
Neurotrophin Signaling Impairment by Viral Infections in the Central Nervous System

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
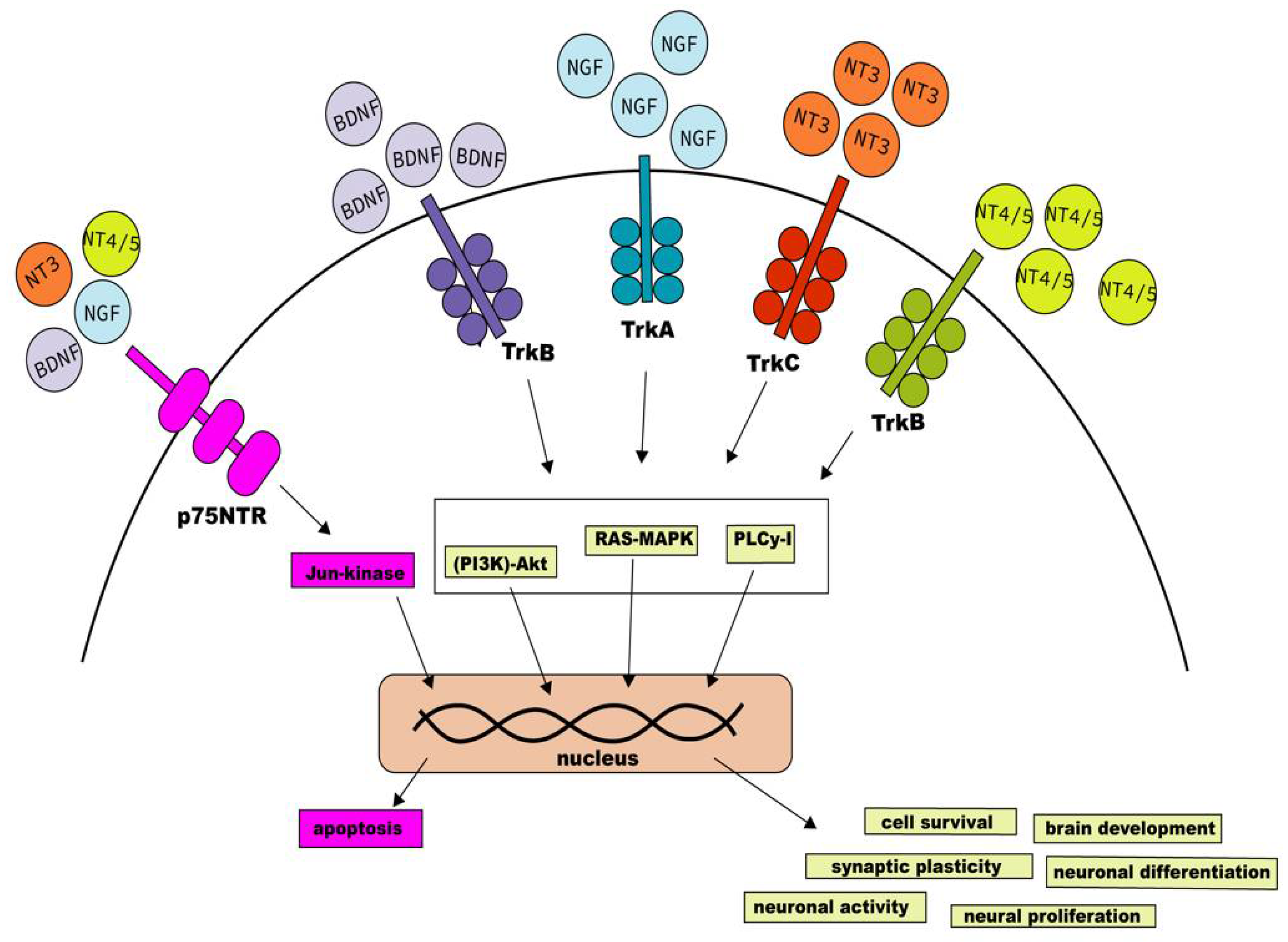
Neurotrophins Receptors Signaling Pathways
2. Role of Neurotrophin Signaling in CNS Homeostasis
2.1. Neurotrophins in the Brain Development
2.2. Synaptic Plasticity
2.3. Role of Neurotrophins in Oligodendrocytes Development and Myelinization
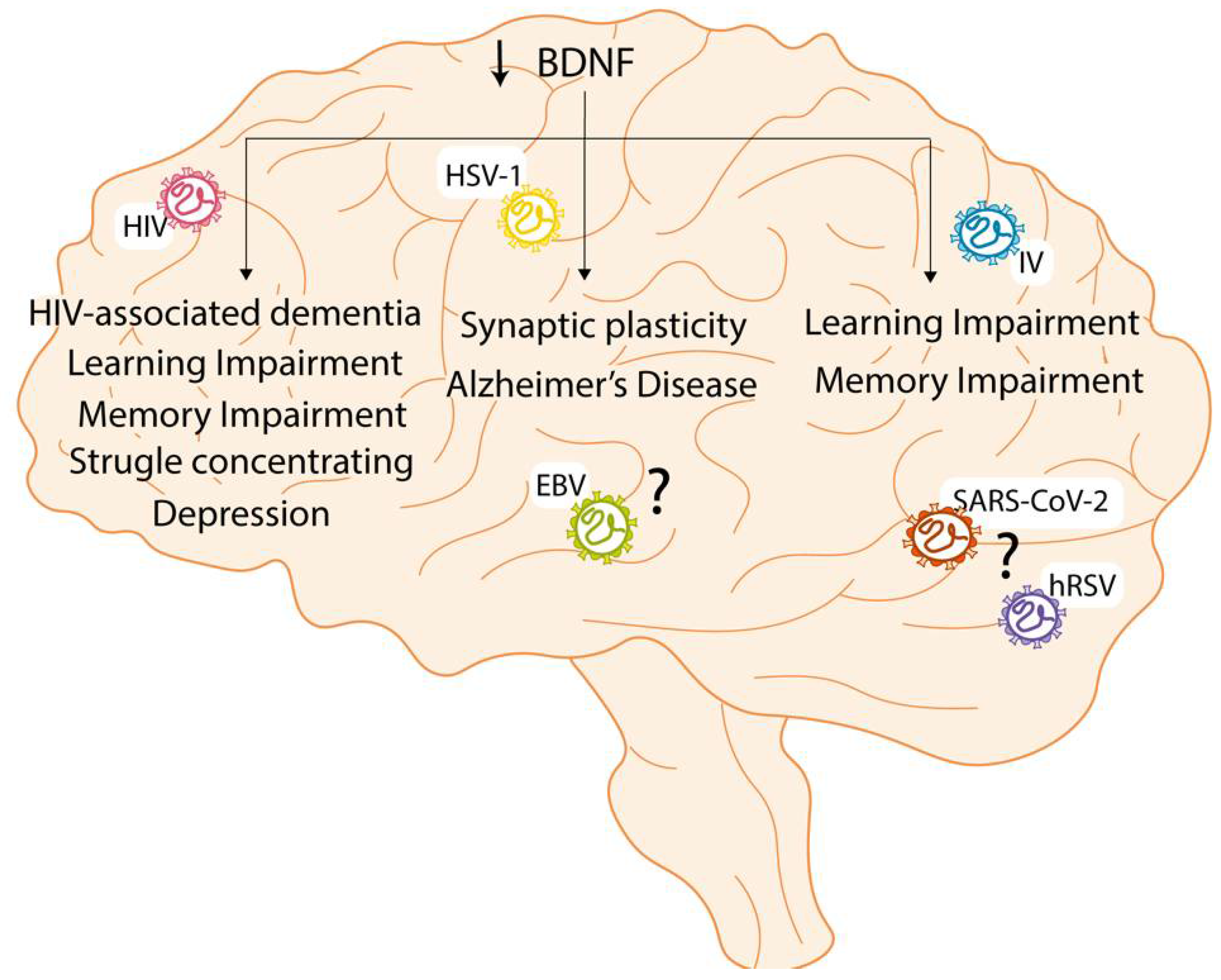
3. Effect of CNS Viral Infections on Neurotrophin Signaling
3.1. Findings and Effects during Viral Infection
3.2. Long-Term Effect
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Arévalo, J.C.; Wu, S.H. Neurotrophin signaling: Many exciting surprises! Cell. Mol. Life Sci. 2006, 63, 1523–1537. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Poo, M.M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2012, 14, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Gibon, J.; Barker, P.A. Neurotrophins and Proneurotrophins: Focus on Synaptic Activity and Plasticity in the Brain. Neuroscientist 2017, 23, 587–604. [Google Scholar] [CrossRef] [PubMed]
- Morel, L.; Domingues, O.; Zimmer, J.; Michel, T. Revisiting the Role of Neurotrophic Factors in Inflammation. Cells 2020, 9, 865. [Google Scholar] [CrossRef] [Green Version]
- Castrén, E.; Antila, H. Neuronal plasticity and neurotrophic factors in drug responses. Mol. Psychiatry 2017, 22, 1085–1095. [Google Scholar] [CrossRef]
- Cappoli, N.; Tabolacci, E.; Aceto, P.; Dello Russo, C. The emerging role of the BDNF-TrkB signaling pathway in the modulation of pain perception. J. Neuroimmunol. 2020, 349, 577406. [Google Scholar] [CrossRef]
- Asaumi, K.; Nakanishi, T.; Asahara, H.; Inoue, H.; Takigawa, M. Expression of neurotrophins and their receptors (TRK) during fracture healing. Bone 2000, 26, 625–633. [Google Scholar] [CrossRef]
- Chavarría, A.; Cárdenas, G. Neuronal influence behind the central nervous system regulation of the immune cells. Front. Integr. Neurosci. 2013, 7, 64. [Google Scholar] [CrossRef] [Green Version]
- Tabakman, R.; Lecht, S.; Sephanova, S.; Arien-Zakay, H.; Lazarovici, P. Interactions between the cells of the immune and nervous system: Neurotrophins as neuroprotection mediators in CNS injury. Prog. Brain Res. 2004, 146, 385–401. [Google Scholar]
- Vega, J.A.; García-Suárez, O.; Hannestad, J.; Pérez-Pérez, M.; Germanà, A. Neurotrophins and the immune system. J. Anat. 2003, 203, 1–19. [Google Scholar] [CrossRef]
- Barouch, R.; Appel, E.; Kazimirsky, G.; Brodie, C. Macrophages express neurotrophins and neurotrophin receptors. J. Neuroimmunol. 2001, 112, 72–77. [Google Scholar] [CrossRef]
- Minnone, G.; De Benedetti, F.; Bracci-Laudiero, L. NGF and its receptors in the regulation of inflammatory response. Int. J. Mol. Sci. 2017, 18, 1028. [Google Scholar] [CrossRef] [PubMed]
- Hoyle, G.W. Neurotrophins and lung disease. Cytokine Growth Factor Rev. 2003, 14, 551–558. [Google Scholar] [CrossRef]
- Al-Qudah, M.A.; Al-Dwairi, A. Mechanisms and regulation of neurotrophin synthesis and secretion. Neurosciences 2016, 21, 306–313. [Google Scholar] [CrossRef] [Green Version]
- Leßmann, V.; Brigadski, T. Mechanisms, locations, and kinetics of synaptic BDNF secretion: An update. Neurosci. Res. 2009, 65, 11–22. [Google Scholar] [CrossRef]
- Kucharczyk, M.; Kurek, A.; Detka, J.; Slusarczyk, J.; Papp, M.; Tota, K.; Basta-Kaim, A.; Kubera, M.; Lason, W.; Budziszewska, B. Chronic mild stress influences nerve growth factor through a matrix metalloproteinase-dependent mechanism. Psychoneuroendocrinology 2016, 66, 11–21. [Google Scholar] [CrossRef]
- Xue, Y.; Liang, H.; Yang, R.; Deng, K.; Tang, M.; Zhang, M. The role of pro- and mature neurotrophins in the depression. Behav. Brain Res. 2021, 404, 113162. [Google Scholar] [CrossRef]
- Harrington, A.W.; Leiner, B.; Blechschmitt, C.; Arevalo, J.C.; Lee, R.; Mörl, K.; Meyer, M.; Hempstead, B.L.; Yoon, S.O.; Giehl, K.M. Secreted proNGF is a pathophysiological death-inducing ligand after adult CNS injury. Proc. Natl. Acad. Sci. USA 2004, 101, 6226–6230. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.M.; Pan, W.; Xu, N.; Zhou, Z.Q.; Zhang, G.F.; Shen, J.C. Environmental enrichment improves long-term memory impairment and aberrant synaptic plasticity by BDNF/TrkB signaling in nerve-injured mice. Neurosci. Lett. 2019, 694, 93–98. [Google Scholar] [CrossRef]
- Kumar, A.; Pareek, V.; Faiq, M.A.; Kumar, P.; Raza, K.; Prasoon, P.; Dantham, S.; Mochan, S. Regulatory role of NGFs in neurocognitive functions. Rev. Neurosci. 2017, 28, 649–673. [Google Scholar] [CrossRef]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Lima Giacobbo, B.; Doorduin, J.; Klein, H.C.; Dierckx, R.A.J.O.; Bromberg, E.; de Vries, E.F.J. Brain-Derived Neurotrophic Factor in Brain Disorders: Focus on Neuroinflammation. Mol. Neurobiol. 2019, 56, 3295–3312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiaretti, A.; Antonelli, A.; Riccardi, R.; Genovese, O.; Pezzotti, P.; Di Rocco, C.; Tortorolo, L.; Piedimonte, G. Nerve growth factor expression correlates with severity and outcome of traumatic brain injury in children. Eur. J. Paediatr. Neurol. 2008, 12, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Failla, M.D.; Juengst, S.B.; Arenth, P.; Wagner, A.K. Preliminary associations between brain derived neurotrophic factor, memory impairment, functional cognition, and depressive symptoms following severe TBI. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Chiaretti, A.; Capozzi, D.; Mariotti, P.; Valentini, P.; Manni, L.; Buonsenso, D.; Fantacci, C.; Ferrara, P. Increased levels of neurotrophins in the cerebrospinal fluid of children with Epstein–Barr virus meningoencephalitis. Int. J. Infect. Dis. 2014, 20, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Meeker, R.; Poulton, W.; Markovis-Plese, S.; Hall, C.; Robertson, K. Protein changes in CSF of HIV-infected patients: Evidence for loss of neuroprotection. Bone 2012, 23, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hirose, M.; Kuroda, Y.; Murata, E. NGF/TrkA Signaling as a Therapeutic Target for Pain. Pain Pract. 2016, 16, 175–182. [Google Scholar] [CrossRef]
- Wysokiński, A. Serum levels of brain-derived neurotrophic factor (BDNF) and neurotrophin-3 (NT-3) in depressed patients with schizophrenia. Nord. J. Psychiatry 2016, 70, 267–271. [Google Scholar] [CrossRef]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef]
- Soppet, D.; Escandon, E.; Maragos, J.; Middlemas, D.S.; Raid, S.W.; Blair, J.; Burton, L.E.; Stanton, B.R.; Kaplan, D.R.; Hunter, T.; et al. The neurotrophic factors brain-derived neurotrophic factor and neurotrophin-3 are ligands for the trkB tyrosine kinase receptor. Cell 1991, 65, 895–903. [Google Scholar] [CrossRef]
- Lewin, G.; Barde, Y.-A. Physiology of the neurotrophins. Annu. Rev. Neurosci. 1979, 2, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.M.; Wu, H.C.; Sun, Z.G.; Lian, F.; Leung, P.C.K. Neurotrophins and glial cell linederived neurotrophic factor in the ovary: Physiological and pathophysiological implications. Hum. Reprod. Update 2019, 25, 224–242. [Google Scholar] [CrossRef] [PubMed]
- Dechant, G.; Neumann, H. Neurotrophins. Mol. Cell. Biol. Neuroprotection CSN 2003, 303–334. [Google Scholar]
- Rodrigues-Amorim, D.; Iglesias-Martínez-almeida, M.; Rivera-Baltanás, T.; Fernández-Palleiro, P.; Freiría-Martínez, L.; Rodríguez-Jamardo, C.; Comís-Tuche, M.; Vallejo-Curto, M.D.C.; Álvarez-Ariza, M.; López-García, M.; et al. The role of the second extracellular loop of norepinephrine transporter, neurotrophin-3 and tropomyosin receptor kinase C in T cells: A peripheral biomarker in the etiology of schizophrenia. Int. J. Mol. Sci. 2021, 22, 8499. [Google Scholar] [CrossRef]
- Tartaglia, L.A.; Ayres, T.M.; Wong, G.H.W.; Goeddel, D.V. A novel domain within the 55 kd TNF receptor signals cell death. Cell 1993, 74, 845–853. [Google Scholar] [CrossRef]
- Zanin, J.P.; Unsain, N.; Anastasia, A. Growth factors and hormones pro-peptides: The unexpected adventures of the BDNF prodomain. J. Neurochem. 2017, 141, 330–340. [Google Scholar] [CrossRef]
- Li, H.; Lin, L.Y.; Zhang, Y.; Lim, Y.; Rahman, M.; Beck, A.; Al-Hawwas, M.; Feng, S.; Bobrovskaya, L.; Zhou, X.F. Pro-BDNF Knockout Causes Abnormal Motor Behaviours and Early Death in Mice. Neuroscience 2020, 438, 145–157. [Google Scholar] [CrossRef]
- Wu, E.H.T.; Wong, Y.H. Pertussis toxin-sensitive Gi/o proteins are involved in nerve growth factor-induced pro-survival Akt signaling cascade in PC12 cells. Cell. Signal. 2005, 17, 881–890. [Google Scholar] [CrossRef]
- Leal, G.; Bramham, C.R.; Duarte, C.B. BDNF and Hippocampal Synaptic Plasticity, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; Volume 104. [Google Scholar]
- Foehr, E.D.; Lin, X.; O’Mahony, A.; Geleziunas, R.; Bradshaw, R.A.; Greene, W.C. NF-κB signaling promotes both cell survival and neurite process formation in nerve growth factor-stimulated PC12 cells. J. Neurosci. 2000, 20, 7556–7563. [Google Scholar] [CrossRef] [Green Version]
- Ascano, M.; Bodmer, D.; Kuruvilla, R. Endocytic trafficking of neurotrophins in neural development. Trends Cell Biol. 2012, 22, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Moya-Alvarado, G.; Gonzalez, A.; Stuardo, N.; Bronfman, F.C. Brain-derived neurotrophic factor (BDNF) regulates Rab5-positive early endosomes in hippocampal neurons to induce dendritic branching. Front. Cell. Neurosci. 2018, 12, 493. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Bucci, C.; Weis, J.; Kruttgen, A. The Small GTPase Rab7 Controls the Endosomal Trafficking and Neuritogenic Signaling of the Nerve Growth Factor Receptor TrkA. J. Neurosci. 2005, 25, 10930. [Google Scholar] [CrossRef]
- Koyuncu, O.O.; Hogue, I.B.; Enquist, L.W. Virus infections in the nervous system. Cell Host Microbe 2013, 13, 379–393. [Google Scholar] [CrossRef] [Green Version]
- Pathak, A.; Carter, B.D. Retrograde apoptotic signaling by the p75 neurotrophin receptor. Neuronal Signal. 2017, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, M.D.; Weil, M.; Raff, M.C. Programmed Cell Death in Animal Development. Cell 1997, 88, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Lachyankar, M.B.; Condon, P.J.; Quesenberry, P.J.; Litofsky, N.S.; Recht, L.D.; Ross, A.H. Embryonic precursor cells that express Trk receptors: Induction of different cell fates by NGF, BDNF, NT-3, and CNTF. Exp. Neurol. 1997, 144, 350–360. [Google Scholar] [CrossRef]
- Ahmed, S.; Reynolds, B.A.; Weiss, S. BDNF enhances the differentiation but not the survival of CNS stem cell- derived neuronal precursors. J. Neurosci. 1995, 15, 5765–5778. [Google Scholar] [CrossRef]
- Sajdel-Sulkowska, E.M.; Xu, M.; McGinnis, W.; Koibuchi, N. Brain region-specific changes in oxidative stress and neurotrophin levels in autism spectrum disorders (ASD). Cerebellum 2011, 10, 43–48. [Google Scholar] [CrossRef]
- Levi-Montalcini, R. Nerve growth factor: 30 years later. Science 1987, 98, 5446–5451. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.; Kermani, P.; Teng, K.K.; Hempstead, B.L. Regulation of cell survival by secreted proneurotrophins. Science 2001, 294, 1945–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.S.; Nishimura, M.C.; Armanini, M.P.; Crowley, C.; Spencer, S.D.; Phillips, H.S. Disruption of a single allele of the nerve growth factor gene results in atrophy of basal forebrain cholinergic neurons and memory deficits. J. Neurosci. 1997, 17, 7288–7296. [Google Scholar] [CrossRef] [PubMed]
- Durany, N.; Michel, T.; Zöchling, R.; Boissl, K.W.; Cruz-Sánchez, F.F.; Riederer, P.; Thome, J. Brain-derived neurotrophic factor and neurotrophin 3 in schizophrenic psychoses. Schizophr. Res. 2001, 52, 79–86. [Google Scholar] [CrossRef]
- Wu, D.; Pardridge, W.M. Neuroprotection with noninvasive neurotrophin delivery to the brain. Proc. Natl. Acad. Sci. USA 1999, 96, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Kohara, K.; Kitamura, A.; Morishima, M.; Tsumoto, T. Activity-dependent transfer of brain-derived neurotrophic factor to postsynaptic neurons. Science 2001, 291, 2419–2423. [Google Scholar] [CrossRef] [Green Version]
- Usui, N.; Watanabe, K.; Ono, K.; Tomita, K.; Tamamaki, N.; Ikenaka, K.; Takebayashi, H. Role of motoneuron-derived neurotrophin 3 in survival and axonal projection of sensory neurons during neural circuit formation. Development 2012, 139, 1125–1132. [Google Scholar] [CrossRef] [Green Version]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Leßmann, V.; Heumann, R. Modulation of unitary glutamatergic synapses by neurotrophin-4/5 or brain-derived neurotrophic factor in hippocampal microcultures: Presynaptic enhancement depends on pre-established paired-pulse facilitation. Neuroscience 1998, 86, 399–413. [Google Scholar] [CrossRef]
- Lewis, G.P.; Linberg, K.A.; Geller, S.F.; Guérin, C.J.; Fisher, S.K. Effects of the neurotrophin brain-derived neurotrophic factor in an experimental model of retinal detachment. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1530–1544. [Google Scholar]
- Rauti, R.; Cellot, G.; D’Andrea, P.; Colliva, A.; Scaini, D.; Tongiorgi, E.; Ballerini, L. BDNF impact on synaptic dynamics: Extra or intracellular long-term release differently regulates cultured hippocampal synapses. Mol. Brain 2020, 13, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilloux, J.-P.; Douillard-Guilloux, G.; Kota, R.; Wang, X.; Gardier, A.M.; Martinowich, K.; Tseng, G.C.; Lewis, D.A.; Sibille, E. Molecular evidence for BDNF- and GABA-related dysfunctions in the amygdala of female subjects with major depression. Mol. Psychiatry 2011, 17, 1130–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginsberg, S.D.; Malek-Ahmadi, M.H.; Alldred, M.J.; Che, S.; Elarova, I.; Chen, Y.; Jeanneteau, F.; Kranz, T.M.; Chao, M.V.; Counts, S.E.; et al. Selective decline of neurotrophin and neurotrophin receptor genes within CA1 pyramidal neurons and hippocampus proper: Correlation with cognitive performance and neuropathology in mild cognitive impairment and Alzheimer’s disease. Hippocampus 2019, 29, 422–439. [Google Scholar] [CrossRef]
- Lee, S.J.; Baek, J.-H.; Kim, Y.-H. Brain-derived Neurotrophic Factor Is Associated with Cognitive Impairment in Elderly Korean Individuals. Clin. Psychopharmacol. Neurosci. 2015, 13, 283. [Google Scholar] [CrossRef] [Green Version]
- Shelley, J.A.; Judy, J.W.; David, D. The Neurotrophins and Their Role in Alzheimer’s Disease. Curr. Neuropharmacol. 2011, 9, 559. [Google Scholar]
- Fields, J.; Dumaop, W.; Langford, T.D.; Rockenstein, E.; Masliah, E. Role of Neurotrophic Factor alterations in the neurodegenerative process in HIV associated neurocognitive disorders. J. Neuroimmune Pharmacol. 2014, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Michael, H.; Mpofana, T.; Ramlall, S.; Oosthuizen, F. The Role of Brain Derived Neurotrophic Factor in HIV-Associated Neurocognitive Disorder: From the Bench-Top to the Bedside. Neuropsychiatr. Dis. Treat. 2020, 16, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Bachis, A.; Avdoshina, V.; Zecca, L.; Parsadanian, M.; Mocchetti, I. Human Immunodeficiency Virus Type 1 Alters Brain-Derived Neurotrophic Factor Processing in Neurons. J. Neurosci. 2012, 32, 9477–9484. [Google Scholar] [CrossRef] [Green Version]
- Sherrard, R.M.; Bower, A.J. BDNF and NT3 extend the critical period for developmental climbing fibre plasticity. Neuroreport 2001, 12, 2871–2874. [Google Scholar] [CrossRef]
- Kobayashi, M.; Matsuoka, I. Enchancement of sympathetic neuron survival by synergistic action of NT3 and GDNF. Neuroreport 2000, 11, 2541–2545. [Google Scholar] [CrossRef]
- Luzuriaga, J.; Pineda, J.R.; Irastorza, I.; Uribe-Etxebarria, V.; García-Gallastegui, P.; Encinas, J.M.; Chamero, P.; Unda, F.; Ibarretxe, G. BDNF and NT3 reprogram human ectomesenchymal dental pulp stem cells to neurogenic and gliogenic neural crest progenitors cultured in serum-free medium. Cell. Physiol. Biochem. 2019, 52, 1361–1380. [Google Scholar] [PubMed]
- Patel, T.D.; Kramer, I.; Kucera, J.; Niederkofler, V.; Jessell, T.M.; Arber, S.; Snider, W.D. Peripheral NT3 signaling is required for ETS protein expression and central patterning of proprioceptive sensory afferents. Neuron 2003, 38, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Ling, Y.L.; Taylor, M.D.; Wright, D.E.; Frank, E. Prenatal exposure to elevated NT3 disrupts synaptic selectivity in the spinal cord. J. Neurosci. 2007, 27, 3686–3694. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.T.; Thompson, B.; Moulton, S.; Newbold, C.; Lum, M.G.; Cameron, A.; Wallace, G.; Kapsa, R.; Clark, G.; O’Leary, S. The effect of polypyrrole with incorporated neurotrophin-3 on the promotion of neurite outgrowth from auditory neurons. Biomaterials 2007, 28, 513–523. [Google Scholar] [CrossRef]
- Jin, L.; Hu, X.; Feng, L. NT3 inhibits FGF2-induced neural progenitor cell proliferation via the PI3K/GSK3 pathway. J. Neurochem. 2005, 93, 1251–1261. [Google Scholar] [CrossRef]
- Lessmann, V.; Gottmann, K.; Heumann, R. BDNF and NT-4/5 enhance glutamatergic synaptic transmission in cultured hippocampal neurones. Neuroreport 1994, 6, 21–25. [Google Scholar] [CrossRef]
- Gao, W.; Zheng, J.L.; Karihaloo, M. Neurotrophin-4/5 (NT-4/5) and Brain-Derived Neutotrophic Factor (BDNF) Act at Later Stages of Cerebellar Granule Cell Differentiation. J. Neurosci. 1995, 15, 2656–2667. [Google Scholar] [CrossRef]
- Widmer, H.R.; Hefti, F. Neurotrophin-4/5 Promotes Survival and Differentiation of Rat Striatal Neurons Developing in Culture. Eur. J. Neurosci. 1994, 6, 1669–1679. [Google Scholar] [CrossRef]
- Cohen, A.; Bray, G.M.; Aguayo, A.J. Neurotrophin-4/5 (NT-4/5) increases adult rat retinal ganglion cell survival and neurite outgrowth in vitro. J. Neurobiol. 1994, 25, 953–959. [Google Scholar] [CrossRef]
- Bosco, A.; Linden, R. BDNF and NT-4 differentially modulate neurite outgrowth in developing retinal ganglion cells. J. Neurosci. Res. 1999, 57, 759–769. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, J.; Li, P.; Ding, Y.; Tang, J.; Chen, G.; Zhang, J.H. NT-4 attenuates neuroinflammation via TrkB/PI3K/FoxO1 pathway after germinal matrix hemorrhage in neonatal rats. J. Neuroinflamm. 2020, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Friedman, B.; Kleinfeld, D.; Ip, N.Y.; Verge, V.M.K.; Moulton, R.; Boland, P.; Zlotchenko, E.; Lindsay, R.M.; Liu, L. BDNF and NT-4/5 exert neurotrophic influences on injured adult spinal motor neurons. J. Neurosci. 1995, 15, 1044–1056. [Google Scholar] [CrossRef] [PubMed]
- Haque, N.S.K.; Hlavin, M.L.; Fawcett, J.W.; Dunnett, S.B. The neurotrophin NT4/5, but not NT3, enhances the efficacy of nigral grafts in a rat model of Parkinson’s disease. Brain Res. 1996, 712, 45–52. [Google Scholar] [CrossRef]
- Altar, C.A.; Boylan, C.B.; Fritsche, M.; Jackson, C.; Hyman, C.; Lindsay, R.M. The neurotrophins NT-4/5 and BDNF augment serotonin, dopamine, and gabaergic systems during behaviorally effective infusions to the substantia nigra. Exp. Neurol. 1994, 130, 31–40. [Google Scholar] [CrossRef]
- Bergami, M.; Rimondini, R.; Santi, S.; Blum, R.; Götz, M.; Canossa, M. Deletion of TrkB in adult progenitors alters newborn neuron integration into hippocampal circuits and increases anxiety-like behavior. Proc. Natl. Acad. Sci. USA 2008, 105, 15570–15575. [Google Scholar] [CrossRef] [Green Version]
- Thoenen, H. Neurotrophins and neuronal plasticity. Science 1995, 270, 593–598. [Google Scholar] [CrossRef]
- Citri, A.; Malenka, R.C. Synaptic plasticity: Multiple forms, functions, and mechanisms. Neuropsychopharmacology 2008, 33, 18–41. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Palacio-Schjetnan, A.; Escobar, M.L. Neurotrophins and synaptic plasticity. Curr. Top. Behav. Neurosci. 2013, 15, 117–136. [Google Scholar]
- Bramham, C.R.; Messaoudi, E. BDNF function in adult synaptic plasticity: The synaptic consolidation hypothesis. Prog. Neurobiol. 2005, 76, 99–125. [Google Scholar] [CrossRef]
- Woo, N.H.; Teng, H.K.; Siao, C.J.; Chiaruttini, C.; Pang, P.T.; Milner, T.A.; Hempstead, B.L.; Lu, B. Activation of p75NTR by proBDNF facilitates hippocampal long-term depression. Nat. Neurosci. 2005, 8, 1069–1077. [Google Scholar] [CrossRef]
- Monteggia, L.M.; Barrot, M.; Powell, C.M.; Berton, O.; Galanis, V.; Gemelli, T.; Meuth, S.; Nagy, A.; Greene, R.W.; Nestler, E.J. Essential role of brain-derived neurotrophic factor in adult hippocampal function. Proc. Natl. Acad. Sci. USA 2004, 101, 10827–10832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Nagappan, G.; Lu, B. Activity-dependent modulation of the BDNF receptor TrkB: Mechanisms and implications. Trends Neurosci. 2005, 28, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Bathina, S.; Das, U.N. Brain-derived neurotrophic factor and its clinical Implications. Arch. Med. Sci. 2015, 11, 1164–1178. [Google Scholar] [CrossRef]
- Mattson, M.P.; Maudsley, S.; Martin, B. BDNF and 5-HT: A dynamic duo in age-related neuronal plasticity and neurodegenerative disorders. Trends Neurosci. 2004, 27, 589–594. [Google Scholar] [CrossRef]
- Tong, L.; Thornton, P.L.; Balazs, R.; Cotman, C.W. β-Amyloid-(1-42) Impairs Activity-dependent cAMP-response Element-binding Protein Signaling in Neurons at Concentrations in Which Cell Survival is Not Compromised. J. Biol. Chem. 2001, 276, 17301–17306. [Google Scholar] [CrossRef] [Green Version]
- Rund, B.R. A review of longitudinal studies of cognitive functions in schizophrenia patients. Schizophr. Bull. 1998, 24, 425–435. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Liang, J.; Chen, D.C.; Xiu, M.H.; De Yang, F.; Kosten, T.A.; Kosten, T.R. Low BDNF is associated with cognitive impairment in chronic patients with schizophrenia. Psychopharmacology 2012, 222, 277–284. [Google Scholar] [CrossRef]
- Mitra, S.; Behbahani, H.; Eriksdotter, M. Innovative therapy for Alzheimer’s disease-with focus on biodelivery of NGF. Front. Neurosci. 2019, 13, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Conner, J.M.; Franks, K.M.; Titterness, A.K.; Russell, K.; Merrill, D.A.; Christie, B.R.; Sejnowski, T.J.; Tuszynski, M.H. NGF is essential for hippocampal plasticity and learning. J. Neurosci. 2009, 29, 10883–10889. [Google Scholar] [CrossRef]
- Shimazu, K.; Zhao, M.; Sakata, K.; Akbarian, S.; Bates, B.; Jaenisch, R.; Lu, B. NT-3 facilitates hippocampal plasticity and learning and memory by regulating neurogenesis. Learn. Mem. 2006, 13, 307–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, A.W.; Xiao, J.; Kemper, D.; Kilpatrick, T.J.; Murray, S.S. Oligodendroglial expression of TrkB independently regulates myelination and progenitor cell proliferation. J. Neurosci. 2013, 33, 4947–4957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Cejudo, J.; Gutiérrez-Fernández, M.; Otero-Ortega, L.; Rodríguez-Frutos, B.; Fuentes, B.; Vallejo-Cremades, M.T.; Hernanz, T.N.; Cerdán, S.; Díez-Tejedor, E. Brain-derived neurotrophic factor administration mediated oligodendrocyte differentiation and myelin formation in subcortical ischemic stroke. Stroke 2015, 46, 221–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McTigue, D.M.; Horner, P.J.; Stokes, B.T.; Gage, F.H. Neurotrophin-3 and brain-derived neurotrophic factor induce oligodendrocyte proliferation and myelination of regenerating axons in the contused adult rat spinal cord. J. Neurosci. 1998, 18, 5354–5365. [Google Scholar] [CrossRef] [PubMed]
- Djalali, S.; Höltje, M.; Große, G.; Rothe, T.; Stroh, T.; Große, J.; Deng, D.R.; Hellweg, R.; Grantyn, R.; Hörtnagl, H.; et al. Effects of brain-derived neurotrophic factor (BDNF) on glial cells and serotonergic neurones during development. J. Neurochem. 2005, 92, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.L.; Murray, S.S.; Xiao, J. Brain-Derived Neurotrophic Factor in Central Nervous System Myelination: A New Mechanism to Promote Myelin Plasticity and Repair. Int. J. Mol. Sci. 2018, 19, 4131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van’t Veer, A.; Du, Y.; Fischer, T.Z.; Boetig, D.R.; Wood, M.R.; Dreyfus, C.F. Brain-derived neurotrophic factor effects on oligodendrocyte progenitors of the basal forebrain are mediated through trkB and the MAP kinase pathway. J. Neurosci. Res. 2009, 87, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Lercher, L.D.; Zhou, R.; Dreyfus, C.F. Mitogen-activated protein kinase pathway mediates effects of brain-derived neurotrophic factor on differentiation of basal forebrain oligodendrocytes. J. Neurosci. Res. 2006, 84, 1692–1702. [Google Scholar] [CrossRef]
- Xiao, J.; Wong, A.W.; Willingham, M.M.; Van Den Buuse, M.; Kilpatrick, T.J.; Murray, S.S. Brain-derived neurotrophic factor promotes central nervous system myelination via a direct effect upon oligodendrocytes. Neurosignals 2010, 18, 186–202. [Google Scholar] [CrossRef]
- Kerschensteiner, M.; Gallmeier, E.; Behrens, L.; Leal, V.V.; Misgeld, T.; Klinkert, W.E.F.; Kolbeck, R.; Hoppe, E.; Oropeza-Wekerle, R.L.; Bartke, I.; et al. Activated human T cells, B cells, and monocytes produce brain-derived neurotrophic factor in vitro and in inflammatory brain lesions: A neuroprotective role of inflammation? J. Exp. Med. 1999, 189, 865–870. [Google Scholar] [CrossRef]
- Xiao, J. Neuroprotection on Multiple Sclerosis: A BDNF Perspective. J. Neurol. Neurophysiol. 2012, 3, 108. [Google Scholar] [CrossRef] [Green Version]
- Frota, E.R.C.; Rodrigues, D.H.; Donadi, E.A.; Brum, D.G.; Maciel, D.R.K.; Teixeira, A.L. Increased plasma levels of brain derived neurotrophic factor (BDNF) after multiple sclerosis relapse. Neurosci. Lett. 2009, 460, 130–132. [Google Scholar] [CrossRef] [PubMed]
- Pöyhönen, S.; Er, S.; Domanskyi, A.; Airavaara, M. Effects of neurotrophic factors in glial cells in the central nervous system: Expression and properties in neurodegeneration and injury. Front. Physiol. 2019, 10, 486. [Google Scholar] [CrossRef] [PubMed]
- Kalinowska-Lyszczarz, A.; Losy, J. The role of neurotrophins in multiple sclerosis-pathological and clinical implications. Int. J. Mol. Sci. 2012, 13, 13713–13725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.R.; Watkins, T.A.; Cosgaya, J.M.; Zhang, C.; Chen, L.; Reichardt, L.F.; Shooter, E.M.; Barres, B.A. NGF Controls Axonal Receptivity to Myelination by Schwann Cells or Oligodendrocytes. Neuron 2004, 43, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capsoni, S.; Covaceuszach, S.; Marinelli, S.; Ceci, M.; Bernardo, A.; Minghetti, L.; Ugolini, G.; Pavone, F.; Cattaneo, A. Taking pain out of NGF: A “painless” NGF mutant, linked to hereditary sensory autonomic neuropathy type V, with full neurotrophic activity. PLoS ONE 2011, 6, e17321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandi, R.; Fabiano, M.; Giorgi, C.; Arisi, I.; La Regina, F.; Malerba, F.; Turturro, S.; Storti, A.E.; Ricevuti, F.; Amadio, S.; et al. Nerve Growth Factor Neutralization Promotes Oligodendrogenesis by Increasing miR-219a-5p Levels. Cells 2021, 10, 405. [Google Scholar] [CrossRef]
- Ladiwala, U.; Lachance, C.; Simoneau, S.J.J.; Bhakar, A.; Barker, P.A.; Antel, J.P. p75 Neurotrophin Receptor Expression on Adult Human Oligodendrocytes: Signaling without Cell Death in Response to NGF. J. Neurosci. 1998, 18, 1297–1304. [Google Scholar] [CrossRef] [Green Version]
- Dowling, P.; Raval, S.; Husar, W.; Casaccia-Bonnefil, P.; Chao, M.; Cook, S.; Blumberg, B. Expression of the p75 neurotrophin receptor in oligodendrocytes in multiple sclerosis brain. Neurology 1997, 48, A25. [Google Scholar]
- Barres, B.A.; Schmid, R.; Sendnter, M.; Raff, M.C. Multiple extracellular signals are required for long-term oligodendrocyte survival. Development 1993, 118, 283–295. [Google Scholar] [CrossRef]
- Barres, B.A.; Raff, M.C.; Gaese, F.; Bartke, I.; Dechant, G.; Barde, Y.A. A crucial role for neurotrophin-3 in oligodendrocyte development. Nature 1994, 367, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Aharoni, R.; Eilam, R.; Domev, H.; Labunskay, G.; Sela, M.; Arnon, R. The immunomodulator glatiramer acetate augments the expression of neurotrophic factors in brains of experimental autoimmune encephalomyelitis mice. Proc. Natl. Acad. Sci. USA 2005, 102, 19045–19050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.M.; Cooksey, E.; Duncan, I.D. Myelin Loss Does Not Lead to Axonal Degeneration in a Long-Lived Model of Chronic Demyelination. J. Neurosci. 2013, 33, 2718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Peskind, E.R.; Millard, S.P.; Chi, P.; Sokal, I.; Yu, C.E.; Bekris, L.M.; Raskind, M.A.; Galasko, D.R.; Montine, T.J. Cerebrospinal fluid concentration of brain-derived neurotrophic factor and cognitive function in non-demented subjects. PLoS ONE 2009, 4, e5424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Failla, M.D.; Conley, Y.P.; Wagner, A.K. Brain-Derived Neurotrophic Factor (BDNF) in Traumatic Brain Injury-Related Mortality: Interrelationships between Genetics and Acute Systemic and Central Nervous System BDNF Profiles. Neurorehabil. Neural Repair 2016, 30, 83–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorley-Lawson, D.A.; Hawkins, J.B.; Tracy, S.I.; Shapiro, M. The Pathogenesis of Epstein-Barr Virus Persistent Infection. Curr. Opin. Virol. 2014, 3, 227–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arduino, P.G.; Porter, S.R. Herpes Simplex Virus Type 1 infection: Overview on relevant clinico-pathological features. J. Oral Pathol. Med. 2008, 37, 107–121. [Google Scholar] [CrossRef]
- Sénécal, V.; Barat, C.; Tremblay, M.J. The delicate balance between neurotoxicity and neuroprotection in the context of HIV-1 infection. Glia 2021, 69, 255–280. [Google Scholar] [CrossRef]
- Wang, Y.; Liao, J.; Tang, S.J.; Shu, J.; Zhang, W. HIV-1 gp120 Upregulates Brain-Derived Neurotrophic Factor (BDNF) Expression in BV2 Cells via the Wnt/β-Catenin Signaling Pathway. J. Mol. Neurosci. 2017, 62, 199–208. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, D.; Feng, J.; Liu, Z.; Hu, Y.; Liu, C.; Kong, X. HIV-1 Protein Tat1–72 Impairs Neuronal Dendrites via Activation of PP1 and Regulation of the CREB/BDNF Pathway. Virol. Sin. 2018, 33, 261–269. [Google Scholar] [CrossRef]
- Darbinian, N.; Darbinyan, A.; Czernik, M.; Peruzzi, F.; Khalili, K.; Reiss, K.; Gordon, J.; Amini, S. HIV-1 tat inhibits NGF-induced Egr-1 transcription activity and consequent p35 expression in neural cells. J. Cell. Physiol. 2008, 216, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, A.; Mehla, R.; Vijayakumar, T.S.; Handy, I. Endocytosis-mediated HIV-1 entry and its significance in the elusive behavior of the virus in astrocytes. Virology 2014, 456–457, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, T.M.L.; Temerozo, J.R.; Giestal-De-Araujo, E.; Bou-Habib, D.C. The effects of neurotrophins and the neuropeptides VIP and PACAP on HIV-1 infection: Histories with opposite ends. Neuroimmunomodulation 2014, 21, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Garaci, E.; Caroleo, M.C.; Aloe, L.; Aquaro, S.; Piacentini, M.; Costa, N.; Amendola, A.; Micera, A.; Caliò, R.; Perno, C.F.; et al. Nerve growth factor is an autocrine factor essential for the survival of macrophages infected with HIV. Proc. Natl. Acad. Sci. USA 1999, 96, 14013–14018. [Google Scholar] [CrossRef] [Green Version]
- Garaci, E.; Aquaro, S.; Lapenta, C.; Amendola, A.; Spada, M.; Covaceuszach, S.; Perno, C.F.; Belardelli, F. Anti-nerve growth factor Ab abrogates macrophage-mediated HIV-1 infection and depletion of CD4+ T lymphocytes in hu-SCID mice. Proc. Natl. Acad. Sci. USA 2003, 100, 8927–8932. [Google Scholar] [CrossRef] [Green Version]
- Camarena, V.; Kobayashi, M.; Kim, J.Y.; Roehm, P.; Perez, R.; Gardner, J.; Wilson, A.C.; Mohr, I.; Chao, M.V. Nature and duration of growth factor signaling through receptor tyrosine kinases regulates HSV-1 latency in neurons. Cell Host Microbe 2010, 8, 320. [Google Scholar] [CrossRef] [Green Version]
- Hollinshead, M.; Johns, H.L.; Sayers, C.L.; Gonzalez-Lopez, C.; Smith, G.L.; Elliott, G. Endocytic tubules regulated by Rab GTPases 5 and 11 are used for envelopment of herpes simplex virus. EMBO J. 2012, 31, 4204. [Google Scholar] [CrossRef] [Green Version]
- Kalil, A.C.; Thomas, P.G. Influenza virus-related critical illness: Pathophysiology and epidemiology. Crit. Care 2019, 23, 258. [Google Scholar] [CrossRef] [Green Version]
- Chiaretti, A.; Pulitanò, S.; Conti, G.; Barone, G.; Buonsenso, D.; Manni, L.; Capozzi, D.; Ria, F.; Riccardi, R. Interleukin and neurotrophin up-regulation correlates with severity of H1N1 infection in children: A case-control study. Int. J. Infect. Dis. 2013, 17, e1186–e1193. [Google Scholar] [CrossRef] [Green Version]
- Giovanni Barone, P.F.; Danilo Buonsenso, L.M. Circulating Plasma Levels of Neurotrophins are Increased in Children with H1N1 Virus Infection. J. Cell Sci. Ther. 2013, 4, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Jurgens, H.A.; Amancherla, K.; Johnson, R.W. Influenza Infection Induces Neuroinflammation, Alters Hippocampal Neuron Morphology, and Impairs Cognition in Adult Mice. J. Neurosci. 2012, 32, 3958–3968. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.; Wilk, E.; Michaelsen-Preusse, K.; Gerhauser, I.; Baumgärtner, W.; Geffers, R.; Schughart, K.; Korte, M. Long-Term Neuroinflammation Induced by Influenza A Virus Infection and the Impact on Hippocampal Neuron Morphology and Function. J. Neurosci. 2018, 38, 3060–3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakadamyali, M.; Rust, M.J.; Zhuang, X. Endocytosis of influenza viruses. Microbes Infect. 2004, 6, 929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohmwald, K.; Espinoza, J.A.; Rey-Jurado, E.; Gómez, R.S.; González, P.A.; Bueno, S.M.; Riedel, C.A.; Kalergis, A.M. Human Respiratory Syncytial Virus: Infection and Pathology. Semin. Respir. Crit. Care Med. 2016, 37, 522–537. [Google Scholar] [CrossRef] [PubMed]
- Tortorolo, L.; Langer, A.; Polidori, G.; Vento, G.; Stampachiacchere, B.; Aloe, L.; Piedimonte, G. Neurotrophin overexpression in lower airways of infants with respiratory syncytial virus infection. Am. J. Respir. Crit. Care Med. 2005, 172, 233–237. [Google Scholar] [CrossRef]
- Nakahara, Y.; Gage, F.H.; Tuszynski, M.H. Grafts of fibroblasts genetically modified to secrete NGF, BDNF, NT-3, or basic FGF elicit differential responses in the adult spinal cord. Cell Transplant. 1996, 5, 191–204. [Google Scholar] [CrossRef]
- Othumpangat, S.; Gibson, L.F.; Samsell, L.; Piedimonte, G. NGF is an essential survival factor for bronchial epithelial cells during respiratory syncytial virus infection. PLoS ONE 2009, 4, e6444. [Google Scholar] [CrossRef]
- Bohmwald, K.; Soto, J.A.; Andrade-Parra, C.; Fernández-Fierro, A.; Espinoza, J.A.; Ríos, M.; Eugenin, E.A.; González, P.A.; Opazo, M.C.; Riedel, C.A.; et al. Lung pathology due to hRSV infection impairs blood–brain barrier permeability enabling astrocyte infection and a long-lasting inflammation in the CNS. Brain. Behav. Immun. 2021, 91, 159–171. [Google Scholar] [CrossRef]
- Lamers, M.M.; Haagmans, B.L. SARS-CoV-2 pathogenesis. Nat. Rev. Microbiol. 2022, 20, 270–284. [Google Scholar] [CrossRef]
- Azoulay, D.; Shehadeh, M.; Chepa, S.; Shaoul, E.; Baroum, M.; Horowitz, N.A.; Kaykov, E. Recovery from SARS-CoV-2 infection is associated with serum BDNF restoration. J. Clean. Prod. 2020, 81, e79. [Google Scholar] [CrossRef]
- Minuzzi, L.G.; Seelaender, M.; Silva, B.S.D.A.; Cunha, E. del B.B.; Deus, M.D.C.; Vasconcellos, F.T.F.; Marqueze, L.F.B.; Gadotti, A.C.; Baena, C.P.; Pereira, T.; et al. COVID-19 Outcome Relates with Circulating BDNF, According to Patient Adiposity and Age. Front. Nutr. 2021, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Usai, C.; Gibbons, J.M.; Pade, C.; Li, W.; Jacobs, S.R.M.; McKnight, A.; Kennedy, P.T.F.; Gill, U.S. The β-NGF/TrkA Signalling Pathway Is Associated with the Production of Anti-Nucleoprotein IgG in Convalescent COVID-19. Front. Immunol. 2022, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bohmwald, K.; Andrade, C.A.; Gálvez, N.M.S.; Mora, V.P.; Muñoz, J.T.; Kalergis, A.M. The Causes and Long-Term Consequences of Viral Encephalitis. Front. Cell. Neurosci. 2021, 15, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Epp, J.R.; Chow, C.; Galea, L.A.M. Hippocampus-dependent learning influences hippocampal neurogenesis. Front. Neurosci. 2013, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Vitaliano, G.D.; Kim, J.K.; Kaufman, M.J.; Adam, C.W.; Zeballos, G.; Shanmugavadivu, A.; Subburaju, S.; McLaughlin, J.P.; Lukas, S.E.; Vitaliano, F. Clathrin-nanoparticles deliver BDNF to hippocampus and enhance neurogenesis, synaptogenesis and cognition in HIV/neuroAIDS mouse model. Commun. Biol. 2022, 5, 1–17. [Google Scholar] [CrossRef]
- Cunha, C.; Brambilla, R.; Thomas, K.L. A simple role for BDNF in learning and memory? Front. Mol. Neurosci. 2010, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Woods, S.P.; Babicz, M.; Shahani, L.; Colpo, G.D.; Morgan, E.E.; Teixeira, A.L. Brain-derived neurotrophic factor (BDNF) is associated with depressive symptoms in older adults with HIV disease. J. Neurovirol. 2021, 27, 70–79. [Google Scholar] [CrossRef]
- Polansky, H.; Goral, B. How an increase in the copy number of HSV-1 during latency can cause Alzheimer’s disease: The viral and cellular dynamics according to the microcompetition model. J. Neurovirol. 2021, 27, 895–916. [Google Scholar] [CrossRef]
- Serrano-Castro, P.J.; Garzón-Maldonado, F.J.; Casado-Naranjo, I.; Ollero-Ortiz, A.; Mínguez-Castellanos, A.; Iglesias-Espinosa, M.; Baena-Palomino, P.; Sánchez-Sanchez, V.; Sánchez-Pérez, R.M.; Rubi-Callejon, J.; et al. The cognitive and psychiatric subacute impairment in severe COVID-19. Sci. Rep. 2022, 12, 3563. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bohmwald, K.; Andrade, C.A.; Mora, V.P.; Muñoz, J.T.; Ramírez, R.; Rojas, M.F.; Kalergis, A.M. Neurotrophin Signaling Impairment by Viral Infections in the Central Nervous System. Int. J. Mol. Sci. 2022, 23, 5817. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105817
Bohmwald K, Andrade CA, Mora VP, Muñoz JT, Ramírez R, Rojas MF, Kalergis AM. Neurotrophin Signaling Impairment by Viral Infections in the Central Nervous System. International Journal of Molecular Sciences. 2022; 23(10):5817. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105817
Chicago/Turabian StyleBohmwald, Karen, Catalina A. Andrade, Valentina P. Mora, José T. Muñoz, Robinson Ramírez, María F. Rojas, and Alexis M. Kalergis. 2022. "Neurotrophin Signaling Impairment by Viral Infections in the Central Nervous System" International Journal of Molecular Sciences 23, no. 10: 5817. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105817