
Quantitative Analysis of the Transcriptome of Two Commonly Used Human Monocytic Cell Lines—THP-1 and Mono Mac 6—Reveals Their Arrest during Early Monocyte/Neutrophil Differentiation


Abstract
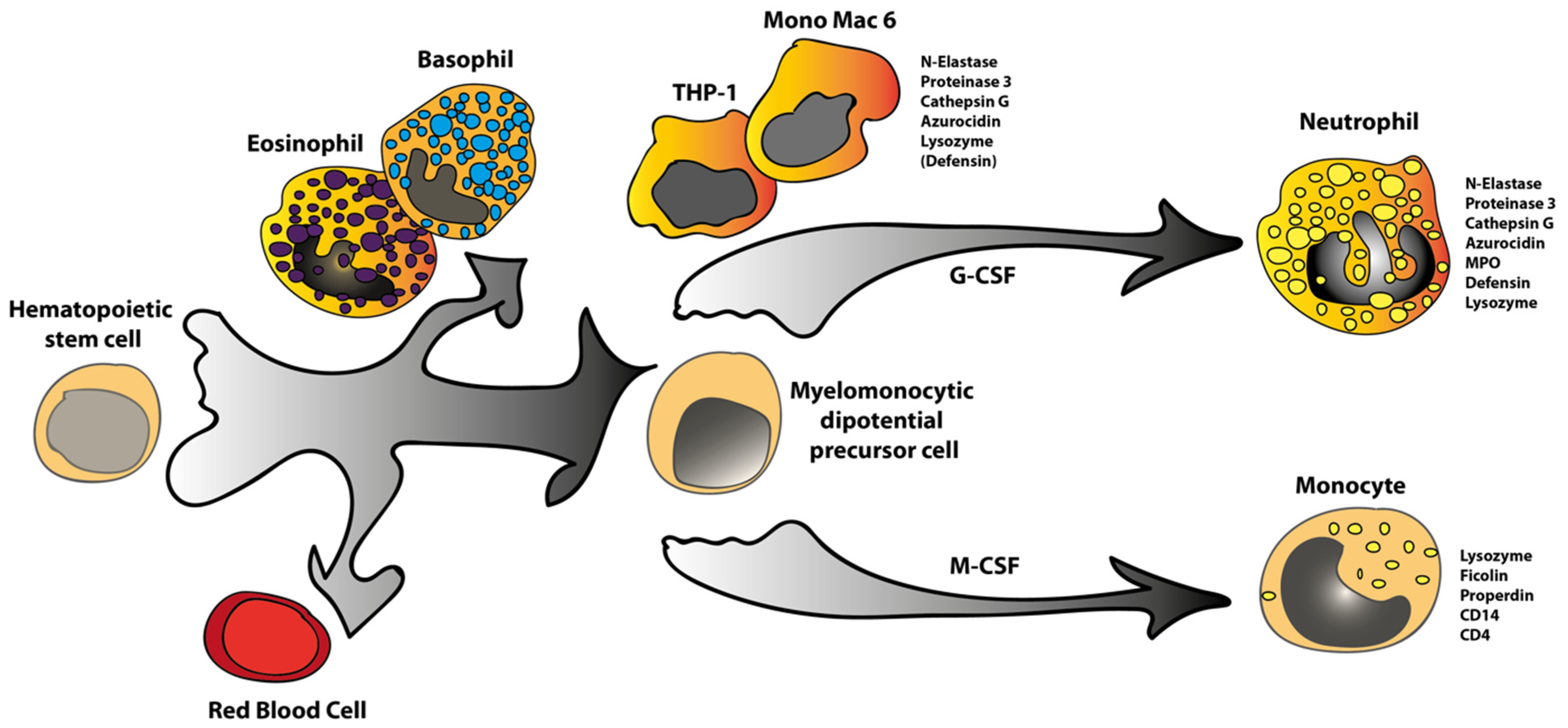
:1. Introduction
2. Results
2.1. Cells, RNA Isolation and Transcriptome Analysis
2.2. Expression Levels of Monocyte/Macrophage Marker Genes
2.3. Expression Levels of Marker Genes Characteristic of Neutrophilic Granulocytes
2.4. Expression of Marker Genes Characteristic of Other Hematopoietic Lineages
3. Discussion
4. Materials and Methods
Cells, RNA Isolation and Transcriptome Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ziegler-Heitbrock, H.W.; Thiel, E.; Fütterer, A.; Herzog, V.; Wirtz, A.; Riethmüller, G. Establishment of a human cell line (Mono Mac 6) with characteristics of mature monocytes. Int. J. Cancer 1988, 41, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef]
- Abrink, M.; E Gobl, A.; Huang, R.; Nilsson, K.; Hellman, L. Human cell lines U-937, THP-1 and Mono Mac 6 represent relatively immature cells of the monocyte-macrophage cell lineage. Leukemia 1994, 8, 1579–1584. [Google Scholar]
- Dahl, R.; Walsh, J.C.; Lancki, D.W.; Laslo, P.; Iyer, S.R.; Singh, H.; Simon, M.C. Regulation of macrophage and neutrophil cell fates by the PU.1:C/EBPα ratio and granulocyte colony-stimulating factor. Nat. Immunol. 2003, 4, 1029–1036. [Google Scholar] [CrossRef]
- Laslo, P.; Spooner, C.J.; Warmflash, A.; Lancki, D.W.; Lee, H.-J.; Sciammas, R.; Gantner, B.N.; Dinner, A.R.; Singh, H. Multilineage Transcriptional Priming and Determination of Alternate Hematopoietic Cell Fates. Cell 2006, 126, 755–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsson, A.; Venkatasubramanian, M.; Chaudhri, V.; Aronow, B.J.; Salomonis, N.; Singh, H.; Grimes, H.L. Single-cell analysis of mixed-lineage states leading to a binary cell fate choice. Nature 2016, 537, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Sieweke, M.H.; Allen, J.E. Beyond Stem Cells: Self-Renewal of Differentiated Macrophages. Science 2013, 342, 1242974. [Google Scholar] [CrossRef] [PubMed]
- Amit, I.; Winter, D.; Jung, S. The role of the local environment and epigenetics in shaping macrophage identity and their effect on tissue homeostasis. Nat. Immunol. 2015, 17, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Perdiguero, E.G.; Klapproth, K.; Schulz, C.; Busch, K.; de Bruijn, M.; Rodewald, H.-R.; Geissmann, F. The Origin of Tissue-Resident Macrophages: When an Erythro-myeloid Progenitor Is an Erythro-myeloid Progenitor. Immunity 2015, 43, 1023–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeffel, G.; Chen, J.; Lavin, Y.; Low, D.; Almeida, F.F.; See, P.; Beaudin, A.E.; Lum, J.; Low, I.; Forsberg, E.C.; et al. C-Myb(+) Erythro-Myeloid Progenitor-Derived Fetal Monocytes Give Rise to Adult Tissue-Resident Macrophages. Immunity 2015, 42, 665–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paivandy, A.; Akula, S.; Lara, S.; Fu, Z.; Olsson, A.-K.; Kleinau, S.; Pejler, G.; Hellman, L. Quantitative In-Depth Transcriptome Analysis Implicates Peritoneal Macrophages as Important Players in the Complement and Coagulation Systems. Int. J. Mol. Sci. 2022, 23, 1185. [Google Scholar] [CrossRef] [PubMed]
- Lara, S.; Akula, S.; Fu, Z.; Olsson, A.-K.; Kleinau, S.; Hellman, L. The Human Monocyte—A Circulating Sensor of Infection and a Potent and Rapid Inducer of Inflammation. Int. J. Mol. Sci. 2022, 23, 3890. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, R.; Collins, S.; Trujillo, J.; McCredie, K.; Ahearn, M.; Tsai, S.; Metzgar, R.; Aulakh, G.; Ting, R.; Ruscetti, F.; et al. Characterization of the continuous, differentiating myeloid cell line (HL-60) from a patient with acute promyelocytic leukemia. Blood 1979, 54, 713–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motakis, E.; Guhl, S.; Ishizu, Y.; Itoh, M.; Kawaji, H.; De Hoon, M.; Lassmann, T.; Carninci, P.; Hayashizaki, Y.; Zuberbier, T.; et al. Redefinition of the human mast cell transcriptome by deep-CAGE sequencing. Blood 2014, 123, e58–e67. [Google Scholar] [CrossRef] [PubMed]
- Wilde, C.G.; Snable, J.L.; E Griffith, J.; Scott, R.W. Characterization of two azurphil granule proteases with active-site homology to neutrophil elastase. J. Biol. Chem. 1990, 265, 2038–2041. [Google Scholar] [CrossRef]
- Sperr, W.R.; Jordan, J.H.; Baghestanian, M.; Kiener, H.P.; Samorapoompichit, P.; Semper, H.; Hauswirth, A.; Schernthaner, G.H.; Chott, A.; Natter, S.; et al. Expression of mast cell tryptase by myeloblasts in a group of patients with acute myeloid leukemia. Blood 2001, 98, 2200–2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Mouse Peritoneal MQ | H-Monoc. Male Age 51 | H-Monoc. Female Age 61 | Mono Mac 6:1 | Mono Mac 6:2 | THP-1 | |
|---|---|---|---|---|---|---|
| A. Monocyte-related transcripts and housekeeping genes | ||||||
| CD14 | 627 | 1697 | 1525 | 43 | 53 | 7 |
| CD4 | 0.6 | 594 | 1422 | 0 | 0 | 182 |
| CD40 | 79 | 6 | 19 | 0.3 | 0.6 | 8 |
| CD86 (B7:2) | 257 | 236 | 108 | 9 | 5 | 0.2 |
| MARCO | 28 | 1 | 4 | 0.2 | 0 | 0 |
| CD68 (Binds oxidized LDL) | 638 | 1273 | 547 | 57 | 55 | 486 |
| LYZ (Lysozyme) | 105,000 | 27,394 | 16,400 | 8795 | 8366 | 1087 |
| FCN1 (Ficolin A) | 1306 | 3198 | 2183 | 0 | 0 | 0 |
| CFP (Compl. factor P, Properdin) | 5225 | 991 | 784 | 5 | 4 | 15 |
| ACTB (beta actin) | 7060 | 19,693 | 23,328 | 13,142 | 12,967 | 17,771 |
| FN1 (Fibronectin) | 25,920 | 0 | 0 | 0 | 0 | 0 |
| PRG4 (Proteoglycan 4, Lubricin) | 3606 | 0 | 0 | 0 | 0 | 0 |
| SRGN (Serglycin) | 2803 | 3855 | 5214 | 19,116 | 19,045 | 4522 |
| SDC3 (Syndecan 3) | 3205 | 5 | 7 | 0 | 0 | 20 |
| ECM1 (Extracellular matrix prot.1) | 3180 | 1 | 0.5 | 18 | 15 | 211 |
| B. Fc receptors | ||||||
| FCGR2A | - | 580 | 1243 | 66 | 70 | 80 |
| FCGR2B | 9 | 59 | 87 | 0.3 | 0 | 1 |
| FCGR3A (FCGR3) | 1968 | 131 | 297 | 0 | 0 | 0.1 |
| FCGR3B | - | 22 | 6 | 0 | 0 | 0 |
| FCGR1A | 35 | 51 | 60 | 24 | 21 | 203 |
| FCER1A | 0 | 7 | 34 | 0 | 0 | 0.2 |
| FCER2 | 0.3 | 2 | 3 | 0.4 | 0.4 | 0.3 |
| FCER1G | 1318 | 1173 | 2125 | 337 | 318 | 704 |
| FCAR (CD89) | - | 92 | 11 | 0 | 0 | 0.2 |
| FCMR (FAIM3) | 1 | 3 | 4 | 0.4 | 0.5 | 0 |
| FCAMR | 0 | 0 | 0 | 0 | 0 | 80 |
| C. MHC class I and II | ||||||
| HLA-A | - | 1548 | 1755 | 634 | 750 | 98 |
| HLA-B | - | 6 | 1329 | 570 | 600 | 2 |
| HLA-C | - | 2979 | 1 | 554 | 706 | 4 |
| B2M | 5521 | 4642 | 1801 | 1536 | 2369 | |
| HLA-DRA | - | 5490 | 3645 | 69 | 61 | 16 |
| HLA-DRB1 | - | 3023 | 2188 | 37 | 35 | 303 |
| HLA-DPA1 | - | 2375 | 4099 | 2 | 2 | 7 |
| HLA-DPB1 | - | 1029 | 1506 | 4 | 5 | 3 |
| D. Lipid mediators and metabolism | ||||||
| ALOX15 (Arachinodate15 lipoxyg.) | 12,680 | 0 | 0 | 0 | 0 | 0.1 |
| PLA2G7 (Phosplipase A2) | 1549 | 15 | 26 | 0.1 | 0.2 | 3 |
| ALOX5AP (Arach.5-lipoxyg.act prot.) | 1296 | 111 | 85 | 5170 | 5429 | 149 |
| PTGIS (Prosaglandin I syntase) | 1283 | 0 | 0 | 90 | 105 | 0 |
| ALOX5 (Arach. 5 -lipoxygenase) | 494 | 190 | 154 | 2 | 0.6 | 14 |
| DPEP2 (Dipeptidase 2 memb. b.) | 1243 | 77 | 85 | 0.5 | 0.5 | 0.4 |
| APOE (Apolipoprotein E) | 2413 | 2 | 4 | 52 | 65 | 51 |
| PLTP (Phospholipidtransfer protein) | 5788 | 1 | 1 | 76 | 101 | 54 |
| E. Anti-microbial and phagocytosis-related proteins | ||||||
| CYBB (Cytoch.b-245 (Nox2) Cytb558) | 2664 | 1081 | 867 | 232 | 159 | 19 |
| PADI4 (Peptidyl arg. deim. type IV) | 1029 | 77 | 31 | 0 | 0 | 0 |
| FLNB (Filamin B phagocytosis) | 2235 | 9 | 9 | 103 | 152 | 53 |
| FLNA (Filamin A) | 1458 | 1175 | 494 | 866 | 1023 | 0 |
| TIMD4 (Binds Phosph. Ser. apoptotic cells) | 1244 | 0 | 0 | 0 | 0 | 0.1 |
| F. Neutrophil, eosinophil, and mast cell-related | ||||||
| MPO (Myeolperoxidase) | 0 | 6 | 18 | 15 | 14 | 5 |
| ELANE (Neutrophil Elastase) | 0 | 0.7 | 1 | 1622 | 1817 | 2542 |
| PRTN3 (Proteinase 3) | 68 | 0 | 0 | 68 | 64 | 2635 |
| CTSG (Cathepsin G) | 0.1 | 0.8 | 7725 | 7712 | 1868 | |
| AZU1 (Azurocidin) | - | 3 | 3 | 941 | 894 | 2410 |
| LTF (Lactoferrin) | 0.3 | 0 | 0.1 | 0 | 0 | 0 |
| BPI (Bacterial permeability incr. pr.) | 0 | 62 | 16 | 0 | 0 | 0 |
| DEFB1 (Defensin beta 1) | 0 | 0 | 0.2 | 5 | 4 | 60 |
| Camp (Cathelicidin) | 0 | 1 | 1 | 0 | 0.1 | 2 |
| TPSB2 (Tryptase B2, MCs) | 0 | 0.3 | 179 | 197 | 0.3 | |
| HDC (Histidine decarboxylase) | 42 | 0 | 0.1 | 0 | 0 | 0 |
| MRGPRX2 (MCs) | - | 0 | 0 | 0 | 0 | 0 |
| EPO (EPX) | 0 | 0 | 0 | 0 | 0 | |
| ECP (RNASE3) | 0 | 0.6 | 128 | 123 | 9 | |
| EDN (RNASE2) | 14 | 40 | 786 | 798 | 1076 | |
| MBP (PRG2) | 0.5 | 2 | 0 | 0.6 | 1 | |
| G. Macrophage and liver-expressed genes | ||||||
| TLN1 (Talin 1 cytosk. memb. con.) | 1971 | 248 | 74 | 172 | 159 | 31 |
| ITSN1 (Intersectin 1 membran traf) | 1359 | 0.1 | 0.6 | 0.3 | 0.2 | 1 |
| GRN (Granulin) | 2744 | 588 | 1041 | 1241 | 1443 | 598 |
| BST1 (ADP-ribosyl cyclase 2) | 1204 | 100 | 19 | 7 | 7 | 31 |
| GDA (Guanine deaminase) | 1232 | 0 | 0 | 0 | 0 | 0.1 |
| HAMP (Hepsidin Iron import) | 876 | 0.7 | 0.1 | 0 | 0 | 0 |
| NINJ1 (Ninjurin 1 apoptosis signal?) | 1122 | 578 | 323 | 28 | 25 | 22 |
| HAL (Histidine amonia lyase) | 1561 | 9 | 33 | 66 | 84 | 29 |
| HRG (Histidine-rich glycoprotein) | 0 | 0 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akula, S.; Lara, S.; Olsson, A.-K.; Hellman, L. Quantitative Analysis of the Transcriptome of Two Commonly Used Human Monocytic Cell Lines—THP-1 and Mono Mac 6—Reveals Their Arrest during Early Monocyte/Neutrophil Differentiation. Int. J. Mol. Sci. 2022, 23, 5818. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105818
Akula S, Lara S, Olsson A-K, Hellman L. Quantitative Analysis of the Transcriptome of Two Commonly Used Human Monocytic Cell Lines—THP-1 and Mono Mac 6—Reveals Their Arrest during Early Monocyte/Neutrophil Differentiation. International Journal of Molecular Sciences. 2022; 23(10):5818. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105818
Chicago/Turabian StyleAkula, Srinivas, Sandra Lara, Anna-Karin Olsson, and Lars Hellman. 2022. "Quantitative Analysis of the Transcriptome of Two Commonly Used Human Monocytic Cell Lines—THP-1 and Mono Mac 6—Reveals Their Arrest during Early Monocyte/Neutrophil Differentiation" International Journal of Molecular Sciences 23, no. 10: 5818. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105818