1 Promotes Progression and Metastasis of Laryngeal Squamous Cell Carcinoma. Epigenetics Chromatin 2019, 12, 35. [Google Scholar] [CrossRef]
- Hu, J.; Shan, Y.; Ma, J.; Pan, Y.; Zhou, H.; Jiang, L.; Jia, L. LncRNA ST3Gal6-AS1/ST3Gal6 Axis Mediates Colorectal Cancer Progression by Regulating A-2,3 Sialylation via PI3K/Akt Signaling. Int. J. Cancer 2019, 145, 450–460. [Google Scholar] [CrossRef]
- Papoutsoglou, P.; Tsubakihara, Y.; Caja, L.; Morén, A.; Pallis, P.; Ameur, A.; Heldin, C.-H.; Moustakas, A. The TGFB2-AS1 LncRNA Regulates TGF-β Signaling by Modulating Corepressor Activity. Cell Rep. 2019, 28, 3182–3198.e11. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yu, D.; Li, H.; Lv, Y.; Li, S. Long Non-coding RNA UCA1 Confers Tamoxifen Resistance in Breast Cancer Endocrinotherapy through Regulation of the EZH2/P21 Axis and the PI3K/AKT Signaling Pathway. Int. J. Oncol. 2019. [Google Scholar] [CrossRef]
- Zhang, J.; Li, W.-Y.; Yang, Y.; Yan, L.-Z.; Zhang, S.-Y.; He, J.; Wang, J.-X. LncRNA XIST Facilitates Cell Growth, Migration and Invasion via Modulating H3 Histone Methylation of DKK1 in Neuroblastoma. Cell Cycle 2019, 18, 1882–1892. [Google Scholar] [CrossRef]
- Wen, Z.; Lian, L.; Ding, H.; Hu, Y.; Xiao, Z.; Xiong, K.; Yang, Q. LncRNA ANCR Promotes Hepatocellular Carcinoma Metastasis through Upregulating HNRNPA1 Expression. RNA Biol. 2020, 17, 381–394. [Google Scholar] [CrossRef]
- Chi, J.; Li, J.; Jia, J.; Zhang, T.; Liu, X.; Yi, L. Long Non-Coding RNA ANRIL in Gene Regulation and Its Duality in Atherosclerosis. Curr. Med. Sci. 2017, 37, 816–822. [Google Scholar] [CrossRef]
- Zhang, C.; Ge, S.; Gong, W.; Xu, J.; Guo, Z.; Liu, Z.; Gao, X.; Wei, X.; Ge, S. LncRNA ANRIL Acts as a Modular Scaffold of WDR5 and HDAC3 Complexes and Promotes Alteration of the Vascular Smooth Muscle Cell Phenotype. Cell Death Dis. 2020, 11, 435. [Google Scholar] [CrossRef]
- Han, D.; Wang, Y.; Wang, Y.; Dai, X.; Zhou, T.; Chen, J.; Tao, B.; Zhang, J.; Cao, F. The Tumor-Suppressive Human Circular RNA CircITCH Sponges MiR-330-5p to Ameliorate Doxorubicin-Induced Cardiotoxicity Through Upregulating SIRT6, Survivin, and SERCA2a. Circ. Res. 2020, 127. [Google Scholar] [CrossRef]
- Jie, M.; Wu, Y.; Gao, M.; Li, X.; Liu, C.; Ouyang, Q.; Tang, Q.; Shan, C.; Lv, Y.; Zhang, K.; et al. CircMRPS35 Suppresses Gastric Cancer Progression via Recruiting KAT7 to Govern Histone Modification. Mol. Cancer 2020, 19, 56. [Google Scholar] [CrossRef]
- Chen, J.; Liu, G.; Wu, Y.; Ma, J.; Wu, H.; Xie, Z.; Chen, S.; Yang, Y.; Wang, S.; Shen, P.; et al. CircMYO10 Promotes Osteosarcoma Progression by Regulating MiR-370-3p/RUVBL1 Axis to Enhance the Transcriptional Activity of β-Catenin/LEF1 Complex via Effects on Chromatin Remodeling. Mol. Cancer 2019, 18, 150. [Google Scholar] [CrossRef] [Green Version]
- Yue, Z.; Shusheng, J.; Hongtao, S.; Shu, Z.; Lan, H.; Qingyuan, Z.; Shaoqiang, C.; Yuanxi, H. Silencing DSCAM-AS1 Suppresses the Growth and Invasion of ER-Positive Breast Cancer Cells by Downregulating Both DCTPP1 and QPRT. Aging 2020, 12, 14754–14774. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Xu, N.; Zhang, H.; Liao, W.; Wang, Y.; Wang, S.; Zhang, S.; Jiang, Y.; Xie, W.; Zhang, Y. Persistent High Glucose Induced EPB41L4A-AS1 Inhibits Glucose Uptake via GCN5 Mediating Crotonylation and Acetylation of Histones and Non-histones. Clin. Transl. Med. 2022, 12, e699. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Xu, B.; Xu, W.; Xia, L.; Xu, Z.; Shen, L.; Peng, W.; Huang, S. Long Noncoding RNA GAS5 Inhibits Cell Proliferation and Fibrosis in Diabetic Nephropathy by Sponging MiR-221 and Modulating SIRT1 Expression. Aging 2019, 11, 8745–8759. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, H.; Fan, Z.; Li, G.; Ma, Q.; Tao, Z.; Wang, R.; Feng, J.; Luo, Y. Long Noncoding RNA H19 Promotes Neuroinflammation in Ischemic Stroke by Driving Histone Deacetylase 1-Dependent M1 Microglial Polarization. Stroke 2017, 48, 2211–2221. [Google Scholar] [CrossRef]
- Zhang, L.; Yu, Z.; Qu, Q.; Li, X.; Lu, X.; Zhang, H. Exosomal LncRNA HOTAIR Promotes the Progression and Angiogenesis of Endometriosis via the MiR-761/HDAC1 Axis and Activation of STAT3-Mediated Inflammation. Int. J. Nanomedicine 2022, Volume 17, 1155–1170. [Google Scholar] [CrossRef]
- Fu, L.; Peng, S.; Wu, W.; Ouyang, Y.; Tan, D.; Fu, X. LncRNA HOTAIRM1 Promotes Osteogenesis by Controlling JNK/AP-1 Signalling-mediated RUNX2 Expression. J. Cell. Mol. Med. 2019, 23, 7517–7524. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Xiang, L.; He, D.; Zhu, R.; Fang, J.; Wang, Z.; Cao, K. The Long Noncoding RNA KTN1-AS1 Promotes Bladder Cancer Tumorigenesis via KTN1 Cis -Activation and the Consequent Initiation of Rho GTPase-Mediated Signaling. Clin. Sci. 2021, 135, 555–574. [Google Scholar] [CrossRef]
- Fan, W.; Wen, X.; Zheng, J.; Wang, K.; Qiu, H.; Zhang, J.; Su, F. LINC00162 Participates in the Pathogenesis of Diabetic Nephropathy via Modulating the MiR-383/HDAC9 Signalling Pathway. Artif. Cells Nanomed. Biotechnol. 2020, 48, 1047–1054. [Google Scholar] [CrossRef]
- He, B.; Pan, H.; Zheng, F.; Chen, S.; Bie, Q.; Cao, J.; Zhao, R.; Liang, J.; Wei, L.; Zeng, J.; et al. Long Noncoding RNA LINC00930 Promotes PFKFB3-Mediated Tumor Glycolysis and Cell Proliferation in Nasopharyngeal Carcinoma. J. Exp. Clin. Cancer Res. 2022, 41, 77. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, P.; Shao, Q.; Wang, J. Lipopolysaccharide Promotes Pulmonary Fibrosis in Acute Respiratory Distress Syndrome (ARDS) via LincRNA-P21 Induced Inhibition of Thy-1 Expression. Mol. Cell. Biochem. 2016, 419, 19–28. [Google Scholar] [CrossRef]
- Zhang, L.; Niu, H.; Ma, J.; Yuan, B.-Y.; Chen, Y.-H.; Zhuang, Y.; Chen, G.-W.; Zeng, Z.-C.; Xiang, Z.-L. The Molecular Mechanism of LncRNA34a-Mediated Regulation of Bone Metastasis in Hepatocellular Carcinoma. Mol. Cancer 2019, 18, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, A.K.; Xi, Y.; McCarthy, R.; Allton, K.; Akdemir, K.C.; Patel, L.R.; Aronow, B.; Lin, C.; Li, W.; Yang, L.; et al. LncPRESS1 Is a P53-Regulated LncRNA That Safeguards Pluripotency by Disrupting SIRT6-Mediated De-Acetylation of Histone H3K56. Mol. Cell 2016, 64, 967–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, H.; Wang, F.; Shi, X.; Ma, H.; Du, Y.; Hou, L.; Xing, N. LncRNA MALAT1 Induces the Dysfunction of β Cells via Reducing the Histone Acetylation of the PDX-1 Promoter in Type 1 Diabetes. Exp. Mol. Pathol. 2020, 114, 104432. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Yang, Y.; Ran, J.; Zhang, L.; Yao, M.; Liu, Z.; Zhang, L. MiR-15a-5p Inhibits Metastasis and Lipid Metabolism by Suppressing Histone Acetylation in Lung Cancer. Free Radic. Biol. Med. 2020, 161, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Li, Z.; Zhang, Z.; Yang, Z.; Kang, Y.; Zhao, X.; Long, D.; Hu, S.; Gu, M.; He, S.; et al. MicroRNA-193b-3p Regulates Chondrogenesis and Chondrocyte Metabolism by Targeting HDAC3. Theranostics 2018, 8, 2862–2883. [Google Scholar] [CrossRef]
- Wu, J.-C.; Jiang, H.-M.; Yang, X.-H.; Zheng, H.-C. ING5-Mediated Antineuroblastoma Effects of Suberoylanilide Hydroxamic Acid. Cancer Med. 2018, 7, 4554–4569. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Zhang, S.; Chen, J.; Zou, M.; Yang, Y.; Lu, W.; Ren, S.; Wang, X.; Dong, W.; Zhang, Z.; et al. The HDAC2/SP1/MiR-205 Feedback Loop Contributes to Tubular Epithelial Cell Extracellular Matrix Production in Diabetic Kidney Disease. Clin. Sci. 2022, 136, 223–238. [Google Scholar] [CrossRef]
- Xu, C.; Liu, C.-H.; Zhang, D.-L. MicroRNA-22 Inhibition Prevents Doxorubicin-Induced Cardiotoxicity via Upregulating SIRT1. Biochem. Biophys. Res. Commun. 2020, 521, 485–491. [Google Scholar] [CrossRef]
- Gondaliya, P.; Dasare, A.P.; Jash, K.; Tekade, R.K.; Srivastava, A.; Kalia, K. MiR-29b Attenuates Histone Deacetylase-4 Mediated Podocyte Dysfunction and Renal Fibrosis in Diabetic Nephropathy. J. Diabetes Metab. Disord. 2020, 19, 13–27. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, H.; Yin, M.; Ye, X.; Chen, G.; Zhou, X.; Yin, L.; Zhang, C.; Ding, B. MiR-376a and Histone Deacetylation 9 Form A Regulatory Circuitry in Hepatocellular Carcinoma. Cell. Physiol. Biochem. 2015, 35, 729–739. [Google Scholar] [CrossRef]
- Yang, B.; Zhao, H.; Dong, R. MiR-449 Improves Cardiac Function by Regulating HDAC1 and CTnI. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 12827–12835. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Chen, L.; Zhang, Z.; Meng, F.; Huang, G.; Sheng, P.; Zhang, Z.; Liao, W. MicroRNA-455-3p Modulates Cartilage Development and Degeneration through Modification of Histone H3 Acetylation. Biochim. Biophys. Acta BBA - Mol. Cell Res. 2016, 1863, 2881–2891. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, G.; Zhang, S.; Li, F.; Wang, R.; Tao, Z.; Ma, Q.; Han, Z.; Yan, F.; Fan, J.; et al. Inhibition of Histone Deacetylase 3 by MiR-494 Alleviates Neuronal Loss and Improves Neurological Recovery in Experimental Stroke. J. Cereb. Blood Flow Metab. 2019, 39, 2392–2405. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.; Zhao, W.; Xie, X.; Pan, Y.; Wang, K.; Zhang, H. MiR-520b Restrains Cell Growth by Targeting HDAC4 in Lung Cancer: MiR-520b Inhibits LC Growth via HDAC4. Thorac. Cancer 2018, 9, 1249–1254. [Google Scholar] [CrossRef] [PubMed]
- Gen, Y.; Muramatsu, T.; Inoue, J.; Inazawa, J. MiR-766-5p Targets Super-Enhancers by Downregulating CBP and BRD4. Cancer Res. 2021, 81, 5190–5201. [Google Scholar] [CrossRef]
- Mao, G.; Zhang, Z.; Huang, Z.; Chen, W.; Huang, G.; Meng, F.; Zhang, Z.; Kang, Y. MicroRNA-92a-3p Regulates the Expression of Cartilage-Specific Genes by Directly Targeting Histone Deacetylase 2 in Chondrogenesis and Degradation. Osteoarthritis Cartilage 2017, 25, 521–532. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Zhang, W.; Song, Z.; Wang, M.; Wu, H.; Yang, Y.; Chen, R. A Novel MiRNA Inhibits Metastasis of Prostate Cancer via Decreasing CREBBP-Mediated Histone Acetylation. J. Cancer Res. Clin. Oncol. 2021, 147, 469–480. [Google Scholar] [CrossRef]
- Hu, M.-J.; Long, M.; Dai, R.-J. Acetylation of H3K27 Activated LncRNA NEAT1 and Promoted Hepatic Lipid Accumulation in Non-Alcoholic Fatty Liver Disease via Regulating MiR-212-5p/GRIA3. Mol. Cell. Biochem. 2022, 477, 191–203. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, P.; Cui, Y. Long Noncoding RNA NEAT1 Inhibits the Acetylation of PTEN through the MiR-524-5p/HDAC1 Axis to Promote the Proliferation and Invasion of Laryngeal Cancer Cells. Aging 2021, 13, 24850–24865. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Y.; Xu, N.; Zhang, S.; Wang, S.; Mao, Y.; Zhu, Y.; Li, B.; Jiang, Y.; Tan, Y.; et al. NEAT1 Regulates Neuroglial Cell Mediating Aβ Clearance via the Epigenetic Regulation of Endocytosis-Related Genes Expression. Cell. Mol. Life Sci. 2019, 76, 3005–3018. [Google Scholar] [CrossRef] [Green Version]
- Guo, T.; Xing, Y.; Chen, Z.; Zhu, H.; Yang, L.; Xiao, Y.; Xu, J. Long Non-Coding RNA NEAT1 Knockdown Alleviates Rheumatoid Arthritis by Reducing IL-18 through P300/CBP Repression. Inflammation 2022, 45, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Q.; Jiang, D.-M.; Hu, S.-S.; Zhao, L.; Wang, L.; Yang, M.-H.; Ai, M.-L.; Jiang, H.-J.; Han, Y.; Ding, Y.-Q.; et al. SATB2-AS1 Suppresses Colorectal Carcinoma Aggressiveness by Inhibiting SATB2-Dependent Snail Transcription and Epithelial–Mesenchymal Transition. Cancer Res. 2019, 79, 3542–3556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Xu, H.-B.; Kurban, E.; Luo, H.-W. LncRNA SNHG14 Promotes Hepatocellular Carcinoma Progression via H3K27 Acetylation Activated PABPC1 by PTEN Signaling. Cell Death Dis. 2020, 11, 646. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Wang, W.; Mo, S.; Liu, Q.; Chen, X.; Chen, R.; Zhang, Y.; Zou, K.; Ye, M.; He, X.; et al. Long Non-Coding RNA SNHG14 Induces Trastuzumab Resistance of Breast Cancer via Regulating PABPC1 Expression through H3K27 Acetylation. J. Cell. Mol. Med. 2018, 22, 4935–4947. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.-Q.; Li, Z.-X.; Guan, J.-L.; Liu, X.; Li, J.-Y.; Chen, Y.; Lin, L.; Kou, J.; Lv, J.-W.; Zhang, L.-L.; et al. Long Noncoding RNA TINCR-Mediated Regulation of Acetyl-CoA Metabolism Promotes Nasopharyngeal Carcinoma Progression and Chemoresistance. Cancer Res. 2020, 80, 5174–5188. [Google Scholar] [CrossRef]
- Su, Q.; Liu, Y.; Lv, X.-W.; Dai, R.-X.; Yang, X.-H.; Kong, B.-H. LncRNA TUG1 Mediates Ischemic Myocardial Injury by Targeting MiR-132-3p/HDAC3 Axis. Am. J. Physiol.-Heart Circ. Physiol. 2020, 318, H332–H344. [Google Scholar] [CrossRef]
- Badal, S.S.; Wang, Y.; Long, J.; Corcoran, D.L.; Chang, B.H.; Truong, L.D.; Kanwar, Y.S.; Overbeek, P.A.; Danesh, F.R. MiR-93 Regulates Msk2-Mediated Chromatin Remodelling in Diabetic Nephropathy. Nat. Commun. 2016, 7, 12076. [Google Scholar] [CrossRef] [Green Version]
- Spolverini, A.; Fuchs, G.; Bublik, D.R.; Oren, M. Let-7b and Let-7c MicroRNAs Promote Histone H2B Ubiquitylation and Inhibit Cell Migration by Targeting Multiple Components of the H2B Deubiquitylation Machinery. Oncogene 2017, 36, 5819–5828. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wang, B.; Zheng, R.; Zhang, J.; Huang, C.; Zheng, R.; Huang, Z.; Qiu, W.; Liu, M.; Yang, K.; et al. Linc-RA1 Inhibits Autophagy and Promotes Radioresistance by Preventing H2Bub1/USP44 Combination in Glioma Cells. Cell Death Dis. 2020, 11, 758. [Google Scholar] [CrossRef]
- Xi, J.; Feng, J.; Li, Q.; Li, X.; Zeng, S. The Long Non-Coding RNA LncFOXO1 Suppresses Growth of Human Breast Cancer Cells through Association with BAP1. Int. J. Oncol. 2017, 50, 1663–1670. [Google Scholar] [CrossRef]
- Singh, R.; Chandel, S.; Dey, D.; Ghosh, A.; Roy, S.; Ravichandiran, V.; Ghosh, D. Epigenetic Modification and Therapeutic Targets of Diabetes Mellitus. Biosci. Rep. 2020, 40, BSR20202160. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Huang, Y.; Zhang, X.; Xu, Y.; Nie, S. Noncoding RNAs Involved in DNA Methylation and Histone Methylation, and Acetylation in Diabetic Vascular Complications. Pharmacol. Res. 2021, 170, 105520. [Google Scholar] [CrossRef] [PubMed]
- Fagan, R.J.; Dingwall, A.K. COMPASS Ascending: Emerging Clues Regarding the Roles of MLL3/KMT2C and MLL2/KMT2D Proteins in Cancer. Cancer Lett. 2019, 458, 56–65. [Google Scholar] [CrossRef]
- Sugeedha, J.; Gautam, J.; Tyagi, S. SET1/MLL Family of Proteins: Functions beyond Histone Methylation. Epigenetics 2021, 16, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, Z.; Ozark, P.A.; Fantini, D.; Marshall, S.A.; Rendleman, E.J.; Cozzolino, K.A.; Louis, N.; He, X.; Morgan, M.A.; et al. Resetting the Epigenetic Balance of Polycomb and COMPASS Function at Enhancers for Cancer Therapy. Nat. Med. 2018, 24, 758–769. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.A.; Kingston, R.E. Mechanisms of Polycomb Gene Silencing: Knowns and Unknowns. Nat. Rev. Mol. Cell Biol. 2009, 10, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-J.; Chun, T. Structural Heterogeneity of the Mammalian Polycomb Repressor Complex in Immune Regulation. Exp. Mol. Med. 2020, 52, 1004–1015. [Google Scholar] [CrossRef]
- Holoch, D.; Margueron, R. Mechanisms Regulating PRC2 Recruitment and Enzymatic Activity. Trends Biochem. Sci. 2017, 42, 531–542. [Google Scholar] [CrossRef]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Rivea Morales, D.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many Human Large Intergenic Noncoding RNAs Associate with Chromatin-Modifying Complexes and Affect Gene Expression. Proc. Natl. Acad. Sci. 2009, 106, 11667–11672. [Google Scholar] [CrossRef] [Green Version]
- Inoue, F.; Sone, K.; Toyohara, Y.; Takahashi, Y.; Kukita, A.; Hara, A.; Taguchi, A.; Tanikawa, M.; Tsuruga, T.; Osuga, Y. Targeting Epigenetic Regulators for Endometrial Cancer Therapy: Its Molecular Biology and Potential Clinical Applications. Int. J. Mol. Sci. 2021, 22, 2305. [Google Scholar] [CrossRef]
- Yang, Y.; Bedford, M.T. Protein Arginine Methyltransferases and Cancer. Nat. Rev. Cancer 2013, 13, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Congrains, A.; Kamide, K.; Ohishi, M.; Rakugi, H. ANRIL: Molecular Mechanisms and Implications in Human Health. Int. J. Mol. Sci. 2013, 14, 1278–1292. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Hsieh, C.-H.; Alonso, L.C. ANRIL: A LncRNA at the CDKN2A/B Locus With Roles in Cancer and Metabolic Disease. Front. Endocrinol. 2018, 9, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yap, K.L.; Li, S.; Muñoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.-M. Molecular Interplay of the Noncoding RNA ANRIL and Methylated Histone H3 Lysine 27 by Polycomb CBX7 in Transcriptional Silencing of INK4a. Mol. Cell 2010, 38, 662–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holdt, L.M.; Hoffmann, S.; Sass, K.; Langenberger, D.; Scholz, M.; Krohn, K.; Finstermeier, K.; Stahringer, A.; Wilfert, W.; Beutner, F.; et al. Alu Elements in ANRIL Non-Coding RNA at Chromosome 9p21 Modulate Atherogenic Cell Functions through Trans-Regulation of Gene Networks. PLoS Genet. 2013, 9, e1003588. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Sun, L.; Song, Y. FEZF1-AS1: A Novel Vital Oncogenic LncRNA in Multiple Human Malignancies. Biosci. Rep. 2019, 39, BSR20191202. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.; Zhou, Z.; Zhang, X. Effects of MiR-195-5p on Cell Proliferation and Apoptosis in Gestational Diabetes Mellitus via Targeting EZH2. Mol. Med. Rep. 2020, 22, 803–809. [Google Scholar] [CrossRef]
- Zheng, J.; Chen, L. Non-Coding RNAs-EZH2 Regulatory Mechanisms in Cervical Cancer: The Current State of Knowledge. Biomed. Pharmacother. 2022, 146, 112123. [Google Scholar] [CrossRef]
- Chang, L.; Zhou, R. Histone Methyltransferase EZH2 in Proliferation, Invasion, and Migration of Fibroblast-like Synoviocytes in Rheumatoid Arthritis. J. Bone Miner. Metab. 2022, 40, 262–274. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, F.; Liu, Y.; Zhao, Q.; Tang, H. DNMT1 Recruited by EZH2-Mediated Silencing of MiR-484 Contributes to the Malignancy of Cervical Cancer Cells through MMP14 and HNF1A. Clin. Epigenetics 2019, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, X.; Ding, X.; Huang, T.; Song, D.; Tao, H. EZH2 Is Associated with Cartilage Degeneration in Osteoarthritis by Promoting SDC1 Expression via Histone Methylation of the MicroRNA-138 Promoter. Lab. Invest. 2021, 101, 600–611. [Google Scholar] [CrossRef] [PubMed]
- Ou, R.; Zhu, L.; Zhao, L.; Li, W.; Tao, F.; Lu, Y.; He, Q.; Li, J.; Ren, Y.; Xu, Y. HPV16 E7-induced Upregulation of KDM2A Promotes Cervical Cancer Progression by Regulating MiR-132–Radixin Pathway. J. Cell. Physiol. 2019, 234, 2659–2671. [Google Scholar] [CrossRef] [PubMed]
- Min, Z.; Xiaomeng, L.; Zheng, L.; Yangge, D.; Xuejiao, L.; Longwei, L.; Xiao, Z.; Yunsong, L.; Ping, Z.; Yongsheng, Z. Asymmetrical Methyltransferase PRMT3 Regulates Human Mesenchymal Stem Cell Osteogenesis via MiR-3648. Cell Death Dis. 2019, 10, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Wang, L.; Xie, S.; Chen, Y.; Song, S.; Lu, Y.; Lu, D. Long Noncoding RNA MEG3 Blocks Telomerase Activity in Human Liver Cancer Stem Cells Epigenetically. Stem Cell Res. Ther. 2020, 11, 518. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Wang, J.; Man, W.-Y.; Zhang, Q.-W.; Xu, W.-G. SiRNA Silencing EZH2 Reverses Cisplatin-Resistance of Human Non-Small Cell Lung and Gastric Cancer Cells. Asian Pac. J. Cancer Prev. 2015, 16, 2425–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tornesello, M.L.; Faraonio, R.; Buonaguro, L.; Annunziata, C.; Starita, N.; Cerasuolo, A.; Pezzuto, F.; Tornesello, A.L.; Buonaguro, F.M. The Role of MicroRNAs, Long Non-Coding RNAs, and Circular RNAs in Cervical Cancer. Front. Oncol. 2020, 10, 150. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, L.; Ma, X. Circ_SPECC1 Enhances the Inhibition of MiR-526b on Downstream KDM4A/YAP1 Pathway to Regulate the Growth and Invasion of Gastric Cancer Cells. Biochem. Biophys. Res. Commun. 2019, 517, 253–259. [Google Scholar] [CrossRef]
- Wakisaka, K.T. The Dawn of Pirna Research in Various Neuronal Disorders. Front. Biosci. 2019, 24, 1440–1451. [Google Scholar] [CrossRef]
- Pezic, D.; Manakov, S.A.; Sachidanandam, R.; Aravin, A.A. PiRNA Pathway Targets Active LINE1 Elements to Establish the Repressive H3K9me3 Mark in Germ Cells. Genes Dev. 2014, 28, 1410–1428. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.-J.; Man, N.; Tan, Y.; Nimer, S.D.; Wang, L. The Role of Histone Acetyltransferases in Normal and Malignant Hematopoiesis. Front. Oncol. 2015, 5, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, G.; Hernández-Illán, E.; Moreira, L.; Balaguer, F.; Goel, A. Epigenetics of Colorectal Cancer: Biomarker and Therapeutic Potential. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-R.; Wang, J.-Q.; Huang, Z.-G.; Chen, R.-N.; Cao, X.; Zhu, D.-C.; Yu, H.-X.; Wang, X.-R.; Zhou, H.-Y.; Xia, Q.; et al. Histone Deacetylase-2: A Potential Regulator and Therapeutic Target in Liver Disease (Review). Int. J. Mol. Med. 2021, 48, 131. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.Y.; Denu, J.M.; Allis, C.D. Histone Acetyltransferases. Annu. Rev. Biochem. 2001, 70, 81–120. [Google Scholar] [CrossRef]
- Attar, N.; Kurdistani, S.K. Exploitation of EP300 and CREBBP Lysine Acetyltransferases by Cancer. Cold Spring Harb. Perspect. Med. 2017, 7, a026534. [Google Scholar] [CrossRef] [Green Version]
- Morgado-Pascual, J.L.; Rayego-Mateos, S.; Tejedor, L.; Suarez-Alvarez, B.; Ruiz-Ortega, M. Bromodomain and Extraterminal Proteins as Novel Epigenetic Targets for Renal Diseases. Front. Pharmacol. 2019, 10, 1315. [Google Scholar] [CrossRef]
- Garcia-Carpizo, V.; Ruiz-Llorente, S.; Sarmentero, J.; Graña-Castro, O.; Pisano, D.G.; Barrero, M.J. CREBBP/EP300 Bromodomains Are Critical to Sustain the GATA1/MYC Regulatory Axis in Proliferation. Epigenetics Chromatin 2018, 11, 30. [Google Scholar] [CrossRef] [Green Version]
- Gujral, P.; Mahajan, V.; Lissaman, A.C.; Ponnampalam, A.P. Histone Acetylation and the Role of Histone Deacetylases in Normal Cyclic Endometrium. Reprod. Biol. Endocrinol. 2020, 18, 84. [Google Scholar] [CrossRef]
- Hadley, M.; Noonepalle, S.; Banik, D.; Villagra, A. Functional Analysis of HDACs in Tumorigenesis. In Protein Acetylation; Brosh, R.M., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; Volume 1983, pp. 279–307. ISBN 978-1-4939-9433-5. [Google Scholar]
- Reichert, N.; Choukrallah, M.-A.; Matthias, P. Multiple Roles of Class I HDACs in Proliferation, Differentiation, and Development. Cell. Mol. Life Sci. 2012, 69, 2173–2187. [Google Scholar] [CrossRef] [Green Version]
- Claveria-Cabello, A.; Colyn, L.; Arechederra, M.; Urman, J.M.; Berasain, C.; Avila, M.A.; Fernandez-Barrena, M.G. Epigenetics in Liver Fibrosis: Could HDACs Be a Therapeutic Target? Cells 2020, 9, 2321. [Google Scholar] [CrossRef]
- Menzies, K.J.; Zhang, H.; Katsyuba, E.; Auwerx, J. Protein Acetylation in Metabolism — Metabolites and Cofactors. Nat. Rev. Endocrinol. 2016, 12, 43–60. [Google Scholar] [CrossRef] [PubMed]
- Etchegaray, J.-P.; Chavez, L.; Huang, Y.; Ross, K.N.; Choi, J.; Martinez-Pastor, B.; Walsh, R.M.; Sommer, C.A.; Lienhard, M.; Gladden, A.; et al. The Histone Deacetylase SIRT6 Controls Embryonic Stem Cell Fate via TET-Mediated Production of 5-Hydroxymethylcytosine. Nat. Cell Biol. 2015, 17, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Zhao, L. Long Non-Coding RNA Histone Deacetylase 4 Antisense RNA 1 (HDAC4-AS1) Inhibits HDAC4 Expression in Human ARPE-19 Cells with Hypoxic Stress. Bioengineered 2021, 12, 2228–2237. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Lv, D.; Li, M.; Zhang, X.; Sun, G.; Bai, Y.; Chang, D. Hypermethylation of MiRNA-589 Promoter Leads to Upregulation of HDAC5 Which Promotes Malignancy in Non-Small Cell Lung Cancer. Int. J. Oncol. 2017, 50, 2079–2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, L.; Hou, C. MiR-138 Activates NF-ΚB Signaling and PGRN to Promote Rheumatoid Arthritis via Regulating HDAC4. Biochem. Biophys. Res. Commun. 2019, 519, 166–171. [Google Scholar] [CrossRef]
- Fu, Q.; Pan, H.; Tang, Y.; Rong, J.; Zheng, Z. MiR-200a-3p Aggravates DOX-Induced Cardiotoxicity by Targeting PEG3 Through SIRT1/NF-ΚB Signal Pathway. Cardiovasc. Toxicol. 2021, 21, 302–313. [Google Scholar] [CrossRef]
- Li, D.; Yang, Y.; Wang, S.; He, X.; Liu, M.; Bai, B.; Tian, C.; Sun, R.; Yu, T.; Chu, X. Role of Acetylation in Doxorubicin-Induced Cardiotoxicity. Redox Biol. 2021, 46, 102089. [Google Scholar] [CrossRef]
- Lau, P.N.I.; Cheung, P. Histone Code Pathway Involving H3 S28 Phosphorylation and K27 Acetylation Activates Transcription and Antagonizes Polycomb Silencing. Proc. Natl. Acad. Sci. 2011, 108, 2801–2806. [Google Scholar] [CrossRef] [Green Version]
- Hayashi-Takanaka, Y.; Yamagata, K.; Nozaki, N.; Kimura, H. Visualizing Histone Modifications in Living Cells: Spatiotemporal Dynamics of H3 Phosphorylation during Interphase. J. Cell Biol. 2009, 187, 781–790. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.C.L.; Goyal, P.; Karimi, M.M.; Abildgaard, M.H.; Kimura, H.; Lorincz, M.C. H3S10ph Broadly Marks Early-Replicating Domains in Interphase ESCs and Shows Reciprocal Antagonism with H3K9me2. Genome Res. 2018, 28, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Albig, C.; Wang, C.; Dann, G.P.; Wojcik, F.; Schauer, T.; Krause, S.; Maenner, S.; Cai, W.; Li, Y.; Girton, J.; et al. JASPer Controls Interphase Histone H3S10 Phosphorylation by Chromosomal Kinase JIL-1 in Drosophila. Nat. Commun. 2019, 10, 5343. [Google Scholar] [CrossRef] [PubMed]
- García-Pichardo, D.; Cañas, J.C.; García-Rubio, M.L.; Gómez-González, B.; Rondón, A.G.; Aguilera, A. Histone Mutants Separate R Loop Formation from Genome Instability Induction. Mol. Cell 2017, 66, 597–609.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simboeck, E.; Sawicka, A.; Zupkovitz, G.; Senese, S.; Winter, S.; Dequiedt, F.; Ogris, E.; Di Croce, L.; Chiocca, S.; Seiser, C. A Phosphorylation Switch Regulates the Transcriptional Activation of Cell Cycle Regulator P21 by Histone Deacetylase Inhibitors. J. Biol. Chem. 2010, 285, 41062–41073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komar, D.; Juszczynski, P. Rebelled Epigenome: Histone H3S10 Phosphorylation and H3S10 Kinases in Cancer Biology and Therapy. Clin. Epigenetics 2020, 12, 147. [Google Scholar] [CrossRef]
- Wiersma, M.; Bussiere, M.; Halsall, J.A.; Turan, N.; Slany, R.; Turner, B.M.; Nightingale, K.P. Protein Kinase Msk1 Physically and Functionally Interacts with the KMT2A/MLL1 Methyltransferase Complex and Contributes to the Regulation of Multiple Target Genes. Epigenetics Chromatin 2016, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, R.A.; Dennis, E.A. Handbook of Cell Signaling, 2nd ed.; Academic Press: Cambridge, MA, USA, 2010; ISBN 978-0-12-374145-5. [Google Scholar]
- Kim, M.; Yi, S.A.; Lee, H.; Bang, S.Y.; Park, E.K.; Lee, M.G.; Nam, K.H.; Yoo, J.H.; Lee, D.H.; Ryu, H.-W.; et al. Reversine Induces Multipotency of Lineage-Committed Cells through Epigenetic Silencing of MiR-133a. Biochem. Biophys. Res. Commun. 2014, 445, 255–262. [Google Scholar] [CrossRef]
- Salas-Lloret, D.; González-Prieto, R. Insights in Post-Translational Modifications: Ubiquitin and SUMO. Int. J. Mol. Sci. 2022, 23, 3281. [Google Scholar] [CrossRef]
- Zhang, Y. Transcriptional Regulation by Histone Ubiquitination and Deubiquitination. Genes Dev. 2003, 17, 2733–2740. [Google Scholar] [CrossRef] [Green Version]
- Kao, C.-F.; Hillyer, C.; Tsukuda, T.; Henry, K.; Berger, S.; Osley, M.A. Rad6 Plays a Role in Transcriptional Activation through Ubiquitylation of Histone H2B. Genes Dev. 2004, 18, 184–195. [Google Scholar] [CrossRef] [Green Version]
- Passmore, L.A.; Barford, D. Getting into Position: The Catalytic Mechanisms of Protein Ubiquitylation. Biochem. J. 2004, 379, 513–525. [Google Scholar] [CrossRef]
- Komander, D. The Emerging Complexity of Protein Ubiquitination. Biochem. Soc. Trans. 2009, 37, 937–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, D.E. Histone Ubiquitylation and Chromatin Dynamics. Front. Biosci. 2012, 17, 1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osley, M.A.; Fleming, A.B.; Kao, C.-F. Histone Ubiquitylation and the Regulation of Transcription. In Chromatin Dynamics in Cellular Function; Laurent, B.C., Ed.; Results and Problems in Cell Differentiation; Springer: Berlin/Heidelberg, Germany, 2006; Volume 41, pp. 47–75. ISBN 978-3-540-33685-3. [Google Scholar]
- Bonnet, J.; Romier, C.; Tora, L.; Devys, D. Zinc-Finger UBPs: Regulators of Deubiquitylation. Trends Biochem. Sci. 2008, 33, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Boyerinas, B.; Park, S.-M.; Hau, A.; Murmann, A.E.; Peter, M.E. The Role of Let-7 in Cell Differentiation and Cancer. Endocr. Relat. Cancer 2010, 17, F19–F36. [Google Scholar] [CrossRef]
- Chen, M.-K.; Zhou, J.-H.; Wang, P.; Ye, Y.; Liu, Y.; Zhou, J.-W.; Chen, Z.-J.; Yang, J.-K.; Liao, D.-Y.; Liang, Z.-J.; et al. BMI1 Activates P-Glycoprotein via Transcription Repression of MiR-3682-3p and Enhances Chemoresistance of Bladder Cancer Cell. Aging 2021, 13, 18310–18330. [Google Scholar] [CrossRef]
- Wang, Y.; Dasso, M. SUMOylation and DeSUMOylation at a Glance. J. Cell Sci. 2009, 122, 4249–4252. [Google Scholar] [CrossRef] [Green Version]
- Sarangi, P.; Zhao, X. SUMO-Mediated Regulation of DNA Damage Repair and Responses. Trends Biochem. Sci. 2015, 40, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Yau, T.-Y.; Molina, O.; Courey, A.J. SUMOylation in Development and Neurodegeneration. Development 2020, 147, dev175703. [Google Scholar] [CrossRef]
- Zheng, B.; Bernier, M.; Zhang, X.; Suzuki, T.; Nie, C.; Li, Y.H.; Zhang, Y.; Song, L.; Shi, H.; Liu, Y.; et al. MiR-200c-SUMOylated KLF4 Feedback Loop Acts as a Switch in Transcriptional Programs That Control VSMC Proliferation. J. Mol. Cell. Cardiol. 2015, 82, 201–212. [Google Scholar] [CrossRef]
- Kothapalli, N.; Camporeale, G.; Kueh, A.; Chew, Y.C.; Oommen, A.M.; Griffin, J.B.; Zempleni, J. Biological Functions of Biotinylated Histones. J. Nutr. Biochem. 2005, 16, 446–448. [Google Scholar] [CrossRef] [Green Version]
- Kobza, K.; Camporeale, G.; Rueckert, B.; Kueh, A.; Griffin, J.B.; Sarath, G.; Zempleni, J. K4, K9 and K18 in Human Histone H3 Are Targets for Biotinylation by Biotinidase: Biotinylation Sites in Human Histone H3. FEBS J. 2005, 272, 4249–4259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballard, T.D.; Wolff, J.; Griffin, J.B.; Stanley, J.S.; van Calcar, S.; Zempleni, J. Biotinidase Catalyzes Debiotinylation of Histones. Eur. J. Nutr. 2002, 41, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Scheerger, S.B.; Zempleni, J. Expression of Oncogenes Depends on Biotin in Human Small Cell Lung Cancer Cells NCI-H69. Int. J. Vitam. Nutr. Res. 2003, 73, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Crisp, S.E.R.H.; Griffin, J.B.; White, B.R.; Toombs, C.F.; Camporeale, G.; Said, H.M.; Zempleni, J. Biotin Supply Affects Rates of Cell Proliferation, Biotinylation of Carboxylases and Histones, and Expression of the Gene Encoding the Sodium-Dependent Multivitamin Transporter in JAr Choriocarcinoma Cells. Eur. J. Nutr. 2004, 43, 23–31. [Google Scholar] [CrossRef]
- Filenko, N.A.; Kolar, C.; West, J.T.; Smith, S.A.; Hassan, Y.I.; Borgstahl, G.E.O.; Zempleni, J.; Lyubchenko, Y.L. The Role of Histone H4 Biotinylation in the Structure of Nucleosomes. PloS One 2011, 6, e16299. [Google Scholar] [CrossRef] [Green Version]
- Camporeale, G.; Oommen, A.M.; Griffin, J.B.; Sarath, G.; Zempleni, J. K12-Biotinylated Histone H4 Marks Heterochromatin in Human Lymphoblastoma Cells. J. Nutr. Biochem. 2007, 18, 760–768. [Google Scholar] [CrossRef] [Green Version]
- Bao, B.; Rodriguez-Melendez, R.; Zempleni, J. Cytosine Methylation in MiR-153 Gene Promoters Increases the Expression of Holocarboxylase Synthetase, Thereby Increasing the Abundance of Histone H4 Biotinylation Marks in HEK-293 Human Kidney Cells. J. Nutr. Biochem. 2012, 23, 635–639. [Google Scholar] [CrossRef] [Green Version]
- Garofoli, M.; Volpicella, M.; Guida, M.; Porcelli, L.; Azzariti, A. The Role of Non-Coding RNAs as Prognostic Factor, Predictor of Drug Response or Resistance and Pharmacological Targets, in the Cutaneous Squamous Cell Carcinoma. Cancers 2020, 12, 2552. [Google Scholar] [CrossRef]
- De Palma, F.D.E.; Salvatore, F.; Pol, J.G.; Kroemer, G.; Maiuri, M.C. Circular RNAs as Potential Biomarkers in Breast Cancer. Biomedicines 2022, 10, 725. [Google Scholar] [CrossRef]
- Yuan, H.-L.; Wang, T.; Zhang, K.-H. MicroRNAs as Potential Biomarkers for Diagnosis, Therapy and Prognosis of Gastric Cancer. OncoTargets Ther. 2018, 11, 3891–3900. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.; Yi, J.; Zhu, X.; Zhang, J.; Zhou, J.; Tang, X.; Lin, J.; Wang, P.; Deng, Z. Circular RNA of Vimentin Expression as a Valuable Predictor for Acute Myeloid Leukemia Development and Prognosis. J. Cell. Physiol. 2019, 234, 3711–3719. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Qian, X.; Wang, Z.; Zhou, X. The HOTAIR LncRNA: A Remarkable Oncogenic Promoter in Human Cancer Metastasis (Review). Oncol. Lett. 2021, 21, 302. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, R.; Gupta, S. Epigenetic Modifications in Cancer. Clin. Genet. 2012, 81, 303–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Yang, Y.; Wang, Y. Predictive Biomarkers and Potential Drug Combinations of Epi-Drugs in Cancer Therapy. Clin. Epigenetics 2021, 13, 113. [Google Scholar] [CrossRef]
- Egger, G.; Liang, G.; Aparicio, A.; Jones, P.A. Epigenetics in Human Disease and Prospects for Epigenetic Therapy. Nature 2004, 429, 457–463. [Google Scholar] [CrossRef]
- Garmpis, N.; Damaskos, C.; Garmpi, A.; Spartalis, E.; Kalampokas, E.; Kalampokas, T.; Margonis, G.-A.; Schizas, D.; Andreatos, N.; Angelou, A.; et al. Targeting Histone Deacetylases in Endometrial Cancer: A Paradigm-Shifting Therapeutic Strategy? Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 950–960. [Google Scholar] [CrossRef]
- Jin, N.; George, T.L.; Otterson, G.A.; Verschraegen, C.; Wen, H.; Carbone, D.; Herman, J.; Bertino, E.M.; He, K. Advances in Epigenetic Therapeutics with Focus on Solid Tumors. Clin. Epigenetics 2021, 13, 83. [Google Scholar] [CrossRef]
- Singh, B.N.; Zhou, H.; Li, J.; Tipton, T.; Wang, B.; Shao, G.; Gilbert, E.N.; Li, Q.; Jiang, S.-W. Preclinical Studies on Histone Deacetylase Inhibitors as Therapeutic Reagents for Endometrial and Ovarian Cancers. Future Oncol. 2011, 7, 1415–1428. [Google Scholar] [CrossRef] [Green Version]
- Mottamal, M.; Zheng, S.; Huang, T.; Wang, G. Histone Deacetylase Inhibitors in Clinical Studies as Templates for New Anticancer Agents. Molecules 2015, 20, 3898–3941. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-L.; Liu, H.-Q.; Li, J.; Mao, C.-Y.; He, J.-T.; Zhao, X. Role of Epigenetic in Leukemia: From Mechanism to Therapy. Chem. Biol. Interact. 2020, 317, 108963. [Google Scholar] [CrossRef]
- Whittaker, S.J.; Demierre, M.-F.; Kim, E.J.; Rook, A.H.; Lerner, A.; Duvic, M.; Scarisbrick, J.; Reddy, S.; Robak, T.; Becker, J.C.; et al. Final Results From a Multicenter, International, Pivotal Study of Romidepsin in Refractory Cutaneous T-Cell Lymphoma. J. Clin. Oncol. 2010, 28, 4485–4491. [Google Scholar] [CrossRef] [PubMed]
- Raedler, L.A. Farydak (Panobinostat): First HDAC Inhibitor Approved for Patients with Relapsed Multiple Myeloma. Am. Health Drug Benefits 2016, 9, 84–87. [Google Scholar] [PubMed]
- Brocks, D.; Schmidt, C.R.; Daskalakis, M.; Jang, H.S.; Shah, N.M.; Li, D.; Li, J.; Zhang, B.; Hou, Y.; Laudato, S.; et al. DNMT and HDAC Inhibitors Induce Cryptic Transcription Start Sites Encoded in Long Terminal Repeats. Nat. Genet. 2017, 49, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
- Morera, L.; Lübbert, M.; Jung, M. Targeting Histone Methyltransferases and Demethylases in Clinical Trials for Cancer Therapy. Clin. Epigenetics 2016, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhu, D.; Zhang, Z.; Kaluz, S.; Yu, B.; Devi, N.S.; Olson, J.J.; Van Meir, E.G. Correction: EZH2 Targeting Reduces Medulloblastoma Growth through Epigenetic Reactivation of the BAI1/P53 Tumor Suppressor Pathway. Oncogene 2020, 39, 1165. [Google Scholar] [CrossRef]
- Shamsian, A.; Sepand, M.R.; Javaheri Kachousangi, M.; Dara, T.; Ostad, S.N.; Atyabi, F.; Ghahremani, M.H. Targeting Tumorigenicity of Breast Cancer Stem Cells Using SAHA/Wnt-b Catenin Antagonist Loaded Onto Protein Corona of Gold Nanoparticles. Int. J. Nanomedicine 2020, 15, 4063–4078. [Google Scholar] [CrossRef]
- Marjanovic, N.D.; Hofree, M.; Chan, J.E.; Canner, D.; Wu, K.; Trakala, M.; Hartmann, G.G.; Smith, O.C.; Kim, J.Y.; Evans, K.V.; et al. Emergence of a High-Plasticity Cell State during Lung Cancer Evolution. Cancer Cell 2020, 38, 229–246.e13. [Google Scholar] [CrossRef]
- Kurreck, J. Design of Antisense Oligonucleotides Stabilized by Locked Nucleic Acids. Nucleic Acids Res. 2002, 30, 1911–1918. [Google Scholar] [CrossRef] [Green Version]
- Titze-de-Almeida, R.; David, C.; Titze-de-Almeida, S.S. The Race of 10 Synthetic RNAi-Based Drugs to the Pharmaceutical Market. Pharm. Res. 2017, 34, 1339–1363. [Google Scholar] [CrossRef]
- Watts, J.K.; Corey, D.R. Silencing Disease Genes in the Laboratory and the Clinic. J. Pathol. 2012, 226, 365–379. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Shen, J.; Xiao, Z.; Li, J.; Zhao, Y.; Zhao, Q.; Cho, C.H.; Li, M. An Overview of the Multifaceted Roles of MiRNAs in Gastric Cancer: Spotlight on Novel Biomarkers and Therapeutic Targets. Biochem. Pharmacol. 2019, 163, 425–439. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Thakral, D.; Gupta, R. Regulatory Noncoding RNAs: Potential Biomarkers and Therapeutic Targets in Acute Myeloid Leukemia. Am. J. Blood Res. 2021, 11, 504–519. [Google Scholar] [PubMed]
- Kristen, A.V.; Ajroud-Driss, S.; Conceição, I.; Gorevic, P.; Kyriakides, T.; Obici, L. Patisiran, an RNAi Therapeutic for the Treatment of Hereditary Transthyretin-Mediated Amyloidosis. Neurodegener. Dis. Manag. 2019, 9, 5–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
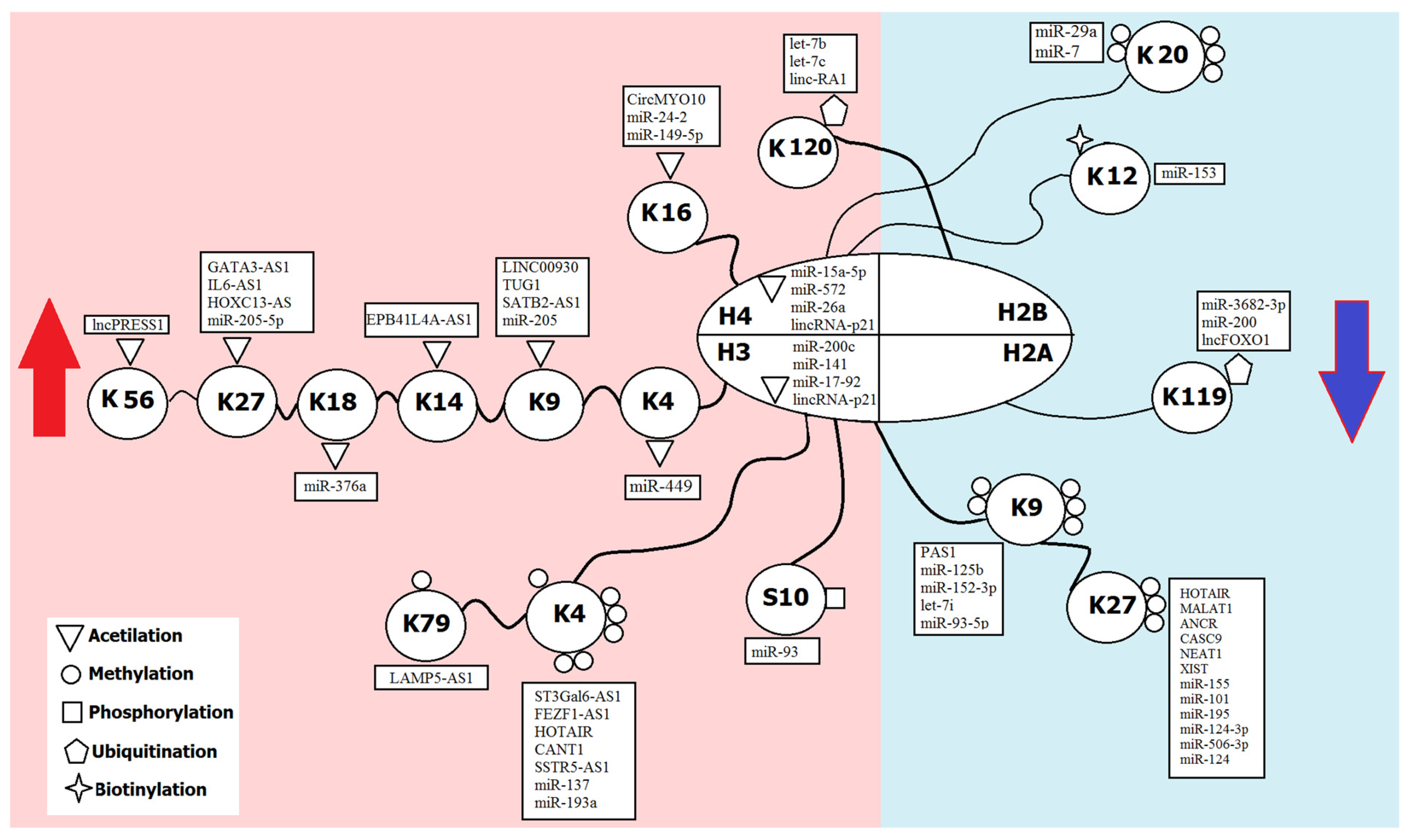
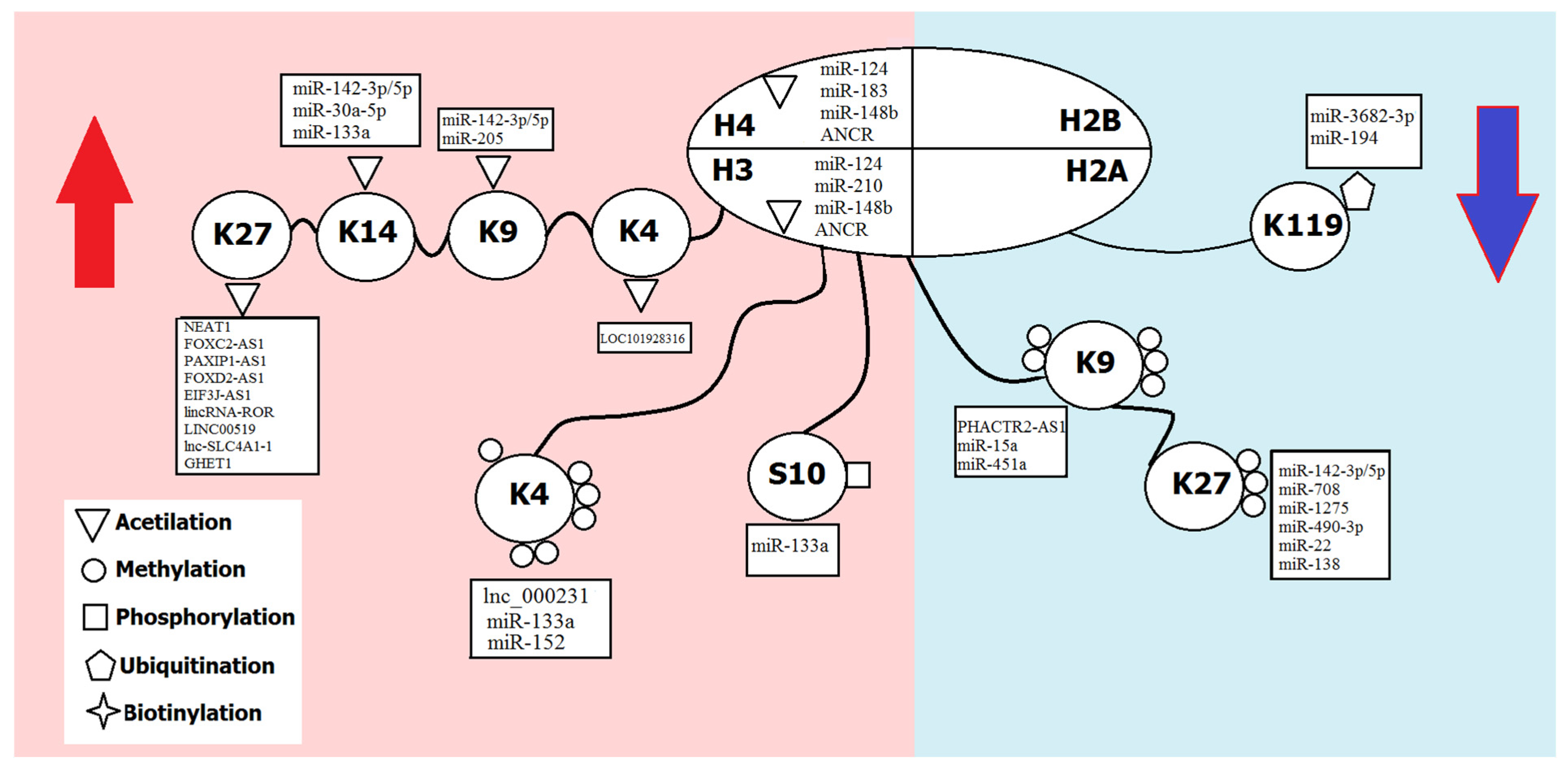
| Modification | Group | Effect on Transcription | Sites of Modification |
|---|---|---|---|
| 1. Modifications by small chemical groups | |||
| Methylation | Methyl group | Activation | H3 (K4, K36, K79) |
| Repression | H3 (K9, K27), H4K20 | ||
| Acetylation | Acetyl group | Activation | H3 (K4, K9, K14, K18, K27, K56); H4 (K5, K8, K12, K16); H2A/H2B (K6, K7, K16, K17) |
| Phosphorylation | Phosphate | Activation | H3 (S10) |
| 2. Modifications by complex molecules | |||
| Ubiquitination | Ubiquitin | Activation | H2B (K120) |
| Repression | H2A (K119) | ||
| Sumoylation | SUMO | Repression | H4 (K5, K8, K12, K16), H2A (K126), H2B (K6, K7, K16, K17) |
| Biotinylation | Biotin | Repression | H2A (K9, K13, K129), H3 (K4, K9, K18), H4 (K8, K12) |
| NcRNA | Target Modification | Epigenetic Regulatory Mechanisms | Role in Pathogenesis | Ref. |
|---|---|---|---|---|
| Methylation/Demethylation | ||||
| ANRIL | H3K27me3 | Directly interacts with PRC2 (EZH2 and SUZ12) | Promotes cholangiocarcinoma progression | [47] |
| CANT1 | H3K4me3 | Prevents hSET1 from binding to the PI3Kγ promoter | Suppresses retinoblastoma progression | [48] |
| Chaer | H3K27me3 | Interacts with PRC2 | Promotes cell proliferation and induces apoptosis in atherosclerosis | [49] |
| circ_0019435 | H3K27me3 | Directly binds EZH2 and silences the expression of DKK1 and PTEN | Promotes proliferation, invasion and EMT in cervical cancer | [50] |
| circAGFG1 | H3K27me3 | Recruits EZH2 to the promoter of p53 and inhibits its expression | Regulates proliferation and apoptosis in cervical cancer | [51] |
| DLEU1 | H3K4me3 | Activates JAK-STAT signaling pathway | Promotes oral squamous cell carcinoma progression | [52] |
| FEZF1-AS1 | H3K4me, H3K4me2 | Specifically binds LSD1, regulating the expression of CDKN1A (P21) | Involved in pathogenesis of colorectal carcinoma and glioma; promotes gastric cancer proliferation | [53,54,55] |
| Firre | H3K27me3 | Interacts with SUZ12 | Maintains X chromosome inactivation | [56] |
| GAS8-AS1 | H3K4me3 | Recruits the MLL1/WDR5 complex and maintains the GAS8 promoter in an open chromatin state | Inhibited malignant transformation of hepatocytes | [57] |
| H19 | H3K27me3, H3K4me3 | Facilitates the PRC2 complex in regulating methylation changes at H3K27me3/H3K4me3 histone sites of genes; represses VASH1 through EZH2-dependent H3K27 trimethylation | Regulates tumor plasticity in neuroendocrine prostate cancer; important regulator in human amniotic mesenchymal stem cells for promoting angiogenesis | [58,59] |
| HOTAIR | H3K27me3 | Recruits PRC2 and mediates H3K27me3 in different genes | Promotes proliferation, invasion, migration in tumors; promotes EMT in gastric cancer; promotes the self-renewal of leukemia stem cells; promotes metastasis of renal cell carcinoma | [60,61,62,63] |
| HOXA-AS3 | H3K27me3 | Facilitates EZH2-mediated H3K27me3, target RUNX2 | Inhibits osteogenic differentiation of mesenchymal stromal cells, regulates degenerative bone diseases | [45] |
| IL6-AS1 | H3K4me | Promotes IL-6 transcription by recruiting early B-cell factor 1 to the IL-6 promoter | Associated with airway inflammation | [64] |
| LAMP5-AS1 | H3K79me2, H3K79me3 | Directly binds DOT1L, promoting the global patterns of H3K79me2/me3 in cells | Regulates the self-renewal program and differentiation block in MLL leukemia | [65] |
| let-7i | H3K9me2 | Targets KDM3A, thus removing the DCLK1 H3K9me2, and suppresses the FXYD3 | Lung cancer progression | [66] |
| LINC01419 | H3K27me3 | Binds to EZH2, leading to histone methylation of the RECK promoter | Hepatocellular carcinoma growth and metastasis | [67] |
| LINP1 | H3K27me3 | Recruits EZH2 to the promoter regions of tumor suppressors KLF2 and PRSS8 | Regulates proliferation and apoptosis in cervical cancer | [68] |
| lnc-ATB | H3K27me3 | Directly interacts with EZH2 | Regulates proliferation, invasion and migration; promotes ovarian cancer | [69] |
| lnc-OB1 | H3K27me3 | Upregulates OSX via the inhibition of H3K27me3 in the OSX promoter; interacts with SUZ12 | Promotes osteogenic differentiation in human osteoblasts, might be a drug target for the treatment of osteoporosis | [70] |
| lncRNA CIR | H3K27me3 | Binds to EZH2 and repressed ATOH8 expression via EZH2-mediated H3K27me3, which promotes the methylation of ATOH8 | Inhibits chondrogenic differentiation | [71] |
| lncRNA ROR | H3K4me3 | Recruits histone transmethylase MLL1 to upregulate TIMP3 expression | Promotes breast cancer progression | [72] |
| lncRNA-p21 | H3K27me3 | Switches the EZH2 function from histone methyltransferase to non-histone methyltransferase, consequently methylating the STAT3 | Promotes the neuroendocrine differentiation in antiandrogen enzalutamide-induced prostate cancer | [73] |
| MALAT1 | H3K27me3 | Recruits EZH2 to promoters of target genes, facilitating H3K27me3 | Potentiates growth and inhibits senescence in gallbladder cancer cells; indicates poor prognosis; releases epigenetic silencing of HIV-1 replication; regulator of inflammation in diabetic retinopathy | [74,75,76] |
| MEG3 | H3K27me3 | Recruits EZH2; increases H3K27me3 via P53; inhibits the expression of TERT by increasing the H3K27me3 in TERT promoter regions and inhibition of the activity of telomerase by reducing the binding of TERT to TERC; increases the expression of TERRA | Reduces the activity of telomerase and blocks telomere | [77,78,79] |
| miR-101 | H4K27me3, H3K9me3, H4K20me3 | Directly targets EZH2 | Regulates autophagy, proliferation and apoptosis in laryngeal squamous cell carcinoma; affects endothelial function and angiogenesis in diabetes mellitus | [80,81] |
| miR-124 | H3K27me3 | Directly targets EZH2 | Promotes tumor growth and is associated with poor prognosis in esophageal squamous cell carcinoma; involved in thyroid carcinoma pathogenesis, reduced in sorafenib insensitive patients | [82,83] |
| miR-125b | H3K9me3 | Targets Suv39h1 | Sustains inflammation in diabetes | [84] |
| miR-137 | H3K4me2, H3K4me3 | Directly targets EZH2, JARID1B, KDM5B | Correlated with the poor prognosis and short-term survival of patients with cervical cancer; inhibits cell proliferation in acute lymphoblastic leukemia; involved in PTEN-Null prostate cancer | [85,86] |
| miR-152-3p | H3K9me3 | Targets SETDB1 | Positively regulates the production of H3K9me3 | [87] |
| miR-155 | H3K27me3 | Crosstalk between miR-155-PRC2 complex-JARID2 and PHF19 | Involved in endometriosis | [88] |
| miR-193a | H3K4me, H3K4me2 and H3K4me3 | Directly targets the MLL1 | Regulates cell growth, migration and apoptosis | [89] |
| miR-195–5p | H3K27me3 | Inhibits EZH2 | Promotes tubular injury in diabetic nephropathy | [90] |
| miR-199b-3p | H3K27me3 | Increases E-cadherin expression and prevents EMT through repressing KDM6A expression | Protects renal tubules from diabetic-induced injury | [91] |
| miR-214 | H3K27me3 | Directly targets EZH2 | Low expression correlates with advanced stage and poor overall survival of patients with cervical cancer | [92] |
| miR-216b | H3K9me2, H3K9me3 | Directly targets JMJD2C, downregulation of the JMJD2C/HIF1α/HES1 signaling axis | Positively correlated with patient survival in osteosarcoma, enhanced cisplatin-induced apoptosis | [93] |
| miR24-2 | H4R3me2, H4R3me3 | Targets PRMT7 | Associated with human tumorigenesis | [94] |
| miR-29 | H4K20me3 | Directly suppress Suv4-20h | Contributes to cellular senescence and cardiac aging | [95] |
| miR-29a | H4K20me3 | Downregulates histone H4K20 trimethylation through directly targeting SUV420H2 | Involved in breast cancer cells epithelial–mesenchymal transition, migration and invasion | [96] |
| miR-506-3p | H3K27me3 | Directly targets EZH2 | Involved in thyroid carcinoma pathogenesis, reduced in sorafenib insensitive patients | [83] |
| miR-7 | H4K20me1 | Negatively regulates SET8 | Suppresses the invasive potential of breast cancer cells and sensitizes cells to DNA damage | [97] |
| miR-93-5p | H3K9me3 | Directly suppresses Bcl-w | Inhibits premature cellular senescence | [98] |
| NEAT1 | H3K9me2 | Transcriptional repression of the c-Fos by H3K9me2 at promoter; binds to EZH2 | Mediates age-related memory impairment; contributes to glioblastoma progression | [99,100] |
| PAN RNA | H3K27me3 | Interacts with UTX and JMJD3, removes the repressive marks on the chromatin | Involved in Kaposi’s sarcoma-associated herpesvirus-mediated malignancies | [101] |
| PART1 | H3K27me3 | Epigenetic silencing of PDGFB via the PLZF-mediated recruitment of EZH2 | Restrains aggressive gastric cancer; low expression is associated with postoperative metastasis and short overall survival | [102] |
| PAS1 | H3K9 | Recruits SUV39H1 to methylate H3K9 of PH20 | Inhibits breast cancer growth and metastasis | [103] |
| PHACTR2-AS1 | H3K9me2, H3K9me3 | Directly binds ribosome DNA genes and recruits SUV39H1 | Promotes growth and metastasis in breast cancer | [104] |
| PVT1 | H3K27me3 | Forms a complex with EZH2, directly binding the promoter region of miR-195 | Related to large tumor size, advanced stage and poor prognosis in cervical cancer; regulates the chemoresistance | [105] |
| SNHG1 | H3K27me3 | Interacts with EZH2 and acts as a sponge for miR-154-5p and miR-143-3p | Oncogenic functions in colorectal and bladder cancer | [106,107] |
| SNHG22 | H3K27me3 | Binds EZH2 and regulates miR-200c-3p/Notch1 axis | Promotes proliferation and invasion, poor prognosis in gastric cancer | [108] |
| SNHG7 | H3K27me3 | Recruits EZH2 to promoter of the inhibitor of the Wnt/β-catenin signaling DKK | Promotes cervical cancer | [109] |
| SNHG8 | H3K27me3 | Directly interacts with EZH2, inhibiting the expression of RECK at the transcriptional level | Promotes cervical cancer | [110] |
| SSTR5-AS1 | H3K4me3 | Interacts with MLL3 and increases the enrichment of MLL3 and H3K4me3 at the promoter region of SSTR5 | Inhibits laryngeal carcinoma cells proliferation, migration and invasion | [111] |
| ST3Gal6-AS1 | H3K4me3 | Binds MLL1 and recruits it to the ST3Gal6 promoter region | Mediates colorectal cancer progression | [112] |
| TGFB2-AS1 | H3K27me3 | Binds to the EED adaptor of the PRC2 | Regulates TGF-β signaling | [113] |
| UCA1 | H3K27me3 | Interacts with the EZH2 and suppresses p21 expression | Promotes breast cancer progression; expression is higher in tamoxifen-resistant breast cancer cells | [114] |
| XIST | H3K27me3 | Interacts with EZH2 and downregulates DKK1 | Facilitates cell growth, migration and invasion in neuroblastoma | [115] |
| Acetylation/deacetylation | ||||
| ANCR | H3ac, H4ac | Inhibits HNRNPA1 degradation and sponging miR-140-3p | Promotes hepatocellular carcinoma metastasis | [116] |
| ANRIL | H3K9ac | Directly recruits PRC1 and PRC2 | Associated with the development of cardiovascular diseases | [117,118] |
| CircITCH | H3K9ac | Sponge of miR-330-5p that upregulates SIRT6, Survivin and SERCA2a | Regulates DNA damage, mitochondrial oxidative stress, cell death, calcium handling defects and contractile dysfunction in DOX-treated human-induced pluripotent stem-cell-derived cardiomyocytes | [119] |
| circMRPS35 | H4K5ac | Enriches H4K5 acetylation at the regions of FOXO1 and FOXO3a promoters by recruiting the histone acetyltransferase KAT7 | Inhibits invasion and proliferation of gastric cancer cells, related to the clinicopathological features and better prognosis of patients | [120] |
| circMYO10 | H4K16Ac | Regulates miR-370-3p/RUVBL1 axis | Promotes osteosarcoma progression | [121] |
| DLEU1 | H3K27ac | Upregulates interferon-stimulated genes (IFIT1, IFI6 and OAS1ISGs) through activation of JAK-STAT signaling | Promotes oral squamous cell carcinoma progression | [52] |
| DSCAM-AS1 | H3K27ac | Promotes DCTPP1 gene transcription by affecting H3K27 acetylation and enhanced DCTPP1 mRNA stability by binding to the 3′ untranslated region | Suppresses the growth and invasion of ER-positive breast cancer cells | [122] |
| EPB41L4A-AS1 | H3K27ac, H3K14ac | Binds to GCN5 and activates transcription of TXNIP | Regulates glucose uptake | [123] |
| GAS5 | H3K9ac | GAS5 upregulates SIRT1 via inhibition of miR-221 | Inhibits cell proliferation and fibrosis in diabetic nephropathy | [124] |
| H19 | H3ac, H4ac | Sponge of miR-19b, which targets SIRT1 | Involved in diabetic retinopathy; promotes neuroinflammation in ischemic stroke | [59,125] |
| HOTAIR | H3K27ac | Downregulates miR-761 and increases HDAC1 expression; HOTAIR/miR-34A axis mediates SIRT1 expression | Promotes progression and angiogenesis of endometriosis; oncogenic role in tumors | [61,126] |
| HOTAIRM1 | H3K27ac | Positively modulates the activity of c-Jun, which recruits the acetyltransferase p300 to RUNX2 promoter and activates the gene | Promotes osteogenesis | [127] |
| IL6-AS1 | H3K4me, H3K27ac | Promotes IL-6 transcription by recruiting early B-cell factor 1 to the IL-6 promoter | Associated with airway inflammation | [64] |
| KTN1-AS1 | H3K27ac | Recruits EP300 to the KTN1 promoter region | Promotes bladder cancer tumorigenesis | [128] |
| LINC-00162 | H3ac | Promotes HDAC9 via inhibition of miR-383 | Participates in the pathogenesis of diabetic nephropathy | [129] |
| LINC00930 | H3K9ac | Scaffold to recruit the RBBP5 and GCN5 complex to the PFKFB3 promoter | Associated with tumorigenesis, lymphatic invasion, metastasis and poor prognosis in nasopharyngeal carcinoma | [130] |
| lincRNA-p21 | H3ac, H4ac | Inhibits acetylation of H3 and H4 at the Thy-1 promoter and Thy-1 | Could lead to pulmonary fibrosis in acute respiratory distress syndrome | [131] |
| Lnc34a | H3ac, H4ac | Suppresses miR-34a through recruiting HDAC1 to promote histones deacetylation | Significantly overexpressed in hepatocellular cancer and associated with bone metastasis | [132] |
| LncPRESS1 | H3K56ac, H3K9ac | Interacts with SIRT6 | Maintains embryonic stem cells’ pluripotency | [133] |
| MALAT1 | H3ac | Reduces the H3 histone acetylation of the PDX-1 promoter and subsequently inhibits the expression of PDX-1, thus suppressing insulin secretion | Induces the dysfunction of β cells in type 1 diabetes | [134] |
| miR-149-5p | H4K16ac | Regulates KAT8 and H4K16ac expression | Involved in an Alzheimer’s disease pathogenesis | [64] |
| miR-15a-5p | H4ac | Suppresses acetyl-CoA activity and decreases histone H4 acetylation by inhibiting ACSS2 expression | Inhibits metastasis and lipid metabolism | [135] |
| miR-193b-3p | H3ac | Directly targets HDAC3 | Regulates chondrogenesis and chondrocyte metabolism | [136] |
| miR-196-b | H3ac, H4ac | Targets ING5 | Closely linked to the tumor size of neuroblastomas | [137] |
| miR-205 | H3K9ac | Directly targets HDAC2 | Regulates extracellular matrix production in tubular epithelial cells in individuals with diabetic kidney disease | [138] |
| miR-22 | H3K56ac | Directly binds to SIRT1 | Regulator of stress-induced heart damage | [139] |
| miR24-2 | H4K16ac | Targets PRMT7; inhibits HDAC3 through miR-675 | Associated with human tumorigenesis | [94] |
| miR-29a | H3K27ac | Binds to RANKL | Represses osteoclast formation and protects against osteoporosis | [96] |
| miR-29b | H3K14ac | Targets HDAC4 | Mediates podocyte dysfunction and renal fibrosis in diabetic nephropathy | [140] |
| miR-376a | H3K18ac | Targets HDAC9 | Contributes to the development of hepatocellular carcinoma | [141] |
| miR-449 | H3K4ac, H3K9ac | Interacts with HDAC1 | Improves cardiac function | [142] |
| miR-455-3p | H3ac | Directly targets HDAC2/8 | Modulates cartilage development and degeneration | [143] |
| miR-494 | H3K9ac | Inhibits HDAC3 in neurons | Involved in ischemic stroke pathogenesis | [144] |
| miR-520b | H3K9ac | Directly targets HDAC4 | Accelerates lung cancer cell proliferation | [145] |
| miR-543 | H3ac, H4ac | Targets ING5 | Closely linked to the tumor size of neuroblastomas | [137] |
| miR-766-5p | H3K27ac | Directly targets CBP and BRD4, reducing levels of H3K27ac at MYC super-enhancers | Protumorigenic | [146] |
| miR-92a-3p | H3ac | Directly targets HDAC2 | Regulates cartilage development and homeostasis | [147] |
| miR-N5 | H3K56ac | Targets CREBBP, mediating H3K56 acetylation at the promoter of EGFR, β-catenin and CDH1 | Inhibits metastasis of prostate cancer | [148] |
| NEAT1 | H3K27ac | Sponges miR-524-5p, miR-221-3p | Promotes the proliferation and invasion of laryngeal cancer cells; associated with neurodegenerative disorders | [100,149,150,151,152] |
| SATB2-AS1 | H3K27ac, H3K9ac | Acts as a scaffold to recruit p300 | Suppresses colorectal carcinoma progression | [153] |
| SNHG14 | H3K27ac | Upregulates PABPC1 through H3K27 acetylation and modulates PTEN signaling; activates Nrf2 signaling pathway | Promotes hepatocellular carcinoma progression; induces trastuzumab resistance of breast cancer | [154,155] |
| TINCR | H3K27ac | Bind ACLY, starting the TINCR-ACLY-PADI1-MAPK-MMP2/9 axis | Promotes nasopharyngeal carcinoma progression and chemoresistance | [156] |
| TUG1 | H3K9ac | Sponges miR-132-3p, activating HDAC3 | Mediates ischemic myocardial injury | [157] |
| Phosphorylation/dephosphorylation | ||||
| miR-93 | HS10 | Targets a histone kinase Msk2 | Diabetic nephropathy progression | [158] |
| Ubiqiutinylation/deubiquitinylation | ||||
| let-7b | H2BK120ub1 | Binds directly and inhibits USP42, USP44, ATXN7L3 | Tumor-suppressive effects | [159] |
| let-7c | H2BK120ub1 | Binds directly and inhibits USP42, USP44, ATXN7L3 | Tumor-suppressive effects | [159] |
| linc-RA1 | H2BK120ub1 | Binds with H2B and inhibits the interaction between H2Bub1 and USP44 | Associated with advanced clinical stage of glioma, promotes glioma radioresistance | [160] |
| lncFOXO1 | H2AK119ub1 | Regulates level ubH2AK119 at FOXO1 promoter | Suppresses growth of human breast cancer cells | [161] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bure, I.V.; Nemtsova, M.V.; Kuznetsova, E.B. Histone Modifications and Non-Coding RNAs: Mutual Epigenetic Regulation and Role in Pathogenesis. Int. J. Mol. Sci. 2022, 23, 5801. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105801
Bure IV, Nemtsova MV, Kuznetsova EB. Histone Modifications and Non-Coding RNAs: Mutual Epigenetic Regulation and Role in Pathogenesis. International Journal of Molecular Sciences. 2022; 23(10):5801. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105801
Chicago/Turabian StyleBure, Irina V., Marina V. Nemtsova, and Ekaterina B. Kuznetsova. 2022. "Histone Modifications and Non-Coding RNAs: Mutual Epigenetic Regulation and Role in Pathogenesis" International Journal of Molecular Sciences 23, no. 10: 5801. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105801